

Abstract
Angiostatin, a circulating inhibitor of angiogenesis, was identified by its ability to maintain dormancy of established metastases in vivo. In vitro, angiostatin inhibits endothelial cell migration, proliferation, and tube formation, and induces apoptosis in a cell type–specific manner. We have used a construct encoding the kringle domains 1–4 of angiostatin to screen a placenta yeast two-hybrid cDNA library for angiostatin-binding peptides. Here we report the identification of angiomotin, a novel protein that mediates angiostatin inhibition of migration and tube formation of endothelial cells. In vivo, angiomotin is expressed in the endothelial cells of capillaries as well as larger vessels of the human placenta. Upon expression of angiomotin in HeLa cells, angiomotin bound and internalized fluorescein-labeled angiostatin. Transfected angiomotin as well as endogenous angiomotin protein were localized to the leading edge of migrating endothelial cells. Expression of angiomotin in endothelial cells resulted in increased cell migration, suggesting a stimulatory role of angiomotin in cell motility. However, treatment with angiostatin inhibited migration and tube formation in angiomotin-expressing cells but not in control cells. These findings indicate that angiostatin inhibits cell migration by interfering with angiomotin activity in endothelial cells.
Keywords: endothelium, neovascularization, migration, plasminogen, receptor
Introduction
Angiogenesis is the process by which new capillaries are formed by sprouting from preexisting vessels. It is a vital function for the growth of normal tissues during embryogenesis as well as for the pathological growth of tumors (Folkman 1995). Direct evidence has shown that the pathological proliferation of cancer cells will not result in a proportional increase in mass without access to the blood circulation. Tumors form their own circulatory system by upregulating angiogenic stimulators and by downregulating angiogenesis inhibitors (Bouck et al. 1996; Hanahan and Folkman 1996).
The inability of seeded metastases to induce an angiogenic response results in a dormant phenotype. This was shown in a Lewis lung carcinoma model where the primary tumor is angiogenic but also generates a circulating angiogenesis inhibitor, angiostatin, that inhibits vascularization of distant metastases (O'Reilly et al. 1994). Systemic treatment with angiostatin maintains dormancy of these metastases after the removal of the primary tumor. This state of dormancy is characterized by a state of no expansion where cell proliferation is balanced by an equal rate of cell death (Holmgren et al. 1995; Parangi et al. 1996).
Angiostatin is a proteolytically derived fragment of plasminogen spanning the first four kringle domains (O'Reilly et al. 1994). In mice, angiostatin inhibits primary tumor growth as well as angiogenesis-dependent growth of metastases (O'Reilly et al. 1994, O'Reilly et al. 1996; Bergers et al. 1999). In vitro, angiostatin inhibits endothelial cell migration and reduces endothelial cell growth in proliferation assays; this effect is specific for endothelial cells (Ji et al. 1998). Furthermore, it has been shown that angiostatin induces endothelial cell–specific apoptosis in vitro (Claesson-Welsh et al. 1998; Lucas et al. 1998). This is a property shared with other angiogenesis inhibitors such as thrombospondin and endostatin (Dhanabal et al. 1999; Jimenez et al. 2000). In addition, angiostatin has been shown to inhibit in endothelial invasion and tube formation in three-dimensional in vitro angiogenesis assays (Barendsz-Janson et al. 1998; Claesson-Welsh et al. 1998; Griscelli et al. 1998). The in vitro activity of angiostatin in endothelial cell proliferation assays resides in kringles 1–3, with kringle 1 being the most potent inhibitor and kringle 4 being relatively ineffective (Cao et al. 1996). In addition, the kringle 5 domain of plasminogen has also been shown exhibit potent anti-angiogenic activity (Cao et al. 1997).
The mechanisms by which angiostatin inhibits angiogenesis are still unclear. Moser et al. have demonstrated that angiostatin binds to the α/β subunits of ATP synthase. This mitochondrial protein was shown to bind angiostatin on the surface of endothelial cells and potentially inducing H+ influx (Moser et al. 1999). Claesson-Welsh and coworkers demonstrated that angiostatin upregulates focal adhesion kinase (FAK) activity specifically in endothelial cells and proposed that deregulation of FAK may result in inhibition of migration and induction of apoptosis (Claesson-Welsh et al. 1998). In addition, the inhibition of angiogenesis in vivo is dependent on wild-type p53, as angiostatin does not inhibit angiogenesis in p53-null mice (Jimenez et al. 2000).
We have used the yeast two-hybrid system to screen for molecules that interact with angiostatin. Here we report the identification of a novel angiostatin-binding protein we named angiomotin (Latin motus = motility). We show that angiomotin-transfected cells bind and respond to angiostatin by inhibition of cell migration and tube formation. These studies present evidence that angiomotin regulates endothelial cell migration and tube formation.
Materials and Methods
Cells and Reagents
Mouse aortic endothelial cells (Bastaki et al. 1997), EaHy926, Phoenix ecotropic packaging cell line (provided by Dr. G. Nolan, Stanford University, Palo Alto, CA), and NIH 3T3 cells were grown in DME 10% FCS (Hyclone) in 10% CO2. The pBABE vector was provided by Dr. H. Land (ICRF, London, UK), and pBKCMV and pGBT9 were acquired from Stratagene and CLONTECH Laboratories, Inc. Angiostatin was generated from elastase-degraded human plasminogen as described previously by O'Reilly et al. in 1994. Each batch of angiostatin was analyzed for endotoxin using the limulus assay (BioWhittaker).
Yeast Two-hybrid Screening
Manipulation of DNA was performed according to standard molecular biological protocols (Sambrook et al. 1989). The two-hybrid library screen was performed according to the protocol of the manufacturer (CLONTECH Laboratories, Inc.). PGBT9 plasmid containing the Gal4-binding domain together with kringle domains 1–4 bait was transformed with a human term placenta cDNA library cloned into a GAL4 activation domain fusion vector (CLONTECH Laboratories, Inc.). About 2 × 106 yeast transformants were plated on synthetic complete medium but lacking histidine, leucine, and tryptophan. 48–96 h later, 244 yeast colonies were picked and assayed for β-galactosidase (β-gal) activity in o-nitrophenyl β-D-galactopyranoside (ONPG)-containing medium. Plasmid DNA was recovered from 37 double-positive colonies, and candidate clones were retransfected into a new yeast strain (Y190). Three clones retained activity in the new yeast strain. Sequence analysis revealed that these were derived from the same gene.
cDNA Cloning
The whole gene was isolated by screening 3 × 106 clones from a placental λ phage cDNA library (Stratagene) using a Hinf1 fragment of the angiomotin sequence derived from the yeast two-hybrid screen. Five independent clones were isolated that overlapped with the probe. The 5′ sequence was used to design gene-specific primers for 5′RACE PCR (Life Technologies) and used to clone the remaining sequence using mRNA from human umbilical vein endothelial cells (HUVECs).
mRNA Expression Analysis
Commercially obtained multiple human and fetal tissue blots (CLONTECH Laboratories, Inc.) were probed with a 1-kb Pst1 fragment of the 5′ end of angiomotin. The blots were hybridized with ExpressHyb solution according to the protocol of the manufacturer. Real time PCR analysis was performed using TaqMan Reverse Transcription Reagent (PE Biosystems) with random hexamers as reverse transcription (RT) primers. The reaction volume for the RT step was 100 μl and the total RNA amount was 1 μg/sample. The PCR reaction of 10 ng of cDNA was performed using the following oligonucleotides: angiomotin forward primer 5′-GTTTGACCTGCAATCCAGACAA-3′, reverse primer 5′-CCCAGGATCTGAATGGGAGTT-3′, and TaqMan probe 5′-CAGATGGGCCTGTGTTCCACTCCAA-3′. The cycling protocol was as follows: 2 min, 50°C; 10 min, 95°C, 1 cycle, 15 s, 95°C; 1 min, 60°C, 40 cycles. The amount of 18s RNA was quantified as an internal control. The results were related to the EaHy926 sample that expressed the lowest levels of angiomotin.
Generation of Angiomotin-expressing Cell Lines
HeLa cells were transfected with angiomotin open reading frame cloned into the pBKCMV expression vector (Stratagene). Cells were transfected using Lipofectin according to the protocol of the manufacturer (Life Technologies). Stable clones were selected by G418 selection. Phoenix Ecotropic packaging cell line was transfected with the pBABE, pBABE-green fluorescent protein (GFP), pBABE-angiomotin, or pBABE-angiomotin GFP constructs using standard calcium phosphate coprecipitation. The culture supernatant was added to mouse aortic endothelial (MAE) or NIH 3T3 cells and incubated for 24 h. The retrofected cells were then selected in 3 μg/ml puromycin (Sigma-Aldrich) for 2 wk.
Immunofluorescence and Immunohistochemistry
Recombinant glutathione S-transferase (GST)-tagged peptide from the BIG3 clones was used for production of rabbit antisera. Rabbit antisera were affinity purified in three cycles in a thiopropyl-Sepharose 6B (Amersham Pharmacia Biotech) column with covalently coupled immunizing peptide. On the last cycle, the depleted serum was collected and saved for use as a control for the affinity-purified antiserum. The specificity of the purified antibodies was verified by immunoprecipitation and Western blot analysis. For immunofluorescence studies, cells were fixed in 3.7% formaldehyde in PBS blocked in horse serum and incubated for 1 h with primary antibodies. Antibody binding was detected with FITC-labeled anti–rabbit antibodies (Dako). Negative controls included preimmune IgG fraction as well as preincubation with the immunizing antigen. Images were collected with a Hamamatsu CCD camera using the OPEN LAB software. For immunohistochemistry, placental tissue was fixed and sectioned according to standard histological procedures. Sections were permeabilized by microwave treatment in sodium citrate buffer. Sections were incubated overnight with the primary antibody and positive staining was visualized with horseradish peroxidase staining using the Vectastain detection kit according to the protocol of the manufacturer (Vector Laboratories).
In Vitro Kinase Activity
In vitro FAK assays were essentially performed as described by Claesson et al., 1998. Cells were lysed in 50 mM Tris-HCl, pH 7.5/150 mM NaCl/0.5% Triton X-100/1 mM phenyl-methyl-sulfonyl-fluoride/10 mM NaF/1 mM sodium orthovanadate on ice, the nuclei were pelleted, and the supernatants were subjected to immunoprecipitation with FAK monoclonal antibodies (Transduction lab.). The resulting immunoprecipitates were incubated in kinase buffer (10 mM Tris-HCl, pH 7.5/0.01% Triton X-100/10 mM MnCl2) in the presence of γ[32P]ATP (immune complex kinase assay). SDS/PAGE was performed in 7% or 10% linear polyacrylamide gel. The gel was dried and radioactive signals were analyzed using a phosphoimager (Molecular Dynamics).
In Vitro Angiogenesis Assays
Endothelial cell migration was studied in modified Boyden chambers containing chemotaxis membranes with an 8-μm pore size (Nucleopore) which were coated with Collagen 1 (BioWhittaker) as described previously (Kundra et al. 1995). MAE cells were pretreated with angiostatin for 60 min at the concentrations indicated. Cells were detached by trypsinization, washed, and resuspended in serum-free medium containing 0.1% BSA. Approximately 30,000 cells were added to the upper compartment of the Boyden chamber with or without the addition of angiostatin. Medium containing 50 ng/ml basic fibroblast growth factor (bFGF) was used as a chemoattractant and 0.1% mg/ml BSA was placed in the lower chamber. After incubation at 37°C for 4 h, filters were fixed and stained with Mayers Hematoxylin (Sigma-Aldrich) and the cells attached to the bottom side of the membrane were counted visually under the microscope. The data were expressed as the total number of cells counted per high power field. In the in vitro tube formation assays, 150 μl of Matrigel matrix was added to each well of an 8-well chamber slide and incubated at 37°C for 30 min to allow for gel formation. MAE cells were preincubated with 5 μg/ml angiostatin before being plated on Matrigel (Becton Dickinson) at 2 × 105 cells/well, and incubated in 10% CO2 at 37°C for 16 h. Tubes were photographed at 40× magnification and quantified tubes using the caliper available in the OPENLAB program. The average tube length was then multiplied with the total number of tubes.
Results
Identification of a Kringle Domain 1–4 Binding Protein by Yeast Two-hybrid Screening
We generated a Gal4-binding domain fusion protein containing the kringle domains 1–4 of angiostatin. This fusion protein was expressed in yeast and did not activate the His or β-gal reporter genes when expressed alone or in combination with control constructs (Fig. 1A and Fig. B). Approximately 2 × 106 clones from a human term placenta cDNA library were screened with the angiostatin-Gal4 construct. Screening under selective conditions (His−, Leu−, and Trp−) generated 37 positive clones in the CG1945 yeast strain. 7 of the 37 clones displayed high β-gal activity after incubation with ONPG at 30°C for 2 h. The DNA from the colonies that contained high β-gal activity was purified and retransfected into yeast strain Y190. Three of the seven of the colonies retained activity in the new yeast strain. Sequence analysis revealed that these three clones were derived from the same gene, and the yeast-derived sequence was subsequently named BIG3. Binding of the BIG3 peptide to angiostatin was verified by coimmunoprecipitation of angiostatin with recombinant GST-tagged BIG3 domain of angiomotin (Fig. 1 C). Angiostatin binding to BIG3 was visualized with antibodies against kringle domains 1–4 by Western blot.
Figure 1.
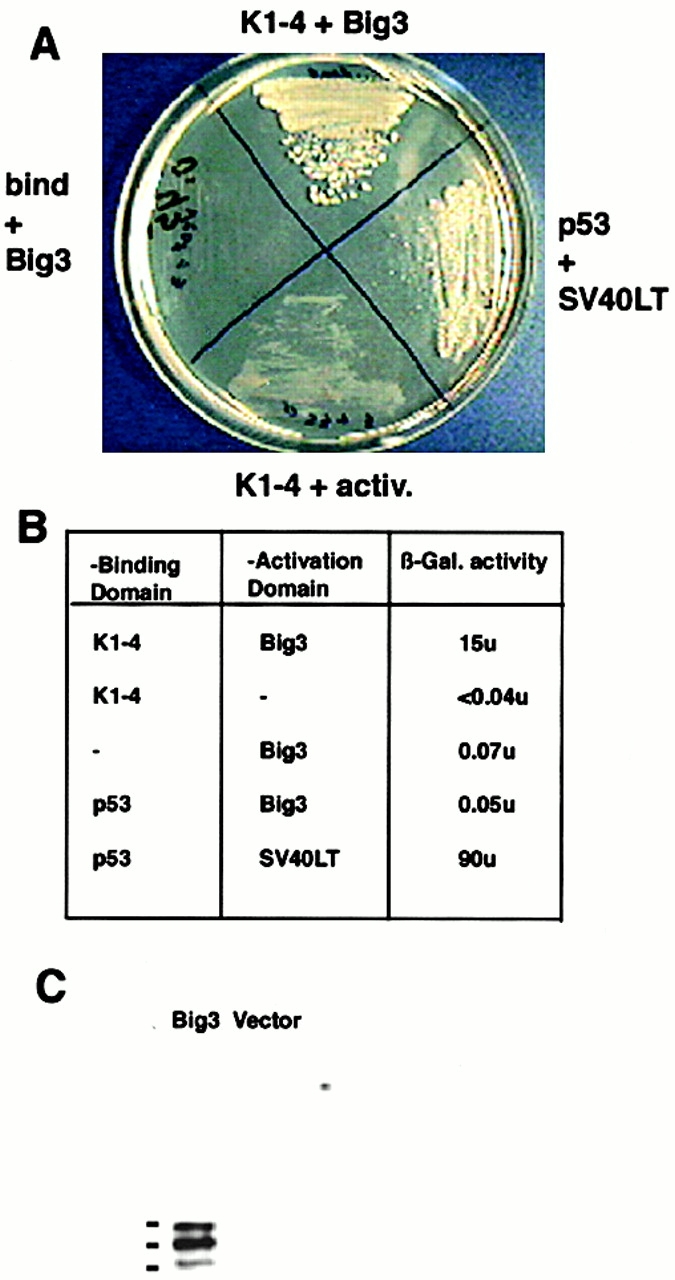
Yeast two-hybrid screening for angiostatin-binding proteins in a human placenta cDNA library. (A) Yeast transfected with the angiostatin-Gal4 binding domain and BIG3-Gal4 activation domain grow under growth-restricted conditions (−Leu, −His, −Trp) and in the presence of 50 mM 3-amino-1, 2,4-triazol. No growth was detected in yeast transfected with both the angiostatin-Gal4 binding domain and the Gal4 activation domain, or the Gal4 binding domain and the BIG3-Gal4 activation domain. The p53-Gal4 binding domain and SV40LT-Gal4 activation domain were used as positive controls. (B) The table shows the activity of the nonselectable β-gal marker in yeast cells transfected with angiostatin-Gal4 binding and the BIG3-Gal4 activation domain and controls. (C) Binding of the 421 amino acid BIG3 sequence derived from yeast two-hybrid screening was verified by coprecipitation of GST-BIG3 with angiostatin.
We cloned and sequenced the whole gene by screening a placenta cDNA phage library with the yeast-derived sequence as a probe in combination with the 5′RACE PCR technique as described in Materials and Methods. The angiomotin cDNA genomic sequence data are available from Genbank/EMBL/DDBJ under accession nos. AF286598 and NT_011819. The cDNA contained a 2,025-bp open reading frame predicted to encode a 675 amino acid protein of an estimated molecular mass of 72 kD (Fig. 2A and Fig. B). Due to its expression in endothelial cells and its effect on cell motility, we named this protein angiomotin. The amino acid sequence did not appear to contain any signal sequence that is usually found in membrane receptors or secreted proteins. This is comparable with other plasminogen- or angiostatin-binding proteins such as alpha enolase, annexin II, or the more recently identified angiostatin-binding protein, ATP synthase (Miles et al. 1991; Das et al. 1994; Hajjar et al. 1996; Moser et al. 1999). All these proteins lack a signal peptide but still bind plasminogen or angiostatin on the endothelial cell surface.
Figure 2.
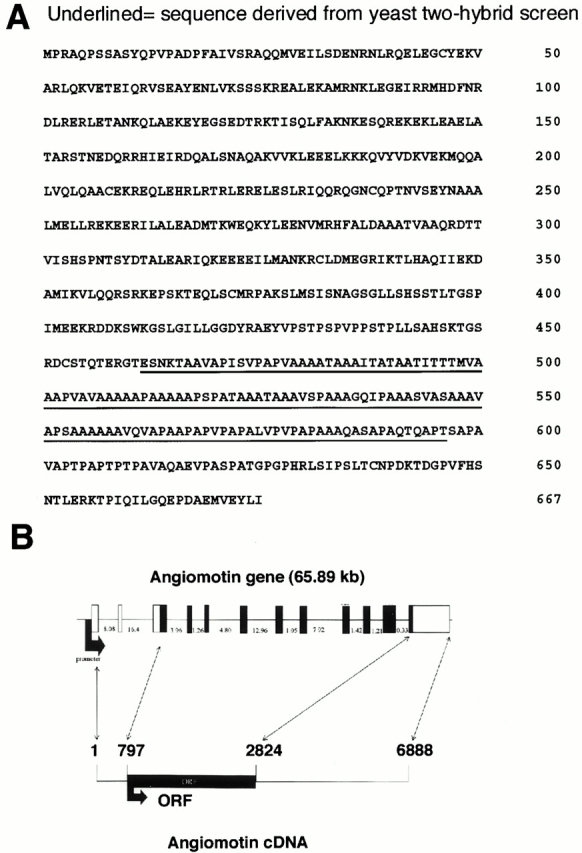
(A) The amino acid sequence of angiomotin. The amino acid sequence of the clone derived from the yeast two-hybrid screening is underlined. (B) Genomic organization of the angiomotin gene, size of cDNA, and the open reading frame (ORF). The Genbank accession number for the angiomotin cDNA genomic sequence data are available from GenBank/EMBL/DDBJ under accession nos. AF286598 and NT_011819.
Binding of Angiostatin to Angiomotin
Next we assessed whether the expression of angiomotin in cells in vitro would confer binding of fluorescein-labeled angiostatin. In our experience, iodination of angiostatin results in loss of activity whereas activity can be retained when fluorescein labeling (FITC) is used (data not shown). Primary bovine endothelial cells were incubated with FITC-angiostatin for 60 min at 4°C before incubation at 37°C for 15 min. Incubation at 37°C resulted in aggregation and internalization of angiostatin (Fig. 3). This pattern of internalization is not detected in fibroblasts where angiostatin only binds to the extracellular matrix. HeLa cells were used for binding experiments, as they do not express angiomotin and exhibit little or no background angiostatin binding (Fig. 3). HeLa cells transfected with angiomotin bound and internalized angiostatin in a similar pattern to that of endothelial cells whereas vector control cells were negative. In addition, internalized FITC-angiostatin colocalized with angiomotin in intracellular vesicles (Fig. 3, bottom).
Figure 3.
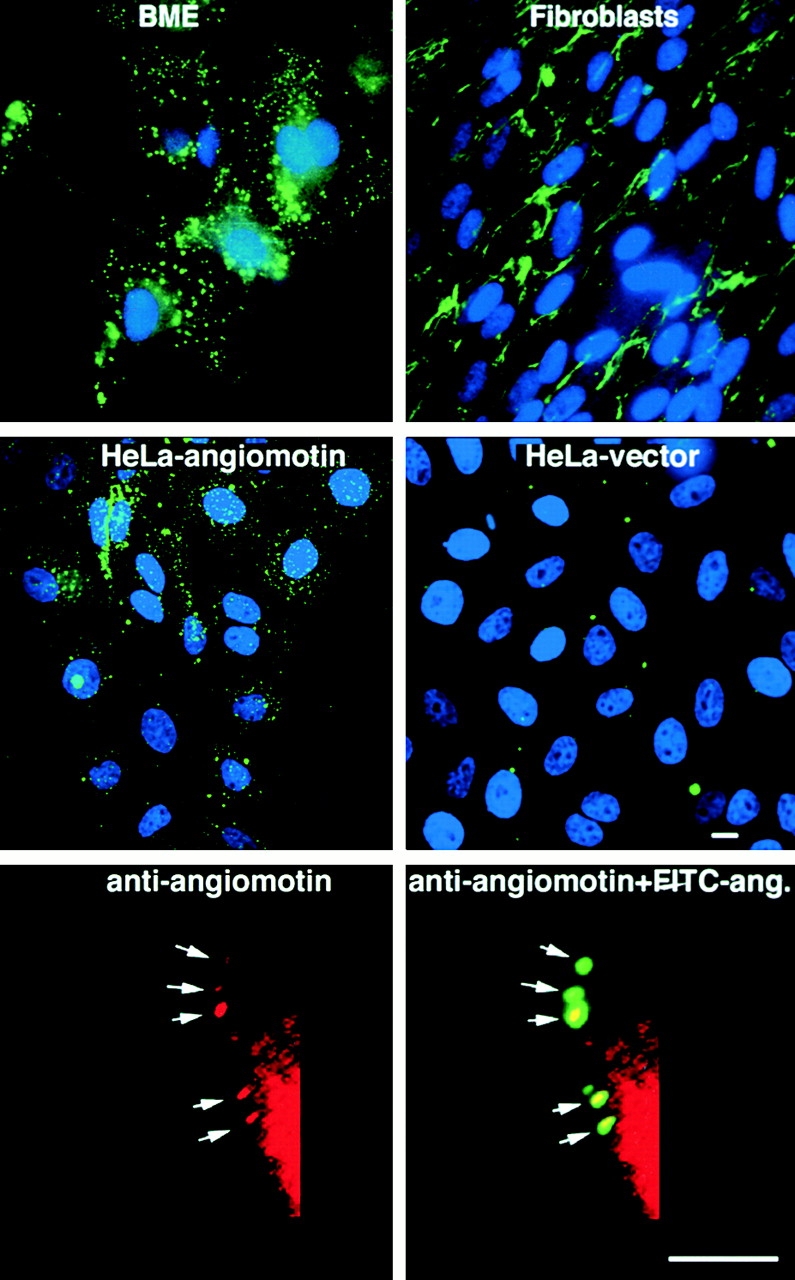
Binding of FITC-labeled angiostatin to living cells in culture. Cells were incubated with 10 μg/ml FITC-labeled angiostatin at 4°C for 60 min. The cells were placed in 37°C incubator 15 min before washing in PBS and fixation in 3.7% formaldehyde. Angiostatin binds to and becomes internalized in bovine microcapillary endothelial (BME) cells resulting in a patchy staining of endosomes (top left). In contrast, angiostatin binds to the matrix of human fetal fibroblasts but is not internalized (top right). HeLa cells transfected with angiomotin bind and internalize angiostatin in a similar pattern to that of endothelial cells (middle left). HeLa-Ctrl cells are transfected with an empty vector and were negative for angiostatin binding (middle right). The bottom panel shows colocalization of FITC-angiostatin (ang.) with angiomotin during internalization after surface binding to HeLa cells transfected with angiomotin. Angiomotin was visualized by immunofluorescent staining using immunoaffinity-purified rabbit polyclonal antibodies against angiomotin as described in Materials and Methods. Bars, 10 μm.
Angiomotin Expression In Vivo and In Vitro
We investigated the expression of angiomotin mRNA by Northern blot analysis of mRNA derived from fetal and adult human tissues. Hybridization with a 1-kb probe from nucleotides 953–2,152 showed positive hybridization in both fetal and adult tissues (Fig. 4 A). Two mRNA bands with approximate sizes of 6.5 and 7.5 kb showed the most abundant expression in placenta and skeletal muscle but were detectable in all analyzed tissues. We also analyzed the expression of Angiomotin mRNA in 15 different cell lines by real time PCR. Highest expression could be detected in human dermal microvascular and HUVECs (Table ). To study angiomotin protein expression in vivo, we raised a polyclonal antiserum directed against the recombinant protein that was derived from the sequence identified from the initial yeast two-hybrid clone. The resulting antiangiomotin serum was subsequently affinity purified against the immunizing peptide over an affinity column. The serum and affinity-purified antibodies detected a protein with a molecular mass of 75 kD in angiomotin-transfected NIH 3T3 cells but not in cells transfected with vector alone (Fig. 4 B).
Figure 4.

Angiomotin is expressed in endothelial cells in vivo. (A) Northern blot analysis of angiomotin expression in fetal and adult human tissues. 2 μg of poly A+ RNA was analyzed for expression of angiomotin using a 1-kb probe from the 5′ region. Two detectable transcripts were detected in both fetal and adult tissues (multitissue blot from CLONTECH Laboratories, Inc.; H, heart; Br, brain; Pl placenta; Lu, lung; Li, liver; Ki, kidney; Pa, pancreas; M, marker). (B) Western blot analysis using immunoaffinity-purified rabbit polyclonal antibodies against angiomotin. The antibodies detected a band with an approximate molecular mass of 75 kD in angiomotin-transfected NIH 3T3 (Am.) cells but not in the vector (vect.) control. (C) Immunohistochemical localization of ABP-1 protein in human placenta. Paraffin-embedded placental tissue from term placenta was stained with immunoaffinity-purified angiomotin polyclonal antibodies. (C, a) Shows angiomotin protein in endothelium of larger vessels in a placental villus. (C, b) Immunostaining using a monoclonal antibody against CD34, an endothelial marker. (C, c) Detection of angiomotin protein in capillaries of placental microvilli. (C, d) IgG negative control. (D) Expression of angiomotin in a dermal Kaposis sarcoma. (D, a) Shows angiomotin staining of a blood vessel within the Kaposis lesion. (D, b) CD34 staining of a consecutive section of D, panel a. (D, c) Angiomotin-negative dermal blood vessel (arrow) in the normal tissue adjacent to the Kaposis lesion. (D, d) Same as D, panel c, but stained with CD34. D, panels e and f, are negative controls stained with nonimmune rabbit IgG as a negative control. Bars, 40 μm.
Table 1.
Real Time RT-PCR Analysis of Angiomotin Expression in Various Cell Lines
| Tissue | Angiomotin/18s mRNA ratio | |
|---|---|---|
| Primary cells | ||
| Dermis | H. dermal MVEC | 12561 |
| Placenta | HUVEC | 320 |
| Dermis | H. dermal fibroblasts | 6 |
| Cell lines | ||
| Lung | H460 | 18 |
| Ovarian | A2780 | 67 |
| Melanoma | A375 | 87 |
| Cervical | HELA | 4 |
| Prostate | DU145 | 30 |
| Bladder | ECV304 | 166 |
| Colon | HT29 | 28 |
| Lymphoid | CEM | 6 |
| Erythroid/Myeloid | K562 | 3 |
| Lymphoid | Jurkat | 2 |
| myeloid | THP-1 | 9 |
| EC/epithelial hybrid | EaHy926 | 1 |
Angiomotin mRNA levels were quantified in 10 ng cDNA generated by RT using random hexamers. The result shows the expression of angiomotin relative to EaHy926 cells that showed the lowest level of expression of the target. The values are represented as the ratio of angiomotin/18s mRNA. EC, endothelial cell; H., human; MVEC, microvascular endothelial cell.
Immunohistochemical analysis of human term placental sections revealed positive staining in the endothelium of larger vessels as well as in capillaries of microvilli (Fig. 4 C). In addition, positive staining was also detected in extravillous cytotrophoblasts (data not shown). We also analyzed the angiomotin expression in tumor vessels of Kaposis sarcoma and vessels in adjacent normal tissue (Fig. 4 D). Angiomotin protein was detected in the endothelial cells of vessels within the tumor but not in the blood vessels of the surrounding normal dermal tissue.
Angiomotin Localizes to Lamellipodia of Migrating Endothelial Cells
Next we assessed the subcellular localization of Angiomotin in transfected cells. The angiomotin open reading frame was fused to GFP in the COOH terminus. Angiomotin was expressed in NIH 3T3 cells using a retroviral expression vector. The cellular localization differed depending on whether cells were migrating or not. In migrating cells, angiomotin localized to the leading edge of the lamellipodia of the migrating cell. Double staining with antibodies against FAK showed overlapping localization of these proteins in migratory cells (Fig. 5, a and b). We also examined the distribution of endogenous angiomotin in human umbilical cord endothelial cells. Cells were incubated with angiomotin serum and rhodamine-labeled phalloidin in order to visualize F-actin. The angiomotin antibodies stained the leading edge of migrating cells (Fig. 5 c) and peripheral as well as circular ruffles in spreading cells. Angiomotin overlaps with F-actin staining in these structures, whereas no positive angiomotin staining was detected in stress fibers (Fig. 5e and Fig. f).
Figure 5.
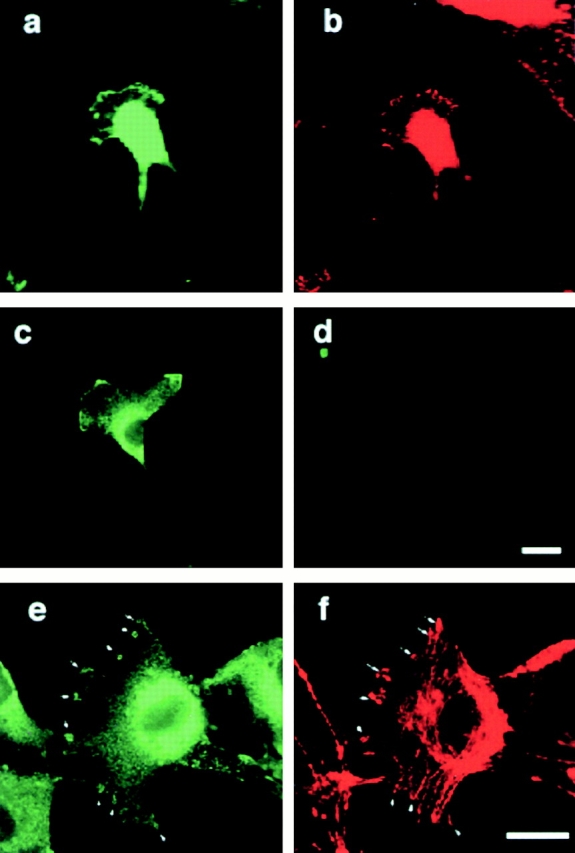
Subcellular localization of transfected and endogenous angiomotin. GFP-tagged angiomotin was expressed in NIH 3T3 cells using a retroviral expression system. Colocalization of GFP-angiomotin (a) with FAK (b) in the extending lamellipodium. (c) Immunofluorescence staining with a polyclonal antibody against angiomotin shows similar localization of endogenous angiomotin in a migrating HUVEC. (d) Control IgG from the same immunized animal. e and f shows the localization of angiomotin (e) to actin ruffles (f) and focal complexes in spreading HUVECs as visualized by phalloidin staining (arrows). Bars, 10 μm.
Angiostatin Induces FAK Activity in Angiomotin-transfected Cells In Vitro
Previous studies have demonstrated that angiostatin induces FAK activity in in vitro assays (Claesson-Welsh et al. 1998). This is, to our knowledge, the only reported signaling pathway that is affected by angiostatin. To assess the effect of angiomotin on FAK activity, we transfected angiomotin into the human cell line EaHy926 (derived from a fusion hybrid between HUVECs and a carcinoma cell line) or into the immortalized MAE cell line. Expression of endogenous angiomotin was not detectable by RT-PCR analysis in neither of these two cell lines (data not shown). Addition of angiostatin to angiomotin-transfected cells increased the in vitro kinase activity of FAK within 1 h (Fig. 6A and Fig. B). By contrast, no effect of angiostatin was detected in cells transfected with the empty vector.
Figure 6.

Angiostatin induces FAK activity in vitro in cells transfected with angiomotin. (A) Angiomotin-negative EaHy926 or (B) MAE cells were transfected with angiomotin or empty vector. Cells were stimulated with angiostatin at the indicated time points. FAK was immunoprecipitated and subjected to in vitro kinase analysis as described in Materials and Methods. Arrows indicate the localization of p125 FAK.
Angiostatin Inhibits Migration and Tube Formation of ABP-1 Transfected Cells
The localization of angiomotin in the leading edge of migrating cells and the effect of angiostatin on FAK kinase activity in angiomotin-transfected cells, suggested that angiomotin might play a role in endothelial cell migration. To test this hypothesis, we studied the effect of angiomotin expression in an angiomotin-negative immortalized MAE cell line. MAE cells were infected with ecotropic retrovirus expressing full-length angiomotin or vector alone. Polyclonal cells were selected by puromycin treatment in order to remove noninfected cells. The effect of angiomotin on cell migration was tested in a modified Boyden chamber. Analysis of spontaneous migration in the absence of chemoattractant showed that angiomotin cells consistently exhibited a higher random migration than the vector control cells (Fig. 7 A). We then tested whether angiostatin had any effect on the increased migratory rate of the MAE-angiomotin cells. Cells were preincubated with 5 μg/ml of angiostatin for 1 h before being added to the migration chamber. Angiostatin did not affect the random or bFGF-induced migration of the control cells. In contrast, angiostatin reduced the motility of MAE-angiomotin cells to background levels (Fig. 7 B) The inhibition of cell migration was dose dependent with maximal inhibition estimated at 500 ng/ml angiostatin (Fig. 7 C).
Figure 7.
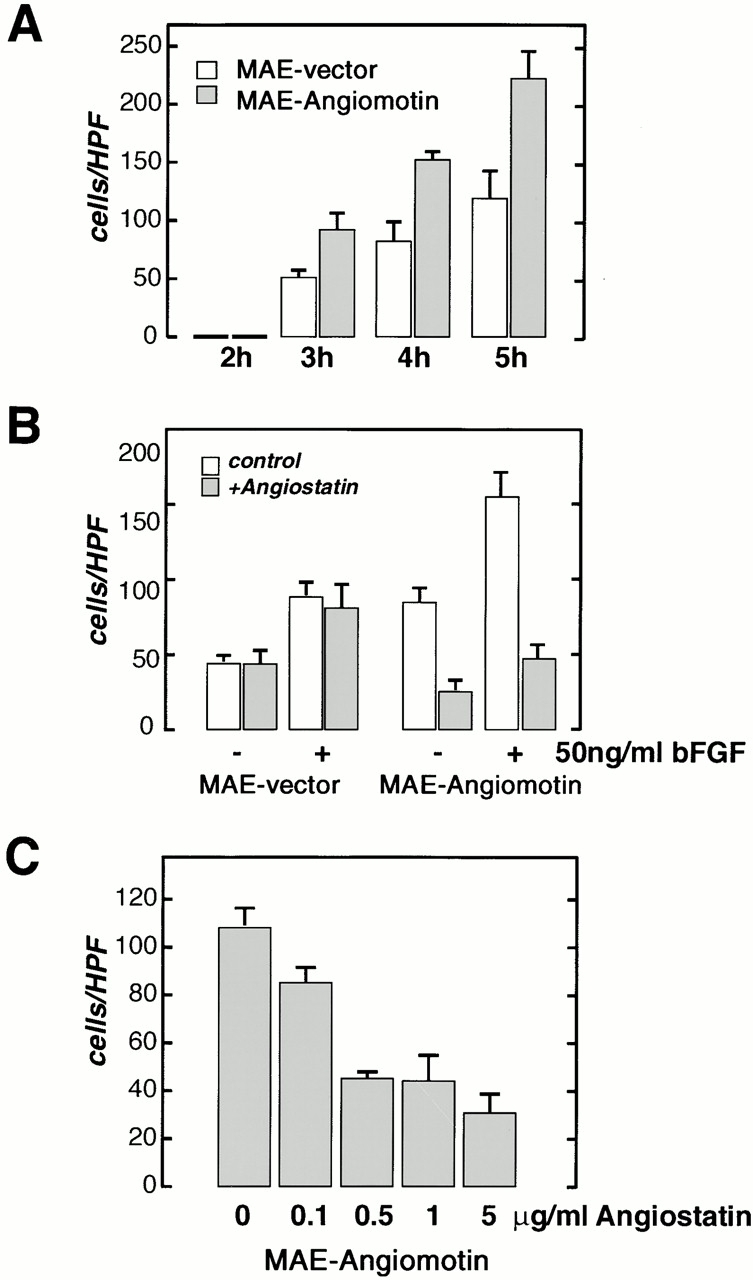
Migration of MAE cells transfected with angiomotin or with the vector alone was studied in the modified Boyden chamber assay. (A) Analysis of spontaneous migration in the absence of chemotactic factor at different time points after start of the experiment. A higher number of angiomotin-expressing cells are migrating through the filter at all time points. Vertical axis, cells migrated per high power field (HPF). (B) Effect of angiostatin on migration of MAE-angiomotin and MAE vector cells with or without stimulation with bFGF. Cells were pretreated for 1 h with angiostatin as indicated. (C) Dose response of angiostatin-mediated inhibition of migration of angiomotin-transfected MAE cells. All samples were performed in quadruplicates (error bars = SD).
To further explore the role of angiomotin in mediating angiostatin inhibition of angiogenesis, we studied the effect of angiostatin on the formation of vascular structures in the matrigel tube formation assay. No detectable difference between angiomotin- and vector-transfected MAE cells in the capacity to form tubes in vitro could be detected (Fig. 8 A). However, pretreating the cells for 16 h with angiostatin inhibited tube formation and caused a 90% inhibition of total tube length (Fig. 8A and Fig. B). This effect was specific for cells transfected with angiomotin, whereas angiostatin did not affect tubulogenesis in control cells.
Figure 8.
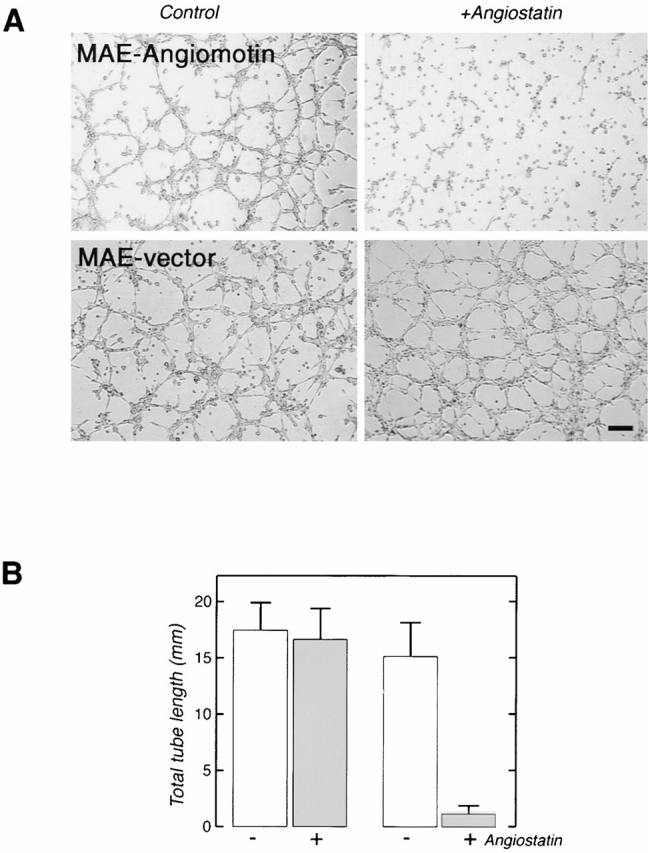
(A) Angiostatin inhibits tube formation of angiomotin-transfected cells plated on matrigel. MAE-vector and MAE-angiomotin transfected cells were pretreated with 5 μg/ml angiostatin for 16 h before trypsinization and plating on matrigel. Images show tube formation 16 h after seeding on matrigel (Bar, 130 μm). (B) Total tube length formed in the presence or absence of angiostatin. The data represent the average from three independent experiments (error bars = SD).
Discussion
Angiostatin was one of the first tumor-generated angiogenesis inhibitors to be identified. The mechanisms of action appears to be highly specific, as angiostatin inhibits migration and induces apoptosis specifically in endothelial cells in vitro and interferes with blood vessel formation in vivo. Here we report the identification of angiomotin, a novel protein mediating angiostatin inhibition of migration and tube formation of endothelial cells. We used the yeast two-hybrid system to identify angiostatin-binding peptides in a human placental cDNA library. This screening procedure has been widely used to identify protein–protein interactions such as proteins that interact with the cytoplasmic parts of receptors. However, it has also been shown to be a useful tool to analyze the interactions of extracellular ligands and their receptors (Zhu and Kahn 1997). The sequence encoding the angiostatin-binding domain of angiomotin was identified in three independent clones after screening a placental cDNA library with the K1–4 domains of angiostatin. The amino acid sequence was rich in alanine (40%) and proline (20%). The proline/alanine rich sequence included 4 (PXXP) motifs which may be potential binding sites for Src homology 3 domains which may be found in a diverse group of signal-transducing molecules (Sudol 1998). The angiomotin amino acid sequence did not appear to contain any of the signal sequences that are normally found in membrane receptors or secreted proteins. This is in analogy with other plasminogen- or angiostatin-binding proteins such as alpha enolase, annexin II, or the more recently identified angiostatin-binding protein, ATP synthase (Miles et al. 1991; Das et al. 1994; Hajjar et al. 1996). These proteins lack the endoplasmic reticulum targeting peptide but are still localized to and are able to bind plasminogen or angiostatin on the endothelial cell surface.
The expression of angiomotin in the endothelium was verified by immunohistochemical staining of placental vessels. In the human term placenta, positive staining was detected in larger vessels as well as in capillaries. In addition, positive staining could also be detected in cytotrophoblasts, a cell type characterized by its ability to invade the endometrium and form a transitional circulatory system during early embryogenesis. Interestingly, these cells have been shown to express other “endothelial cell–specific” proteins such as the vascular endothelial growth factor (VEGF)-receptor 1 and tyrosine kinase with immunoglobin and EGF homology domains (TIE)-2 (Ahmed et al. 1995; Dunk et al. 2000). We also analyzed the expression pattern of angiomotin in Kaposis sarcoma lesion, a tumor with endothelial origin. Positive staining could be detected in vessels infiltrating the tumor. Interestingly, dermal capillaries in adjacent normal tissue in the same section were negative, thus indicating that there is a differential expression of angiomotin in vessels of normal and pathological tissues.
The endothelial cell migration assay has proven to be a consistent predictor of inhibition of neovascularization in vivo. Indeed, angiostatin, endostatin, and thrombospondin all inhibit endothelial cell migration in vitro (Good et al. 1990; Ji et al. 1998; Yamaguchi et al. 1999). Thrombospondin is probably the best characterized angiogenesis inhibitor with regard to signaling pathways. It binds the CD36 receptor and activates p38 mitogen-activated protein kinase (MAPK) via the src-related kinase, c-fyn (Jimenez et al. 2000). In contrast, angiostatin-induced inhibition of angiogenesis is not dependent on either CD36 or c-fyn, as was recently shown in the mouse cornea angiogenesis assay (Jimenez et al. 2000).
Our findings indicate a direct functional role for angiomotin in endothelial cell migration, as angiomotin is located in areas of actin reorganization and cells expressing angiomotin migrate significantly faster than control cells. Treatment of angiomotin-transfected cells with angiostatin efficiently inhibited basal as well as bFGF-stimulated migration of these cells, indicating that there is a direct functional link between angiomotin and angiostatin. Furthermore, we also showed that angiostatin reduced the total tube length generated by angiomotin-transfected cells in the matrigel assay by 90%. However, endothelial cells that are plated on Matrigel rapidly migrate to form cell aggregates. Endothelial tubes are then extended between these aggregates of cells. Angiostatin-treated MAE-angiomotin cells remained primarily as single cells on the matrigel and did not form aggregates. This observation suggests that angiostatin interferes with the early processes of tube formation involving cell motility.
The angiomotin sequence provides little information into exactly how it may be involved in mediating angiostatin inhibition of in vitro angiogenesis. The lack of a signal peptide and transmembrane domain argues that angiomotin does not act as a typical membrane receptor. The NH2-terminal coil–coil domain as well as the proline rich sequences in the angiostatin-binding domain suggest that angiomotin forms protein complexes. Considering the stimulatory effect of angiomotin on endothelial cell migration, we speculate that angiostatin antagonizes angiomotin function by inhibiting the formation of complexes with other proteins.
Acknowledgments
We thank Dr. Arturo Galvani for generating angiomotin antiserum and for real time PCR screening of angiomotin mRNA levels in cell lines and tissues, Dr. Peter Biberfeld for help with immunocytochemical analysis of angiomotin in placenta, and Dr. Vladimir Kashuba for helpful advice.
This study was supported from grants from the Swedish Cancer Society, The Gunnar Nilsson Cancer Foundation, and from Pharmacia & Upjohn, Oncology, Nerviano. O. Matvijenko was supported by grants from Svenska Institutet.
Footnotes
B. Troyanovsky and T. Levchenko contributed equally to this work.
Abbreviations used in this paper: β-gal, β-galactosidase; bFGF, basic fibroblast growth factor; FAK, focal adhesion kinase; GFP, green fluorescent protein; GST; glutathione S-transferase; HUVEC, human umbilical vein endothelial cell; MAE, mouse aortic endothelial; RT, reverse transcription.
References
- Ahmed A., Li X.F., Dunk C., Whittle M.J., Rushton D.I., Rollason T. Colocalisation of vascular endothelial growth factor and its Flt-1 receptor in human placenta. Growth Factors. 1995;12:235–243. doi: 10.3109/08977199509036883. [DOI] [PubMed] [Google Scholar]
- Barendsz-Janson A.F., Griffioen A.W., Muller A.D., van Dam-Mieras M.C., Hillen H.F. In vitro tumor angiogenesis assaysplasminogen lysine binding site 1 inhibits in vitro tumor-induced angiogenesis. J. Vasc. Res. 1998;35:109–114. doi: 10.1159/000025572. [DOI] [PubMed] [Google Scholar]
- Bastaki M., Nelli E.E, Dell'Era P., Rusnati M., Molinari-Tosatti M.P., Parolini S., Auerbach R., Ruco L.P., Possati L., Presta M. Basic fibroblast growth factor-induced angiogenic phenotype in mouse endothelium. A study of aortic and microvascular endothelial cell lines. Arterioscler. Thromb. Vasc. Biol. 1997;17:454–464. doi: 10.1161/01.atv.17.3.454. [DOI] [PubMed] [Google Scholar]
- Bergers G., Javaherian K., Lo K.M., Folkman J., Hanahan D. Effects of angiogenesis inhibitors on multistage carcinogenesis in mice. Science. 1999;284:808–812. doi: 10.1126/science.284.5415.808. [DOI] [PubMed] [Google Scholar]
- Bouck N., Stellmach V., Hsu S.C. How tumors become angiogenic. Adv. Cancer Res. 1996;69:135–174. doi: 10.1016/s0065-230x(08)60862-3. [DOI] [PubMed] [Google Scholar]
- Cao Y., Ji R.W., Davidson D., Schaller J., Marti D., Sohndel S., McCance S.G., O'Reilly M.S., Llinas M., Folkman J. Kringle domains of human angiostatin. Characterization of the anti-proliferative activity on endothelial cells. J. Biol. Chem. 1996;271:29461–29467. doi: 10.1074/jbc.271.46.29461. [DOI] [PubMed] [Google Scholar]
- Cao Y., Chen A., An S.S.A., Ji R.W., Davidson D., Llinás M. Kringle 5 of plaminogen is a novel inhibitor of endothelial cell growth. J. Biol. Chem. 1997;272:22924–22928. doi: 10.1074/jbc.272.36.22924. [DOI] [PubMed] [Google Scholar]
- Claesson-Welsh L., Welsh M., Ito N., Anand-Apte B., Soker S., Zetter B., O'Reilly M., Folkman J. Angiostatin induces endothelial cell apoptosis and activation of focal adhesion kinase independently of the integrin-binding motif RGD. Proc. Natl. Acad. Sci. USA. 1998;95:5579–5583. doi: 10.1073/pnas.95.10.5579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Das B., Mondragon M.O., Sadeghian M., Hatcher V.B., Norin A.J. A novel ligand in lymphocyte-mediated cytotoxicityexpression of the beta subunit of H+ transporting ATP synthase on the surface of tumor cell lines. J. Exp. Med. 1994;180:273–281. doi: 10.1084/jem.180.1.273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dhanabal M., Ramchandran R., Waterman M.J., Lu H., Knebelmann B., Segal M., Sukhatme V.P. Endostatin induces endothelial cell apoptosis. J. Biol. Chem. 1999;274:11721–11726. doi: 10.1074/jbc.274.17.11721. [DOI] [PubMed] [Google Scholar]
- Dunk C., Shams M., Nijjar S., Rhaman M., Bussolati B., Ahmed A. Angiopoietin-1 and Angiopoietin-2 activate trophoblast Tie-2 to promote growth and migration during placental development. Am. J. Pathol. 2000;156:2185–2199. doi: 10.1016/S0002-9440(10)65089-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Folkman J. Angiogenesis in cancer, vascular, rheumatoid and other diseases. Nat. Med. 1995;1:27–31. doi: 10.1038/nm0195-27. [DOI] [PubMed] [Google Scholar]
- Griscelli F., Li H., Bennaceur-Griscelli A., Soria J., Opolon P., Soria C., Perricaudet M., Yeh P., Lu H. Angiostatin gene transferinhibition of tumor growth in vivo by blockage of endothelial cell proliferation associated with a mitosis arrest. Proc. Natl. Acad. Sci. USA. 1998;95:6367–6372. doi: 10.1073/pnas.95.11.6367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Good D.J., Polverini P., Rastinejad F., Le Beau M.M., Lemons R.S., Frazier W.A., Bouck N.P. A tumor suppressor-dependent inhibitor of angiogenesis is immunologically and functionally indistinguishable from a fragment of thrombospondin. Proc. Natl. Acad. Sci. USA. 1990;87:6624–6628. doi: 10.1073/pnas.87.17.6624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hajjar K.A., Guevara C.A., Lev E., Dowling K., Chacko J. Interaction of the fibrinolytic receptor, annexin II, with the endothelial cell surface. Essential role of endonexin repeat 2. J. Biol. Chem. 1996;271:21652–21659. doi: 10.1074/jbc.271.35.21652. [DOI] [PubMed] [Google Scholar]
- Hanahan D., Folkman J. Patterns and emerging mechanisms of the angiogenic switch during tumorigenesis. Cell. 1996;86:353–364. doi: 10.1016/s0092-8674(00)80108-7. [DOI] [PubMed] [Google Scholar]
- Holmgren L., O'Reilly M.S., Folkman J. Dormancy of micrometastasesbalanced proliferation and apoptosis in the presence of angiogenesis suppression. Nat. Med. 1995;1:149–153. doi: 10.1038/nm0295-149. [DOI] [PubMed] [Google Scholar]
- Ji W.R., Castellino F.J., Chang Y., Deford M.E., Gray H., Villarreal X., Kondri M.E., Marti D.N., Llinas M., Schaller J. Characterization of kringle domains of angiostatin as antagonists of endothelial cell migration, an important process in angiogenesis. FASEB J. 1998;12:1731–1738. doi: 10.1096/fasebj.12.15.1731. [DOI] [PubMed] [Google Scholar]
- Jimenez B., Volpert O.V., Crawford S.E., Febbraio M., Silverstein R.L., Bouck N. Signals leading to apoptosis-dependent inhibition of neovascularization by thrombospondin-1. Nat. Med. 2000;6:41–48. doi: 10.1038/71517. [DOI] [PubMed] [Google Scholar]
- Kundra V., Anand-Apte B., Feig L.A., Zetter B.R. The chemotactic response to PDGF-BBevidence of a role for Ras. J. Cell Biol. 1995;130:725–731. doi: 10.1083/jcb.130.3.725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lucas R., Holmgren L., Garcia I., Jimenez B., Mandriota S.J., Borlat F., Sim B.K., Wu Z., Grau G.E., Shing Y. Multiple forms of angiostatin induce apoptosis in endothelial cells. Blood. 1998;92:4730–4741. [PubMed] [Google Scholar]
- Miles L.A., Dahlberg C.M., Plescia J., Felez J., Kato K., Plow E.F. Role of cell-surface lysines in plasminogen binding to cellsidentification of alpha-enolase as a candidate plasminogen receptor. Biochemistry. 1991;30:1682–1691. doi: 10.1021/bi00220a034. [DOI] [PubMed] [Google Scholar]
- Moser T.L., Stack M.S., Asplin I., Enghild J.J., Hojrup P., Everitt L., Hubchak S., Schnaper H.W., Pizzo S.V. Angiostatin binds ATP synthase on the surface of human endothelial cells. Proc. Natl. Acad. Sci. USA. 1999;96:2811–2816. doi: 10.1073/pnas.96.6.2811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Reilly M.S., Holmgren L., Chen C., Folkman J. Angiostatin induces and sustains dormancy of human primary tumors in mice. Nat. Med. 1996;2:689–692. doi: 10.1038/nm0696-689. [DOI] [PubMed] [Google Scholar]
- O'Reilly M.S., Holmgren L., Shing Y., Chen C., Rosenthal R.A., Moses M., Lane W.S., Cao Y., Sage E.H., Folkman J. Angiostatina novel angiogenesis inhibitor that mediates the suppression of metastases by a Lewis lung carcinoma. Cell. 1994;79:315–328. doi: 10.1016/0092-8674(94)90200-3. [DOI] [PubMed] [Google Scholar]
- Parangi S., O'Reilly M., Christofori G., Holmgren L., Grosfeld J., Folkman J., Hanahan D. Antiangiogenic therapy of transgenic mice impairs de novo tumor growth. Proc. Natl. Acad. Sci. USA. 1996;93:2002–2007. doi: 10.1073/pnas.93.5.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sambrook, J., E.F. Fritsch, and T. Maniatis. 1989. Molecular Cloning. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
- Sudol M. From Src Homology domains to other signaling modulesproposal of the ‘protein recognition code’. Oncogene. 1998;17:1469–1474. doi: 10.1038/sj.onc.1202182. [DOI] [PubMed] [Google Scholar]
- Yamaguchi N., Anand-Apte B., Lee M., Sasaki T., Fukai N., Shapiro R., Que I., Lowik C., Timpl R., and Olsen B.R. Endostatin inhibition of VEGF-induced endothelial cell migration and tumor growth independently of zinc binding. EMBO (Eur. Mol. Biol. Organ.) J. 1999;18:4414–4423. doi: 10.1093/emboj/18.16.4414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu J., Kahn R.C. Analysis of a peptide hormone-receptor interaction in the yeast two-hybrid system. Proc. Natl. Acad. Sci. USA. 1997;94:13063–13068. doi: 10.1073/pnas.94.24.13063. [DOI] [PMC free article] [PubMed] [Google Scholar]