

Abstract
Although most leukocytes, T lymphocytes in particular, respond to several different chemokines, there is virtually no information on chemokine activities and chemokine receptors in B lymphocytes. A putative chemokine receptor, BLR1, that is expressed in Burkitt's lymphoma cells and B lymphocytes was cloned a few years ago. Deletion of the gene for BLR1 yielded mice with abnormal primary follicles and germinal centers of the spleen and Peyer's patches, reflecting the inability of B lymphocytes to migrate into B cell areas. By screening expressed sequence tag DNA sequences, we have identified a CXC chemokine, termed B cell–attracting chemokine 1 (BCA-1), that is chemotactic for human B lymphocytes. BCA-1 cDNA encodes a protein of 109 amino acids with a leader sequence of 22 residues. The mature protein shares 23–34% identical amino acids with known CXC chemokines and is constitutively expressed in secondary lymphoid organs. BCA-1 was chemically synthesized and tested for activity on murine pre–B cells 300-19 transfected with BLR1 and on human blood B lymphocytes. In transfected cells, BCA-1 induced chemotaxis and Ca2+ mobilization demonstrating that it acts via BLR1. Under the same conditions, no activity was obtained with 10 CXC and 19 CC chemokines, lymphotactin, neurotactin/fractalkine and several other peptide ligands. BCA-1 was also a highly effective attractant for human blood B lymphocytes (which express BLR1), but was inactive on freshly isolated or IL-2–stimulated T lymphocytes, monocytes, and neutrophils. In agreement with the nomenclature rules for chemokine receptors, we propose the term CXCR5 for BLR1. Together with the observed disturbance of B cell colonization in BLR1/ CXCR5-deficient mice, the present results indicate that chemotactic recruitment by locally produced BCA-1 is important for the development of B cell areas of secondary lymphoid tissues.
In the past few years, major progress has been made in the understanding of the role of chemokines in T lymphocyte recruitment (1). T cells respond to CXC and CC chemokines and express many different chemokine receptors, some of which are upregulated by IL-2 (2). Of particular interest is a new group of CC chemokines that were identified recently, such as TARC (3), ELC/MIP-3β (4, 5), LARC/MIP-3α/exodus (5–7), and SLC (8), which are expressed preferentially in lymphoid tissues and are chemotactic for T lymphocytes. Another CC chemokine, DC-CK1/ PARC (9, 10) is expressed by dendritic cells of germinal centers and was reported to attract naive T lymphocytes (9), suggesting a role for the onset of immune reactions.
There is little information, however, on chemokine activities and chemokine receptor expression on B lymphocytes. Two newly identified CC chemokine receptors, CCR6 (11) and CCR7 (4), were reported to be present in T and B cells, but no ligand was found that induces chemotaxis of B lymphocytes. SDF-1, a CXC chemokine that was originally described as a growth factor for progenitor B cells, is chemotactic for human pre– and pro–B cell lines, but does not attract mature B lymphocytes (12). The search for chemokines that attract B cells was stimulated by the cloning of a receptor termed BLR1 (13) or MDR15 (14) that is highly expressed in Burkitt's lymphoma cells and B lymphocytes and has considerable structural similarity to known chemokine receptors. Deletion of the gene for BLR1 in mice (15) yielded a remarkable phenotype. The −/− animals lack inguinal lymph nodes and have a defective formation of primary follicles and germinal centers of the spleen and of Peyer's patches. Receptor-deficient B lymphocytes enter T cell areas, but do not migrate into B cell areas suggesting that the BLR1 ligand is essential for the homing of B lymphocytes and the proper development of B cell–rich regions of lymphoid tissues.
To search for the ligand of BLR1/MDR15, we screened the expressed sequence tag (EST) data bank for CXC chemokine motifs and cloned a yet uncharacterized CXC chemokine termed B cell–attracting chemokine (BCA)-1, which attracts cells transfected with BLR1. The present paper shows that BCA-1 is expressed in lymphoid tissues and is a selective and highly efficacious chemoattractant for human blood B lymphocytes.
Materials and Methods
Agonists.
BCA-1, reference chemokines, and neuropeptide Y (NPY) were chemically synthesized according to established protocols (16). The novel chemokines I-TAC and HCC-2 were supplied by Dr. K. Neote (Pfizer Central Research, Groton, CT) and Dr. W.G. Forssmann (Lower Saxony Institute for Peptide Research (IPF), Hannover, Germany). C5a and C3a were provided by Dr. C.A. Dahinden (University Hospital, Bern, Switzerland). Somatostatin, substance P, and calcitonin were purchased from Sigma Chemical Co. (St. Louis, MO).
Cells.
Human neutrophils (17), monocytes (18), and PBLs were isolated from donor blood buffy coats, and PBLs were activated by culturing with IL-2 (2). B lymphocytes were isolated from blood mononuclear cells by positive selection with a mouse anti–human CD20 mAb (Immunotech, Marseille, France) and MACS goat anti–mouse IgG microbeads (19) after depletion of monocytes by adherence to plastic. Purity was assessed by FACS® analysis (mouse anti–human CD19, Sigma Chemical Co.; mouse anti–human CD3, Becton Dickinson, Mountain View, CA). Before use in chemotaxis and Ca2+ mobilization assays, B lymphocytes were incubated for 30 min at 37°C.
Cloning of BCA-1.
The coding sequence of BCA-1 was generated by PCR using human spleen cDNA as template and the primers BCA-1SE, 5′-GGGCTGCAGCTCCAGACAGAATGAAGTTC, containing the start codon, and BCA-1AS, 5′-GGACTCGAGTGGAAATATCAGCATCAGGG, including the stop codon. PCR amplification was performed in volumes of 30 μl, containing 1× PCR buffer including 1.5 mM MgCl2 (GIBCO BRL, Paisley, UK), 0.4 μM primers, 280 μM dNTPs, and 2 μl of cDNA preparation. To start DNA synthesis, 1.5 U Taq-polymerase (GIBCO BRL) was added after cDNA denaturation and allowed to progress for 30 cycles (1 min at 57°C, 45 s at 72°C, 15 s at 95°C) in a master cycler (5330; Eppendorf-Netheler-Hinz, Hamburg, Germany). The amplified DNA was purified, digested with PstI and XhoI, subcloned into the vector pBluescript SK (Stratagene Corp., La Jolla, CA), and three independent clones were sequenced to completion.
BLR1 Transfectants.
BLR1 DNA was generated by PCR with cDNA from freshly isolated PBLs as template and the primers BLR1SE, 5′-TTAGAATTCGACTCACAGCCGGCACAGC, and BLR1AS, 5′-GTGAGCCCTTAGGATCCCAGC, corresponding to positions 65–83 and 1,276–1,296 of the published sequence of BLR1 (13), spanning the entire coding region of 372 amino acids. PCR amplification was performed as described (20). The amplified DNA was purified, digested with EcoRI and BamHI, subcloned into the mammalian expression vector SRαpuro (provided by Dr. F. Arenzana-Seisdedos, Pasteur Institute, Paris, France), and sequenced. Stable transfectants were generated with 20 μg of linearized SRαpuro-BLR1 plasmid DNA in murine pre–B cells 300–19 (5 × 106 cells in 400 μl RPMI 1640; reference 21). Clones were established by limiting dilution in the presence of 1.5 μg/ml puromycin (Sigma Chemical Co.) and screened for BLR1 expression by RNA dot blot analysis.
Ca2+ Responses to Chemokines.
Changes in the cytosolic free intracellular Ca2+ concentration ([Ca2+]i) were measured in cells loaded with fura-2 by incubation for 25 min at 37°C with 0.1 nmol fura-2/AM per 106 cells in a buffer containing 136 mM NaCl, 4.8 mM KCl, 1 mM CaCl2, 5 mM glucose, and 20 mM Hepes, pH 7.4. After centrifugation, loaded cells were resuspended in the same buffer (2 × 106 cells in 800 μl), stimulated with the indicated chemokine at 37°C, and the [Ca2+]i-related fluorescence changes were recorded (22).
In Vitro Chemotaxis.
Cell migration was assessed in triplicate in 48-well chambers as described by Loetscher et al. (23). RPMI 1640 supplemented with 20 mM Hepes, pH 7.4, and 1% pasteurized human plasma protein solution (Swiss Red Cross Laboratory, Bern, Switzerland) was used to dissolve the chemokines and to suspend the cells. The duration of the assay was 120 min for BLR1 transfectants, parental 300-19 cells, and B lymphocytes; 60 min for T lymphocytes; and 25 min for neutrophils and monocytes. Migrating cells were stained on the lower side of the filters and counted at ×1,000 magnification in five randomly selected fields.
Expression was measured by Northern and dot blot analysis using a human RNA Master BlotTM and a human Immune System Multiple Tissue Blot IITM (Clontech, Palo Alto, CA) as described (2, 23). A 32P-labeled complete BCA-1 DNA (2 × 109 cpm/μg) was used as hybridization probe at 5 × 106 cpm/ml hybridization solution.
Results and Discussion
BCA-1 DNA and Protein.
A partial sequence of a putative human CXC chemokine was recently derived from the EST DNA sequence T39765 (24). Using T39765 as probe we searched the EST cDNA databank and identified two additional sequences, AA298459 and T55152, providing the missing 5′- and 3′-sequence information. The resulting DNA of 663 nucleotides has a putative open reading frame of 327 nucleotides preceded by a stretch with high similarity to the consensus of translation initiation sequences of vertebrates (25). DNA containing the whole coding region of BCA-1 was generated by PCR using spleen cDNA and specific oligonucleotide primers. Three independent clones were sequenced to completion and found to be identical. They differ from the EST-derived sequence at position 47–48 (GC instead of CG) yielding a threonine instead of a serine at position 16. The translated cDNA consists of 109 amino acids (Fig. 1 a) and contains a hydrophobic sequence followed by a putative cleavage site between residues 22 and 23. The predicted secreted form of 87 residues was chemically synthesized and shown to activate cells expressing BLR1, indicating that it is a functional form of the chemokine. In view of the biological activities described below the chemokine was termed BCA-1. The four cysteines of BCA-1 align with those of CXC chemokines. In addition BCA-1 has an arginine residue immediately preceding the first cysteine, a feature that is conserved in all CXC chemokines except platelet factor 4. Overall, it shares 24–34% sequence identity with CXC chemokines. However, evolutionary distance comparison indicates that BCA-1 is not closely related to any particular group of chemokines. Like SDF-1, it appears to be segregated within the CXC chemokine subfamily (Fig. 1 b).
Figure 1.
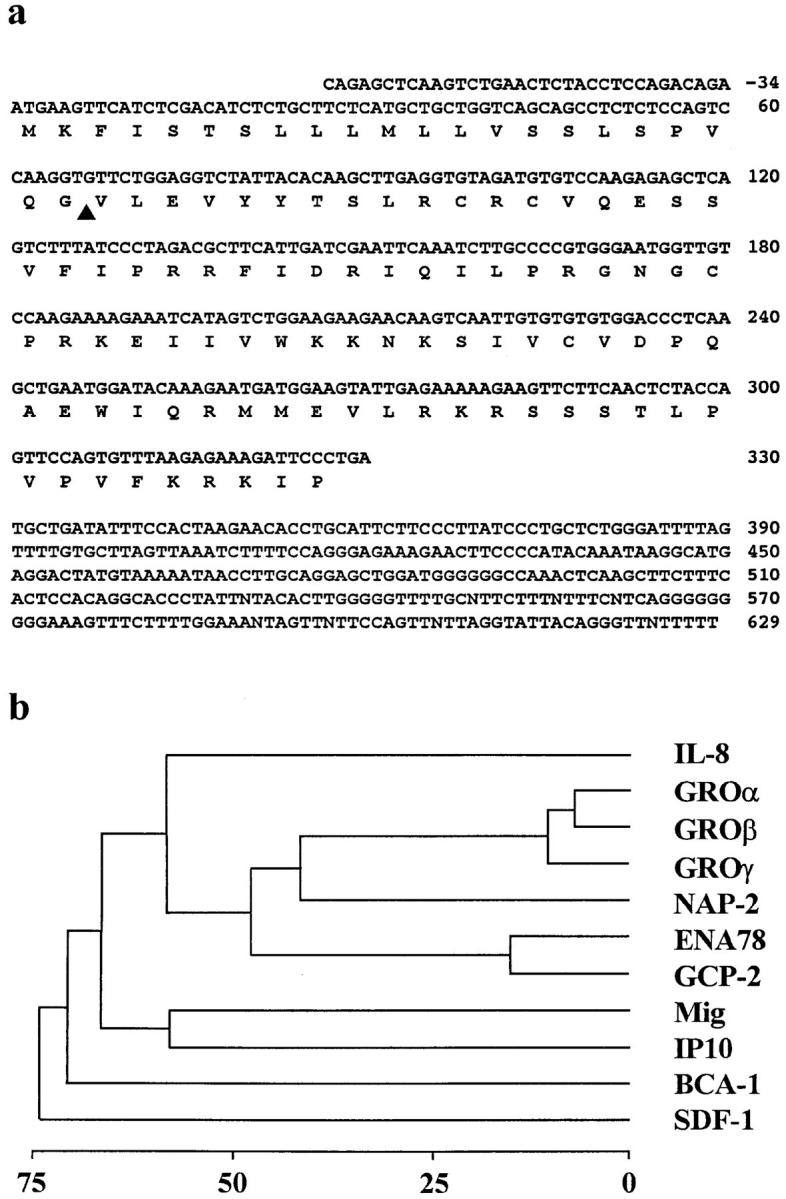
(a) Nucleotide and deduced amino acid sequences of BCA-1. Numbering starts at the first nucleotide of the start codon. The predicted open reading frame is indicated below the nucleotide sequence, and the putative signal peptide cleavage site is marked by an arrowhead. These sequence data are available from EMBL/Genbank/DDBJ under accession number AJ002211. (b) Phylogenetic tree diagram of human CXC chemokines. Similarity scores of the proteins were determined by average linkage cluster analysis (33). Gap penalty, window size, filtration level, and K-tulpe size parameters for pairwise alignments were set at 3, 10, 2.5, and 1, respectively. Distance to the branching points indicates percent sequence divergence.
Expression of BCA-1 was assessed with a DNA hybridization probe of 355 bp spanning the entire coding region. In a dot blot analysis of 50 human tissue RNAs (Fig. 2 a), the highest expression was found in the liver followed by the spleen, lymph nodes, appendix, and stomach. Some expression was also detected in the salivary and mammary glands, whereas all other tissues tested were negative. Survey of a number of fetal tissues yielded some expression in the spleen only (data not shown). A Northern blot with a selection of lymphoid and myeloid RNAs showed high levels of a single transcript of 1.4 kb in spleen and lymph nodes, but not in the thymus, bone marrow, and blood lymphocytes (Fig. 2 b). Since the RNA for dot and Northern blot analysis was obtained from normal tissues, BCA-1 expression in lymphoid tissues appears to be constitutive.
Figure 2.

Expression of BCA-1 in human tissues. (a) Dot blot analysis using a DNA probe spanning the whole open reading frame of BCA-1. The strongest signal, obtained with liver RNA (140,575 cpm), was taken as 100%, and the relative expression in other tissues was calculated. The lowest significant signal (2,759 cpm) was detected in the mammary gland. Most tissues yielded counts around background (∼50 cpm). Tissue samples that did not express BCA-1 transcripts included total brain, amygdala, caudate nucleus, cerebellum, cerebral cortex, frontal lobe, hippocampus, medulla oblongata, occipital lobe, putamen, substantia nigra, temporal lobe, thalamus, subthalamic nucleus, spinal cord, heart, aorta, skeletal muscle, colon, bladder, uterus, prostate, testis, ovary, pancreas, pituitary gland, adrenal gland, thyroid gland, kidney, small intestine, thymus, blood leukocytes, bone marrow, lung, trachea, placenta, fetal brain, fetal heart, fetal kidney, fetal liver, fetal thymus, and fetal lung. (b) Northern blot of human tissue RNAs hybridized with the same DNA probe.
BCA-1 Is the Ligand for BLR1.
An elegant study by Förster et al. (15) showed that disruption of the BLR1 gene in mice results in a defective formation of primary follicles and germinal centers of the spleen and of Peyer's patches and the lack of inguinal lymph nodes. Receptor-deficient B lymphocytes enter the T cell areas, but fail to migrate into the areas that are normally populated by B cells. These observations suggested that chemokines generated in homing tissues could direct the influx and the retention of BLR1-bearing B cells. Since BCA-1 is highly expressed in secondary lymphoid tissues, it was considered a promising candidate ligand for BLR1. Synthetic BCA-1 was first tested on clones of the murine pre–B cell line 300-19 stably transfected with BLR1 DNA. This cell line has been successfully used in our laboratory to express human chemokine receptors and to assess chemokine-dependent functions (20, 23). As shown in Fig. 3 a, BCA-1 induced chemotaxis of BLR1-expressing cells with maximum migration at 100 nM, whereas the parental, nontransfected cells did not respond. Furthermore a transient rise in [Ca2+]i was elicited by stimulation of BLR1 transfectants, but not of parental 300-19 cells (Fig. 3 b). A response was observed already at 1 nM BCA-1, and the rate and magnitude of the [Ca2+]i rise increased with the agonist concentration. 10 CXC and 19 CC chemokines, lymphotactin, neurotactin, as well as several other peptide ligands (Table 1) were tested and found to be inactive. These results demonstrate that the CXC chemokine BCA-1 is the ligand for BLR1, and show that no other known chemokine is recognized by this receptor. According to the current chemokine receptor nomenclature, we propose the term CXCR5 for the orphan receptor formerly known as BLR1.
Figure 3.


BCA-1 responses of murine pre–B cells 300-19 stably transfected with BLR1/ CXCR5. (a) Chemotaxis of parental (open circles) and BLR1/ CXCR5-expressing cells (closed circles). Migrated cells (mean counts ± SD of triplicate assays per five high power fields) from one experiment that is representative of a total of six. (b) [Ca2+]i changes in BLR1/CXCR5-expressing 300-19 cells. BCA-1 was added at 0.1, 1, 10, and 100 nM to fura-2–loaded cells (arrowheads), and [Ca2+]i-dependent fluorescence changes were recorded. Parental 300-19 cells (control) were stimulated with 100 nM BCA-1. One out of five independent experiments with identical results is shown.
Table 1.
Chemokines and Other Protein Agonists Tested for Activity on Cells Expressing BLR1/CXCR5*
| CXC chemokines | CC chemokines | Other chemokines | Other peptides | |||
|---|---|---|---|---|---|---|
| BCA-1 | RANTES | Lymphotactin | C3a | |||
| IL-8 | MCP-1 | Neurotactin | C5a | |||
| GROα | MCP-2 | Calcitonin | ||||
| NAP-2 | MCP-3 | NPY | ||||
| GCP-2 | MCP-4 | Somatostatin | ||||
| ENA78 | Eotaxin | Substance P | ||||
| Mig | Eotaxin-2 | |||||
| IP10 | MIP-1α | |||||
| I-TAC | MIP-1β | |||||
| SDF-1 | I-309 | |||||
| PF4 | HCC-1 | |||||
| HCC-2 | ||||||
| TARC | ||||||
| LARC | ||||||
| DC-CK1 | ||||||
| CKβ8 | ||||||
| ELC | ||||||
| SLC | ||||||
| MDC |
[Ca2+]i changes were determined in stably transfected 300-19 cells after stimulation with 100 nM agonist.
BCA-1 Is Selective for B Lymphocytes.
The chemotactic activity of BCA-1 was then tested on B and T cells, monocytes, and neutrophils. As shown in Fig. 4 a, B lymphocytes, which express high levels of BLR1/CXCR5 (13), migrated in a concentration-dependent manner in response to BCA-1. The efficacy of BCA-1 was extraordinarly high, as >60% of the input cells were recovered on the lower surface of the filter or in the lower well at the highest agonist concentration. The potency, however, was moderate since concentrations of at least 100 nM were needed. The reason for the superior efficacy but lower potency of BCA-1 in B lymphocytes as compared to BLR1-transfected 300-19 cells is not clear at present. BCA-1 was not chemotactic for T lymphocytes, monocytes, and neutrophils (Fig. 4 a). Controls showed that these leukocytes migrated normally in response to SDF-1, MCP-1, and IL-8, respectively (data not shown). As shown in Fig. 4 b, BCA-1 did not induce [Ca2+]i changes in B lymphocytes, even at concentrations as high as 1 μM. As expected, BCA-1 also did not induce Ca2+ mobilization in T lymphocytes, monocytes, and neutrophils, and did not prevent the effects of the chemokines that induce chemotaxis in these cells (Fig. 4 b).
Figure 4.


Functional responses of human blood leukocytes to BCA-1. (a) In vitro chemotaxis of human B lymphocytes (closed circles), T lymphocytes cultured for 19 d in the presence of 250 U/ml IL-2 (open circles), monocytes (open squares), and neutrophils (open triangles) in response to increasing concentrations of BCA-1. (b) [Ca2+]i changes in human blood leukocytes. Fura-2–loaded cells were stimulated sequentially with 100 nM BCA-1 followed by another chemokine, and changes in [Ca2+]i were recorded. The data are representative for four independent experiments.
The pattern of biological activity of BCA-1 is somewhat unusual. It is selective for B lymphocytes that are attracted with high efficacy and relatively low potency. In terms of efficacy, BCA-1 is similar to SDF-1, which was shown to attract unexpectedly high numbers of T lymphocytes in vitro (26). Another feature is its constitutive rather than induced expression, and in this respect, BCA-1 is again similar to SDF-1 (27). BCA-1, is unique among chemokines in its selectivity for B cells, whereas SDF-1 acts via a broadly distributed receptor and attracts T lymphocytes, monocytes, neutrophils, as well as B cell precursors (12, 26, 28, 29). Despite its high chemotactic efficacy, BCA-1 did not induce [Ca2+]i changes in B cells. It is known from studies with neutrophils that the shape change induced by chemoattractants, which is regarded as a correlate to cellular motion, does not require intracellular Ca2+ mobilization (30). The mechanism of signaling by chemoattractants in B cells has not been studied, and experiments with BCA-1 may uncover differences with respect to other leukocytes.
It has been recently recognized that certain chemokines are involved in the regulation of lymphocyte homing rather than in the recruitment of leukocytes into inflamed tissues. It was suggested that DC-CK1/PARC, a chemokine that is selectively expressed by dendritic cells of germinal centers, attracts naive T lymphocytes and may thus be involved in the initiation of an immune response (9, 10). SDF-1 is consitutively produced by stromal cells of the bone marrow and was originally identified as a factor that stimulates the growth of immature B cells (31). Since SDF-1 is chemotactic for pro– and pre–B lymphocytes, it may act in lymphopoiesis by attracting these progenitors into close contact with stromal cells where growth and differentiation factors are released (12). It was also proposed that SDF-1 may direct the homing of progenitor cells to their bone marrow maturation sites (32), and may support bone marrow colonization by hematopoietic precursor cells during embryogenesis (26). The finding that the ligand for BLR1 is a chemokine provides further evidence for chemokine functions that are not related to inflammation and immune pathology. Together with the observed disturbance of B cell colonization in BLR1/CXCR5-deficient mice (15), the present results indicate that chemotactic recruitment by locally produced BCA-1 and possibly other B cell–attracting chemokines is important for the development of B cell areas of secondary lymphoid tissues.
Footnotes
We thank Dr. Luca Piali for FACS® analysis of the cell populations, Dr. Beatrice Dewald for critical reading of the manuscript, and the “Hochschulstiftung” of the University of Bern (Bern, Switzerland) for financing a Master Cycler PCR instrument. Donor blood buffy coats were provided by the Swiss Central Laboratory Blood Transfusion Service, Bern, Switzerland.
This work was supported by grant 31-39744.93 of the Swiss National Science Fundation to M. Baggiolini and B. Moser, and the Protein Engineering Network of Centers of Excellence (PENCE), Canada (to I. Clark-Lewis). The Professor Max Cloëtta Foundation, Zürich, supports B. Moser with a career development award.
Address correspondence to Marco Baggiolini, Theodor-Kocher Institute, University of Bern, PO Box 99, CH-3000 Bern 9, Switzerland. Phone: 41-31-631-4141; Fax: 41-31-631-3799; E-mail: marco.baggiolini@tki.unibe.ch
References
- 1.Baggiolini M, Dewald B, Moser B. Human chemokines: an update. Annu Rev Immunol. 1997;15:675–705. doi: 10.1146/annurev.immunol.15.1.675. [DOI] [PubMed] [Google Scholar]
- 2.Loetscher P, Seitz M, Baggiolini M, Moser B. Interleukin-2 regulates CC chemokine receptor expression and chemotactic responsiveness in T lymphocytes. J Exp Med. 1996;184:569–577. doi: 10.1084/jem.184.2.569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Imai T, Yoshida T, Baba M, Nishimura M, Kakizaki M, Yoshie O. Molecular cloning of a novel T cell–directed CC chemokine expressed in thymus by signal sequence trap using Epstein-Barr virus vector. J Biol Chem. 1996;271:21514–21521. doi: 10.1074/jbc.271.35.21514. [DOI] [PubMed] [Google Scholar]
- 4.Yoshida R, Imai T, Hieshima K, Kusuda J, Baba M, Kitaura M, Nishimura M, Kakizaki M, Nomiyama H, Yoshie O. Molecular cloning of a novel human CC chemokine EBI1-ligand chemokine that is a specific functional ligand for EBI1, CCR7. J Biol Chem. 1997;272:13803–13809. doi: 10.1074/jbc.272.21.13803. [DOI] [PubMed] [Google Scholar]
- 5.Rossi DL, Vicari AP, Franz-Bacon K, McClanahan TK, Zlotnik A. Identification through bioinformatics of two new macrophage proinflammatory human chemokines–MIP-3α and MIP-3β. J Immunol. 1997;158:1033–1036. [PubMed] [Google Scholar]
- 6.Hieshima K, Imai T, Opdenakker G, Van Damme J, Kusuda J, Tei H, Sakaki Y, Takatsuki K, Miura R, Yoshie O, et al. Molecular cloning of a novel human CC chemokine liver and activation-regulated chemokine (LARC) expressed in liver–chemotactic activity for lymphocytes and gene localization on chromosome. J Biol Chem. 1997;272:5846–5853. doi: 10.1074/jbc.272.9.5846. [DOI] [PubMed] [Google Scholar]
- 7.Hromas R, Gray PW, Chantry D, Godiska R, Krathwohl M, Fife K, Bell GI, Takeda J, Aronica S, Gordon M, et al. Cloning and characterization of exodus, a novel β-chemokine. Blood. 1997;89:3315–3322. [PubMed] [Google Scholar]
- 8.Nagira M, Imai T, Hieshima K, Kusuda J, Ridanpää M, Takagi S, Nishimura M, Kakizaki M, Nomiyama H, Yoshie O. Molecular cloning of a novel human CC chemokine secondary lymphoid-tissue chemokine that is a potent chemoattractant for lymphocytes and mapped to chromosome 9p13. J Biol Chem. 1997;272:19518–19524. doi: 10.1074/jbc.272.31.19518. [DOI] [PubMed] [Google Scholar]
- 9.Adema GJ, Hartgers F, Verstraten R, De Vries E, Marland G, Menon S, Foster J, Xu YM, Nooyen P, McClanahan T, et al. A dendritic-cell–derived C-C chemokine that preferentially attracts naive T cells. Nature. 1997;387:713–717. doi: 10.1038/42716. [DOI] [PubMed] [Google Scholar]
- 10.Hieshima K, Imai T, Baba M, Shoudai K, Ishizuka K, Nakagawa T, Tsuruta J, Takeya M, Sakaki Y, Takatsuki K, et al. A novel human CC chemokine PARC that is most homologous to macrophage-inflammatory protein-1α/ LD78α and chemotactic for T lymphocytes, but not for monocytes. J Immunol. 1997;159:1140–1149. [PubMed] [Google Scholar]
- 11.Baba M, Imai T, Nishimura M, Kakizaki M, Takagi S, Hieshima K, Nomiyama H, Yoshie O. Identification of CCR6, the specific receptor for a novel lymphocyte-directed CC chemokine LARC. J Biol Chem. 1997;272:14893–14898. doi: 10.1074/jbc.272.23.14893. [DOI] [PubMed] [Google Scholar]
- 12.D'Apuzzo M, Rolink A, Loetscher M, Hoxie JA, Clark-Lewis I, Melchers F, Baggiolini M, Moser B. The chemokine SDF-1, stromal cell–derived factor 1, attracts early stage B cell precursors via the chemokine receptor CXCR4. Eur J Immunol. 1997;27:1788–1793. doi: 10.1002/eji.1830270729. [DOI] [PubMed] [Google Scholar]
- 13.Dobner T, Wolf I, Emrich T, Lipp M. Differentiation-specific expression of a novel G protein–coupled receptor from Burkitt's lymphoma. Eur J Immunol. 1992;22:2795–2799. doi: 10.1002/eji.1830221107. [DOI] [PubMed] [Google Scholar]
- 14.Barella L, Loetscher M, Tobler A, Baggiolini M, Moser B. Sequence variation of a novel heptahelical leucocyte receptor through alternative transcript formation. Biochem J. 1995;309:773–779. doi: 10.1042/bj3090773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Förster R, Mattis AE, Kremmer E, Wolf E, Brem G, Lipp M. A putative chemokine receptor, BLR1, directs B cell migration to defined lymphoid organs and specific anatomic compartments of the spleen. Cell. 1996;87:1037–1047. doi: 10.1016/s0092-8674(00)81798-5. [DOI] [PubMed] [Google Scholar]
- 16.Clark-Lewis I, Moser B, Walz A, Baggiolini M, Scott GJ, Aebersold R. Chemical synthesis, purification, and characterization of two inflammatory proteins, neutrophil activating peptide 1 (interleukin-8) and neutrophil activating peptide 2. Biochemistry. 1991;30:3128–3135. doi: 10.1021/bi00226a021. [DOI] [PubMed] [Google Scholar]
- 17.Peveri P, Walz A, Dewald B, Baggiolini M. A novel neutrophil-activating factor produced by human mononuclear phagocytes. J Exp Med. 1988;167:1547–1559. doi: 10.1084/jem.167.5.1547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Uguccioni M, D'Apuzzo M, Loetscher M, Dewald B, Baggiolini M. Actions of the chemotactic cytokines MCP-1, MCP-2, MCP-3, RANTES, MIP-1α and MIP-1β on human monocytes. Eur J Immunol. 1995;25:64–68. doi: 10.1002/eji.1830250113. [DOI] [PubMed] [Google Scholar]
- 19.Miltenyi S, Mueller W, Weichel W, Radbruch A. High gradient magnetic cell separation with MACS. Cytometry. 1990;11:231–238. doi: 10.1002/cyto.990110203. [DOI] [PubMed] [Google Scholar]
- 20.Roos RS, Loetscher M, Legler DF, Clark-Lewis I, Baggiolini M, Moser B. Identification of CCR8, the receptor for the human CC chemokine I-309. J Biol Chem. 1997;272:17251–17254. doi: 10.1074/jbc.272.28.17251. [DOI] [PubMed] [Google Scholar]
- 21.Moser B, Barella L, Mattei S, Schumacher C, Boulay F, Colombo MP, Baggiolini M. Expression of transcripts for two interleukin 8 receptors in human phagocytes, lymphocytes and melanoma cells. Biochem J. 1993;294:285–292. doi: 10.1042/bj2940285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.von Tscharner V, Prod'hom B, Baggiolini M, Reuter H. Ion channels in human neutrophils activated by a rise in free cytosolic calcium concentration. Nature. 1986;324:369–372. doi: 10.1038/324369a0. [DOI] [PubMed] [Google Scholar]
- 23.Loetscher M, Gerber B, Loetscher P, Jones SA, Piali L, Clark-Lewis I, Baggiolini M, Moser B. Chemokine receptor specific for IP10 and Mig: structure, function, and expression in activated T-lymphocytes. J Exp Med. 1996;184:963–969. doi: 10.1084/jem.184.3.963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wells TNC, Peitsch MC. The chemokine information source: identification and characterization of novel chemokines using the world wide web and expressed sequence tag databases. J Leukocyte Biol. 1997;61:545–550. doi: 10.1002/jlb.61.5.545. [DOI] [PubMed] [Google Scholar]
- 25.Kozak M. An analysis of 5′-noncoding sequences from 699 vertebrate messenger RNAs. Nucleic Acids Res. 1987;15:8125–8148. doi: 10.1093/nar/15.20.8125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Bleul CC, Fuhlbrigge RC, Casasnovas JM, Aiuti A, Springer TA. A highly efficacious lymphocyte chemoattractant, stromal cell–derived factor 1 (SDF-1) J Exp Med. 1996;184:1101–1109. doi: 10.1084/jem.184.3.1101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Tashiro K, Tada H, Heilker R, Shirozu M, Nakano T, Honjo T. Signal sequence trap: a cloning strategy for secreted proteins and type I membrane proteins. Science. 1993;261:600–603. doi: 10.1126/science.8342023. [DOI] [PubMed] [Google Scholar]
- 28.Oberlin E, Amara A, Bachelerie F, Bessia C, Virelizier JL, Arenzana-Seisdedos F, Schwartz O, Heard JM, Clark-Lewis I, Legler DF, et al. The CXC chemokine SDF-1 is the ligand for LESTR/fusin and prevents infection by T-cell-line–adapted HIV-1. Nature. 1996;382:833–835. doi: 10.1038/382833a0. [DOI] [PubMed] [Google Scholar]
- 29.Bleul CC, Farzan M, Choe H, Parolin C, Clark-Lewis I, Sodroski J, Springer TA. The lymphocyte chemoattractant SDF-1 is a ligand for LESTR/fusin and blocks HIV-1 entry. Nature. 1996;382:829–833. doi: 10.1038/382829a0. [DOI] [PubMed] [Google Scholar]
- 30.Baggiolini M, Dewald B, Moser B. Interleukin-8 and related chemotactic cytokines–CXC and CC chemokines. Adv Immunol. 1994;55:97–179. [PubMed] [Google Scholar]
- 31.Nagasawa T, Kikutani H, Kishimoto T. Molecular cloning and structure of a pre–B-cell growth-stimulating factor. Proc Natl Acad Sci USA. 1994;91:2305–2309. doi: 10.1073/pnas.91.6.2305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Aiuti A, Webb IJ, Bleul C, Springer T, Gutierrez-Ramos JC. The chemokine SDF-1 is a chemoattractant for human CD34+ hematopoietic progenitor cells and provides a new mechanism to explain the mobilization of CD34+progenitors to peripheral blood. J Exp Med. 1997;185:111–120. doi: 10.1084/jem.185.1.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Sneath, P.H.A., and R.R. Sokal. 1973. Numerical Taxonomy. Freeman, New York.