

Abstract
Migration of mature B lymphocytes within secondary lymphoid organs and recirculation between these sites are thought to allow B cells to obtain T cell help, to undergo somatic hypermutation, to differentiate into effector cells, and to home to sites of antibody production. The mechanisms that direct migration of B lymphocytes are unknown, but there is evidence that G protein–coupled receptors, and possibly chemokine receptors, may be involved. Stromal cell– derived factor (SDF)-1α is a CXC chemokine previously characterized as an efficacious chemoattractant for T lymphocytes and monocytes in peripheral blood. Here we show with purified tonsillar B cells that SDF-1α also attracts naive and memory, but not germinal center (GC) B lymphocytes. Furthermore, GC B cells could be converted to respond to SDF-1α by in vitro differentiation into memory B lymphocytes. Conversely, the migratory response in naive and memory B cells was significantly reduced after B cell receptor engagement and CD40 signaling. The receptor for SDF-1, CXC chemokine receptor 4 (CXCR4), was found to be expressed on responsive as well as unresponsive B cell subsets, but was more rapidly downregulated on responsive cells by ligand. Finally, messenger RNA for SDF-1 was detected by in situ hybridization in a layer of cells surrounding the GC. These findings show that responsiveness to the chemoattractant SDF-1α is regulated during B lymphocyte activation, and correlates with positioning of B lymphocytes within a secondary lymphoid organ.
Mature B lymphocytes are known to localize to distinct microenvironments within secondary lymphoid organs such as the spleen, peripheral and mesenteric lymph nodes, and Peyer's patches in the gut. In the spleen, these microenvironments are located in the white pulp and support antigen- and costimulus-dependent proliferation in the T cell zone (periarteriolar lymphocyte sheath, PALS) and affinity maturation in the germinal center (GC)1 within the B cell area or follicle. Corresponding compartments can be found in the other secondary lymphoid organs. Early studies in the rat have shown that recirculating B cells collected from the thoracic duct home to the follicular mantle zone, i.e., the B cell area surrounding GCs, but not to the GC itself (1, 2). A large body of work on the generation of T cell–dependent antibody responses suggests that B lymphocytes after encounter of antigen and B cell receptor (BCR) engagement are initially activated by cognate interaction with T lymphocytes in the T cell zone (3–5). B cells then move to the follicle where they form germinal centers in which they undergo somatic hypermutation and affinity maturation. After the GC reaction wanes, some memory B cells are thought to remain in close proximity to follicular dendritic cells within the B cell area, whereas others recirculate or migrate together with antibody-secreting cells into the bone marrow (6).
The regulated movement of B lymphocytes into supportive niches is thought to prevent autoreactivity. Goodnow and coworkers have demonstrated that in the presence of a polyclonal B cell repertoire, a process termed follicular exclusion hinders autoreactive B cells from entering the B cell zone, which is a prerequisite for their survival in the absence of T cell help (7–9). Follicular exclusion has been proposed to be due to subtle differences between autoreactive and nonautoreactive B cells in responsiveness to positioning cues and is thought to be a checkpoint for censoring autoreactive B cells from the preimmune repertoire (9). This concept has recently been challenged by studies showing that BCR engagement and not the presence or absence of a polyclonal B cell repertoire is the key factor that leads to arrest of B cells in the outer periarteriolar lymphoid sheath (10, 11).
The molecular mechanisms that govern localization of B cells within secondary lymphoid organs and between these sites are unknown, but some studies suggest the involvement of seven transmembrane receptors and signaling through pertussis-toxin–sensitive G proteins. Goodnow's group has shown in transfer experiments that pretreatment with pertussis-toxin inhibits the migration of B and T cells into the splenic white pulp indicating that a G protein– coupled receptor, and perhaps one that binds a chemotactic factor, may be necessary for attraction of lymphocytes into the white pulp (12). A recent study provides evidence for a role of the putative chemokine receptor Burkitt's lymphoma receptor 1 (BLR1) in B cell migration. The phenotype of BLR1-deficient mice suggests the importance of this receptor in the localization of B lymphocytes within follicles in the spleen (13).
The role of chemotactic factors in directing granulocytes, macrophages, and T lymphocytes from the bloodstream into sites of inflammation has been extensively investigated (14, 15), but very little is known about B lymphocyte chemoattractants and their receptors. To characterize chemoattractant-induced B cell migration, we carried out chemotaxis assays with stromal cell–derived factor (SDF)-1α, which we have shown to be an efficacious chemoattractant for monocytes and lymphocytes, including the CD4+ and CD8+ T lymphocyte subsets, and the CD45RA+ naive and CD45RO+ memory T lymphocyte subsets in peripheral blood (16). SDF-1 is unique among chemotactic cytokines. Formally, SDF-1 belongs to the CXC subfamily of a family of small, 60–80 amino acid chemotactic cytokines termed chemokines. All other human CXC chemokine genes cluster on chromosome 4; the sdf-1 gene is located on chromosome 10 (17). SDF-1 is with 99% identity between mouse and human the most highly conserved cytokine described to date (16, 17). Mice genetically deficient in SDF-1 lack B lymphocytes, lack myelopoiesis in the bone marrow, and show defects in heart development (18). Such a severe phenotype has not been described for any other chemokine and demonstrates wide importance for SDF-1 in both hematopoietic and nonhematopoietic organs. The defect in B cell development is in agreement with the finding that SDF-1 functions as a pre–B cell growth-stimulating factor (19).
It has recently been shown that SDF-1α binds the CXC chemokine receptor 4 (CXCR4; references 20, 21). CXCR4, also known as LESTR/fusin, facilitates entry of T cell line– tropic strains of HIV-1 into CD4-expressing cells (22). By binding to CXCR4, SDF-1α blocks infection by these strains of HIV-1 (20, 21). Expression of CXCR4 messenger RNA (mRNA) has been detected in a wide variety of tissues (23–25) and we have found expression of CXCR4 on T and B lymphocytes on the protein level (26).
B lymphocytes differ from T lymphocytes in that they can undergo a second round of antigen receptor editing. During development, both T and B lymphocytes go through VDJ recombination in a primary lymphoid organ. However, after encounter of antigen, B lymphocytes undergo somatic hypermutation, and also further recombination, in a specialized microenvironment in secondary lymphoid organs, the GC. The surface phenotype of tonsillar B cells are characteristic for the localization in the different compartments of the human tonsil and have been well characterized. Here, we investigate the responsiveness to SDF-1α of tonsillar B lymphocytes with distinct surface phenotypes corresponding to different microenvironments within this secondary lymphoid organ.
Materials and Methods
Cytokines, Chemokines, and Antibodies.
IL-2 was a kind gift of Dr. J. Ritz (Dana Farber Cancer Institute, Boston, MA) and IL-10 was purchased from Genzyme (Cambridge, MA). Synthetic, human SDF-1α(1–67) (20) was a gift of Dr. I. Clark-Lewis (University of British Columbia, Vancouver, Canada). IL-8, macrophage inflammatory protein (MIP)-1α, MIP-1β, monocyte chemoattractant (MCP)-1, and regulated on activation normal T cell expressed and secreted (RANTES) were obtained from Genzyme, monokine induced by IFN-γ (Mig) from PharMingen (San Diego, CA), interferon-inducible protein 10 (IP10) from R&D Sys. Inc. (Minneapolis, MN), and lymphotactin from Peprotec (Rocky Hill, NJ). The following mAbs and polyclonal antibodies were used for staining of surface antigens: anti-kappa-FITC was obtained from Dako (Carpinteria, CA); anti–intercellular adhesion molecule (ICAM)-1–PE, and anti-CD20-FITC and -PerCP, anti-CD44-FITC, anti-CD56-FITC, anti-CD62 ligand (L)-FITC, anti-CD3-PerCP, anti-CD14-PE, and anti-CD38-PE from Becton Dickinson (San Jose, CA); anti-CD38-TC from CALTAG Labs. (Burlingame, CA); anti–lymphocyte function-associated antigen (LFA)-3–FITC (CD58) from Amac (Westbrook, ME); anti-CXCR4-PE (12G5) from PharMingen; anti-B7.1 (CD80) was obtained from Repligen (Cambridge, MA) and conjugated with FITC in our laboratory. The CCR3-specific mAb 7B11 (27) was a gift from Dr. C.R. Mackay (LeukoSite Inc., Cambridge, MA), and the mAb 12G5 against CXCR4 (28) was supplied by Dr. J. Hoxie (University of Pennsylvania, Philadelphia, PA). Unconjugated mAbs directed against CD38, CD44, kappa, and lambda light chain were obtained from Immunotech (Westbrook, ME), and mAbs against CD14 and CD3 from Coulter Corp. (Miami, FL). An mAb against CD40 was purchased from the American Type Culture Collection (Rockville, MD). The annexin V apoptosis detection assay was carried out according to the manufacturer's recommendations (R&D Sys. Inc.).
B Lymphocyte Purification.
Fresh tonsils were obtained from the Massachusetts Eye and Ear Infirmary (Boston, MA). Tonsils were mechanically homogenized. Tonsillar B lymphocytes were obtained from the single cell suspension by rosetting with sheep red blood cells (BioWhittaker, Walkersville, MD). The purity of tonsillar B cells obtained with this method was routinely >96% as estimated by FACScan® analysis (Becton Dickinson, San Jose, CA) using mAb to CD20, CD19, CD3, CD4, CD8, and CD14. Subsets of tonsillar B cells were purified by negative selection using a magnetic cell separation protocol (MACS; Miltenyi Biotec, Sunnyvale, CA). In brief, tonsillar B cells were incubated with mAbs specific for CD3, CD14, and either CD38 (for naive and memory B cell preparation), or CD44 (for GC B cell preparation), and subsequently with magnetic beads coated with anti–mouse IgG antibodies. Unstained cells were separated on a MACS column applying a magnetic field. The purity of CD38+CD44− GC B cells and CD38−CD44+ naive and memory B cells was routinely >97% as estimated by FACScan® analysis. Three-color flow cytometry was carried out as previously described (26). For chemokine receptor downmodulation, tonsillar B cells were incubated for 30 min at 37°C or 4°C in RPMI-1640 containing 0.25% human serum albumin in the presence or absence of 2 μg/ml SDF-1α, washed twice, and stained using the appropriate mAbs.
Chemotaxis Assays.
Chemotaxis assays were carried out as described (16) with the exception that tonsillar B lymphocytes were migrated for a period of 4 h and B cell subsets for a period of 3 h. In brief, 5 × 105 B cells in 100 μl RPMI-1640 containing 0.25% human serum albumin were transmigrated through 5-μm pore size bare filter Transwell inserts (Costar, Cambridge, MA). Migrated cells were counted by FACS® analysis scatter gating on lymphocytes. For antibody inhibition, cells were incubated for 15 min with differing concentrations of mAb to CXCR4 (12G5) or CCR3 (7B11) before addition to the top chamber of the chemotaxis assay. Chemotaxis was then carried out in the presence of mAb to an optimal concentration of 1.5 μg/ml SDF-1α. For pertussis toxin treatment, CD38− naive and memory B lymphocytes were incubated for 2 h at 37°C with 200 ng/ml pertussis toxin (GIBCO BRL, Gaithersburg, MD), washed twice, and subsequently added to the top chamber of the chemotaxis assay. Results are shown as percentage of total input or specific migration after subtraction of background migration to control medium.
Actin polymerization.
Actin polymerization was tested as described (16, 29). In brief, human B cell subsets (1.25 × 106/ml) were incubated in L15 medium at 37°C with or without SDF-1α at 1.5 μg/ml. At the indicated time points, cells (400 μl) were added to 100 μl of a solution containing 4 × 10−7 M FITC-labeled phalloidin, 0.5 mg/ml l-α-lysophosphatidylcholine (both Sigma Chemical Co., Milwaukee, WI), and 18% formaldehyde in PBS. The fixed cells were subjected to fluorescence flow cytometry and the mean relative fluorescence of each sample determined. All data points are plotted relative to the mean relative fluorescence of the sample before addition of chemoattractant.
In Vitro Generation of Memory B Cells.
Purified GC B cells were differentiated into memory type B cells in vitro as described (30). In brief, irradiated CD40L-transfected NIH3T3 cells (transfected [t]-CD40L; reference 31) were plated onto 24-well plates at 105 cells per well and adhered overnight. GC B cells were plated on t-CD40L in Iscove's modified Dulbecco's medium supplemented with 2% human AB serum, 1% human serum albumin, 50 μg/ml transferrin, 5 μg/ml insulin, and 15 μg/ml gentamicin (B cell medium) containing IL-2 (10 U/ml) and IL-10 (10 ng/ml). B cells were cultured at 37°C in 5% CO2 for 3 d followed by 4 d of reculture on freshly prepared t-CD40L cells in the presence of IL-2 and IL-10.
Short-term BCR Cross-linking.
BCR and CD40 on CD38− naive and memory B cells were cross-linked for 2 h on FcγRII (CD32) transfectants. Irradiated CD32-transfected NIH3T3 cells (t-CD32, a gift of Dr. J. Fingeroff, Dana-Farber Cancer Institute, Boston MA) were plated onto 24-well plates at 1–1.5 × 105 cells/well and adhered overnight. mAbs to human kappa and lambda light chain (2 μg/ml; both IgG1) with or without a mAb to CD40 (10 μg/ml) were preincubated on t-CD32 cells in B cell medium for 15 min before purified CD38− naive and memory B cells were added at 106 cells/ml/well. An irrelevant mAb (IgG1) was used as control. BCR/CD40 cross-linking was carried out for 2 h at 37°C. Treated cells were assayed in the chemotaxis assay to an optimal concentration of SDF-1α of 1.5 μg/ml.
In Situ Hybridization.
Digoxigenin (DIG)-labeled SDF-1 RNA probes were prepared by cloning a reverse transcriptase PCR product from total human tonsillar RNA (5′-primer: GCCATGAACGCCAAGGTCGT and 3′-primer: CGGGTCAATGCACACTTGTC) into pCR-II (Invitrogen, San Diego, CA). This PCR product contains sequence that is common to both SDF-1α and SDF-1β. Two clones carrying the insert in either orientation were identified and DIG-labeled sense and antisense RNA probes were generated after linearization with HindIII using T7 RNA polymerase according to the manufacturer's recommendations (Boehringer Mannheim GmbH, Mannheim, Germany). Control RNA/DNA hybridization on nitrocellulose filters using the DIG nucleic acid detection kit (Boehringer Mannheim GmbH) showed equal incorporation of DIG label in the two probes.
In situ hybridization was carried out on 10-μm frozen sections of human tonsil as described (32). In brief, freshly prepared sections were delipidized in chloroform for 5 min and fixed in PBS containing 4% paraformaldehyde. After fixation, sections were washed briefly once in PBS and twice in 2× standard saline citrate (SSC), and prehybridized with hybridization buffer containing 4× SSC, 10% dextran sulfate, 1× Denhardt's solution, 2 mM EDTA, 50% deionized formamide, and 500 μg/ml herring sperm DNA for 1 h at 37°C. Sections were subsequently covered with hybridization buffer containing 200 ng/ml DIG-labeled sense or antisense RNA probe at 37°C for 16 h. After washing the slides at 37°C with 2× SSC for 5 min and 60% formamide, 0.2× SSC for 15 min and subsequently at room temperature with 2× SSC for 10 min sections were blocked with a saturated solution of blocking reagent (Boehringer Mannheim GmbH) in 100 mM Tris-HCl (pH 7.5) and 150 mM NaCl for 30 min. Sections were then incubated with a 1:200 dilution of alkaline phosphatase–conjugated anti-DIG antibody in the saturated blocking solution for 2 h. After washing with 100 mM Tris-HCl (pH 7.5) and 150 mM NaCl for 20 min, sections were developed over 16 h using nitroblue tetrazolium salt/5-bromo-4-chloro-3-indolyl phosphate (NBT/BCIP) according to the manufacturer's recommendations (Boehringer Mannheim GmbH).
Results
Chemotaxis in B Lymphocytes.
To characterize chemotactic responses in B lymphocytes we used purified human tonsillar B cells in a chemotaxis assay with bare polycarbonate filters with 5-μm pores as part of a Transwell insert (16, 33). In contrast to the Boyden chamber microchemotaxis assay, this assay allows the determination of the absolute number of responsive cells and the collection and characterization of transmigrated cells. Tonsillar B lymphocytes were chosen because they comprise subpopulations in multiple states of activation and differentiation that have been well characterized according to their surface antigen expression and localization in tonsil (34–37). Using the bare filter chemotaxis assay, we detected specific migration of a small fraction of tonsillar B cells towards SDF-1α, which peaked at a concentration of 1.5 μg/ml of the chemokine and showed the typical biphasic curve that is characteristic for chemoattractant induced migration (Fig. 1 A, top). The same concentration was found to be optimal for T lymphocyte chemotaxis (16). Depending on the donor, the percentage of responsive cells varied in nine independent experiments between 6 and 20% of total input (data not shown). Chemotaxis of tonsillar B cells was partially inhibited by preincubation of input cells with the mAb 12G5 (28) directed against the receptor for SDF-1, CXCR4 (Fig. 1 A, bottom). Preincubation with a control mAb directed against CCR3; reference 27) was without any effect. Since SDF-1α–stimulated Ca2+ flux in CXCR4 transfectants is only partially inhibited by mAb 12G5 (26) only partial inhibition of B lymphocyte chemotaxis to SDF-1α would be expected. It also is possible that SDF-1α may use additional, as yet unidentified, receptors on B lymphocytes.
Figure 1.
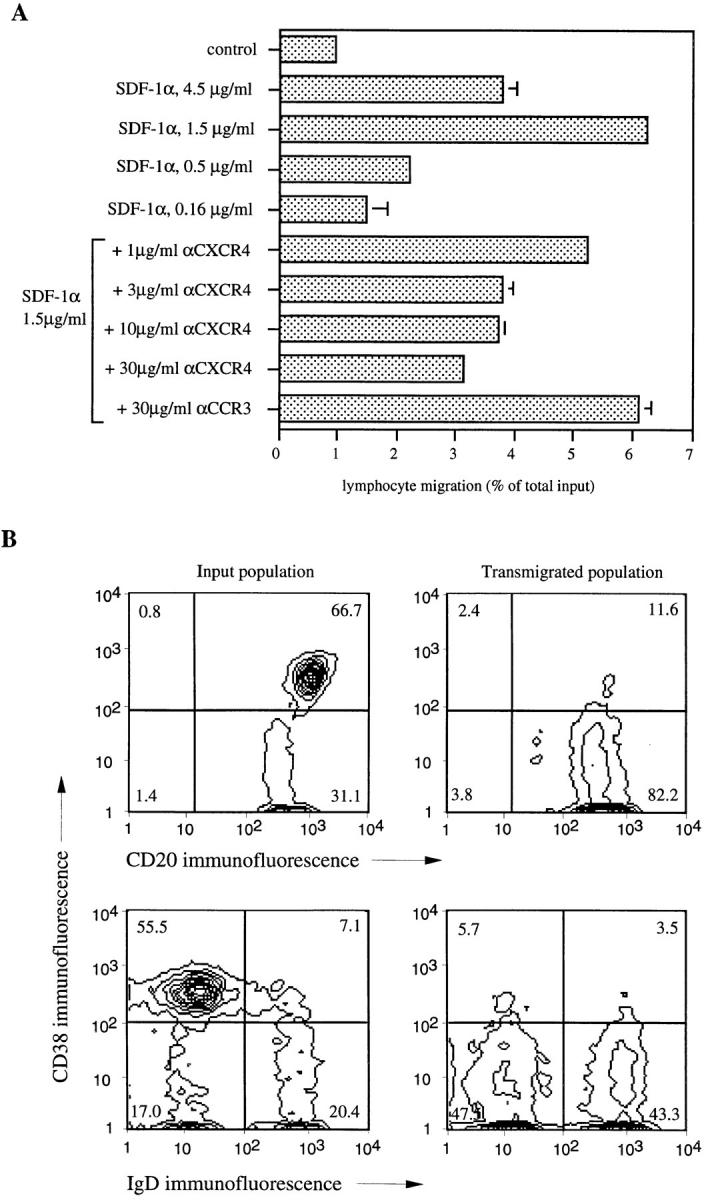
SDF-1α induces chemotaxis in CD38− naive and memory B lymphocytes, but not GC B cells from human tonsil. (A) Human tonsillar B lymphocytes were assayed in the bare filter chemotaxis assay with polycarbonate filters as part of a Transwell insert for migration to different concentrations of SDF-1α. Control shows migration to buffer alone. For antibody inhibition, human tonsillar B lymphocytes were preincubated for 15 min at the given concentrations of mAbs against the chemokine receptors CXCR4 (12G5 mAb) and CCR3 (7B11 mAb), respectively, before addition to the chemotaxis assay. Results are indicated as percent of total input, and error bars show the range of duplicates. The results are representative of four independent experiments. (B) Human tonsillar B lymphocytes were subjected to chemotaxis, and input and transmigrated populations were phenotyped by flow cytometry. Percentages of cells in the respective quadrants are indicated.
The CXC chemokines IL-8, Mig, and IP10, the CC chemokines MIP-1α, MIP-1β, monocyte chemoattractant protein 1, and regulated on activation normal T cell expressed and secreted (RANTES), the C chemokine lymphotactin, and the chemoattractant C5a were also tested for their ability to induce chemotaxis in tonsillar B lymphocytes over a 1,000-fold concentration range in the same assay. None of the tested chemokines induced significant migration in at least two independent experiments. Although a low amount of activity was seen with Mig and IP10 in some experiments, this was not a consistent finding (data not shown).
Characterization of SDF-1α Responsive B Lymphocyte Subsets.
We collected and phenotyped transmigrated tonsillar B cells. Three main subpopulations have been described (3, 34–39). GC B cells express CD38, high levels of CD20, and little to no CD44 and surface immunoglobulin. B cells outside the GC are negative for CD38, and express CD44 and intermediate levels of CD20. B cells outside the GC can be divided into CD38−CD44+IgD+ naive and CD38−CD44+IgD− memory B lymphocytes. The input population of purified tonsillar B cells contained about two-fold more CD38+ GC B cells than CD38− cells (Fig. 1 B, upper left). By contrast, almost all transmigrated cells were negative for CD38 (Fig. 1 B, upper right). Among transmigrated B lymphocytes, half were CD38−IgD+ naive and the other half were CD38−IgD− memory B cells (Fig. 1 B, lower right). Both subpopulations were present in the input population in a similar 1:1 ratio (Fig. 1 B, lower left). CD38+ GC B cells were consistently unresponsive in 11 independent experiments.
GC B cells are large, blast-like lymphocytes that are thought to undergo somatic hypermutation within the specialized microenvironment of the GC. It is unlikely, though, that the unresponsiveness to SDF-1α is due to the increased size of GC B lymphocytes, since phytohemagglutinin-stimulated T lymphocyte blasts that are even larger in size than GC B cells as estimated by forward light scatter readily respond to SDF-1α in the same assay (26). It has been shown that isolated GC B cells rapidly undergo apoptosis (40). To monitor apoptosis among input tonsillar B lymphocytes, we tested input cells at different time points over the 4-h duration of the chemotaxis assay for binding of annexin V. Loss of membrane phospholipid asymmetry, which leads to surface binding of annexin V, is thought to be an early marker of programmed cell death (41). We found that after 2 h, ∼20% and after 4 h, ∼30% of input cells bound annexin V, indicating that the majority of input GC cells remained intact during the course of the assay.
To confirm the unresponsiveness of GC B lymphocytes to SDF-1α we carried out chemotaxis assays with isolated B cell subsets. Purified CD38+ GC B cells showed little, if any, specific migration above background (Fig. 2 A), whereas 50–80% of isolated CD38− naive and memory B lymphocytes migrated to an optimal concentration of SDF-1α of 1.5 μg/ml in the same assay (Fig. 2 A). Specific migration of these cells was completely abolished after pretreatment with pertussis toxin suggesting that the chemotactic signal was dependent on a Gαi-coupled receptor (Fig. 2 B).
Figure 2.


SDF-1α efficiently attracts purified CD38− naive and memory B lymphocytes by a pertussis toxin–sensitive mechanism. (A) B lymphocyte subsets were isolated by negative selection with an mAb specific for CD38 (to obtain naive and memory B cells) or CD44 (to obtain GC B cells) and tested in the chemotaxis assay to an optimal concentration of SDF-1α of 1.5 μg/ml. Results are shown as specific migration, i.e., migration above background migration to medium alone, and represent mean and standard deviation of three independent experiments. (B) Isolated CD38− naive and memory B lymphocytes were transmigrated to an optimal concentration of SDF-1α after pretreatment with or without pertussis toxin (PT) over 2 h at 37°C. Control shows migration of sham treated cells to buffer alone. Columns and error bars show the mean and standard deviation of three independent experiments.
As a second, independent assay that determines responsiveness to SDF-1α we carried out actin polymerization experiments with CD38+ GC B cells versus CD38− B cells. In this assay, changes in intracellular filamentous actin (F-actin) were measured after exposure to SDF-1α over time (29). There was a significant, transient increase in intracellular F-actin in purified CD38− naive and memory B cells within 15 s after addition of SDF-1α, whereas F-actin in purified CD38+ GC B cells showed little change compared to sham-treated cells (Fig. 3). The observed rapid increase in filamentous actin is characteristic for chemokine induced actin polymerization and differs from stimulation with phorbol esters and calcium ionophores (29, 42, 43). The presented experiments suggest that B lymphocytes participating in the GC reaction are unresponsive to chemotactic stimulation by SDF-1α.
Figure 3.

Naive and memory, but not GC B lymphocytes respond to SDF-1α by reorganization of the actin skeleton. Intracellular F-actin was measured using FITC-labeled phalloidin in purified CD38− naive and memory B cells (boxes) and CD38+ GC B cells (circles) after addition of SDF-1α at time 0 at a concentration of 1.5 μg/ml or in untreated CD38− naive and memory B cells (diamonds). Results are shown as percent of intracellular F-actin relative to the value before addition of chemokine, and are the mean and standard deviation of three independent experiments.
Differentiation State and Chemotaxis.
The finding that the differentiation state of a B lymphocyte correlates with its responsiveness to chemokine prompted us to test if in vitro differentiation of isolated cells would restore migratory responses in the same cells. To this end we purified GC B cells and differentiated them into memory type B lymphocytes by culture on NIH3T3 cells expressing CD40L over 7 d in the presence of IL-2 and IL-10 (30). After 7 d, the majority of cultured cells had acquired the CD38−CD20intermediateCD44+IgD− phenotype that is characteristic for memory B lymphocytes in vivo (Fig. 4 A). L-selectin was induced and expression of intercellular adhesion molecule 1, lymphocyte function-associated antigen 3, and B7.1 was increased on cultured GC B cells when compared to GC B cells before culture (Fig. 4 B). These surface antigens are known to participate in lymphocyte homing and lymphocyte–lymphocyte interaction (44, 45). In parallel, specific migration to SDF-1α of these cells increased on average 27-fold during in vitro differentiation (average of three independent experiments; Fig. 4 C). The percentage of responsive cells among in vitro generated memory B cells was consistently lower than in experiments using isolated CD38− naive and memory B lymphocytes (Fig. 2 A). This may suggest that additional signals other than CD40L, IL-2, and IL-10 are necessary to fully reconstitute migratory properties in memory B lymphocytes.
Figure 4.


In vitro differentiation of GC B lymphocytes into memory B cells restores responsiveness to SDF-1α. (A) Two-color flow cytometry of isolated GC B lymphocytes (left) and in vitro generated memory B cells (right). GC B lymphocytes were cultured on CD40L expressing NIH3T3 cells in the presence of IL-2 and IL-10 for 7 d. Percentages of cells in the respective quadrants are indicated. (B) Flow cytometric analysis of GC and in vitro–generated memory B lymphocytes for homing receptors, adhesion, and costimulatory molecules. Histograms show the number of cells (y-axis) at each fluorescence intensity (log scale, x-axis). Specific mAb staining and nonbinding control IgG1 staining are shown as open and filled curves, respectively. (C) Migratory response of GC and in vitro–generated memory B lymphocytes. Results are shown as specific migration to 1.5 μg/ml of SDF-1α after subtraction of nonspecific migration to medium alone. The phenotype (Fig. 4, A and B) and migratory response (Fig. 4 C) are from a single experiment representative of three independent experiments.
BCR Cross-linking and Chemotaxis.
Since recent findings in the mouse suggest a key role for BCR engagement in the localization of B cells within the white pulp in the spleen (10, 11), we wished to determine what influence signaling through the BCR would have on the responsiveness of naive and memory B cells. Therefore, we cross-linked the BCR by incubating purified CD38− naive and memory cells for 2 h with anti-kappa and -lambda light chain mAbs in the presence of NIH3T3 cells that express FcγRII (CD32). Compared to incubation with control IgG, this treatment reduced SDF-1α–mediated chemotaxis consistently by 50% (Fig. 5 A). Treatment for longer than 2 h did not result in additional reduction in migration indicating that BCR cross-linking regulates responsiveness to SDF-1α within hours. Adding an mAb specific for CD40 to the combination of anti-kappa and -lambda light chain mAbs under the same conditions led to an additional reduction in SDF-1α–induced migration (Fig. 5 A). In contrast, the anti-CD40 mAb alone had no effect on B cell chemotaxis to SDF-1α (Fig. 5 A). To demonstrate the specificity of the inhibitory effect of BCR cross-linking on SDF-1α–induced migration, we incubated CD38− naive and memory B cells for 2 h in the presence of NIH3T3 cells expressing FcγRII with a lambda light chain–specific mAb alone. After chemotaxis to an optimal concentration of SDF-1α, input and transmigrated cells were stained for kappa light chain expression (Fig. 5 B). BCR cross-linking with an mAb specific for lambda light chain consistently led to a selective reduction in the percentage of lambda-expressing B lymphocytes in the transmigrated population compared to the input population, as shown by the increase in the percentage of transmigrated B cells that expressed kappa chain.
Figure 5.


B cell receptor engagement reduces the migratory response in CD38− naive and memory B lymphocytes. (A) CD38− naive and memory B cells were added to the chemotaxis assay after a 2-h incubation on FcγRII (CD32) expressing NIH3T3 cells in the presence of the indicated mAbs and migrated to an optimal concentration of SDF-1α of 1.5 μg/ml. mAbs to lambda and kappa light chain and the control IgG1 were used at 2 μg/ml, and the mAb to CD40 at 10 μg/ml. Results are expressed relative to the migration after incubation with control IgG1 and represent mean and standard deviation of three independent experiments. (B) CD38− naive and memory B cells were incubated on CD32-NIH3T3 cells in the presence of 2 μg/ml mAb to lambda light chain and transmigrated to an optimal concentration of SDF-1α of 1.5 μg/ml. Input (left) and transmigrated cells (right) were stained for expression of kappa light chain.
Differential Downregulation of CXCR4 by SDF-1α.
Chemokine directed migration is mediated through G protein–coupled receptors. To investigate if the unresponsiveness of GC B lymphocytes was due to a lack of expression of CXCR4, the receptor for SDF-1α, we stained tonsillar B lymphocytes containing responsive and unresponsive subsets with the CXCR4-specific mAb 12G5. We found that both subsets express high levels of the receptor (Fig. 6 A). Exposure to SDF-1α at 37°C downregulated surface expression of CXCR4. Interestingly, receptors on CD38− naive and memory B cells were more rapidly downregulated than on CD38+ GC B cells in multiple experiments, with a 73 and 54% decrease, respectively, after 30 min (Fig. 6 B). The same treatment with SDF-1α carried out at 4°C did not result in diminished binding of mAb 12G5 (not shown). This finding indicates that the reduced binding of CXCR4-specific mAb after treatment at 37°C is due to true downregulation of the receptor and not competition of the antibody and the ligand for the same epitope.
Figure 6.

CXCR4 is more rapidly downregulated on responsive B lymphocytes than on GC B cells. Phenotyping of human tonsillar B lymphocytes after treatment without (A) or with (B) 2 μg/ml SDF-1α for 30 min at 37°C. The CD38+ population corresponds to GC B cells, and the CD38− population to naive and memory B cells. A reference dashed line at 102 fluorescence units is shown for comparison between the panels.
Expression of SDF-1 in the Human Tonsil.
To locate sites of SDF-1 production within the human tonsil, we carried out in situ hybridization studies on frozen sections of human tonsil. Using a RNA probe specific for both SDF-1α and SDF-1β we detected message for SDF-1 in cells present in the connective tissue that forms the stationary scaffold of the tonsil and in nonlymphoid cells that surround GCs (Fig. 7). Expression of SDF-1 was found to be excluded from GCs and not to be associated with vascular or epithelial components in the tonsillar tissue. The staining found in the tonsil therefore suggests expression of SDF-1 in a specialized reticulum cell that lines the GC.
Figure 7.
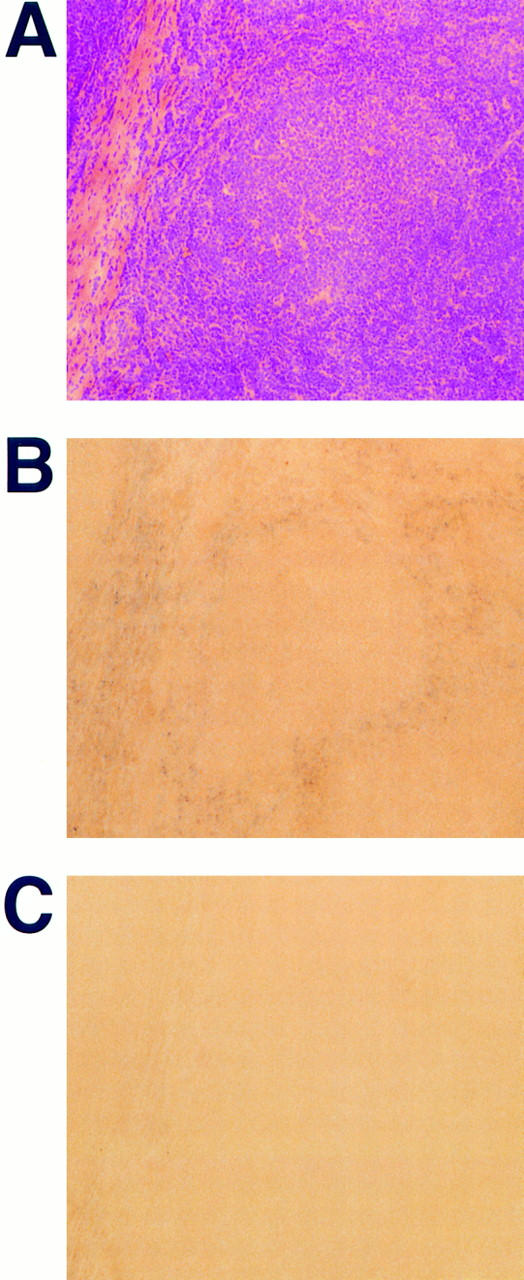
Detection of SDF-1 mRNA in human tonsil by in situ hybridization. Parallel frozen sections of human tonsil were either stained with hematoxylin and eosin (A) or hybridized with digoxigenin-labeled antisense (B) and sense (C) RNA probes specific for both SDF-1α and SDF-1β. mRNA expression was visualized using alkaline phosphatase–conjugated antidigoxigenin antibody and nitroblue tetrazolium salt/5-bromo-4-chloro-3-indolyl phosphate. Sections are shown using ×10 magnification.
Discussion
SDF-1α induces migration in naive and memory B cells, whereas GC B cells are unresponsive in assays measuring chemotaxis and actin polymerization. B lymphocytes within the GC maintain high levels of the SDF-1 receptor CXCR4. Despite the expression of the appropriate chemokine receptor on these cells, there is no ligand-induced chemotaxis. Our studies further indicate that in comparison to naive and memory B cells, on GC B cells CXCR4 is more slowly removed from the surface after addition of SDF-1α. Internalization of chemokine receptors within minutes after addition of agonist, as shown here for CXCR4 on naive and memory B lymphocytes after addition of SDF-1α, has been described for the fMLP and C5a receptors and is thought to be an important mechanism to allow the continuous sampling of ambient chemoattractant concentration that is necessary to follow a chemotactic gradient (46–48). It seems that in GC B cells, CXCR4 is less coupled to downstream regulators that induce internalization of receptors than in migratory cells; there may be overlap between these downstream regulators and those that signal chemotaxis.
The function of CXCR4 on GC B cells remains enigmatic. SDF-1 has previously been described to promote growth of pre–B cells and has therefore been termed PBSF for pre–B cell growth-stimulating factor (19). We have carried out preliminary experiments measuring apoptosis in isolated GC B lymphocytes in the presence or absence of SDF-1α and have failed to demonstrate a significant effect on inhibition of apoptosis in these cells. In addition, our in situ hybridization experiments indicate that expression of SDF-1 is excluded from GCs. Thus, we have no indication that SDF-1 serves as a growth factor or survival signal to GC B cells that express its specific receptor CXCR4. It has to be kept in mind that additional not yet identified factors may use CXCR4 on B cells within GCs.
SDF-1α induces migration in naive and memory B cells, but not in GC B cells. This observation suggests that B lymphocytes undergoing somatic hypermutation within GCs are unresponsive to cues that direct migration of naive and memory B lymphocytes. In the mouse, Butcher's group has shown that purified PNAhi GC B cells compared to B cells outside of the GC do not home to secondary lymphoid organs when transferred into syngeneic recipients (49). Although the lack of expression of the homing receptor L-selectin is thought to be a factor, other mechanisms are likely to be involved since homing to the spleen, which is L-selectin independent (50), was also impaired (49). The authors have proposed that the nonmigratory phase is a transient phenomenon that is related to a specific differentiation state within the confines of the GC microenvironment and our observation that in vitro differentiation of GC B cells towards a memory phenotype restores responsiveness to SDF-1 supports this notion.
Responsiveness of B lymphocytes to SDF-1α is regulated by differentiation state and BCR signaling. In contrast to the transition from unresponsive GC B cells to migratory effector B lymphocytes that developed over days in culture, the responsiveness to SDF-1α in CD38− naive and memory cells decreased rapidly after BCR signaling. Only hours after BCR cross-linking, naive and memory B cells migrated significantly slower than control cells, and this effect was increased by concomitant signaling through CD40. CD40 signaling by itself was ineffective. Thus, while responsiveness of naive and memory B lymphocytes to SDF-1α is reduced after BCR cross-linking, these cells are thought to migrate to the T cell areas of secondary lymphoid organs after encounter of antigen (5, 51). This finding should not be interpreted as contradictory to our results since SDF-1 is unlikely to be the only chemotactic factor acting on B lymphocytes in lymphoid organs. Our finding may therefore suggest that the reduced responsiveness to SDF-1 allows a B cell after encounter of antigen to migrate along a gradient of a different chemotactic factor leading it into the T cell area. In this context, our observation that SDF-1 mRNA is found in a specialized reticulum cell that lines GCs in the tonsil is particularly striking. The expression of mRNA for SDF-1 in nonlymphoid cells surrounding GCs correlates with the position of small recirculating naive and memory B lymphocytes in the tonsil (38). Our finding is consistent with the previous isolation of SDF-1 protein or mRNA from stromal cell lines, in this case derived from the bone marrow (16, 19, 52). Our in situ hybridization results, therefore, further support the finding that responsiveness to SDF-1 correlates with the localization of B cell subsets within the tonsil.
Much is known about the events that allow emigration of lymphocytes from the bloodstream, through high endothelial venules into secondary lymphoid organs (14, 15), whereas little is known about the subsequent steps that direct migration of B and T lymphocytes into their respective microenvironments. Our study shows that subsets of B lymphocytes within a secondary lymphoid organ respond to SDF-1α and that this responsiveness correlates with a specific localization within this lymphoid organ and is regulated by the differentiation state of the cell and by BCR engagement. Migration of pro– and pre–B cells to SDF-1 has also recently been described in the mouse (53). Our previous work demonstrated that naive and memory T lymphocytes in the peripheral blood efficiently migrate towards an SDF-1α gradient (16). Chemokines may be important to direct migration of lymphocytes into supportive microenvironments and to maintain the specialized architecture of secondary lymphoid organs. Our work suggests that SDF-1α may participate in this process, but further work will be necessary to investigate the importance of SDF-1 relative to other chemokines. The unresponsiveness of GC B cells to SDF-1 may be due to alterations in downstream signaling pathways, and thus this may generalize to other chemoattractant receptors. From work on the putative chemokine receptor BLR1, it becomes clear that distinct ligand–receptor pairs may be responsible for the positioning of B cells in the spleen and Peyer's patches versus mesenteric and peripheral lymph nodes, since GC architecture is compromised in BLR1-deficient mice only in the spleen and Peyer's patches (13). The situation is complicated by the fact that lymphocytes express multiple relevant chemokine receptors that allow detection of multiple concentration gradients, and they may influence each other within a given microenvironment. Further work will be necessary to identify other B lymphocyte chemoattractants to address these questions.
Footnotes
We are indebted to Sabine Michalak for excellent technical support. The authors thank Qing Ma for help with RNA probes and in situ hybridization and Cheryl Greene at the Massachusetts Eye and Ear Infirmary (Boston, MA) for supplying human tonsils.
C.C. Bleul was supported by a fellowship of the Deutsche Forschungsgemeinschaft, J.L. Schultze is a fellow of the Lymphoma Research Foundation of America. This work was supported by grant HL48675 of the National Institutes of Health.
Address correspondence to Timothy A. Springer, The Center for Blood Research and Harvard Medical School, Department of Pathology, 200 Longwood Ave., Boston, MA 02115. Phone: 617-278-3200; Fax: 617-278-3232; E-mail: springer@sprsgi.med.harvard.edu
Abbreviations used in this paper: BCR, B cell receptor; BLR1, Burkitt's lymphoma receptor 1; DIG, digxigenin; F-actin, filamentous actin; GC, germinal center; IP10, interferon-inducible protein 10; L, ligand; Mig, monokine-induced by IFN-γ; MIP, macrophage inflammatory protein; mRNA, messenger RNA; SDF, stromal cell–derived factor; SSC, standard saline citrate; t, transfected.
References
- 1.Nieuwenhuis P, Ford WL. Comparative migration of B- and T-lymphocytes in the rat spleen and lymph-nodes. Cell Immunol. 1976;23:254–267. doi: 10.1016/0008-8749(76)90191-x. [DOI] [PubMed] [Google Scholar]
- 2.Howard JC, Hunt SV, Gowans JL. Identification of marrow-derived small lymphocytes in the lymphoid tissue and thoracic duct lymph of normal rats. J Exp Med. 1972;135:200–219. doi: 10.1084/jem.135.2.200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.MacLennan ICM. Germinal centers. Annu Rev Immunol. 1994;12:117–139. doi: 10.1146/annurev.iy.12.040194.001001. [DOI] [PubMed] [Google Scholar]
- 4.Rajewsky K. Clonal selection and learning in the antibody system. Nature. 1996;381:751–758. doi: 10.1038/381751a0. [DOI] [PubMed] [Google Scholar]
- 5.Kelsoe G. Life and death in germinal centers (Redux) Immunity. 1996;4:107–111. doi: 10.1016/s1074-7613(00)80675-5. [DOI] [PubMed] [Google Scholar]
- 6.Paramithiotis E, Cooper MD. Memory B lymphocytes migrate to bone marrow in humans. Proc Natl Acad Sci USA. 1997;94:208–212. doi: 10.1073/pnas.94.1.208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Cyster JG, Hartley SB, Goodnow CC. Competition for follicular niches excludes self-reactive cells from the recirculating B-cell repertoire. Nature. 1994;371:389–395. doi: 10.1038/371389a0. [DOI] [PubMed] [Google Scholar]
- 8.Cyster JG, Goodnow CC. Antigen-induced exclusion from follicles and anergy are separate and complementary processes that influence peripheral B cell fate. Immunity. 1995;3:691–701. doi: 10.1016/1074-7613(95)90059-4. [DOI] [PubMed] [Google Scholar]
- 9.Goodnow CC, Cyster JG, Hartley SB, Bell SE, Cooke MP, Healy JI, Akkaraju S, Rathmell JC, Pogue SL, Shokat KP. Self-tolerance checkpoints in B lymphocyte development. Adv Immunol. 1995;59:279–368. doi: 10.1016/s0065-2776(08)60633-1. [DOI] [PubMed] [Google Scholar]
- 10.Fulcher DA, Lyons AB, Korn SL, Cook MC, Koleda C, Parish C, Fazekas de St B, Groth, Basten A. The fate of self-reactive B cells depends primarily on the degree of antigen receptor engagement and availability of T cell help. J Exp Med. 1996;183:2313–2328. doi: 10.1084/jem.183.5.2313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Cook MC, Basten A, Fazekas de St B, Groth Outer periarteriolar lymphoid sheath arrest and subsequent differentiation of both naive and tolerant immunoglobulin transgenic B cells is determined by B cell receptor occupancy. J Exp Med. 1997;186:631–643. doi: 10.1084/jem.186.5.631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Cyster JG, Goodnow CC. Pertussis toxin inhibits migration of B and T lymphocytes into splenic white pulp cords. J Exp Med. 1995;182:581–586. doi: 10.1084/jem.182.2.581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Forster R, Mattis AE, Kremmer E, Wolf E, Brem G, Lipp M. A putative chemokine receptor, BLRI, directs B cell migration to defined lymphoid organs and specific anatomic compartments of the spleen. Cell. 1996;87:1037–1047. doi: 10.1016/s0092-8674(00)81798-5. [DOI] [PubMed] [Google Scholar]
- 14.Springer TA. Traffic signals on endothelium for lymphocyte recirculation and leukocyte emigration. Annu Rev Physiol. 1995;57:827–872. doi: 10.1146/annurev.ph.57.030195.004143. [DOI] [PubMed] [Google Scholar]
- 15.Butcher EC, Picker LJ. Lymphocyte homing and homeostasis. Science. 1996;272:60–66. doi: 10.1126/science.272.5258.60. [DOI] [PubMed] [Google Scholar]
- 16.Bleul CC, Fuhlbrigge RC, Casasnovas JM, Aiuti A, Springer TA. A highly efficacious lymphocyte chemoattractant, stromal cell–derived factor 1 (SDF-1) J Exp Med. 1996;184:1101–1110. doi: 10.1084/jem.184.3.1101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Shirozu M, Nakano T, Inazawa J, Tashiro K, Tada H, Shinohara T, Honjo T. Structure and chromosomal localization of the human stromal cell-derived factor 1 (SDF1) gene. Genomics. 1995;28:495–500. doi: 10.1006/geno.1995.1180. [DOI] [PubMed] [Google Scholar]
- 18.Nagasawa T, Hirota S, Tachibana K, Takakura N, Nishikawa S-I, Kitamura Y, Yoshida N, Kikutani H, Kishimoto T. Defects of B-cell lymphopoiesis and bone marrow myelopoiesis in mice lacking the CXC chemokine PBSF/SDF-1. Nature. 1996;382:635–638. doi: 10.1038/382635a0. [DOI] [PubMed] [Google Scholar]
- 19.Nagasawa T, Kikutani H, Kishimoto T. Molecular cloning and structure of a pre–B-cell growth-stimulating factor. Proc Natl Acad Sci USA. 1994;91:2305–2309. doi: 10.1073/pnas.91.6.2305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Bleul CC, Farzan M, Choe H, Parolin C, Clark-Lewis I, Sodroski J, Springer TA. The lymphocyte chemoattractant SDF-1 is a ligand for LESTR/fusin and blocks HIV-1 entry. Nature. 1996;382:829–833. doi: 10.1038/382829a0. [DOI] [PubMed] [Google Scholar]
- 21.Oberlin E, Amara A, Bachelerie F, Bessia C, Virelizier J-L, Arenzana-Seisdedos A, Schwartz O, Heard J-M, Clark-Lewis I, Legler DF, et al. The CXC chemokine, stromal cell derived factor 1 (SDF-1), is the ligand for LESTR/fusin and prevents infection by T-cell-line–adapted HIV-1. Nature. 1996;382:833–835. doi: 10.1038/382833a0. [DOI] [PubMed] [Google Scholar]
- 22.Feng Y, Broder CC, Kennedy PE, Berger EA. HIV-1 entry cofactor: functional cDNA cloning of a seven-transmembrane, G protein–coupled receptor. Science. 1996;272:872–877. doi: 10.1126/science.272.5263.872. [DOI] [PubMed] [Google Scholar]
- 23.Federsppiel B, Melhado IG, Duncan AM, Delaney A, Schappert K, Clark-Lewis I, Jirik FR. Molecular cloning of the cDNA and chromosomal localization of the gene for a putative seven-transmembrane segment (7-TMS) receptor isolated from human spleen. Genomics. 1993;16:707–712. doi: 10.1006/geno.1993.1251. [DOI] [PubMed] [Google Scholar]
- 24.Nomura H, Nielsen BW, Matsushima K. Molecular cloning of cDNAs encoding a LD78 receptor and putative leukocyte chemotactic peptide receptors. Int Immunol. 1993;5:1239–1249. doi: 10.1093/intimm/5.10.1239. [DOI] [PubMed] [Google Scholar]
- 25.Loetscher M, Geiser T, O'Reilly T, Zwahlen R, Baggiolini M, Moser B. Cloning of a human seven-transmembrane domain receptor, LESTR, that is highly expressed in leukocytes. J Biol Chem. 1994;269:232–237. [PubMed] [Google Scholar]
- 26.Bleul CC, Wu L, Hoxie JA, Springer TA, Mackay CR. The HIV coreceptors CXCR4 and CCR5 are differentially expressed and regulated on human T lymphocytes. Proc Natl Acad Sci USA. 1997;94:1925–1930. doi: 10.1073/pnas.94.5.1925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Heath H, Qin S, Rao P, Wu L, LaRosa G, Kassam N, Ponath PD, Mackay CR. Chemokine receptor usage by human eosinophils. The importance of CCR3 demonstrated using an antagonistic monoclonal antibody. J Clin Invest. 1997;99:178–184. doi: 10.1172/JCI119145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Endres MJ, Clapham PR, Marsh M, Ahuja M, Davis-Turner J, McKnight A, Thomas JF, Stoebenau-Haggarty B, Choe S, Vance PJ, et al. CD4-independent infection by HIV-2 is mediated by fusin/CXCR4. Cell. 1996;87:745–756. doi: 10.1016/s0092-8674(00)81393-8. [DOI] [PubMed] [Google Scholar]
- 29.Howard TH, Meyer WH. Chemotactic peptide modulation of actin assembly and locomotion in neutrophils. J Cell Biol. 1984;98:1265–1271. doi: 10.1083/jcb.98.4.1265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Arpin C, Dechanet J, Van Kooten C, Merville P, Grouard G, Briere F, Banchereau J, Liu Y-J. Generation of memory B cells and plasma cells in vitro. Science. 1995;268:720–722. doi: 10.1126/science.7537388. [DOI] [PubMed] [Google Scholar]
- 31.Schultze JL, Cardoso AA, Freeman GJ, Seamon MJ, Daley J, Pinkus GS, Gribben JG, Nadler LM. Follicular lymphomas can be induced to present alloantigen efficiently: a conceptual model to improve their tumor immunogenicity . Proc Natl Acad Sci USA. 1995;92:8200–8204. doi: 10.1073/pnas.92.18.8200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Dijkman HBPM, Mentzel S, de Jong AS, Assmann KJM. RNA in situhybridization using digoxigenin-labeled cRNA probes. Biochemica. 1995;2:23–27. [Google Scholar]
- 33.Roth SJ, Carr MW, Rose SS, Springer TA. Characterization of transendothelial chemotaxis of T lymphocytes. J Immunol Methods. 1995;100:97–116. doi: 10.1016/0022-1759(95)00208-1. [DOI] [PubMed] [Google Scholar]
- 34.Lagresle C, Bella C, DeFrance T. Phenotypic and functional heterogeneity of the IgD−B cell compartment: identification of two major tonsillar B cell subsets. Int Immunol. 1993;5:1259–1268. doi: 10.1093/intimm/5.10.1259. [DOI] [PubMed] [Google Scholar]
- 35.Liu Y-J, Barthélémy C, de Bouteiller O, Arpin C, Durand I, Banchereau J. Memory B cells from human tonsils colonize mucosal epithelium and directly present antigen to T cells by rapid up-regulation of B7-1 and B7-2. Immunity. 1995;2:239–248. doi: 10.1016/1074-7613(95)90048-9. [DOI] [PubMed] [Google Scholar]
- 36.Galibert L, Burdin N, de Saint-Vis B, Garrone P, Van Kooten C, Banchereau J, Rousset F. CD40 and B cell antigen receptor dual triggering of resting B lymphocytes turns on a partial germinal center phenotype. J Exp Med. 1996;183:77–85. doi: 10.1084/jem.183.1.77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Galibert L, Burdin N, Barthelemy C, Meffre G, Durand I, Garcia E, Garrone P, Rousset F, Banchereau J, Liu Y-J. Negative selection of human germinal center B cells by prolonged BCR cross-linking. J Exp Med. 1996;183:2075–2085. doi: 10.1084/jem.183.5.2075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hardie DL, Johnson GD, Khan M, MacLennan ICM. Quantitative analysis of molecules which distinguish functional compartments within germinal centers. Eur J Immunol. 1993;23:997–1004. doi: 10.1002/eji.1830230502. [DOI] [PubMed] [Google Scholar]
- 39.Pascual V, Liu Y-J, Magalski A, de Bouteiller O, Banchereau J, Capra JD. Analysis of somatic mutation in five B cell subsets of human tonsil. J Exp Med. 1994;180:329–339. doi: 10.1084/jem.180.1.329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Liu YJ, Joshua DE, Williams GT, Smith CA, Gordon J, MacLennan ICM. Mechanism of antigen-driven selection in germinal centres. Nature. 1989;342:929–931. doi: 10.1038/342929a0. [DOI] [PubMed] [Google Scholar]
- 41.Koopman G, Reutelingsperger CPM, Kuijten GAM, Keehnen RMJ, Pals ST, van Oers MHJ. Annexin V for flow cytometric detection of phosphatidylserine expression on B cells undergoing apoptosis. Blood. 1994;5:1415–1420. [PubMed] [Google Scholar]
- 42.Howard TM, Wang D. Calcium ionophore, phorbol ester, and chemotactic peptide-induced cytoskeleton reorganization in human neutrophils. J Clin Invest. 1987;79:1359–1364. doi: 10.1172/JCI112962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Sham RL, Phatak PD, Ihne TP, Abboud CN, Packman CH. Signal pathway regulation of interleukin-8-induced actin polymerization in neutrophils. Blood. 1993;82:2546–2551. [PubMed] [Google Scholar]
- 44.Springer TA. Adhesion receptors of the immune system. Nature. 1990;346:425–433. doi: 10.1038/346425a0. [DOI] [PubMed] [Google Scholar]
- 45.June CH, Bluestone JA, Nadler LM, Thompson CB. The B7 and CD28 receptor families. Immunol Today. 1994;15:321–331. doi: 10.1016/0167-5699(94)90080-9. [DOI] [PubMed] [Google Scholar]
- 46.Van Epps DE, Simpson S, Bender JG, Chenoweth DE. Regulation of C5a and formyl peptide receptor expression on human polymorphonuclear leukocytes. J Immunol. 1990;144:1062–1068. [PubMed] [Google Scholar]
- 47.Gerard C, Gerard NP. C5a anaphylatoxin and its seven transmembrane–segment receptor. Annu Rev Immunol. 1994;12:775–808. doi: 10.1146/annurev.iy.12.040194.004015. [DOI] [PubMed] [Google Scholar]
- 48.Giannini E, Boulay F. Phosphorylation, dephosphorylation, and recycling of the C5a receptor in differentiated HL60 cells. J Immunol. 1995;154:4055–4064. [PubMed] [Google Scholar]
- 49.Reichert RA, Gallatin WM, Weissman IL, Butcher EC. Germinal center B cells lack homing receptors necessary for normal lymphocyte recirculation. J Exp Med. 1983;157:813–827. doi: 10.1084/jem.157.3.813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Arbones ML, Ord DC, Ley K, Ratech H, Maynard-Curry C, Otten G, Capon DJ, Tedder TF. Lymphocyte homing and leukocyte rolling and migration are impaired in L-selectin–deficient mice. Immunity. 1994;1:247–260. doi: 10.1016/1074-7613(94)90076-0. [DOI] [PubMed] [Google Scholar]
- 51.Liu Y-J, Arpin C. Germinal center development. Immunol Rev. 1997;156:111–126. doi: 10.1111/j.1600-065x.1997.tb00963.x. [DOI] [PubMed] [Google Scholar]
- 52.Tashiro K, Tada H, Heilker R, Shirozu M, Nakano T, Honjo T. Signal sequence trap: A cloning strategy for secreted proteins and type I membrane proteins. Science. 1993;261:600–603. doi: 10.1126/science.8342023. [DOI] [PubMed] [Google Scholar]
- 53.D'Apuzzo M, Rolink A, Loetscher M, Hoxie JA, Clark-Lewis I, Melchers F, Baggiolini M, Moser B. The chemokine SDF-1, stromal cell–derived factor 1, attracts early stage B cell precursors via the chemokine receptor. Eur J Immunol. 1997;27:1788–1793. doi: 10.1002/eji.1830270729. [DOI] [PubMed] [Google Scholar]