

Abstract
We have discovered a new and specific cell-killing mechanism mediated by the selective uptake of the antitumor drug 1-O-octadecyl-2-O-methyl-rac-glycero-3-phosphocholine (ET-18-OCH3, Edelfosine) into lipid rafts of tumor cells, followed by its coaggregation with Fas death receptor (also known as APO-1 or CD95) and recruitment of apoptotic molecules into Fas-enriched rafts. Drug sensitivity was dependent on drug uptake and Fas expression, regardless of the presence of other major death receptors, such as tumor necrosis factor (TNF) receptor 1 or TNF-related apoptosis-inducing ligand R2/DR5 in the target cell. Drug microinjection experiments in Fas-deficient and Fas-transfected cells unable to incorporate exogenous ET-18-OCH3 demonstrated that Fas was intracellularly activated. Partial deletion of the Fas intracellular domain prevented apoptosis. Unlike normal lymphocytes, leukemic T cells incorporated ET-18-OCH3 into rafts coaggregating with Fas and underwent apoptosis. Fas-associated death domain protein, procaspase-8, procaspase-10, c-Jun amino-terminal kinase, and Bid were recruited into rafts, linking Fas and mitochondrial signaling routes. Clustering of rafts was necessary but not sufficient for ET-18-OCH3–mediated cell death, with Fas being required as the apoptosis trigger. ET-18-OCH3–mediated apoptosis did not require sphingomyelinase activation. Normal cells, including human and rat hepatocytes, did not incorporate ET-18-OCH3 and were spared. This mechanism represents the first selective activation of Fas in tumor cells. Our data set a framework for the development of more targeted therapies leading to intracellular Fas activation and recruitment of downstream signaling molecules into Fas-enriched rafts.
Keywords: CD95, Fas signaling, death receptor, antitumor ether lipid, ET-18-OCH3
Introduction
A group of synthetic ether-linked lysophospholipid analogs, collectively named antitumor ether lipids, represent a promising class of anticancer agents that, unlike most conventional chemotherapeutic drugs, do not target the DNA but act at the level of cell membranes, and therefore their effects are independent of the proliferative state of target cells (1–4). The synthetic ether phospholipid 1-O-octadecyl-2-O-methyl-rac-glycero-3-phosphocholine (ET-18-OCH3, Edelfosine) selectively induces apoptosis in cancer cells (5) and has become the effective prototype of antitumor ether lipids. Because of its ether bonds, ET-18-OCH3 is metabolically stable, resistant to phospholipases, and accumulates in the plasma membrane (6). The mechanism by which ET-18-OCH3 selectively engages the suicide apparatus in tumor cells is still unclear. Persistent activation of the c-Jun amino-terminal kinase (JNK) pathway (7, 8) and mitochondrial-related processes (9–11) have been involved in the apoptotic response induced by ET-18-OCH3. We have recently found that ET-18-OCH3–induced apoptosis is mediated by the activation of the death receptor Fas (also known as APO-1 or CD95) independently of its ligand (FasL, also named CD95L; references 12 and 13) through its coclustering with membrane rafts (14). This Fas association with membrane rafts, further confirmed in subsequent studies following Fas–FasL interaction (15–17), can be of major importance in amplifying Fas signaling as lipid rafts might serve as foci for recruitment of signaling molecules at the plasma membrane (18). Fas is a member of the TNFR family that triggers cell death through the presence of a death domain in its cytoplasmic portion after receptor engagement with FasL or agonistic anti-Fas antibodies (19). Stimulation of Fas results in receptor oligomerization and recruitment of the adaptor molecule Fas-associated death domain protein (FADD) through interaction between its own death domain and the clustered receptor death domains. In turn, FADD recruits procaspase-8 via a death effector domain interaction forming the so-called death-inducing signaling complex (DISC; reference 20). The close proximity of procaspase-8 molecules in the DISC drives its activation by self-cleavage, triggering downstream effector caspases and leading to apoptosis (21). Thus, formation of membrane platforms where Fas molecules are brought together is suggested to increase DISC formation and therefore potentiate Fas signaling (13, 14, 16, 17, 22–25). In this regard, sphingomyelinase (SMase)-derived ceramide has been recently reported to amplify the signaling of Fas at the membrane level after the initial Fas–FasL interaction, acting as a mediator of the clustering of Fas (23, 24, 26).
Here, we show a novel mechanism of drug action in which ET-18-OCH3 is selectively incorporated into tumor cell membrane rafts promoting their clustering as well as translocation of Fas and downstream signaling molecules into clustered rafts in a SMase-independent manner. ET-18-OCH3–mediated apoptosis was dependent on intracellular activation of Fas, independently of other major death receptors such as TNF and TNF-related apoptosis-inducing ligand (TRAIL) receptors. Normal cells, including hepatocytes and PBLs, were spared because they failed to take up the ether lipid and thereby failed to trigger the ensuing apoptotic events.
Materials and Methods
Cells and Fas Transfectants.
The human cell lines HL-60 (acute myeloid leukemia), U937 (histiocytic lymphoma), and Jurkat (acute T cell leukemia), obtained from the American Type Culture Collection, were grown as described previously in RPMI 1640 culture medium supplemented with 10% heat-inactivated FCS, 2 mM l-glutamine, 100 U/ml penicillin, and 24 μg/ml gentamicin (5).
To isolate PBLs, mononuclear cells were obtained from fresh human peripheral blood by centrifugation on Ficoll-Paque density gradients, washed with PBS, and resuspended in RPMI 1640 containing 10% heat-inactivated FCS. Monocytes were depleted by culture dish adherence after overnight incubation. Nonadherent cells (lymphocytes) were washed with PBS and collected. Lymphocyte preparations were typically 69–74% CD3+, 25–28% CD19+, and <0.4% CD14+ (12).
The human fibrosarcoma HT1080 cell line, obtained from the German Collection of Microorganisms and Cell Cultures, as well as the murine fibroblasts L929 and L929-Fas, stably transfected with human Fas cDNA (provided by P.H. Krammer, DKFZ, Heidelberg, Germany; reference 27), were grown in DMEM culture medium supplemented with 10% heat-inactivated FCS, 2 mM l-glutamine, 100 U/ml penicillin, and 24 μg/ml gentamicin (12). L929-Fas cells were grown in the presence of 500 μg/ml hygromycin. A PCR-generated DNA fragment encoding a 57C terminally truncated Fas version (FasΔ57C) was prepared using the plasmid pKEX-Fas (28) as a template, cloned into the eukaryotic expression vector pKEX-2-XR, and the sequence was verified by double-strand DNA sequencing. Transfection of L929 cells with pKEX-FasΔ57C plasmid was performed using Lipofectin reagent (Life Technologies) and hygromycin for clone selection. As a control, transfection was also performed with empty pKEX-2-XR plasmid.
Human and Rat Hepatocytes.
Human hepatocytes were obtained from small surgical liver biopsies by a two-step collagenase perfusion method (29). Rat hepatocytes were prepared from 20-d-old fetal rat livers by a nonperfusion collagenase dispersion method (30).
Immunofluorescence Flow Cytometry.
Cell surface expression of death receptors was analyzed by immunofluorescence flow cytometry in 4 × 105 cells as described previously (12, 31) in a Becton Dickinson FACSCalibur™ flow cytometer using anti-Fas SM1/1 mAb (Bender MedSystems) and specific antibodies against TNFR1 and TRAIL-R2/DR5 (Qbiogene, Oncogene Research Products, and Santa Cruz Biotechnology, Inc.). P3X63 myeloma culture supernatant, provided by F. Sanchez-Madrid (Hospital de La Princesa, Madrid, Spain), was used as a negative control.
Apoptosis Assay.
Quantitation of apoptotic cells was determined by flow cytometry as the percentage of cells in the sub-G1 region (hypodiploidy) in cell cycle analysis as described previously (12). Death receptor–mediated apoptosis was induced with rhTRAIL or rhFasL (Qbiogene), active in human and murine systems, murine TNF-α (provided by M. Modolell, Max-Planck-Institut für Immunbiologie, Freiburg, Germany), rhTNF-α (Qbiogene), and cytotoxic anti-Fas CH-11 mAb (Upstate Biotechnology; reference 12).
ET-18-OCH3 Uptake.
Drug uptake was measured as described previously (5) after incubating 106/ml cells with 10 μM ET-18-OCH3 plus 0.05 μCi/ml [3H]ET-18-OCH3 for 6 h, and subsequent exhaustive washing (five times) with 1% BSA-PBS. ET-18-OCH3 stock solutions (INKEYSA) were prepared as described previously (5). [3H]ET-18-OCH3 (specific activity, 42 Ci/mmol) was synthesized by tritiation of the 9-octadecenyl derivative (Amersham Buchler).
Microinjection Assays and Detection of DNA Fragmentation by TdT-mediated dUTP Nick-End Labeling (TUNEL).
Cells were grown on 24-mm diameter poly-l-lysine–coated glass coverslips and microinjected with glass microelectrodes (150–200 MΩ) filled with 1.2% rhodamine-conjugated dextran (Molecular Probes) in 1 M KCl as a marker or with rhodamine-conjugated dextran plus 190 μM ET-18-OCH3, which renders final intracellular drug concentrations comparable to the normally used doses (12). Cells were microinjected using a hydraulic 3D micromanipulator (Narishige) under visual control (40-kPa for 0.1–1 s; Diaphot microscope; Nikon; reference 12). Cells were then incubated in 10% FCS-DMEM for 6 h, fixed with 4% formaldehyde, permeabilized with 0.2% Triton X-100, and analyzed for DNA fragmentation in situ by the TUNEL technique using the Fluorescein Apoptosis Detection System kit (Promega) according to the manufacturer's instructions. Fluorescence of rhodamine and fluorescein was examined with a laser scan confocal microscope (LSM 510; Carl Zeiss MicroImaging, Inc.). Rhodamine stains red throughout the cytoplasm of all the microinjected cells, whereas fluorescein-12-dUTP is incorporated at the 3′-OH ends of fragmented DNA, resulting in localized green fluorescence within the nucleus of apoptotic cells.
Confocal Microscopy.
Untreated and drug-treated cells were settled onto poly-l-lysine–coated slides and analyzed with a Zeiss LSM 510 laser scan confocal microscope for raft and protein visualization as described previously (14). Colocalization assays were analyzed by excitation of the corresponding fluorochromes in the same section of permeabilized or nonpermeabilized samples. Negative controls, lacking the primary antibody or using an irrelevant antibody, showed no staining.
Lipid Raft Isolation and Western Blotting.
Lipid rafts were isolated from 7 × 107 cells by nonionic detergent lysis and centrifugation on discontinuous sucrose gradients as described previously (14, 32). 1-ml fractions were collected from the top of the gradient and 20 μl of each fraction was subjected to SDS-PAGE, immunoblotting, and enhanced chemiluminescence detection. Location of GM1-containing lipid rafts was determined using cholera toxin (CTx) B subunit conjugated to horseradish peroxidase (Sigma-Aldrich) as described previously (32). Proteins were identified using the following specific antibodies: anti–48-kD Fas (C-20) and anti–46-kD JNK1 (C-17) rabbit polyclonal antibodies, and anti–23-kD Bid (N-19) and anti–45-kD DR5 (N-19) goat polyclonal antibodies (Santa Cruz Biotechnology, Inc.); anti–29-kD FADD mAb (clone-1; BD Transduction Laboratories); anti–60-kD TNFR1 (H398) mAb (Qbiogene); and anti–45-kD DR5 (Ab-1) rabbit polyclonal antibody, anti–55-kD procaspase-8 (Ab-3), and anti–58-kD procaspase-10 (Ab-2) mAb (Oncogene Research Products).
Sphingomyelin and Ceramide Measurement.
Cellular lipid pools were metabolically radiolabeled by culturing cells for 3 d in the presence of 0.5 μCi/ml [methyl-3H]choline or L-[14C]serine, and the radiolabeled pools of sphingomyelin and ceramide were analyzed by thin layer chromatography (TLC) and scintillation counting after the distinct cellular treatments (33). [3H]choline-labeled extracts were separated by TLC using chloroform/methanol/25% NH4OH (60:30:5) as the solvent system, allowing separation of sphingomyelin. [3H]serine-labeled extracts were subjected to an alkaline hydrolysis procedure. Lipids were dissolved into chloroform/methanol (1:1) containing 0.1 M NaOH. After an incubation of 1 h at 37°C, the remaining lipids were reextracted and subdivided on two TLC plates, which were developed in chloroform/acetic acid (9:1), separating ceramide, or chloroform/methanol/25% NH4OH (60:30:5), separating sphingomyelin. Radiolabeled lipids were identified by autoradiography as well as by the use of unlabeled standards, scraped from the plates, and counted for radioactivity.
Ceramide was also quantified by the diacylglycerol kinase assay as described previously (33) using 5 × 105 cells and 300 μCi/ml [γ-32P]ATP in each experimental point. Radiolabeled lipids were visualized by TLC and autoradiography, and the ceramide-1-phosphate spots were scraped from the plates and counted for radioactivity.
SMase Activities.
Neutral and acidic SMase was assayed in the absence of serum using [choline-methyl-14C]-sphingomyelin as described previously (34). In brief, volumes were adjusted to 125 μl (105 Jurkat cells) and reactions were started by adding 125 μl of substrate solution. For the measurement of acidic SMase activity, the substrate solution consisted of [methyl-choline-14C]-sphingomyelin (105 dpm/assay) and 0.1% (wt/vol) Triton X-100 in 250 mM sodium acetate buffer, pH 5.0, containing 10 mM EDTA. For neutral SMase activity assays, the substrate solution consisted of [methyl-choline-14C]-sphingomyelin (105 dpm/assay) and 0.1% (wt/vol) Triton X-100 in 250 mM Tris HCl buffer, pH 7.4, containing 10 mM dithiothreitol and 10 mM MgCl2. After 2 h of incubation at 37°C, reactions were terminated by extracting the lipids by subsequent addition of methanol/chloroform/H2O (100:100:50), and the sphingomyelin degradation product [14C]phosphorylcholine was quantified in the aqueous upper phase by liquid scintillation counting.
PTE-ET-18-OCH3 Synthesis and Uptake.
The fluorescent analogue PTE-ET-18-OCH3 was synthesized through cross-coupling between 1-bromopolyene and 1,3-glycerol acetal with a terminal acetylene group, followed by partial reduction (unpublished data). Cells (2.5 × 105 cells/ml) were treated with 20 μM PTE-ET-18-OCH3 for 7 h in 10% FCS-RPMI 1640, washed three times with 1% BSA-PBS, and the incorporated fluorescence was measured with a microplate fluorometer (TECAN). 2 × 105 PTE-ET-18-OCH3–treated cells were settled onto poly-l-lysine–coated slides, fixed in 4% formaldehyde for 25 min at room temperature, and analyzed using a fluorescence microscope (Axioplan 2; Carl Zeiss MicroImaging, Inc.) and a digital camera (ORCA-ER-1394; Hamamatsu).
Results
ET-18-OCH3–induced Apoptosis Involves Drug Uptake and Intracellular Activation of Fas, but Is Independent of TNF and TRAIL Functional Receptors.
Our previous reports indicated that Fas activation was involved in ET-18-OCH3–mediated apoptosis (12, 14). To gain further insight and to discern whether additional death receptors could be involved in ET-18-OCH3–induced apoptosis, we used mouse fibrosarcoma Fas-deficient L929 cells and generated stable transfectants with the pKEX-2-XR expression vector containing human full Fas (L929-Fas) or a truncated version of human Fas (L929-FasΔ57C), lacking the 57 COOH-terminal amino acids (amino acids 279–335) that included part of the Fas cytoplasmic death domain (35). L929-Fas and L929-FasΔ57C cells expressed high levels of human Fas protein on their cell surface (Fig. 1 A). L929, L929-Fas, and L929-FasΔ57C were sensitive to TNF-α and TRAIL (Fig. 1 B), indicating the functional presence of their corresponding receptors. rhFasL or the agonistic cytotoxic CH-11 anti-Fas mAb induced apoptosis in L929-Fas cells, but not in L929-FasΔ57C cells (Fig. 1 B). Parental and transfected L929 cells hardly incorporated ET-18-OCH3 (<0.2 nmol/106 cells after 6 h of incubation with 10 nmol [3H]ET-18-OCH3) and were resistant to ET-18-OCH3 incubation (<1% apoptosis after a 2-d treatment with 10 μM ET-18-OCH3), indicating that Fas has no role in the incorporation of the ether lipid. Microinjection of ET-18-OCH3 into L929-Fas cells induced a potent apoptotic response, but not in L929 or L929-FasΔ57C cells (Fig. 1 C). L929 cells transfected with empty pKEX-2-XR expression vector behaved as parental L929 cells (not depicted). These results demonstrate that ET-18-OCH3 must be inside the cell to promote Fas-mediated apoptosis and suggest that the proapoptotic action of the ether lipid is mediated by the intracellular domain of Fas. These data also demonstrate that the presence of Fas in the target cell, independently of other TNF-α– or TRAIL-activated death receptors, is essential for ET-18-OCH3–mediated apoptosis.
Figure 1.
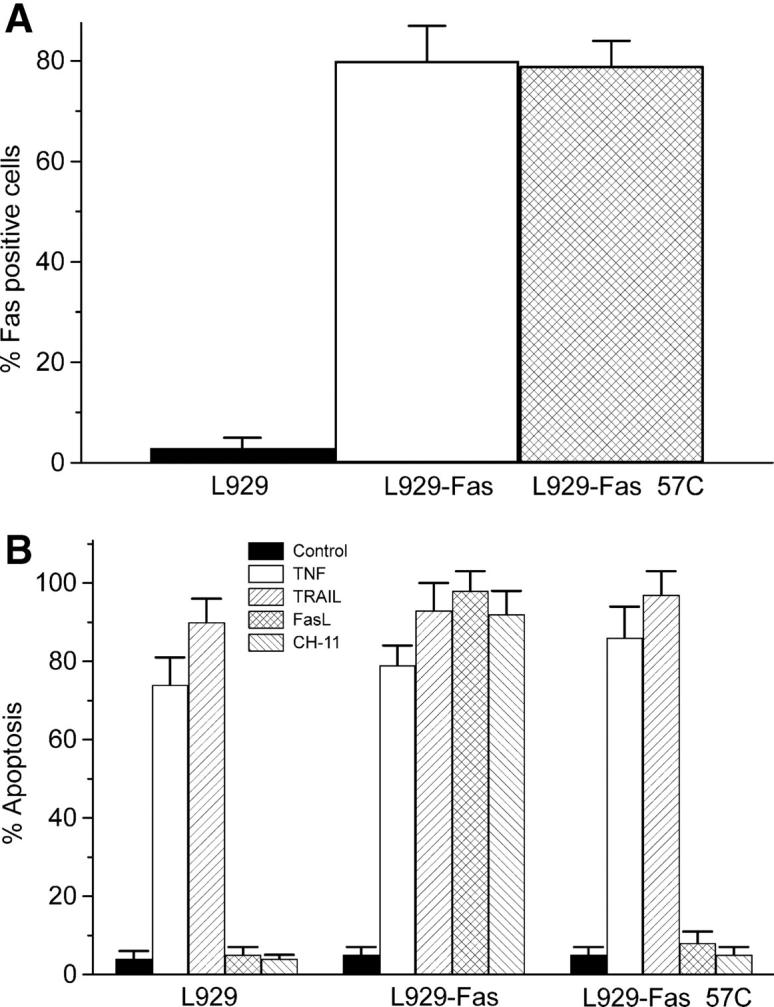

Intracellular Fas activation in ET-18-OCH3–mediated apoptosis. L929, L929-Fas, and L929-FasΔ57C cells were assayed by flow cytometry for their respective cell surface content of Fas using anti-Fas SM1/1 mAb (A), and for the ability of 100 U/ml murine TNF-α (TNF), 50 ng/ml rhTRAIL, 100 ng/ml rhFasL, and 50 ng/ml cytotoxic anti-Fas CH-11 mAb to induce apoptosis after a 24-h incubation in the presence of 500 ng/ml actinomycin D (B). Untreated control cells (Control) were run in parallel. Data shown are means ± SD of three independent determinations. (C) Apoptosis was determined by TUNEL analysis in L929, L929-Fas, and L929-FasΔ57C cells after microinjection of rhodamine-labeled dextran alone (Control), used to visualize the microinjected cells by red fluorescence, or in combination with ET-18-OCH3 (ET-18-OCH3). Microinjected cells are identified by cytoplasmic red staining, and apoptotic nuclei are visualized by the TUNEL reaction in green. Images shown are representative of four independent experiments. Bar, 10 μm.
ET-18-OCH3 Induces Recruitment and Clustering of Fas and Downstream Signaling Molecules into Membrane Rafts.
Human T lymphoid leukemic Jurkat cells take up high amounts of ET-18-OCH3 (3.1 ± 0.4 nmol/106 cells after 6 h of incubation with 10 nmol [3H]ET-18-OCH3; n = 3), resulting in Fas translocation and clustering into membrane rafts followed by apoptosis (12, 14). Jurkat cells express a number of functional death receptors, including Fas, TNFR1, and TRAIL-R2/DR5 (12, 36, 37), but not TRAIL-R1/DR4 (38). We asked whether the intracellular-driven engagement and translocation of Fas into membrane rafts by ET-18-OCH3 was specific for Fas or could involve additional death receptors. We found that ET-18-OCH3 induced aggregation of Fas, but not of TNFR1 or DR5, into “caps” on one pole of the cell as evidenced by confocal microscopy (Fig. 2 and not depicted). These Fas clusters colocalized with membrane rafts (Fig. 2) as assessed by the raft marker FITC-CTx B subunit that binds ganglioside GM1 (39), mainly found in rafts (40). ET-18-OCH3–induced clustering of lipid rafts was detectable after only 15–30 min of 10 μM ET-18-OCH3 incubation, whereas recruitment of Fas into the raft clusters was observed after a 6-h treatment, preceding apoptosis that was detectable after a 9-h treatment. Translocation of Fas into rafts was further confirmed by isolation of membrane rafts in both untreated and ET-18-OCH3–treated Jurkat cells in sucrose gradients (Fig. 3). Lipid rafts were isolated based on their insolubility in Triton X-100 detergent and buoyant density on sucrose density gradients (41). Thus, Jurkat cells were lysed in a 1% Triton X-100 lysis buffer at 4°C and fractionated by discontinuous sucrose gradient centrifugation (14, 32). The distinct fractions from the gradient were analyzed by SDS-PAGE and Western blotting. The position of the membrane rafts in the sucrose gradient was determined by the presence of the ganglioside GM1, detected using the GM1-specific ligand CTx B subunit (Fig. 3). GM1 was enriched in the upper part of the sucrose gradient (fractions 3–6), with a secondary localization at the bottom of the gradient (fractions 9–12), indicating a separation of lipid rafts (fractions 3–6) from the Triton X-100-soluble membranes (Fig. 3). Using a specific anti-Fas mAb, we found that Fas was located in the soluble fractions (fractions 9–12) of the sucrose gradient and not in the detergent-insoluble lipid raft region in untreated Jurkat cells, indicating that Fas is excluded from the lipid rafts in untreated Jurkat cells (Fig. 3). However, ET-18-OCH3 treatment induced the recruitment of Fas into the lipid raft region (fractions 4–6) of the sucrose gradient (Fig. 3). These data showed a rather remarkable specificity for Fas in the action of ET-18-OCH3, as two additional major death receptors, namely TNFR1 and DR5, were not mobilized to rafts. We also found that FADD and procaspase-8, major components of the DISC (42), were recruited to the Fas-containing clustered rafts after ET-18-OCH3 treatment (Fig. 3). This colocalization was further confirmed by confocal microscopy (not depicted). Procaspase-10 was also recruited into rafts to a higher level than procaspase-8 (Fig. 3). However, the active caspase-10 and caspase-8 cleavage forms localized mostly in nonraft fractions (Fig. 3). Surprisingly, we found that JNK and Bid were also recruited into the lipid raft region upon ET-18-OCH3 treatment (Fig. 3). This is of interest as JNK and mitochondrial signaling have been involved in ET-18-OCH3–mediated apoptosis (7–11), and Bid has been shown to act as a bridge between Fas signaling and mitochodria (43, 44). Disruption of rafts after a 30-min pretreatment with 2.5 mg/ml methyl-β-cyclodextrin prevented ET-18-OCH3–induced apoptosis (51.5 ± 5.2 and 6.8 ± 0.9% apoptosis after a 24-h incubation with 10 μM ET-18-OCH3 in untreated or methyl-β-cyclodextrin–treated Jurkat cells, respectively; n = 3), Fas aggregation, and the translocation of the apoptotic signaling molecules (not depicted). Thus, ET-18-OCH3–induced clustering of Fas-containing rafts recruits and concentrates to the cytoplasmic side of the plasma membrane a number of molecules required to mount an efficient Fas-dependent apoptotic response, forming clusters of rafts enriched in apoptotic signaling molecules.
Figure 2.

ET-18-OCH3–induced clustering of Fas, but not of TNFR1, in human leukemic cells. Jurkat cells were either untreated (Control) or treated with 10 μM ET-18-OCH3 for 6 h, and then stained with FITC-CTx B subunit to identify rafts (green fluorescence) and with specific antibodies followed by CY3-conjugated antibodies (red fluorescence) to identify Fas (A) or TNFR1 (B). Areas of colocalization between membrane rafts and the indicated proteins in the merge panels are yellow. Images shown are representative of three independent experiments. Bar, 10 μm.
Figure 3.

Recruitment of Fas and downstream signaling molecules into membrane rafts after ET-18-OCH3 treatment. Untreated Jurkat cells (Control) and Jurkat cells treated with 10 μM ET-18-OCH3 (ET-18-OCH3) for 6 h were lysed in 1% Triton X-100 and subjected to discontinuous sucrose density gradient centrifugation. Individual fractions were subjected to SDS-PAGE and Western blotting. Location of GM1-containing rafts (fractions 3–6) was determined using CTx B subunit conjugated to horseradish peroxidase. Location of the indicated receptors and signaling proteins were examined using specific antibodies. Active p18 caspase-8 and p23 caspase-10 cleavage forms are indicated. Representative blots of three separate experiments are shown.
Although Fas and the components of the DISC have been reported to be recruited to lipid rafts after Fas ligation in mouse thymocytes and human T cell lines (16, 17), a recent report has shown that FasL induces cell death in human fibrosarcoma HT1080 cells without translocation of Fas to lipid rafts (45). This suggests that translocation of Fas to lipid rafts is cell type dependent. Next, we investigated whether ET-18-OCH3 could induce apoptosis as well as Fas translocation to lipid rafts in HT1080 cells that were reluctant to mobilize Fas to rafts. We found that HT1080 cells were resistant to the apoptotic action of ET-18-OCH3 (<3% apoptosis after a 2-d treatment with 10 μM ET-18-OCH3), and no raft or Fas clustering was observed in ET-18-OCH3–treated HT1080 cells (not depicted). On the other hand, treatment of HT1080 with the agonistic CH-11 mAb induced apoptosis (39.4 ± 4.8% apoptosis after a 48-h incubation with 50 ng/ml CH-11; n = 3), but no raft or Fas clustering was detected (not depicted), in agreement with previous observations (45). Thus, these data further support the notion that ET-18-OCH3–induced apoptosis is mediated by Fas translocation to lipid rafts.
ET-18-OCH3–induced Fas and Raft Clustering Do Not Require Protein Synthesis.
Next, we analyzed whether ET-18-OCH3 could induce a marked up-regulation of Fas on the cell surface, resulting in a high density of Fas and subsequent spontaneous clustering of the receptor. However, ET-18-OCH3 did not induce any increase on the Fas cell surface expression (Fig. 4 A), and cycloheximide pretreatment, at protein synthesis inhibitory concentrations (not depicted), did not affect raft and Fas coclustering after ether lipid incubation (Fig. 4 B). These results demonstrate that ET-18-OCH3–induced raft and Fas clustering do not depend on protein synthesis, and thereby ET-18-OCH3 induces translocation and clustering of Fas in lipid rafts from the pool of Fas already present in the tumor cell. This is in agreement with our previous observations that ET-18-OCH3–induced apoptosis does not depend on protein synthesis (10), and that ET-18-OCH3 did not induce any change in the expression of fas mRNA level (12).
Figure 4.


Clustering of Fas is independent of protein synthesis. (A) Cell surface expression of Fas was determined by immunofluorescence flow cytometry in untreated Jurkat cells (Control) and cells treated with 10 μM ET-18-OCH3 for 6 h. Data shown are means ± SD of three independent experiments. (B) Jurkat cells, preincubated with 5 μM cycloheximide (CHX) for 1 h and then treated with 10 μM ET-18-OCH3 for 6 h, were stained with FITC-CTx B subunit (green fluorescence for rafts) and anti-Fas mAb, followed by CY3-conjugated anti–mouse antibody (red fluorescence for Fas). Areas of colocalization between membrane rafts and Fas in the merge panels are yellow. Images shown are representative of three independent experiments. Bar, 10 μm.
Raft Clustering in the Absence of Fas Does Not Induce Apoptosis in ET-18-OCH3–treated Cells.
Because rafts were necessary for both Fas cell surface aggregation and ET-18-OCH3–induced apoptosis, and clustering of rafts was rapidly induced after ET-18-OCH3 addition, we asked whether raft clustering was sufficient to trigger apoptosis. We generated a subline from the leukemic Jurkat cell line that lacked Fas cell surface expression after protracted cultures (>10 mo) of parental cells. Fas-deficient Jurkat cells preserved the expression of TNFR1 and DR5 at similar levels as the parental Jurkat cells, but lacked Fas (Fig. 5 A), resulting in CH-11–, rhFasL-, and ET-18-OCH3–resistant cells (Fig. 5 B). However, Fas-deficient Jurkat cells underwent apoptosis after TNF-α or TRAIL incubation (not depicted). Interestingly, ET-18-OCH3 promoted clustering of rafts in Fas-deficient Jurkat cells in a similar way as in the parental Jurkat cells (Fig. 5 C), indicating that clustering of rafts was not sufficient to trigger an apoptotic response. These data further indicate that Fas is a major mediator in ET-18-OCH3 proapoptotic action and that the abilities of the ether lipid in promoting raft clustering and apoptosis can be dissociated.
Figure 5.


Clustering of Fas-deficient rafts is not sufficient for apoptosis. (A) Cell surface expression of Fas, TNFR1, and DR5 in Jurkat (JK) and Fas-deficient Jurkat cells was determined by flow cytometry. Percent of positive cells for each death receptor was estimated using the P3X63 (X63) myeloma culture supernatant as negative control. (B) Induction of apoptosis in Jurkat (JK) and Fas-deficient Jurkat cells was determined by flow cytometry after a 24-h incubation with 50 ng/ml cytotoxic anti-Fas CH-11 mAb, 100 ng/ml rhFasL (FasL), or 10 μM ET-18-OCH3 (ET). Untreated control cells (C) were run in parallel. Data shown are means ± SD of three independent experiments. (C) Fas-deficient Jurkat cells were untreated (Control) or incubated with 10 μM ET-18-OCH3 (ET-18-OCH3) for 6 h, and then stained with FITC-CTx B subunit and analyzed by confocal microscopy to visualize membrane rafts. Images shown are representative of three independent experiments. Bar, 10 μm.
ET-18-OCH3 Does Not Activate SMase.
Because SMase-released ceramide has been involved in Fas aggregation and killing (15, 24), we analyzed its putative involvement in ET-18-OCH3–induced apoptosis. Serine is a metabolic precursor for the biosynthesis of all sphingolipids, including ceramide and sphingomyelin. Choline forms part of the headgroup of both sphingomyelin and phosphatidylcholine. By growing cells in the presence of [3H]choline or [14C]serine, all mentioned lipids become radiolabeled. This allows sensitive detection of any change in the cellular levels of these lipids. As shown in Fig. 6 A, [3H]choline-labeled sphingomyelin pools were not affected in ET-18-OCH3–treated Jurkat cells. ET-18-OCH3 did not affect [3H]serine-labeled sphingomyelin or ceramide cell levels (Fig. 6, B and C). In addition, ET-18-OCH3 did not induce significant elevations in endogenous ceramide levels determined through the diacylglycerol kinase assay that allows mass measurement of ceramide (Fig. 6 D). Furthermore, we did not find any significant change in neutral and acidic SMase activities after ET-18-OCH3 treatment using [14C]sphingomyelin as substrate in time course experiments that included 1-, 5-, 15-, 30-, and 60-min time points (not depicted). Moreover, ET-18-OCH3 failed to activate SMase in human leukemic HL-60 and U937 cells (not depicted), which undergo ET-18-OCH3–mediated apoptosis (10, 14). These data suggest that SMase activation is not required in ET-18-OCH3–induced apoptosis.
Figure 6.
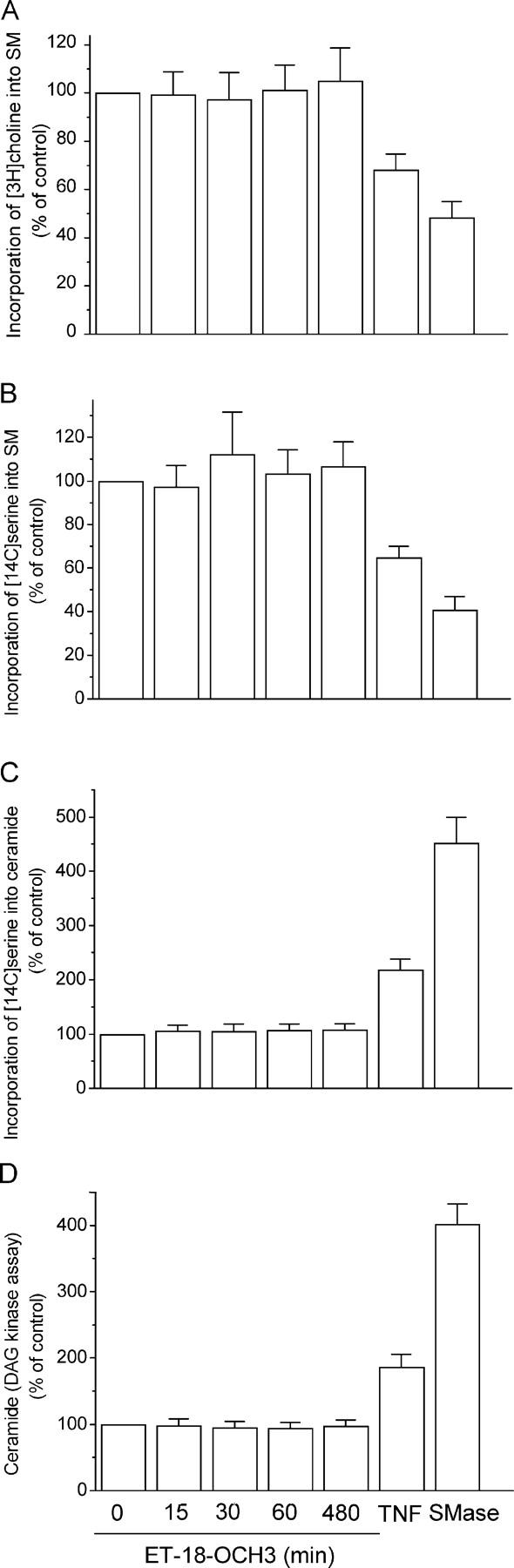
Lack of SMase activation and ceramide generation by ET-18-OCH3. Jurkat cells were labeled with [methyl-3H]choline and L-[14C]serine, and then treated with 10 μM ET-18-OCH3 for the indicated time points. The incorporation of [methyl-3H]choline and L-[14C]serine into sphingomyelin (SM) (A and B) and the incorporation of L-[14C]serine into ceramide (C) was determined as a percentage to untreated control cells. Ceramide content was also calculated by the diacylglycerol (DAG) kinase assay (D). Jurkat cells treated with 500 U/ml TNF-α (TNF) for 30 min or with 0.3 U/ml Staphylococcus aureus SMase for 1 h were used as positive controls. Data shown are means ± SD of three independent determinations.
ET-18-OCH3 Accumulates in Membrane Rafts and Colocalizes with Fas Clusters.
Next, we asked whether ET-18-OCH3 itself could colocalize with Fas in membrane rafts. We synthesized a novel ET-18-OCH3 fluorescent analogue able to preserve the selectivity for cancer cells of the parental drug. Because the sn-1 position of the ET-18-OCH3 molecule can hold some modifications preserving drug bioactivity (5), we synthesized an ET-18-OCH3 analogue replacing the sn-1 C18 aliphatic chain of ET-18-OCH3 by a lipophilic fluorescent group of similar length containing a conjugated ϖ-phenyltetraene group (all-[E]-1-O-[15′-phenylpentadeca-8′,10′,12′,14′-tetraenyl]-2-O-methyl-rac-glycero-3-phosphocholine, PTE-ET-18-OCH3, mol wt, 550.33) that shows maximum absorption bands at 334, 350, and 368 nm, and emission at 442 and 467 nm. This fluorescent analogue induced apoptosis in T lymphoid leukemia Jurkat cells (Fig. 7 A), whereas it spared normal human PBLs in a similar way to ET-18-OCH3 (<2% apoptosis in 10–30 μM ET-18-OCH3– or PTE-ET-18-OCH3–treated resting PBLs for 24 h). Incubation of human leukemic HL-60 and Jurkat cells with this fluorescent analogue, followed by exhaustive washing with a BSA-containing buffer to eliminate the loosely cell surface–bound ether lipid, led to an accumulation of the fluorescent analogue into well-defined dense patches, whereas a faint labeling was hardly detected in normal PBLs (Fig. 7 B). Incorporation of the fluorescent ET-18-OCH3 analogue was blocked by adding the parental drug ET-18-OCH3 (not depicted). The selective uptake of the ether lipid in tumor cells was further corroborated by quantitative fluorescence spectroscopy measurements (Fig. 7 C). Thus, the new fluorescent compound behaves as a reliable active fluorescent analogue of ET-18-OCH3 that allows us to visualize the location of ET-18-OCH3 in situ. We found that fluorescent PTE-ET-18-OCH3 colocalized with Fas-containing membrane rafts into caps on one or two poles of the Jurkat cells (Fig. 8). Time course experiments showed that ET-18-OCH3 rapidly accumulated in lipid raft clusters after 15–30 min of incubation (not depicted). Interestingly, PTE-ET-18-OCH3 remained accumulated in Fas-containing membrane clusters even after 15 h of treatment in Jurkat cells, when cells undergo apoptosis (not depicted), indicating that ET-18-OCH3 remained plasma membrane bound.
Figure 7.



Selective incorporation of the fluorescent analogue PTE-ET-18-OCH3 in human leukemic cells. Human leukemic Jurkat cells were incubated with the indicated concentrations of ET-18-OCH3 or PTE-ET-18-OCH3 for 24 h and apoptosis was measured by flow cytometry (A). Normal PBLs and leukemic HL-60 and Jurkat cells (JK) were treated with 20 μM PTE-ET-18-OCH3 for 7 h, washed with 1% BSA-PBS, and fluorescent drug uptake was analyzed by fluorescence microscopy (blue fluorescence) (B) or by fluorescence quantitation (C). Data shown are means ± SD of three independent determinations.
Figure 8.

Accumulation of the fluorescent analogue PTE-ET-18-OCH3 in Fas-enriched raft caps in leukemic cells. Jurkat cells were incubated with 20 μM PTE-ET-18-OCH3 (blue fluorescence) for 6 h, and then its colocalization with membrane rafts (A) and Fas (B) was examined using FITC-CTx B subunit (green fluorescence for rafts) and anti-Fas SM1/1 mAb, followed by CY3-conjugated anti–mouse antibody (red fluorescence for Fas). Images shown are representative of four independent experiments. Bar, 10 μm.
ET-18-OCH3 Spares Normal Human and Rat Hepatocytes.
Treatment of mice with an anti-Fas monoclonal agonistic antibody induces massive apoptosis of hepatocytes, which express abundant Fas, leading to fulminant hepatic failure and to the death of animals within a few hours (46). These hepatic effects prevent clinical use of exogenous Fas activation. Unlike agonistic cytotoxic anti-Fas CH-11 antibody that induced apoptosis in human hepatocytes, ET-18-OCH3 was not cytotoxic (Fig. 9). Rat hepatocytes were also spared (<3% apoptosis) after a 24-h treatment with either 10 or 20 μM ET-18-OCH3. We found that neither normal human nor rat hepatocytes incorporated significant amounts of ET-18-OCH3 (<0.2 nmol/106 cells after 6 h of incubation with 10 nmol [3H]ET-18-OCH3; n = 3), and therefore intracellular Fas activation was prevented.
Figure 9.

ET-18-OCH3 spares normal human hepatocytes. Human hepatocytes were untreated (C) or treated with 200 ng/ml cytotoxic anti-Fas CH-11 mAb or with 10 or 20 μM ET-18-OCH3 for 24 h, and then apoptosis was determined by flow cytometry. Data shown are means ± SD of three independent determinations.
Discussion
Here we describe a novel mechanism of action for an antitumor drug that involves: (a) drug insertion into lipid rafts promoting their clustering, (b) intracellular activation of Fas through its translocation and clustering into membrane raft caps on one pole of the cell, and (c) translocation of downstream Fas signaling molecules into Fas caps forming clusters of rafts enriched in apoptotic signaling molecules. This unique mode of action of the antitumor ether lipid ET-18-OCH3 (Edelfosine) illustrates the first use of raft-dependent and intracellular activated Fas-mediated killing in cancer chemotherapy, representing a new way to target tumor cells that can be of interest as a new framework in designing novel antitumor drugs. Evidence for the intracellular activation of the death receptor Fas by ET-18-OCH3 is manifold: (a) ET-18-OCH3 must be incorporated into the cells, likely into the cytoplasmic leaflet of the plasma membrane, to activate Fas and induce apoptosis; (b) a partial deletion of the intracellular domain of Fas blocks ET-18-OCH3–mediated apoptosis; (c) cells sensitive to ET-18-OCH3 become resistant when Fas is lost; and (d) Fas-deficient cells are resistant to ET-18-OCH3, but become sensitive upon enforced Fas expression. Furthermore, our data indicate that ET-18-OCH3 activates and translocates Fas into clustered membrane rafts, whereas other major death receptors, such as TNFR1 and TRAIL-R2/DR5 receptors, are not affected, suggesting a specificity for Fas. Thus, it is suggested that Fas is engaged by ET-18-OCH3, although whether the ether lipid interacts directly with Fas or activates the receptor through a more indirect way remains to be elucidated.
The synthesis of the first bioactive ET-18-OCH3 fluorescent analogue allowed us to visualize the location of the ether lipid in membrane rafts, in agreement with a recent report using [3H]ET-18-OCH3 (47). Here we found that fluorescent PTE-ET-18-OCH3 colocalized with Fas and membrane rafts, suggesting that ET-18-OCH3 could interact with both rafts and Fas intracellularly, and then ET-18-OCH3 self-aggregates translocating Fas into the lipid rafts. It could be argued that the presence of an additional lipid, ET-18-OCH3, might affect isolation of rafts through their resistance to solubilization by nonionic Triton X-100 at 4°C. This latter is the most widely used approach for raft isolation, despite its limitations and the fact that the highly asymmetric nature of the bilayer likely could lead to anomalous processes during detergent solubilization (41, 48). However, the differential localization of some proteins between raft-enriched and nonraft fractions in untreated versus ET-18-OCH3–treated cells (Fig. 3), the agreement between confocal and biochemical data on Fas and TNFR1 localization after ET-18-OCH3 treatment (compare Figs. 2 and 3), and the fact that ET-18-OCH3 is expected to be in much lower amounts than endogenous raft lipids, suggest that this method is reliable for the analysis of protein translocation to lipid rafts after ether lipid treatment and that ET-18-OCH3 does not cause significant changes in the isolation behavior of rafts.
An interesting finding from this work is that ET-18-OCH3 must be inside the cell, likely the inner leaflet of the plasma membrane, to alter rafts that are supposed to be mainly present in the outer leaflet of the cell membrane. The behavior of rafts in the cytoplasmic leaflet of the bilayer is an important, unresolved issue, but it is suggested that raft-like structures are also present in the inner leaflet and that rafts in the two leaflets are coupled (41, 48). The difference in length of the acyl chains of sphingolipids can allow them to interdigitate with the lipids in the inner leaflet (18, 49) forming putative inner leaflet rafts. Confocal imaging and fluorescence spectroscopy have shown a correlation between like phases in the two leaflets of model membranes containing two phospholipid species (50). In this regard, it could be envisaged that the insertion of ET-18-OCH3 into these putative inner leaflet rafts might induce local and transient changes in the interactions with their outer leaflet counterparts.
The clinical usefulness of exogenous activation of Fas as well as of the other major death receptors, namely TNF and TRAIL receptors, by their respective ligands or agonists has been hampered by toxic side effects. Systemic administration of TNF causes a severe inflammatory response syndrome that resembles septic shock (51), administration of agonistic antibody to Fas in mice is lethal because of liver failure through massive hepatocyte apoptosis (46), and TRAIL induces apoptosis of human hepatocytes (52). Here, we found that both human and rat hepatocytes are spared after ET-18-OCH3 treatment. Our present and previous findings (5, 12) show that Fas engagement by ET-18-OCH3 selectively triggers apoptosis in tumor cells, whereas normal cells are spared. In this regard, additional normal cells, such as resting PBLs, neutrophils, normal bone marrow human hematopoietic progenitors, human umbilical vein endothelial cells, and bovine aortic endothelial cells, are resistant to treatment with ET-18-OCH3 (5, 8, 12). This is due to the fact that ET-18-OCH3 does not activate Fas extracellularly, but from inside the cell in a FasL-independent manner, and the ether lipid is taken up by tumor cells but not by normal cells. The molecular explanation for the selective uptake of ET-18-OCH3 by tumor cells, leading to drug selectivity for tumors, remains to be elucidated. ET-18-OCH3 contains just one long alkyl chain as well as a polar head group, conferring the molecule its amphiphilic properties, which implies that the lipid is water soluble and can be administered orally (3). The particular amphiphilic molecular structure of ET-18-OCH3 suggests that this ether phospholipid readily inserts into the outer leaflet of the plasma membrane, but due to its water-soluble head group, the lipid will not flip-flop spontaneously across the plasma membrane. Indirect evidence suggests that a protein structure could be involved in the ether lipid uptake. Suramin, a broad specificity membrane–impermeable inhibitor of ligand receptor interactions (53), inhibits both ET-18-OCH3 uptake and ET-18-OCH3–induced apoptosis in leukemic cells (5). A photoactivatable analogue of ET-18-OCH3 is able to interact with two proteins of MCF-7 breast cancer cells (54). Lem3p (ligand effect modulator 3 protein; reference 55) and LdMT (Leishmania donovani putative Miltefosine Transporter; reference 56) have been recently found to be required for the uptake of alkylphosphocholine drugs, including ET-18-OCH3, into Saccharomyces cerevisiae yeast and L. donovani parasite, respectively.
Fas and mitochondria have been involved previously in the ET-18-OCH3 apoptotic response (9–12, 14). The translocation of Bid, a Fas mitochondria linker (43, 44), into Fas-containing membrane rafts suggests that Bid could act as a bridge between Fas activation and mitochondria in ET-18-OCH3–induced apoptosis, providing a molecular explanation for the involvement of mitochondria into the Fas-mediated apoptotic response triggered by ET-18-OCH3. This mechanism of action brings together Fas- and mitochondria-mediated signaling pathways potentiating the efficient triggering of apoptosis. Our data also indicate that procaspase-10 is activated at a higher extent than procaspase-8 in ET-18-OCH3–treated cells, suggesting that caspase-10 could be a relatively stronger mediator than caspase-8 in the apoptotic response induced by ET-18-OCH3.
Our data indicate that ET-18-OCH3 is rapidly incorporated into membrane rafts forming caps at the cell surface, and then Fas and downstream Fas effectors are recruited into rafts enriched in apoptotic signaling molecules, which seem to function as concentrating platforms for receptors, adaptors, and signaling molecules required to elicit an efficient apoptotic response. Thus, the herein reported recruitment of Fas and downstream Fas effectors into membrane rafts can provide a mechanism for amplifying Fas signaling by reorganization of membrane microdomains and bringing molecules together in a well-defined and reduced space, facilitating interactions among different signaling molecules and pathways. Our results demonstrate that rafts can be targets for therapeutic intervention. However, the results reported here also indicate that clustering of rafts in the absence of Fas is not sufficient to mount an apoptotic response in ET-18-OCH3–treated cells. Thus, rafts behave as recruitment platforms to facilitate and potentiate downstream signaling events, and Fas functions as a critical trigger of the apoptotic process.
This ET-18-OCH3–mediated aggregation of Fas-containing raft membrane domains resembles ceramide-mediated clustering of Fas. Initial activation of Fas by FasL leads to caspase-8–mediated SMase translocation into membrane rafts and its subsequent activation-generating ceramide, which in turn promotes Fas clustering in ceramide-enriched membrane platforms and further Fas activation (15, 24). Ceramide-mediated Fas clustering requires ligand-bound Fas as C16-ceramide is unable to trigger Fas clustering in the absence of stimulatory anti-Fas antibody or FasL (24). Here we failed to detect SMase activation or any increase in ceramide production after ET-18-OCH3 treatment, and the action of ET-18-OCH3 is independent of FasL (12). Furthermore, we found ceramide inhibited ET-18-OCH3–induced apoptosis (unpublished data). Overall, these data suggest that the action of ET-18-OCH3 is independent of SMase-derived ceramide.
Our data indicate that ET-18-OCH3 is the first drug that selectively activates Fas in tumor cells and also constitutes the first death receptor–engaged drug without harmful side effects. These features are of great potential in cancer therapy. Elucidation of the mechanisms regulating ET-18-OCH3 uptake, intracellular Fas activation, and the formation of rafts enriched in apoptotic signaling molecules, may lead to the development of novel strategies in cancer therapy with no adverse effects on normal tissues. ET-18-OCH3, beyond its putative clinical importance, is an interesting model compound for the development of more selective anticancer drugs.
Acknowledgments
This work was supported by grants FIS-02/1199 and FIS-01/1048 from the Fondo de Investigación Sanitaria, grant SA-087/01 from Junta de Castilla y León, and grant BQU2000-1500 from the Ministerio de Ciencia y Tecnología of Spain.
The authors have no conflicting financial interests.
Abbreviations used in this paper: CTx, cholera toxin; DISC, death-inducing signaling complex; ET-18-OCH3, 1-O-octadecyl-2-O-methyl-rac-glycero-3-phosphocholine; FADD, Fas-associated death domain protein; FasL, Fas ligand; JNK, c-Jun amino-terminal kinase; SMase, sphingomyelinase; TLC, thin layer chromatography; TRAIL, TNF-related apoptosis-inducing ligand; TUNEL, TdT-mediated dUTP nick-end labeling.
References
- 1.Munder, P.G., and O. Westphal. 1990. Antitumoral and other biomedical activities of synthetic ether lysophospholipids. Chem. Immunol. 49:206–235. [PubMed] [Google Scholar]
- 2.Houlihan, W.J., M. Lohmeyer, P. Workman, and S.H. Cheon. 1995. Phospholipid antitumor agents. Med. Res. Rev. 15:157–223. [DOI] [PubMed] [Google Scholar]
- 3.Gajate, C., and F. Mollinedo. 2002. Biological activities, mechanisms of action and biomedical prospect of the antitumor ether phospholipid ET-18-OCH(3) (edelfosine), a proapoptotic agent in tumor cells. Curr. Drug Metab. 3:491–525. [DOI] [PubMed] [Google Scholar]
- 4.Jendrossek, V., and R. Handrick. 2003. Membrane targeted anticancer drugs: potent inducers of apoptosis and putative radiosensitisers. Curr. Med. Chem. Anti-Canc. Agents. 3:343–353. [DOI] [PubMed] [Google Scholar]
- 5.Mollinedo, F., J.L. Fernandez-Luna, C. Gajate, B. Martin-Martin, A. Benito, R. Martinez-Dalmau, and M. Modolell. 1997. Selective induction of apoptosis in cancer cells by the ether lipid ET-18-OCH3 (edelfosine): molecular structure requirements, cellular uptake, and protection by Bcl-2 and Bcl-X(L). Cancer Res. 57:1320–1328. [PubMed] [Google Scholar]
- 6.van Blitterswijk, W.J., H. Hilkmann, and G.A. Storme. 1987. Accumulation of an alkyl lysophospholipid in tumor cell membranes affects membrane fluidity and tumor cell invasion. Lipids. 22:820–823. [DOI] [PubMed] [Google Scholar]
- 7.Gajate, C., A. Santos-Beneit, M. Modolell, and F. Mollinedo. 1998. Involvement of c-Jun NH2-terminal kinase activation and c-Jun in the induction of apoptosis by the ether phospholipid 1-O-octadecyl-2-O-methyl-rac-glycero-3-phosphocholine. Mol. Pharmacol. 53:602–612. [DOI] [PubMed] [Google Scholar]
- 8.Ruiter, G.A., S.F. Zerp, H. Bartelink, W.J. van Blitterswijk, and M. Verheij. 1999. Alkyl-lysophospholipids activate the SAPK/JNK pathway and enhance radiation-induced apoptosis. Cancer Res. 59:2457–2463. [PubMed] [Google Scholar]
- 9.Cuvillier, O., E. Mayhew, A.S. Janoff, and S. Spiegel. 1999. Liposomal ET-18-OCH(3) induces cytochrome c-mediated apoptosis independently of CD95 (APO-1/Fas) signaling. Blood. 94:3583–3592. [PubMed] [Google Scholar]
- 10.Gajate, C., A.M. Santos-Beneit, A. Macho, M. Lazaro, A. Hernandez-De Rojas, M. Modolell, E. Munoz, and F. Mollinedo. 2000. Involvement of mitochondria and caspase-3 in ET-18-OCH(3)-induced apoptosis of human leukemic cells. Int. J. Cancer. 86:208–218. [DOI] [PubMed] [Google Scholar]
- 11.Vrablic, A.S., C.D. Albright, C.N. Craciunescu, R.I. Salganik, and S.H. Zeisel. 2001. Altered mitochondrial function and overgeneration of reactive oxygen species precede the induction of apoptosis by 1-O-octadecyl-2-methyl-rac-glycero-3-phosphocholine in p53-defective hepatocytes. FASEB J. 15:1739–1744. [DOI] [PubMed] [Google Scholar]
- 12.Gajate, C., R.I. Fonteriz, C. Cabaner, G. Alvarez-Noves, Y. Alvarez-Rodriguez, M. Modolell, and F. Mollinedo. 2000. Intracellular triggering of Fas, independently of FasL, as a new mechanism of antitumor ether lipid-induced apoptosis. Int. J. Cancer. 85:674–682. [DOI] [PubMed] [Google Scholar]
- 13.Mollinedo, F., and C. Gajate. 2004. FasL-independent activation of Fas. Fas Signaling. H. Wajant, editor. Landes Bioscience, Georgetown, TX. 1–15.
- 14.Gajate, C., and F. Mollinedo. 2001. The antitumor ether lipid ET-18-OCH(3) induces apoptosis through translocation and capping of Fas/CD95 into membrane rafts in human leukemic cells. Blood. 98:3860–3863. [DOI] [PubMed] [Google Scholar]
- 15.Grassme, H., A. Jekle, A. Riehle, H. Schwarz, J. Berger, K. Sandhoff, R. Kolesnick, and E. Gulbins. 2001. CD95 signaling via ceramide-rich membrane rafts. J. Biol. Chem. 276:20589–20596. [DOI] [PubMed] [Google Scholar]
- 16.Hueber, A.O., A.M. Bernard, Z. Herincs, A. Couzinet, and H.T. He. 2002. An essential role for membrane rafts in the initiation of Fas/CD95-triggered cell death in mouse thymocytes. EMBO Rep. 3:190–196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Scheel-Toellner, D., K. Wang, R. Singh, S. Majeed, K. Raza, S.J. Curnow, M. Salmon, and J.M. Lord. 2002. The death-inducing signalling complex is recruited to lipid rafts in Fas-induced apoptosis. Biochem. Biophys. Res. Commun. 297:876–879. [DOI] [PubMed] [Google Scholar]
- 18.Simons, K., and D. Toomre. 2000. Lipid rafts and signal transduction. Nat. Rev. Mol. Cell Biol. 1:31–39. [DOI] [PubMed] [Google Scholar]
- 19.Nagata, S. 1997. Apoptosis by death factor. Cell. 88:355–365. [DOI] [PubMed] [Google Scholar]
- 20.Peter, M.E., and P.H. Krammer. 1998. Mechanisms of CD95 (APO-1/Fas)-mediated apoptosis. Curr. Opin. Immunol. 10:545–551. [DOI] [PubMed] [Google Scholar]
- 21.Ashkenazi, A., and V.M. Dixit. 1998. Death receptors: signaling and modulation. Science. 281:1305–1308. [DOI] [PubMed] [Google Scholar]
- 22.Algeciras-Schimnich, A., L. Shen, B.C. Barnhart, A.E. Murmann, J.K. Burkhardt, and M.E. Peter. 2002. Molecular ordering of the initial signaling events of CD95. Mol. Cell. Biol. 22:207–220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kolesnick, R. 2002. The therapeutic potential of modulating the ceramide/sphingomyelin pathway. J. Clin. Invest. 110:3–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Grassme, H., A. Cremesti, R. Kolesnick, and E. Gulbins. 2003. Ceramide-mediated clustering is required for CD95-DISC formation. Oncogene. 22:5457–5470. [DOI] [PubMed] [Google Scholar]
- 25.Hueber, A.O. 2003. Role of membrane microdomain rafts in TNFR-mediated signal transduction. Cell Death Differ. 10:7–9. [DOI] [PubMed] [Google Scholar]
- 26.Cremesti, A.E., F.M. Goni, and R. Kolesnick. 2002. Role of sphingomyelinase and ceramide in modulating rafts: do biophysical properties determine biologic outcome? FEBS Lett. 531:47–53. [DOI] [PubMed] [Google Scholar]
- 27.Ratter, F., M. Germer, T. Fischbach, K. Schulze-Osthoff, M.E. Peter, W. Droge, P.H. Krammer, and V. Lehmann. 1996. S-adenosylhomocysteine as a physiological modulator of Apo-1-mediated apoptosis. Int. Immunol. 8:1139–1147. [DOI] [PubMed] [Google Scholar]
- 28.Oehm, A., I. Behrmann, W. Falk, M. Pawlita, G. Maier, C. Klas, M. Li-Weber, S. Richards, J. Dhein, B.C. Trauth, et al. 1992. Purification and molecular cloning of the APO-1 cell surface antigen, a member of the tumor necrosis factor/nerve growth factor receptor superfamily. Sequence identity with the Fas antigen. J. Biol. Chem. 267:10709–10715. [PubMed] [Google Scholar]
- 29.Gomez-Lechon, M.J., P. Lopez, T. Donato, A. Montoya, A. Larrauri, P. Gimenez, R. Trullenque, R. Fabra, and J.V. Castell. 1990. Culture of human hepatocytes from small surgical liver biopsies. Biochemical characterization and comparison with in vivo. In Vitro Cell. Dev. Biol. 26:67–74. [DOI] [PubMed] [Google Scholar]
- 30.Casado, M., M.J. Diaz-Guerra, L. Bosca, and P. Martin-Sanz. 1997. Differential regulation of nitric oxide synthase mRNA expression by lipopolysaccharide and pro-inflammatory cytokines in fetal hepatocytes treated with cycloheximide. Biochem. J. 327:819–823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Mollinedo, F., C. Burgaleta, G. Velasco, A.G. Arroyo, A. Acevedo, and I. Barasoain. 1992. Enhancement of human neutrophil functions by a monoclonal antibody directed against a 19-kDa antigen. J. Immunol. 149:323–330. [PubMed] [Google Scholar]
- 32.Cheng, P.C., M.L. Dykstra, R.N. Mitchell, and S.K. Pierce. 1999. A role for lipid rafts in B cell antigen receptor signaling and antigen targeting. J. Exp. Med. 190:1549–1560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Veldman, R.J., E.I. Pécheur, S.C.D. van IJzendoorn, J.W. Kok, and D. Hoekstra. 2003. Cell lipids: from isolation to functional dynamics. Essential Cell Biology. J. Davey and M. Lord, editors. Oxford University Press, Oxford. 317–347.
- 34.Veldman, R.J., N. Maestre, O.M. Aduib, J.A. Medin, R. Salvayre, and T. Levade. 2001. A neutral sphingomyelinase resides in sphingolipid-enriched microdomains and is inhibited by the caveolin-scaffolding domain: potential implications in tumour necrosis factor signalling. Biochem. J. 355:859–868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Itoh, N., and S. Nagata. 1993. A novel protein domain required for apoptosis. Mutational analysis of human Fas antigen. J. Biol. Chem. 268:10932–10937. [PubMed] [Google Scholar]
- 36.Dudich, E., L. Semenkova, I. Dudich, E. Gorbatova, N. Tochtamisheva, E. Tatulov, M. Nikolaeva, and G. Sukhikh. 1999. alpha-fetoprotein causes apoptosis in tumor cells via a pathway independent of CD95, TNFR1 and TNFR2 through activation of caspase-3-like proteases. Eur. J. Biochem. 266:750–761. [DOI] [PubMed] [Google Scholar]
- 37.Suliman, A., A. Lam, R. Datta, and R.K. Srivastava. 2001. Intracellular mechanisms of TRAIL: apoptosis through mitochondrial-dependent and -independent pathways. Oncogene. 20:2122–2133. [DOI] [PubMed] [Google Scholar]
- 38.Schneider, P., M. Thome, K. Burns, J.L. Bodmer, K. Hofmann, T. Kataoka, N. Holler, and J. Tschopp. 1997. TRAIL receptors 1 (DR4) and 2 (DR5) signal FADD-dependent apoptosis and activate NF-kappaB. Immunity. 7:831–836. [DOI] [PubMed] [Google Scholar]
- 39.Schon, A., and E. Freire. 1989. Thermodynamics of intersubunit interactions in cholera toxin upon binding to the oligosaccharide portion of its cell surface receptor, ganglioside GM1. Biochemistry. 28:5019–5024. [DOI] [PubMed] [Google Scholar]
- 40.Harder, T., P. Scheiffele, P. Verkade, and K. Simons. 1998. Lipid domain structure of the plasma membrane revealed by patching of membrane components. J. Cell Biol. 141:929–942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Brown, D.A., and E. London. 2000. Structure and function of sphingolipid- and cholesterol-rich membrane rafts. J. Biol. Chem. 275:17221–17224. [DOI] [PubMed] [Google Scholar]
- 42.Kischkel, F.C., S. Hellbardt, I. Behrmann, M. Germer, M. Pawlita, P.H. Krammer, and M.E. Peter. 1995. Cytotoxicity-dependent APO-1 (Fas/CD95)-associated proteins form a death-inducing signaling complex (DISC) with the receptor. EMBO J. 14:5579–5588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Li, H., H. Zhu, C.J. Xu, and J. Yuan. 1998. Cleavage of BID by caspase 8 mediates the mitochondrial damage in the Fas pathway of apoptosis. Cell. 94:491–501. [DOI] [PubMed] [Google Scholar]
- 44.Luo, X., I. Budihardjo, H. Zou, C. Slaughter, and X. Wang. 1998. Bid, a Bcl2 interacting protein, mediates cytochrome c release from mitochondria in response to activation of cell surface death receptors. Cell. 94:481–490. [DOI] [PubMed] [Google Scholar]
- 45.Legler, D.F., O. Micheau, M.A. Doucey, J. Tschopp, and C. Bron. 2003. Recruitment of TNF receptor 1 to lipid rafts is essential for TNFalpha-mediated NF-kappaB activation. Immunity. 18:655–664. [DOI] [PubMed] [Google Scholar]
- 46.Ogasawara, J., R. Watanabe-Fukunaga, M. Adachi, A. Matsuzawa, T. Kasugai, Y. Kitamura, N. Itoh, T. Suda, and S. Nagata. 1993. Lethal effect of the anti-Fas antibody in mice. Nature. 364:806–809. [DOI] [PubMed] [Google Scholar]
- 47.van der Luit, A.H., M. Budde, P. Ruurs, M. Verheij, and W.J. van Blitterswijk. 2002. Alkyl-lysophospholipid accumulates in lipid rafts and induces apoptosis via raft-dependent endocytosis and inhibition of phosphatidylcholine synthesis. J. Biol. Chem. 277:39541–39547. [DOI] [PubMed] [Google Scholar]
- 48.Munro, S. 2003. Lipid rafts: elusive or illusive? Cell. 115:377–388. [DOI] [PubMed] [Google Scholar]
- 49.Simons, K., and E. Ikonen. 1997. Functional rafts in cell membranes. Nature. 387:569–572. [DOI] [PubMed] [Google Scholar]
- 50.Korlach, J., P. Schwille, W.W. Webb, and G.W. Feigenson. 1999. Characterization of lipid bilayer phases by confocal microscopy and fluorescence correlation spectroscopy. Proc. Natl. Acad. Sci. USA. 96:8461–8466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Watanabe, S., N. Mukaida, N. Ikeda, M. Akiyama, A. Harada, I. Nakanishi, H. Nariuchi, Y. Watanabe, and K. Matsushima. 1995. Prevention of endotoxin shock by an antibody against leukocyte integrin beta 2 through inhibiting production and action of TNF. Int. Immunol. 7:1037–1046. [DOI] [PubMed] [Google Scholar]
- 52.Jo, M., T.H. Kim, D.W. Seol, J.E. Esplen, K. Dorko, T.R. Billiar, and S.C. Strom. 2000. Apoptosis induced in normal human hepatocytes by tumor necrosis factor-related apoptosis-inducing ligand. Nat. Med. 6:564–567. [DOI] [PubMed] [Google Scholar]
- 53.van der Bend, R.L., J. Brunner, K. Jalink, E.J. van Corven, W.H. Moolenaar, and W.J. van Blitterswijk. 1992. Identification of a putative membrane receptor for the bioactive phospholipid, lysophosphatidic acid. EMBO J. 11:2495–2501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Li, G., P. Samadder, G. Arthur, and R. Bittman. 2001. Synthesis and antiproliferative properties of a photoactivatable analogue of ET-18-OCH3. Tetrahedron. 57:8925–8932. [Google Scholar]
- 55.Hanson, P.K., L. Malone, J.L. Birchmore, and J.W. Nichols. 2003. Lem3p is essential for the uptake and potency of alkylphosphocholine drugs, edelfosine and miltefosine. J. Biol. Chem. 278:36041–36050. [DOI] [PubMed] [Google Scholar]
- 56.Perez-Victoria, F.J., F. Gamarro, M. Ouellette, and S. Castanys. 2003. Functional cloning of the miltefosine transporter. A novel P-type phospholipid translocase from Leishmania involved in drug resistance. J. Biol. Chem. 278:49965–49971. [DOI] [PubMed] [Google Scholar]