

Abstract
It has been hypothesized that in the mature nerve terminal, interactions between synapsin and actin regulate the clustering of synaptic vesicles and the availability of vesicles for release during synaptic activity. Here, we have used immunogold electron microscopy to examine the subcellular localization of actin and synapsin in the giant synapse in lamprey at different states of synaptic activity. In agreement with earlier observations, in synapses at rest, synapsin immunoreactivity was preferentially localized to a portion of the vesicle cluster distal to the active zone. During synaptic activity, however, synapsin was detected in the pool of vesicles proximal to the active zone. In addition, actin and synapsin were found colocalized in a dynamic filamentous cytomatrix at the sites of synaptic vesicle recycling, endocytic zones. Synapsin immunolabeling was not associated with clathrin-coated intermediates but was found on vesicles that appeared to be recycling back to the cluster. Disruption of synapsin function by microinjection of antisynapsin antibodies resulted in a prominent reduction of the cytomatrix at endocytic zones of active synapses. Our data suggest that in addition to its known function in clustering of vesicles in the reserve pool, synapsin migrates from the synaptic vesicle cluster and participates in the organization of the actin-rich cytomatrix in the endocytic zone during synaptic activity.
Keywords: synapsin; actin; synapse; neurotransmission; endocytosis
Introduction
Synaptic transmission depends on a highly regulated, rapid, and sustainable cycle of vesicle exo- and endocytosis. In neurons, secretory vesicles filled with neurotransmitter are clustered at release sites. After fusion at the active zone, vesicles are reformed locally, primarily via clathrin-mediated endocytosis, and then rejoin the synaptic vesicle cluster (for review see Slepnev and De Camilli, 2000). Evidence linking actin dynamics to presynaptic functions has been reported. Actin was localized to nerve terminals and implicated in several stages of the synaptic vesicle cycle, such as vesicle clustering, release, and endocytosis (Doussau and Augustine, 2000). In recent imaging studies, it has been shown that the presynaptic actin cytoskeleton undergoes plastic modification during synaptic activity (Morales et al., 2000; Colicos et al., 2001; Sankaranarayanan et al., 2003). At the ultrastructural level, perturbation of actin dynamics revealed an activity-dependent cytoskeletal matrix involved in the delivery of recycled synaptic vesicles back to the vesicle cluster (Shupliakov et al., 2002).
One of the first actin-binding proteins to be identified in nerve terminals was synapsin (for review see Greengard et al., 1994). Synapsins are a family of neuron-specific proteins that are bound to synaptic vesicles. Three highly conserved mammalian synapsin genes give rise to several protein isoforms via alternative splicing (Kao et al., 1999). Although distinct functions of the synapsin gene products are not known, the dominant isoforms in the mature nerve terminal are synapsins Ia, Ib, IIa, and IIb (Südhof et al., 1989; Pieribone et al., 2002). Acute perturbation of synapsin function in a variety of in vivo systems has demonstrated its critical role in regulating neurotransmission (Llinas et al., 1985, 1991; Pieribone et al., 1995; Hilfiker et al., 1998). Mice genetically deficient in synapsin I, synapsin II, or both are viable, but synaptic vesicle clustering is almost completely abolished and the number of synaptic vesicles is drastically reduced (Li et al., 1995; Takei et al., 1995). These morphological defects result in significant deficits in synaptic plasticity and behavior (Rosahl et al., 1993, 1995; Li et al., 1995; Ryan et al., 1996; Terada et al., 1999). To understand the mechanism by which synapsin regulates neurotransmitter release, its interactions with actin have been examined in vitro (Greengard et al., 1994). As an actin-nucleating agent, synapsin lowers the critical concentration of actin necessary for filament formation (Benfenati et al., 1992; Fesce et al., 1992; Valtorta et al., 1992). Furthermore, synaptic vesicles containing synapsin are able to promote globular actin (G-actin)* nucleation and filamentous actin (F-actin) polymerization, whereas vesicles stripped of synapsin are unable to do so (Benfenati et al., 1989). Synapsin also stabilizes and bundles F-actin in a phosphorylation-dependent manner (Bahler et al., 1989; Jovanovic et al., 1996). Finally, synapsin binds simultaneously to both actin and synaptic vesicles (Benfenati et al., 1992). Based upon a variety of in vivo and in vitro data, these synapsin–actin interactions have been proposed to take place in vesicle clusters of synapses. Upon depolarization, synapsin changes its conformation and dissociates from actin, thereby freeing synaptic vesicles for release (Greengard et al., 1994; Esser et al., 1998). Additional functions of synapsins have also been proposed (Esser et al., 1998; Hosaka and Südhof, 1999).
Previously, we examined the effects of synapsin- (Pieribone et al. 1995) and actin-perturbing compounds (Shupliakov et al., 2002) on the structure and function of a central nervous system synapse. Reagents that interfere with synapsin function caused a disruption of the synaptic vesicle cluster. Surprisingly, actin-perturbing reagents did not directly disrupt the organization of the synaptic vesicle cluster. Rather, these reagents were more effective in the area lateral to the active zone, where they altered the trafficking of recycled vesicles back to the cluster. Thus, our experiments revealed a dynamic cytoskeletal matrix surrounding the vesicle cluster that assembles during synaptic activity and participates in the recycling of synaptic vesicles. This cytomatrix contained actin, as demonstrated by light microscopy after injection of Oregon green phalloidin into the synapse (Shupliakov et al., 2002). In light of these findings, we sought to localize both actin and synapsin in a central nervous system synapse using immunogold electron microscopic techniques. We performed these studies in the reticulospinal synapse of the lamprey, which is uniquely suited to relate ultrastructural synaptic organization with function (Shupliakov and Brodin, 2000). Within the same axon, synapses are often separated by large regions of axoplasm, making it possible to attribute recycling intermediates to a particular synapse. These features make this preparation particularly well suited for the subcellular localization of synaptic proteins.
Results
Immunolocalization of actin in the synapse
A monoclonal anti-actin antibody was used to label lamprey synapses at rest and during synaptic activity. Fig. 1 A shows a giant reticulospinal synapse at rest, labeled using postembedding immunogold techniques. Immunogold particles are predominantly associated with the postsynaptic density of the synapse. Only a small number of gold particles were detected within the vesicle cluster and the axoplasmic matrix. Labeling was also consistently associated with regions lateral to the active zones (Fig. 1 A). Small amounts of filamentous material were often observed in these regions. On several sections containing dense projections, gold particles were associated with the active zone (Fig. 1 A). This is in agreement with recent findings indicating that actin may be associated with these structures (Phillips et al., 2001). Preabsorption of the antibody with actin abolished the labeling (not depicted). Quantitative evaluation of actin immunoreactivity in resting synapses is shown in Table I.
Figure 1.
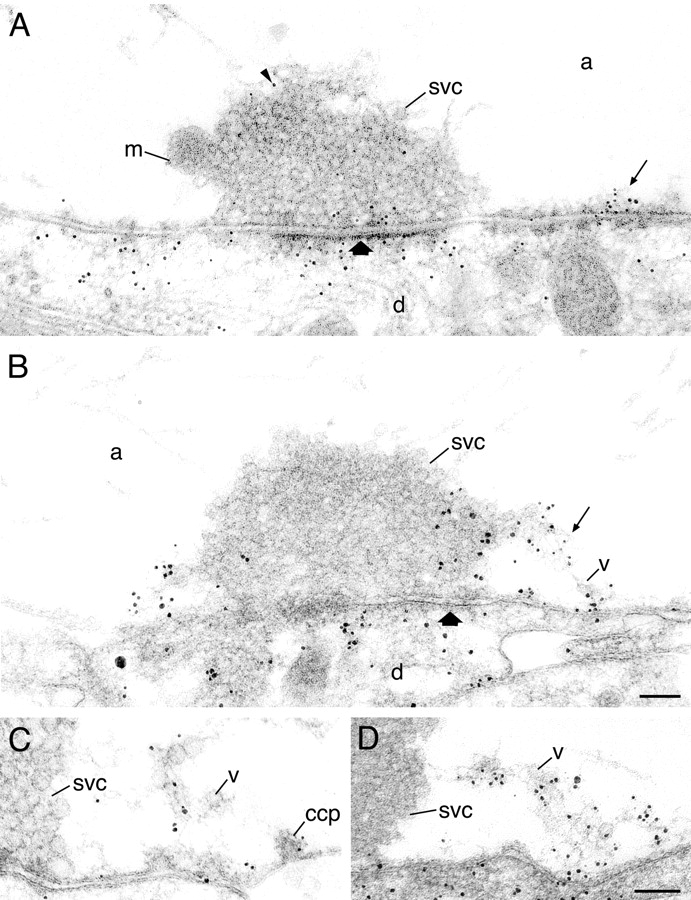
Ultrastructural localization of actin in resting and stimulated reticulospinal synapses. (A) Electron micrograph of a synapse in an unstimulated specimen labeled with an anti-actin antibody. Arrowhead indicates gold particle. Thick arrow indicates active zone. (B) Immunogold localization of actin in a synapse stimulated with action potentials at 5 Hz. Thin arrow indicates filamentous matrix in the endocytic zone. (C and D) Endocytic zones in two different stimulated synapses. Note clathrin-coated intermediates (ccp) at the presynaptic membrane in stimulated axons. m, mitochondrion; svc, synaptic vesicle cluster; v, vesicles in the filamentous matrix; d, dendrite; a, axoplasmic matrix. Bars, 100 nm.
Table I. Quantitative analysis of actin immunogold labeling.
| Profile | Unstimulated | 5 Hz stimulated | ||
|---|---|---|---|---|
|
n
|
Mean ± SEM
|
n
|
Mean ± SEM
|
|
| particles/μm2 | particles/μm2 | |||
| Endocytic zone | 9 | 21.0 ± 11 | 9 | 312 ± 63 |
| Synaptic vesicle cluster | 5 | 6.22 ± 2.2 | 6 | 6.45 ± 3.5 |
| Axoplasm | 5 | 1.25 ± 0.38 | 6 | 1.33 ± 0.54 |
| Postsynaptic density | 5 | 338 ± 23 | 5 | 459 ± 93 |
The subcellular distribution of actin was examined in synapses stimulated at 5 Hz for 20 min (Table I). As shown previously with this stimulation protocol (Shupliakov et al., 1997), coated endocytic intermediates were observed lateral to the active zone (Fig. 1, B–D). These intermediates, as well as uncoated small clear vesicles, were often found in contact with the filamentous material that was decorated by anti-actin antibodies (Fig. 1, C and D). A significant increase in actin immunolabeling at the lateral sides of the active zone was observed, as compared with synapses fixed at rest (P < 0.0001, n = 9, unpaired t test). This increase was associated with a proliferation of the filamentous material, which was heavily decorated with gold particles (Fig. 1, B–D). As in synapses fixed at rest, gold particles were associated with the synaptic vesicle cluster; most of this labeling, however, was found in the peripheral regions of the cluster (Fig. 1 B).
Immunolocalization of synapsin at rest and during synaptic activity
The antisynapsin antibodies used in our experiments produced robust punctate immunoreactivity on cryostat sections of the lamprey spinal cord, reflecting the discrete localization of synapses in this tissue. Single puncta were detected consistently on the internal surface of reticulospinal axons, suggesting a synaptic labeling (Fig. 2 A).
Figure 2.
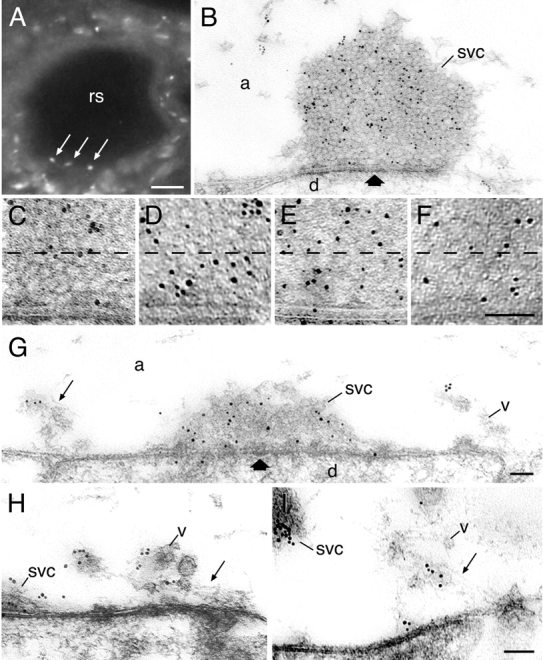
Redistribution of synapsin in resting and stimulated reticulospinal synapses. (A) Transverse section of a reticulospinal axon (rs) stained with lamprey-specific synapsin antibodies using immunofluorescence. Punctate labeling (arrows) is localized close to the surface of the axon. (B) The ultrastructural localization of synapsin at rest. Gold particles are associated with the synaptic vesicle cluster. Thick arrows indicate active zone. (C–F) An increase in synapsin labeling in the area within 100 nm from the presynaptic membrane in the vesicle cluster at different states: (C) rest; (D) 5 Hz action potential stimulation; (E) high K+ stimulation; (F) control, SV2 immunolabeling at rest. (G–I) Electron micrographs of synapses stimulated by action potentials at 5 Hz for 20 min and labeled for synapsin. Thin arrow indicates filamentous matrix. Note that synapsin is associated with the filamentous cytomatrix and vesicles in the endocytic zone. svc, synaptic vesicle cluster; v, vesicles in the filamentous matrix; d, dendrite; a, axoplasmic matrix. Bars: (A) 20 μm; (F) 100 nm, also for C–E; (G) 100 nm, also for B; (I) 100 nm, also for H.
To examine the distribution of synapsin immunoreactivity at various stages of the synaptic vesicle cycle at the ultrastructural level, we stained reticulospinal synapses with synapsin antibodies at rest and during stimulation. Under resting conditions, synapsin was present predominantly on clustered vesicles located at a distance >100 nm from the presynaptic active zone (Fig. 2, B and C; Pieribone et al., 1995). In synapses stimulated at 5 Hz or exposed to elevated potassium Ringer's solution for 15 min, the distribution of synapsin immunolabeling in the 100-nm zone of the vesicle cluster immediately adjacent to the presynaptic membrane increased twofold (Fig. 2, D and E; Fig. 3; P < 0.01, unpaired t test). An even distribution of gold particles was observed in the distal portion of the cluster. By contrast, labeling of an integral vesicle membrane protein (synaptic vesicle protein 2 [SV2]) was uniformly distributed throughout the cluster at rest (Fig. 2 F; Fig. 3; P > 0.05, n = 16, unpaired t test). These data indicate that synapsin redistributes within the cluster during synaptic activity.
Figure 3.

Quantitative evaluation of gold particle distribution at active zones at rest and during synaptic activity. The bar graph shows the ratio of gold particle densities between 0–100 nm and 100–200 nm from the active zone. At rest, n = 8; 5 Hz, n = 13, and K+ stimulation, n = 12.
In stimulated synapses, synapsin immunoreactivity was observed within the filamentous cytomatrix formed at the lateral edges of active zones during stimulation (Fig. 2 G; Fig. 4, A and B). Synapsin labeling was associated with synaptic vesicles present in the cytomatrix, which appeared to be in transit to the vesicle cluster (Fig. 2, G–I; Fig. 4, A and B). Only occasional gold particles were found at synaptic endocytic zones at rest.
Figure 4.


3-D distribution of synapsin in stimulated reticulospinal synapse. (A) Electron micrograph of one serial section of a reticulospinal synapse stimulated at 5 Hz for 20 min, shown reconstructed in B. Synapsin is present on vesicles (v) in the filamentous cytomatrix (arrow). svc, synaptic vesicle cluster; v, vesicles in the filamentous matrix; d, dendrite; a, axoplasmic matrix. ccp, clathrin-coated pit. This arrow indicates active zone. (B) 3-D reconstruction of 10 serial ultrathin sections. The area occupied by actin-rich filamentous cytomatrix lateral to the vesicle cluster is shown in yellow. The plasma membrane is depicted in green. All synaptic vesicles are shown in red. Colloidal gold particles are indicated by white spheres. Clathrin-coated endocytic intermediates are shown as flat blue discs on the plasma membrane. Bar, 100 nm.
To overview the spatial distribution of actin and synapsin in the active synapse, we performed a three-dimensional (3-D) reconstruction of ten 90-nm serial sections (Fig. 4, A and B). The reconstruction contained ∼900 clustered synaptic vesicles, roughly equivalent to one half of an average reticulospinal synaptic vesicle cluster, and 40 synaptic vesicles enmeshed within the filamentous cytomatrix. The filaments were distinctly localized to endocytic zones of the plasma membrane and formed a halo-like structure spanning from the plasma membrane to the vesicle cluster. The ultrastructural architecture of the actin-rich cytomatrix thus correlates well with fluorescent phalloidin rings previously observed around vesicle clusters (Shupliakov et al., 2002; Sankaranarayanan et al., 2003). The majority of synaptic vesicles enmeshed in the cytomatrix were labeled for synapsin, as were the clustered synaptic vesicles.
To directly test if synapsin and actin are colocalized around the synapse, double labeling experiments were performed. Sections were incubated in a mixture of rabbit anti–synapsin domain D antibodies and a mouse anti-actin antibody. The primary antibodies were detected with the corresponding secondary antibodies coupled to 5- and 10-nm gold particles, respectively. In stimulated synapses, labeling for both synapsin and actin was observed within the filamentous cytomatrix and on synaptic vesicles recycling back to the cluster (Fig. 5, A–D).
Figure 5.
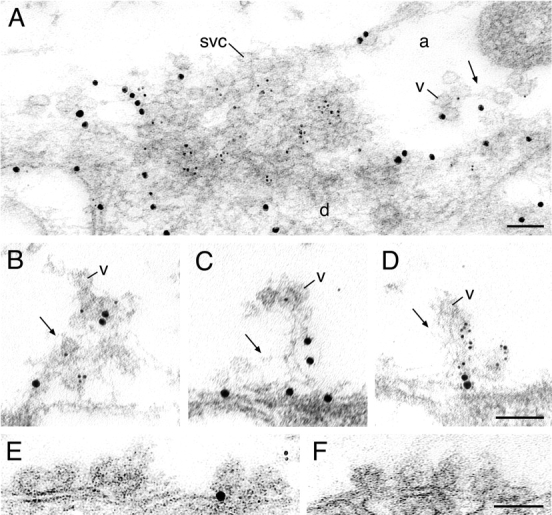
Distribution of synapsin and actin in active synapses as revealed by double labeling. (A) Electron micrograph of a synapse stimulated with high K+ for 30 min. Large gold particles (enhanced 10 nm) denote actin and small (enhanced 5 nm) denote synapsin immunoreactivity. (B–D) Regions of endocytic zones, containing filamentous cytomatrix in three synapses stimulated at 5 Hz for 20 min and labeled with immunogold as in A. (E and F) Electron micrographs of endocytic regions containing clathrin-coated intermediates in synapses stimulated with high K+ for 30 min. svc, synaptic vesicle cluster; v, vesicles in the filamentous matrix; d, dendrite; a, axoplasmic matrix. Bars, 100 nm.
To address the possible pathway by which synapsin becomes associated with the actin cytomatrix and recycling vesicles, we investigated if synapsin labeling is associated with clathrin-coated intermediates. Both action potential–stimulated (5 Hz) and K+-stimulated synapses were examined. In neither case did we detect labeling of coated intermediates or of the plasma membrane. This was particularly evident in preparations stimulated with high K+, in which numerous endocytic intermediates occurred at the endocytic zone (Fig. 5, E and F; see also Fig. 2 G). Ultrathin sections were stained with both synapsin and SV2 antibodies (Fig. 6, A–C). As in previous experiments, SV2 antibodies efficiently labeled synaptic vesicle clusters (Fig. 6 A). In addition, SV2 immunolabeling was observed on coated intermediates as well as on the plasma membrane in the endocytic zone (Fig. 6, B and C). These results indicate that synapsin is not linked to the vesicle membrane before endocytosis, but associates with the recycled vesicle after it has been released into the cytoplasm.
Figure 6.

Localization of synapsin in the endocytic zone. Electron micrographs from synapses double labeled with synapsin (small particles) and SV2 (large particles) antibodies. Note an even distribution of both antigens in the vesicle cluster in A and the accumulation of only SV2 immunoreactivity on the plasma membrane and endocytic intermediates in B and C. Bar, 100 nm.
Microinjection of synapsin antibodies results in a decrease of the actin-rich filamentous matrix at endocytic zones
The activity-dependent migration of synapsin to the endocytic zone and its colocalization with actin suggest that it may be involved in the organization of actin in this region. To test this possibility, antisynapsin antibodies were microinjected into giant reticulospinal axons to perturb the function of synapsin. G-304 antibodies, which disrupt the distal pool of synaptic vesicles in lamprey synapses (Pieribone et al., 1995), were used in these experiments. After antibody microinjection, control and injected axons were subjected to high-frequency action potential stimulation (18 Hz) for 6 min to induce synaptic vesicle recycling and then immediately fixed. The ultrastructure of synapses from injected and control axons was studied in serial ultrathin sections. In agreement with our previous studies, a significant decrease in the number of vesicles in injected synapses was observed, whereas large synaptic vesicle clusters were still present in control synapses (Fig. 7, A and B). In both groups of synapses, a similar number of clathrin-coated pits was observed within the endocytic zone (Fig. 7, A–D). In control synapses, a distinct filamentous cytomatrix was consistently observed in the vicinity of the clathrin-coated intermediates (Fig. 7 C). In antibody-injected synapses, however, this cytomatrix was significantly reduced and in many sections completely absent (Fig. 7 D).
Figure 7.
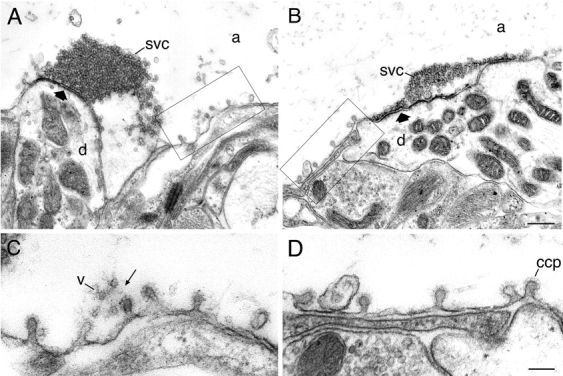
Disruption of the actin-rich cytomatrix by microinjection of synapsin antibodies. (A) A reticulospinal synapse in an uninjected axon subjected to high frequency stimulation at 18 Hz for 6 min. Boxed area is shown in C at higher magnification. (B) A synapse from an axon from the same preparation injected with synapsin antibodies. Boxed area is shown in D at higher magnification. Thick arrow indcates active zone, and thin arrow indicates filamentous matrix. svc, synaptic vesicle cluster; v, vesicles in the filamentous matrix; d, dendrite; a, axoplasmic matrix. Bars, 100 nm.
Discussion
The present results support a novel role for actin and synapsin in neurotransmission. Using immunogold electron microscopy to study active synapses, we have identified actin in the endocytic zone surrounding vesicle clusters. Furthermore, we have demonstrated an increase in actin immunoreactivity in this zone during neurotransmitter release. These observations are in agreement with our previous demonstration of an accumulation of phalloidin labeling in the periphery of the vesicle cluster during synaptic activity (Shupliakov et al., 2002) and with recent light-microscopic studies on cultured hippocampal neurons transfected with GFP–actin (Sankaranarayanan et al., 2003).
We also detected low concentrations of actin within the cluster. As the antibody we used to detect actin does not distinguish between G- and F-actin, we speculate that the relatively low levels of actin immunolabeling in the cluster may be due to a failure to detect epitopes of G-actin hidden by complex-bound monomers. However, as filamentous material at both the endocytic and active zones was efficiently labeled by these anti-actin antibodies, it seems unlikely that actin filaments are concentrated within the cluster.
Previous studies have shown synapsin to dissociate from clustered synaptic vesicles in an activity-dependent manner (Torri-Tarelli et al., 1992; Hilfiker et al., 1999). Most recently, this was imaged in real-time using GFP–synapsin chimeras in mammalian hippocampal neurons (Chi et al., 2001, 2003). However, the precise fate of dissociated synapsin in these studies could not be determined, as the studies were limited by the resolution of light microscopy. Presently, we find that upon activity-dependent dissociation of synapsin from vesicles, its migration is delimited by the actin-rich cytomatrix at the endocytic zones surrounding vesicle clusters. Furthermore, during synaptic activity, synapsin was associated with the actin-rich cytomatrix in both the presence and absence of synaptic vesicles. Disruption of synapsin function by antibody microinjection resulted in a marked reduction of the actin-rich cytomatrix in the endocytic zone.
Our results lead us to propose a model for the interaction of synapsin and actin at the sites of synaptic vesicle recycling. Synapsin–synapsin and synapsin–G-actin interactions may be predominantly responsible for vesicle clustering in the reserve pool (Greengard et al., 1994). Upon activation and vesicle fusion, synapsin disassociates from synaptic vesicles. Synapsin then accumulates in the endocytic region and interacts with a soluble pool of G-actin, where the actin-rich cytomatrix is formed. After endocytosis and uncoating of clathrin from nascent vesicles, a pool of synapsin bound to the actin-rich cytomatrix reassociates with vesicle membranes. Synapsin's ability to nucleate, bind to, and bundle actin may be important in regulating the dynamic formation of the actin matrix, thereby coordinating its own reassociation with synaptic vesicles. Therefore, we propose that the well-established synapsin–actin interactions may be more relevant to processes outside of the vesicle cluster. This does not exclude the possibility that synapsin and actin also function in the regulation of release. In stimulated synapses, a migration of synapsin within the vesicle cluster was observed close to the active zone. Actin immunoreactivity was also found in this region of active synapses, thus supporting this possibility.
The absence of synapsin immunoreactivity on endocytic intermediates in our experiments is consistent with previous biochemical studies of the nerve terminal. In subcellular fractionation experiments, the distributions of synapsin and clathrin were shown to be nonoverlapping (Huttner et al., 1983; Maycox et al., 1992). The uncoating of vesicles has been shown to take place within the endocytic zone, and perturbation of synaptojanin–endophilin interactions results in an increased number of coated vesicles within the lateral actin matrix, implying a role for these proteins in normal uncoating and the translocation of vesicles to the actin matrix (Gad et al., 2000). The present colocalization of actin and synapsin within the endocytic zone of the active nerve terminal places all necessary components of the model together in a spatially and temporally regulated manner.
How could this putative mechanism work? It has been shown that synapsin's interaction with actin is regulated by phosphorylation. Synapsin is phosphorylated by calcium/calmodulin-dependent protein kinase II (CaMKII). In addition, it is also phosphorylated by MAPK at three known sites (Jovanovic et al., 1996), which regulate neurotransmitter release (Jovanovic et al., 2000). Phosphorylation of synapsin by MAPK, like CaMKII, decreases synapsin's abilities to bind to actin (Jovanovic et al., 1996). Inhibition of MAPK by pharmacological agents inhibits synapsin phosphorylation and concomitantly reduces neurotransmitter release (Jovanovic et al., 2000). However, in contrast to the CaMKII sites, the MAPK sites on synapsin appear to be phosphorylated under basal conditions and dephosphorylated upon nerve terminal depolarization (Jovanovic et al., 2001). This dephosphorylation is dependent on calcineurin. Calcineurin has been demonstrated to be a key regulator of several proteins involved in the endocytic pathway (Cousin et al., 2001). Our present findings of synapsin associated with actin outside of vesicle clusters during synaptic activity may therefore correlate to a pool of synapsin regulated by the MAPK pathway. This pool of synapsin would be dephosphorylated during synaptic activity and therefore retain its affinity for actin during vesicle recycling.
Several proteins regulated by calcineurin have been shown to interact with the proline-rich domain of synapsin in vitro. Many of these proteins have been implicated in or localized to stages of the synaptic vesicle cycle outside of the vesicle cluster. These proteins include PI3 kinase (Onofri et al., 2000), Grb2 (McPherson et al., 1994; Vaccaro et al., 1997), Src (Onofri et al., 1997; Foster-Barber and Bishop, 1998; Zhao et al., 2000), SH3p4/endophilin 1 (Onofri et al., 2000), SH3p13/endophilin 3 (Ringstad et al., 1997, 1999), amphiphysins I and II (Onofri et al., 2000), and syndapin (Qualmann et al., 1999). In some cases, these interactions can modulate the association of actin and synapsin. For example, the binding of amphiphysins to synapsin decreased synapsin's ability to promote actin polymerization (Onofri et al., 2000). Furthermore, synapsin can interact with other actin-binding proteins. Synapsin was shown to associate with the brain-specific form of profilin, together with dynamin and actin, whereas clathrin was found to associate with the more ubiquitous profilin I (Witke et al., 1998). Abp1 is a protein that binds directly to both dynamin and actin, thus providing an additional link between the actin cytoskeleton and the endocytic machinery (Kessels et al., 2001). Previously, it was difficult to postulate the relevance of synapsin's interactions with proteins involved in endocytosis to the mature nerve terminal, as synapsin was implicated only in vesicle clustering. Localizing synapsin to the endocytic region of the synapse provides a physiological context for many of these biochemical interactions. One of the possible functions of the actin/synapsin-rich cytomatrix could be the compartmentalization of endocytic proteins during synaptic vesicle recycling.
Our studies in lamprey and recent experiments in mammalian synapses show that an actin-rich matrix is formed during synaptic activity, but the precise role of the cytomatrix in the transport of the newly formed vesicles currently remains unclear (Shupliakov et al., 1998; Sankaranarayanan et al., 2003). A role of actin filaments in vesicle propulsion has been suggested by studies in nonneuronal systems (Merrifield et al., 1999; Rozelle et al., 2000), whereas actin in the nerve terminal has long been postulated to serve as a scaffold for synaptic vesicles or regulatory proteins (Greengard et al., 1994). Although more experiments are needed to clarify the role of actin in synaptic vesicle recycling, our experiments indicate that synaptic vesicles are associated with the actin/synapsin-rich cytomatrix on their way back to the vesicle cluster. This association may promote the proper translocation of vesicles from sites of endocytosis to sites of release. Small synaptic boutons are probably less dependent on this mechanism because their active zones are surrounded by the presynaptic membrane, creating a physical border that prevents the diffusion of vesicles during the synaptic vesicle cycle.
Several studies indicate that the perturbation of synapsins affects the reserve pool of synaptic vesicles (Pieribone et al., 1995; Hilfiker et al., 1998). Additional studies suggest that synapsin may also play a role in regulating the readily releasable pool (Rosahl et al., 1995; Hilfiker et al., 1999). The most robust effects of functional or genetic deletion of synapsin in vivo have been observed with stimulation conditions that require sustained or high frequency neurotransmitter release, indicating a role for synapsin in regulating the efficiency of synaptic vesicle cycling (Li et al., 1995; Pieribone et al., 1995; Rosahl et al., 1995). In the present study, disruption of synapsin function by microinjection of antisynapsin antibodies in active synapses resulted in a dramatic reduction of the lateral actin matrix and a concomitant reduction in the number of synaptic vesicles in the cluster, thus demonstrating that synapsin is intimately associated not only with the vesicles in the cluster but also with the actin machinery of synaptic vesicle recycling. Taken together with the extensive literature on biochemical interactions of synapsin, these data support the existence of several pools of synapsin within the nerve terminal that may serve functionally distinct roles at various stages of the synaptic vesicle cycle.
Materials and methods
Antibodies
Two different antibodies were used to study the distribution of synapsin. Lamprey-specific antibodies were raised against the D domain of synapsin I. Lamprey synapsin domain D (Kao et al., 1999) was subcloned into a pGEX vector, and a GST fusion protein was produced. The antibodies were generated by immunizing rabbits with the recombinant protein (Cocalico Biologicals). The antibodies were affinity purified and tested by immunoblotting using standard procedures (Shupliakov et al., 1997). On immunoblots of lamprey spinal cord protein extract, the antibodies detected two bands of the expected molecular weights corresponding to lamprey synapsin Ia and Ib (not depicted; Kao et al., 1999).
Previously characterized G-304 antibodies recognizing lamprey synapsin Ia protein were also used in these experiments (Pieribone et al., 1995). Although both antibodies used showed a similar immunolabeling pattern, the lamprey-specific D-domain antibodies were more efficient in immunogold experiments.
A mouse monoclonal anti-actin antibody (C4) was purchased from ICN Biomedicals and Chemicon International. The antibody was tested on immunoblots of lamprey spinal cord protein extract and revealed a single band of 42 kD molecular mass, the expected size for actin (not depicted). Preabsorption of the C4 antibody with chicken muscle actin (1 mg/ml) overnight at 4°C dramatically decreased labeling of actin-containing structures (not depicted; n = 3 synapses).
The monoclonal mouse anti-SV2 antibody was developed by K.M. Buckley (Buckley and Kelly, 1985) and obtained from the Developmental Studies Hybridoma Bank, developed under the auspices of the National Institute of Child Health and Human Development and maintained by the University of Iowa, Department of Biological Sciences. On a Western blot of lamprey spinal cord extract, it recognized one band of ∼100 kD, consistent with the previously reported size of SV2 (not depicted).
Secondary FITC-conjugated donkey anti–rabbit antibodies were purchased from Jackson ImmunoResearch Laboratories. Immunogold reagents were purchased from Amersham Biosciences.
Dissection, stimulation, microinjection, and fixation procedures
Spinal cords of adult lampreys (Lampetra fluviatilis) were used for all experiments. Animals were housed in a fresh water aerated aquarium maintained at 4°C. Animals were anesthetized and decapitated, and trunk segments of the spinal cord were dissected as described previously (Shupliakov et al., 2002).
Stimulation was applied by an extracellular suction electrode placed at the caudal end of the spinal cord (Brodin et al., 1994). An additional extracellular electrode was placed at the rostral end of the spinal cord to monitor the extracellular spike volley during the stimulation period. Several preparations were stimulated by elevated K+ as described elsewhere (Wickelgren et al., 1985; Gad et al., 1998).
Intraaxonal microinjections of G-304 anti-synapsin antibodies were performed as described previously (Pieribone et al., 1995). Axons were stimulated at a frequency of 18 Hz.
Specimens were fixed in 4% paraformaldehyde (immunofluorescence) or 4% paraformaldehyde/0.5% glutaraldehyde/4% tannic acid in 0.1 M cacodylate buffer at 4°C for 1 h and then incubated in the same fixative without tannic acid at 4°C for 3 h. Preparations were washed in 0.1 M cacodylate buffer, stained en bloc with 2% uranyl acetate, dehydrated with a graded ethanol series, and embedded at −25°C in LR Gold Resin (London Resin Co.). For microinjection experiments, specimens were fixed in 3% glutaraldehyde in 0.1 M phosphate buffer, embedded in Durcupan ACM (Fluka), and processed as described elsewhere (Pieribone et al., 1995; Shupliakov et al., 1997).
Immunofluorescence
14-μm-thick sections were cut on a cryostat, mounted on slides, and stained with antibodies following standard protocols (for example see Schotland et al., 1996).
Postembedding immunogold labeling of ultrathin sections and electron microscopy
Serial ultrathin sections were cut from the LR Gold–embedded tissue with a diamond knife and mounted on mesh grids and Formvar-coated nickel slot grids. Sections were incubated overnight at 4°C with primary antibodies diluted in Tris-phosphate–buffered saline with 1% human serum albumin. Synapsin antibodies were used at 5 μg/ml. The actin antibody was used at a dilution of 1:50. The SV2 antibody was obtained as hybridoma supernatant, concentrated 50-fold, and then used undiluted.
Secondary goat anti–rabbit and goat anti–mouse antibodies conjugated to 5- or 10-nm gold particles were used at a dilution of 1:25–1:50. Grids incubated without primary antibodies were used as controls unless otherwise specified in the text. Gold particles were enhanced using the IntenSE Silver Enhancement Kit (Amersham Biosciences). Sections were counterstained with uranyl acetate and lead citrate and examined in a CM12, CM10 Philips, or Tecnai 12 transmission electron microscope.
Quantitative analysis
Density of gold particles in synaptic regions was obtained as described previously (Pieribone et al., 1995). For quantification of gold particle labeling within the vesicle cluster, the middle section of the synapse was defined as that section in a series where the active zone was at its maximum length. Boxes of 100 nm were drawn from the active zone through the cluster to quantify gold particles in different pools of vesicles. Gold particles were quantified manually in each 100-nm box. Statistical evaluation of the data was performed using Microsoft Excel software.
3-D reconstruction
Serial ultrathin sections of a synapse were subjected to postembedding immunogold electron microscopy, as described above. Sections were photographed, and the contours of objects of interest were traced onto transparent overlays. The contours were then digitized using a Wacom digitizing tablet connected to a G4 Power Macintosh computer. 3-D reconstructions were rendered using the FormZ software (AutoDeSys, Inc.) and imported into Adobe Photoshop 6.0® for final color adjustments and printing.
Acknowledgments
The authors wish to thank L. Daniell and Dr. E. Zotova for technical advice and Drs. J. Morgan, E.A. Lumpkin, and F. Capani for their comments on the manuscript.
This work was supported by Swedish Research Council grants 13473 (O. Shupliakov) and 11287 (L. Brodin), National Institutes of Health grants MH 39327 (P. Greengard), GM07982 (O. Bloom), and NS037823 (V. Pieribone), and the Rockefeller University-Karolinska Institute Exchange Program and Rockefeller University institutional funds (O. Bloom).
O. Bloom's present address is Department of Cell Biology, Yale University School of Medicine, New Haven, CT 06520.
O. Kjaerulff's present address is Institute of Medical Physiology, Copenhagen University, Blegdamsvej 3, DK-2200, Copenhagen, Denmark.
V.A. Pieribone's present address is The John B. Pierce Laboratory, Cellular and Molecular Physiology, Yale University School of Medicine, New Haven, CT 06519.
Footnotes
Abbreviations used in this paper: 3-D, three-dimensional; CaMKII, calmodulin-dependent protein kinase II; F-actin, filamentous actin; G-actin, globular actin; SV2, synaptic vesicle protein 2.
References
- Bahler, M., F. Benfenati, F. Valtorta, A.J. Czernik, and P. Greengard. 1989. Characterization of synapsin I fragments produced by cysteine-specific cleavage: a study of their interactions with F-actin. J. Cell Biol. 108:1841–1849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benfenati, F., F. Valtorta, M. Bahler, and P. Greengard. 1989. Synapsin I, a neuron-specific phosphoprotein interacting with small synaptic vesicles and F-actin. Cell Biol. Int. Rep. 13:1007–1021. [DOI] [PubMed] [Google Scholar]
- Benfenati, F., F. Valtorta, E. Chieregatti, and P. Greengard. 1992. Interaction of free and synaptic vesicle-bound synapsin I with F-actin. Neuron. 8:377–386. [DOI] [PubMed] [Google Scholar]
- Brodin, L., O. Shupliakov, V.A. Pieribone, J. Hellgren, and R.H. Hill. 1994. The reticulospinal glutamate synapse in lamprey: plasticity and presynaptic variability. J. Neurophysiol. 72:592–604. [DOI] [PubMed] [Google Scholar]
- Buckley, K., and R.B. Kelly. 1985. Identification of a transmembrane glycoprotein specific for secretory vesicles of neural and endocrine cells. J. Cell Biol. 100:1284–1294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chi, P., P. Greengard, and T.A. Ryan. 2001. Synapsin dispersion and reclustering during synaptic activity. Nat. Neurosci. 4:1187–1193. [DOI] [PubMed] [Google Scholar]
- Chi, P., P. Greengard, and T.A. Ryan. 2003. Synaptic vesicle mobilization is regulated by distinct synapsin I phosphorylation pathways at different frequencies. Neuron. 38:69–78. [DOI] [PubMed] [Google Scholar]
- Colicos, M.A., B.E. Collins, M.J. Sailor, and Y. Goda. 2001. Remodeling of synaptic actin induced by photoconductive stimulation. Cell. 107:605–616. [DOI] [PubMed] [Google Scholar]
- Cousin, M.A., T.C. Tan, and P.J. Robinson. 2001. Protein phosphorylation is required for endocytosis in nerve terminals: potential role for the dephosphins dynamin I and synaptojanin, but not AP180 or amphiphysin. J. Neurochem. 76:105–116. [DOI] [PubMed] [Google Scholar]
- Doussau, F., and G.J. Augustine. 2000. The actin cytoskeleton and neurotransmitter release: an overview. Biochimie. 82:353–363. [DOI] [PubMed] [Google Scholar]
- Esser, L., C.R. Wang, M. Hosaka, C.S. Smagula, T.C. Südhof, and J. Deisenhofer. 1998. Synapsin I is structurally similar to ATP-utilizing enzymes. EMBO J. 17:977–984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fesce, R., F. Benfenati, P. Greengard, and F. Valtorta. 1992. Effects of the neuronal phosphoprotein synapsin I on actin polymerization. II. Analytical interpretation of kinetic curves. J. Biol. Chem. 267:11289–11299. [PubMed] [Google Scholar]
- Foster-Barber, A., and J.M. Bishop. 1998. Src interacts with dynamin and synapsin in neuronal cells. Proc. Natl. Acad. Sci. USA. 95:4673–4677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gad, H., P. Löw, E. Zotova, L. Brodin, and O. Shupliakov. 1998. Dissociation between Ca2+-triggered synaptic vesicle exocytosis and clathrin-mediated endocytosis at a central synapse. Neuron. 21:607–616. [DOI] [PubMed] [Google Scholar]
- Gad, H., N. Ringstad, P. Low, O. Kjaerulff, J. Gustafsson, M. Wenk, G. Di Paolo, Y. Nemoto, J. Crun, M.H. Ellisman, et al. 2000. Fission and uncoating of synaptic clathrin-coated vesicles are perturbed by disruption of interactions with the SH3 domain of endophilin. Neuron. 27:301–312. [DOI] [PubMed] [Google Scholar]
- Greengard, P., F. Benfenati, and F. Valtorta. 1994. Synapsin I, an actin-binding protein regulating synaptic vesicle traffic in the nerve terminal. Molecular and Cellular Mechanisms of Neurotransmitter Release. L. Stjärne, P. Greengard, S. Grillner, T. Hökfelt, and D. Ottoson, editors. Raven Press, New York. 31–45. [DOI] [PubMed]
- Hilfiker, S., F.E. Schweizer, H.T. Kao, A.J. Czernik, P. Greengard, and G.J. Augustine. 1998. Two sites of action for synapsin domain E in regulating neurotransmitter release. Nat. Neurosci. 1:29–35. (published erratum appears in Nat. Neurosci. 1998. 1:329). [DOI] [PubMed] [Google Scholar]
- Hilfiker, S., V.A. Pieribone, A.J. Czernik, H.T. Kao, G.J. Augustine, and P. Greengard. 1999. Synapsins as regulators of neurotransmitter release. Philos. Trans. R. Soc. Lond. B Biol. Sci. 354:269–279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hosaka, M., and T.C. Südhof. 1999. Homo- and heterodimerization of synapsins. J. Biol. Chem. 274:16747–16753. [DOI] [PubMed] [Google Scholar]
- Huttner, W.B., W. Schiebler, P. Greengard, and P. De Camilli. 1983. Synapsin I (protein I), a nerve terminal–specific phosphoprotein. III. Its association with synaptic vesicles studied in a highly purified synaptic vesicle preparation. J. Cell Biol. 96:1374–1388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jovanovic, J., T.S. Sihra, A.C. Nairn, H.C. Hemmings, Jr., P. Greengard, and A.J. Czernik. 2001. Bi-directional changes in synapsin I phosphorylation during voltage-dependent Ca2+ entry and neurotransmitter release in isolated nerve terminals (synaptosomes). J. Neurosci. 21:7944–7953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jovanovic, J.N., F. Benfenati, Y.L. Siow, T.S. Sihra, J.S. Sanghera, S.L. Pelech, P. Greengard, and A.J. Czernik. 1996. Neurotrophins stimulate phosphorylation of synapsin I by MAP kinase and regulate synapsin I-actin interactions. Proc. Natl. Acad. Sci. USA. 93:3679–3683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jovanovic, J.N., A.J. Czernik, A.A. Fienberg, P. Greengard, and T.S. Sihra. 2000. Synapsins as mediators of BDNF-enhanced neurotransmitter release. Nat. Neurosci. 3:323–329. [DOI] [PubMed] [Google Scholar]
- Kao, H.T., B. Porton, S. Hilfiker, G. Stefani, V.A. Pieribone, R. DeSalle, and P. Greengard. 1999. Molecular evolution of the synapsin gene family. J. Exp. Zool. 285:360–377. [PubMed] [Google Scholar]
- Kessels, M.M., A.E. Engqvist-Goldstein, D.G. Drubin, and B. Qualmann. 2001. Mammalian Abp1, a signal-responsive F-actin-binding protein, links the actin cytoskeleton to endocytosis via the GTPase dynamin. J. Cell Biol. 153:351–366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, L., L.S. Chin, O. Shupliakov, L. Brodin, T.S. Sihra, O. Hvalby, V. Jensen, D. Zheng, J.O. McNamara, P. Greengard, et al. 1995. Impairment of synaptic vesicle clustering and of synaptic transmission, and increased seizure propensity, in synapsin I-deficient mice. Proc. Natl. Acad. Sci. USA. 92:9235–9239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Llinas, R., T.L. McGuinness, C.S. Leonard, M. Sugimori, and P. Greengard. 1985. Intraterminal injection of synapsin I or calcium/calmodulin-dependent protein kinase II alters neurotransmitter release at the squid giant synapse. Proc. Natl. Acad. Sci. USA. 82:3035–3039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Llinas, R., J.A. Gruner, M. Sugimori, T.L. McGuinness, and P. Greengard. 1991. Regulation by synapsin I and Ca(2+)-calmodulin-dependent protein kinase II of the transmitter release in squid giant synapse. J. Physiol. 436:257–282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maycox, P.R., E. Link, A. Reetz, S.A. Morris, and R. Jahn. 1992. Clathrin-coated vesicles in nervous tissue are involved primarily in synaptic vesicle recycling. J. Cell Biol. 118:1379–1388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McPherson, P.S., A.J. Czernik, T.J. Chilcote, F. Onofri, F. Benfenati, P. Greengard, J. Schlessinger, and P. De Camilli. 1994. Interaction of Grb2 via its Src homology 3 domains with synaptic proteins including synapsin I. Proc. Natl. Acad. Sci. USA. 91:6486–6490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Merrifield, C.J., S.E. Moss, C. Ballestrem, B.A. Imhof, G. Giese, I. Wunderlich, and W. Almers. 1999. Endocytic vesicles move at the tips of actin tails in cultured mast cells. Nat. Cell Biol. 1:72–74. [DOI] [PubMed] [Google Scholar]
- Morales, M., M.A. Colicos, and Y. Goda. 2000. Actin-dependent regulation of neurotransmitter release at central synapses. Neuron. 27:539–550. [DOI] [PubMed] [Google Scholar]
- Onofri, F., S. Giovedi, P. Vaccaro, A.J. Czernik, F. Valtorta, P. De Camilli, P. Greengard, and F. Benfenati. 1997. Synapsin I interacts with c-Src and stimulates its tyrosine kinase activity. Proc. Natl. Acad. Sci. USA. 94:12168–12173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Onofri, F., S. Giovedi, H.T. Kao, F. Valtorta, L.B. Borbone, P. De Camilli, P. Greengard, and F. Benfenati. 2000. Specificity of the binding of synapsin I to Src homology 3 domains. J. Biol. Chem. 275:29857–29867. [DOI] [PubMed] [Google Scholar]
- Phillips, G., J. Huang, Y. Wang, H. Tanaka, L. Shapiro, W. Zhang, W. Shan, K. Arndt, M. Frank, R. Gordon, et al. 2001. The presynaptic particle web: ultrastructure, composition, dissolution, and reconstitution. Neuron. 32:63–77. [DOI] [PubMed] [Google Scholar]
- Pieribone, V., O. Shupliakov, L. Brodin, S. Hilfiker-Rosenfluh, A. Czernik, and P. Greengard. 1995. Distinct pools of synaptic vesicles in neurotransmitter release. Nature. 375:493–497. [DOI] [PubMed] [Google Scholar]
- Pieribone, V., B. Porton, B. Rendon, J. Feng, P. Greengard, and H. Kao. 2002. Expression of synapsin III in nerve terminals and neurogenic regions of the adult brain. J. Comp. Neurol. 454:105–114. [DOI] [PubMed] [Google Scholar]
- Qualmann, B., J. Roos, P.J. DiGregorio, and R.B. Kelly. 1999. Syndapin I, a synaptic dynamin-binding protein that associates with the neural Wiskott-Aldrich syndrome protein. Mol. Biol. Cell. 10:501–513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ringstad, N., Y. Nemoto, and P. De Camilli. 1997. The SH3p4/Sh3p8/SH3p13 protein family: binding partners for synaptojanin and dynamin via a Grb2-like Src homology 3 domain. Proc. Natl. Acad. Sci. USA. 94:8569–8574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ringstad, N., H. Gad, P. Löw, G. Di Paolo, L. Brodin, O. Shupliakov, and P. De Camilli. 1999. Endophilin/SH3p4 is required for the transition from early to late stages in clathrin-mediated synaptic vesicle endocytosis. Neuron. 24:143–154. [DOI] [PubMed] [Google Scholar]
- Rosahl, T.W., M. Geppert, D. Spillane, J. Herz, R.E. Hammer, R.C. Malenka, and T.C. Südhof. 1993. Short-term synaptic plasticity is altered in mice lacking synapsin I. Cell. 75:661–670. [DOI] [PubMed] [Google Scholar]
- Rosahl, T.W., D. Spillane, M. Missler, J. Herz, D.K. Selig, J.R. Wolff, R.E. Hammer, R.C. Malenka, and T.C. Südhof. 1995. Essential functions of synapsins I and II in synaptic vesicle regulation. Nature. 375:488–493. [DOI] [PubMed] [Google Scholar]
- Rozelle, A.L., L.M. Machesky, M. Yamamoto, M.H. Driessens, R.H. Insall, M.G. Roth, K. Luby-Phelps, G. Marriott, A. Hall, and H.L. Yin. 2000. Phosphatidylinositol 4,5-bisphosphate induces actin-based movement of raft-enriched vesicles through WASP-Arp2/3. Curr. Biol. 10:311–320. [DOI] [PubMed] [Google Scholar]
- Ryan, T.A., L. Li, L.S. Chin, P. Greengard, and S.J. Smith. 1996. Synaptic vesicle recycling in synapsin I knock-out mice. J. Cell Biol. 134:1219–1227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sankaranarayanan, S., P. Atluri, and T. Ryan. 2003. Actin has a molecular scaffolding, not propulsive, role in presynaptic function. Nat. Neurosci. 6:127–135. [DOI] [PubMed] [Google Scholar]
- Schotland, J., O. Shupliakov, S. Grillner, and L. Brodin. 1996. Synaptic and non-synaptic monoaminergic neuronal systems in the lamprey spinal cord. J. Comp. Neurol. 372:229–244. [DOI] [PubMed] [Google Scholar]
- Shupliakov, O., and L. Brodin. 2000. A model glutamate synapse: the lamprey giant reticulospinal axon. Handbook of Chemical Neuroanatomy: Glutamate. O.P. Ottersen and J.S. Mathisen, editors. Elsevier Science Publishing Co. Inc., Amsterdam. 273–288.
- Shupliakov, O., P. Löw, D. Grabs, H. Gad, H. Chen, C. David, K. Takei, P. De Camilli, and L. Brodin. 1997. Synaptic vesicle endocytosis impaired by disruption of dynamin-SH3 domain interactions. Science. 276:259–263. [DOI] [PubMed] [Google Scholar]
- Shupliakov, O., O. Bloom, J. Gustafsson, P. Löw, P. Greengard, and L. Brodin. 1998. Two roles of actin in synaptic vesicle recycling. The 3rd meeting of the Swedish Association for Cytoskeletal Research: The Cytoskeleton and Cell Signalling; 1998 October 16–18; Kristineberg, Sweden. Stockholm University Press, Stockholm, Sweden. p. 10.
- Shupliakov, O., O. Bloom, J.S. Gustafsson, O. Kjaerulff, P. Löw, N. Tomilin, V.A. Pieribone, P. Greengard, and L. Brodin. 2002. Impaired recycling of synaptic vesicles after acute perturbation of the presynaptic actin cytoskeleton. Proc. Natl. Acad. Sci. USA. 99:14476–14481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Slepnev, V.I., and P. De Camilli. 2000. Accessory factors in clathrin-dependent synaptic vesicle endocytosis. Nat. Rev. Neurosci. 1:161–172. [DOI] [PubMed] [Google Scholar]
- Südhof, T.C., M. Baumert, M.S. Perin, and R. Jahn. 1989. A synaptic vesicle membrane protein is conserved from mammals to Drosophila. Neuron. 2:1475–1481. [DOI] [PubMed] [Google Scholar]
- Takei, Y., A. Harada, S. Takeda, K. Kobayashi, S. Terada, T. Noda, T. Takahashi, and N. Hirokawa. 1995. Synapsin I deficiency results in the structural change in the presynaptic terminals in the murine nervous system. J. Cell Biol. 131:1789–1800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Terada, S., T. Tsujimoto, Y. Takei, T. Takahashi, and N. Hirokawa. 1999. Impairment of inhibitory synaptic transmission in mice lacking synapsin I. J. Cell Biol. 145:1039–1048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torri-Tarelli, F., M. Bossi, R. Fesce, P. Greengard, and F. Valtorta. 1992. Synapsin I partially dissociates from synaptic vesicles during exocytosis induced by electrical stimulation. Neuron. 9:1143–1153. [DOI] [PubMed] [Google Scholar]
- Vaccaro, P., L. Dente, F. Onofri, A. Zucconi, S. Martinelli, F. Valtorta, P. Greengard, G. Cesareni, and F. Benfenati. 1997. Anti-synapsin monoclonal antibodies: epitope mapping and inhibitory effects on phosphorylation and Grb2 binding. Brain Res. Mol. Brain Res. 52:1–16. [DOI] [PubMed] [Google Scholar]
- Valtorta, F., P. Greengard, R. Fesce, E. Chieregatti, and F. Benfenati. 1992. Effects of the neuronal phosphoprotein synapsin I on actin polymerization. I. Evidence for a phosphorylation-dependent nucleating effect. J. Biol. Chem. 267:11281–11288. [PubMed] [Google Scholar]
- Wickelgren, W., J. Leonard, M. Grimes, and R. Clark. 1985. Ultrastructural correlates of transmitter release in presynaptic areas of lamprey reticulospinal axons. J. Neurosci. 5:1188–1201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Witke, W., A.V. Podtelejnikov, A. Di Nardo, J.D. Sutherland, C.B. Gurniak, C. Dotti, and M. Mann. 1998. In mouse brain profilin I and profilin II associate with regulators of the endocytic pathway and actin assembly. EMBO J. 17:967–976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao, W., S. Cavallaro, P. Gusev, and D.L. Alkon. 2000. Nonreceptor tyrosine protein kinase pp60c-src in spatial learning: synapse-specific changes in its gene expression, tyrosine phosphorylation, and protein-protein interactions. Proc. Natl. Acad. Sci. USA. 97:8098–8103. [DOI] [PMC free article] [PubMed] [Google Scholar]