

Abstract
T helper cell type 1 (Th1) and 2 (Th2) are central to immune regulation. However, no stable cell surface marker capable of distinguishing and separating these two subsets of CD4+ cells has yet been found. Using differential display PCR, we have identified a gene encoding a cell membrane bound molecule, originally designated ST2L, T1, DER4, or Fit, expressed constitutively and stably on the surface of murine Th2s, but not Th1s even after stimulation with a range of immunological stimuli. Antibody against a peptide derived from ST2L strongly and stably labeled the surface of cloned Th2s but not Th1s, and Th2s but not Th1s derived from naive T cells of ovalbumin T cell receptor–α/β transgenic mice. Three-color single cell flow cytometric analysis shows that cell surface ST2L coexpressed with intracellular interleukin (IL)-4, but not with interferon (IFN)-γ. The antibody selectively lysed Th2s in vitro in a complement-dependent manner. In vivo, it enhanced Th1 responses by increasing IFN-γ production and decreasing IL-4 and IL-5 synthesis. It induced resistance to Leishmania major infection in BALB/c mice and exacerbated collagen-induced arthritis in DBA/1 mice. Thus, ST2L is a stable marker distinguishing Th2s from Th1s and is also associated with Th2 functions. Hence, it may be a target for therapeutic intervention.
CD4+ T cells can be divided into Th1 and Th2 subsets, the balance of which frequently determines the outcome of infectious and autoimmune diseases (for review see references 1 and 2). Th1s characteristically produce IFN-γ and activate macrophages to kill intracellular pathogens and can cause inflammatory diseases. Th2s produce IL-4 and IL-5 and are associated with allergic reactions involving IgE, eosinophils, and basophils (for review see reference 3). However, a stable cell surface marker distinguishing Th1s and Th2s thus far has not been found nor is the mechanism for selective cytokine induction known.
Recently, CD30 was reported to be expressed transiently and preferentially on cloned human Th2s compared with Th1s (4). Message for IL-12 receptor β2 subunit was found to be induced on Th1s by IL-12 and IFN-γ (5, 6). However, the stability and in vivo relevance of these molecules remained to be ascertained. Th1s but not Th2s, are able to bind P-selectin and L-selectin (7), but the ligands for the binding are unknown. The eotaxin receptor CCR3 was found to be selectively expressed by human Th2s (8). However, this marker is rapidly downregulated after T cell activation. A stable cell surface marker distinguishing Th1s and Th2s will not only finally settle the debate on the validity of the classification of these T cell subsets, but will greatly facilitate the investigation of Th1–Th2 interaction by selective sorting of pure population of cells for functional experimentations.
We now report the identification of a stable cell surface marker expressed on Th2s, but not Th1s. Using differential display PCR (9) comparing a panel of Th1 and Th2 clones, we identified a gene that is expressed on Th2s but not Th1s. This gene has complete homology with a gene originally designated as ST2L (10), T1 (11), DER4 (12), or Fit-1 (13). This gene (henceforward called ST2L) is transcribed in the bone marrow of adult mice and in cultured hematopoietic cells as a 5-kb messenger RNA (mRNA)1 in response to proliferation-inducing stimuli. It is expressed in major hematopoietic organs, but restricted to only a few cell types. The gene encodes a receptor-like molecule of the Ig superfamily homologous to the type I IL-1 receptor, but does not bind any of the three known IL-1 species (14). Antibody against the extracellular domain of ST2L stably labeled Th2s, but not Th1s. It lysed Th2s with complement in vitro and enhanced Th1 responses in vivo. The antibody increased resistance to Leishmania major infection in BALB/c mice and exacerbated collagen-induced arthritis in DBA/1 mice. Thus, ST2L could be an important cell surface target for the manipulation of immune responses.
Materials and Methods
Cell Lines and Tissue Culture.
T cell clones used were as follows: Dorris (specific for hen egg lysozyme, H-2k) and D10 (specific for conalbumin, H-2k) were obtained from American Type Culture Collection (Atlanta, GA). X4, X9, X12, X17, D2.2, and D2.3 were cloned and maintained as described previously (15). The X and D series of clones are all H-2d restricted and specific against group A streptococcal M protein. The cell lines were maintained by periodic antigen stimulation with the appropriate irradiated antigen-presenting cells followed by expansion in medium containing IL-2. For the derivation of naive T cell lines, CD4+ cells were purified from the spleen of OVA TCR-α/β transgenic mice (D011.10; reference 16) by negative selection as described previously (17). They were cultured with OVA peptide (OVA323-339) and irradiated BALB/c spleen cells in the presence of IL-12 plus anti–IL-4 (for Th1 line) or IL-4 (for Th2 line) for 6 d (17).
Differential Display PCR.
This was performed (9) using RNAimage kit (GenHunter Corp., Brookline, MA). Total RNA was purified from Th1 or Th2 clones and treated with DNase I (GenHunter Corp.) to remove any trace of DNA. Purified RNA (1 μg) was transcribed into cDNA using MMLV reverse transcriptase. The cDNA was amplified by AmpliTaq DNA polymerase (Perkin-Elmer Corp., Norwalk, CT) with 50 pairs of primers (GenHunter). 40 cycles of PCR reaction were performed in the presence of [35S]dATP (New England Nuclear, Boston, MA), and products separated in 6% sequencing gels under denaturing conditions. DNA fragments from Th1s and Th2s were then compared. Bands unique to Th1 or Th2 clones and longer than 150 bp were excised from the dried gel, extracted by boiling in 100 μl of double-distilled H20, and precipitated with ethanol in the presence of glycogen. DNA was further amplified with the original set of primers used to generate the particular band by the same thermal cycle conditions. PCR products were electrophoresed in 1.5% agarose ethidium bromide gel to confirm the band size before cloning into PCRII vector (Invitrogen, Carlsbad, CA) and sequenced using M13R and M13(-20)F primers.
Reverse Northern Blot.
This was used to screen and confirm the differentially expressed mRNA (18) using a ReversePrime Kit (GenHunter Corp.). Denatured PCR products (100 ng) were dot blotted onto duplicate nylon membranes with the slot-blot filtration manifolds (Hoefer, San Francisco, CA), using murine IL-5 and β-actin cDNA as controls for specificity and equal loading, respectively. Membranes were UV cross-linked and hybridized with equal amounts of Th1 or Th2 cDNA probes (106 cpm). The probes were prepared from 10–50 μg of each of Th1 or Th2 RNA by reverse transcription using 1,000 units of MMLV reverse transcriptase in the presence of α-[32P]dCTP and T20 primer. The duplicate membranes were hybridized with Th1 or Th2 probes in 6 × SSC, 5x Denhardt's solution, 0.5% SDS, and 100 μg/ml of denatured salmon sperm DNA at 60°C overnight. The membranes were then washed, exposed for 4–8 h to a x-ray film, and the hybridization patterns compared.
Northern Blot.
Total RNA was isolated from cells by RNAZolB and was separated (20 μg/lane) in 1% formaldehyde agarose gels, blotted to nylon membrane, and hybridized with the 3′ terminal cDNA fragment of ST2L, and labeled with α-[32P]dATP. The cDNA fragment was cloned from D10 cells by differential display PCR (DD-PCR) using the primer pair: AAGCTTTTTTTTTTTA and AAGCTTCAGCAGC (designated H-T11A and H-AP34, respectively; GenHunter Corp.).
Reverse Transcriptase PCR Southern Blot.
Primers for amplifying ST2L were: sense, ACTTTGTTCACCACACTCTGC; antisense, AACAGATGCCGTCTTGGAGGC (product of ∼450 bp). Primers for β-actin were: sense, GTGGGCCGCTCTAGGCACCAA; antisense, CTCTTTGATGTCACGCACGATTTC (product of ∼540 bp). PCR products were separated in 1% agarose gels. Membranes were hybridized with internal probes (ST2L, ATTGAAATGGAGCCTCTGGGTGAGGCAAGC; β-actin, CAGAGCAAGAGAGGTATCCTGACCC) labeled with γ-[32P]dATP using T4 polynucleotide kinase.
Anti-ST2L Antibody.
A rabbit was immunized with the peptide corresponding to residues 267–287 of ST2L. The peptide was selected to have no homology with IL-1 receptor, spanning the hydrophilic region and within the extracellular domain of ST2L. The rabbit was immunized subcutaneously with 100 μg of the peptide conjugated to KLH emulsified in complete Freund's adjuvant, and boosted sequentially with the peptide-KLH conjugate (100 μg) in incomplete Freund's adjuvant and in saline. Specificity of the antibody was tested by Western blot against recombinant ST2L and by ELISA with peptide-bound 96-well plates. Total IgG was purified from the immune serum and preimmunized serum by protein A column or by ammonium sulphate precipitation. Purity of the IgG was >95% on SDS-PAGE Coomassie blue staining.
Flow Cytometric Analysis.
Cells were incubated with anti-ST2L antibody (40 μg/ml) followed by FITC-conjugated donkey anti–rabbit IgG (SAPU, Paisley, Scotland). All reactions and washing were carried out at 4°C in PBS containing FCS (5%) and NaN3 (0.1%), and all antibodies were centrifuged (11,000 g for 5 min) immediately before use. For three-color staining, cells were suspended at 105–106/ml and stimulated with PMA (50 ng/ml, Calbiochem Corp., La Jolla, CA) plus ionomycin (500 ng/ml; Calbiochem Corp.) for 4 h. Brefeldin A (10 μg/ml, Sigma Chemical Co., St. Louis, MO) was added during the last 2 h. Cells were then incubated with anti-ST2L antibody or control IgG followed by biotinylated goat anti–rabbit IgG (Dako Corp.) and then developed with PerCP-streptavidin (Becton Dickinson, Mountain View, CA). The cells were then fixed with paraformaldehyde (2%), and, after washing, permeabilized at room temperature with 0.5% saponin (Sigma Chemical Co.) in PBS and FCS (5%), and stained with FITC-conjugated anti–murine IFN-γ (IgG2a; PharMingen, San Diego, CA) and PE-conjugated anti– murine IL-4 (IgG2a; PharMingen), or FITC- and PE-conjugated isotype control antibodies (Becton Dickinson). Samples were analyzed on a FACScan® flow cytometer.
Anti-ST2L and Complement-mediated Lysis of T Cells.
Cloned T cell lines (106 cells in 0.2 ml of culture medium) were incubated for 30 min on ice with various concentrations of anti-ST2L antibody or normal rabbit IgG (clarified by centrifugation). The cells were washed once and then incubated with guinea pig complement (1/10 dilution; Harlan Sera-lab, Oxon, UK) for 45 min at 37°C. Cell viability was estimated microscopically (counting >500 cells) by staining with acridine orange and ethidium bromide.
Leishmania Infection and Immunological Assays.
BALB/c mice (female, 6–8-wk-old; Harlan Olac, Bicester, UK) were infected in the right hind footpad with 5 × 105 stationary phase promastigotes of L. major (LV39) and injected intraperitoneally with 250 μg of rabbit anti-ST2L antibody or normal rabbit Ig. The antibody was administered for the next 2 d, and then every 3–4 d. Lesion development was followed by measuring the footpad swelling (difference between the infected right and uninfected left hind footpad). The maintenance of parasites, infection, and measurement of disease progression were as described previously (19). At the end of the experiment, mice were killed by cervical dislocation. Footpad was removed to assay for parasite load by limiting dilution (20). Draining lymph node cells were harvested and cultured (106/ml) in vitro with frozen and thawed parasite antigen (105–107 parasite/ml equivalent). Culture supernatant was harvested at 48 h and assayed for cytokines by ELISA using paired antibodies (PharMingen).
Induction and Treatment in Collagen-induced Arthritis in Mice.
Male DBA/1 mice (6–8-wk-old, Harlan Olac) were injected intradermally with 200 μg of bovine type II collagen (Sigma Chemical Co.) in Freund's complete adjuvant (day −21). Collagen (200 μg in PBS) was given again on day 0 by intraperitoneal injection. Daily injections (intraperitoneal, 200 μg/mouse) of anti-ST2L antibody or normal rabbit IgG were administered from day 1 for 5 d. Mice were monitored daily for signs of arthritis for which severity scores were derived as follows: 0, normal; 1, erythema; 2, erythema and swelling; 3, extension/loss of function; total score = sum of four limbs. Paw thickness was measured with a dial-calliper (Kroeplin, Munich, Germany). Quantification of arthritis was performed as previously described (21, 22). In some experiments, mice were killed 48 h after antibody treatment and spleen cells collected and cultured with graded concentrations of collagen for 48 h in vitro. Supernatants were collected for cytokine assays by ELISA using paired antibodies. T cell proliferation was analyzed by [3H]thymidine uptake.
Results
Identification of Differentially Expressed mRNA in Th1s and Th2s.
Total RNA from a panel of four different cloned Th1 and Th2 lines were compared by DD-PCR. Using 50 primer pairs, >50 distinct bands were detected (Fig. 1 a), of which 10 fragments have so far been confirmed to be differentially expressed between Th1s and Th2s using reverse Northern blot (data not shown) and Northern blot analysis. Nucleotide sequence of one band (Fig. 1 b) has complete homology with a gene originally designated as ST2L (10). Northern blot analysis, using the cloned ST2L fragment as cDNA probe, demonstrated the constitutive expression of ST2L in all four of the Th2, but not in Th1, lines (Fig. 1 c). Using the more sensitive reverse transcription PCR Southern blot, we again failed to detect ST2L message in a panel of Th1 lines. In contrast, the message was readily demonstrated in four Th2 lines.
Figure 1.

Detection of ST2L message in Th2s. RNA from a panel of four each of different Th1 (Dorris, X4, X9, and X17) and Th2 (D10, D2.2, D2.3, and X12) clones (resting cells, two wk after antigen stimulation) were extracted and compared by DD-PCR. (a) Two representative patterns comparing the PCR products from Dorris (Th1) and D10 (Th2) cells, using two different primer pairs (lanes 1 and 2, H-T11A/H-AP34; lanes 3 and 4, H-T11G/H-AP34). Arrowhead, the ST2L band. (b) Nucleotide sequence of one cDNA present in Th2s, but not in Th1s. This has complete homology with the 3′ terminus of ST2L cDNA. (Data are available from EMBL/GenBank/DDBJ under accession number D13695.) The primer (H-T11A/H-AP34) binding sites are underlined. (c) Northern blot analysis detected ST2L in D10, but not in Dorris cells. Similar results were obtained for the other three Th1 and Th2 clones (not shown). (d) Reverse transcriptase PCR Southern demonstrates the presence of ST2L message in Th2, but not in Th1, clones. c and d are representative of three experiments.
Regulation and Expression of ST2L Gene in Th1s and Th2s.
We next investigated the regulation and expression of ST2L in Ths by various immunological stimuli. In contrast to Th2s that expressed ST2L constitutively and stably, Th1s did not express ST2L even when cultured with Con A, IL-1β, IL-2, IL-4, IFN-γ, or with anti–IFN-γ or anti– IL-4 antibodies. After incubation with IL-5 or anti-CD3 antibody, Th1s expressed a weak ST2L message transiently at 3 and 6 h after activation, respectively (Fig. 2 a); but this disappeared completely by 12 h (data not shown). The expression of ST2L in Th2s was not affected by these stimuli (Fig. 2 a). ST2L is a delayed, early proliferative response gene in BALB/c-3T3 cells induced by serum factors (10– 13). We therefore examined the regulation of ST2L expression in T cells by serum. Cells were rested for 48 h in serum-free medium and then cultured in medium containing 20% FCS. The expression of ST2L in Th2s was diminished when cultured in serum-free medium, but restored soon after the addition of serum. In contrast, no ST2L message was detectable in Th1s before or 6 h after addition of serum (Fig. 2 b).
Figure 2.


(a) Regulation of ST2L gene expression in vitro analyzed by reverse transcriptase PCR Southern. Representative Th1 (X9) and Th2 (D10) clones were stimulated for 3, 6, or 12 h with: lane 1, Con A (2.5 μg/ml); lane 2, IL-1β; lane 3, IL-5; lane 4, IL-2; lane 5, IL-4; lane 6, IFN-γ (all cytokines were recombinant murine proteins obtained from Genzyme [Cambridge, MA] used at 5 ng/ml); lane 7, anti–IFN-γ (1 μg/ml); lane 8, anti–IL-4 (1 μg/ml); lane 9, medium alone; lane 10, anti-CD3 (1.5 μg/ml). Results for 12 h (not shown) were the same as for 3 h, except no message was detected in X9. D10 cells expressed ST2L constitutively and stably, whereas X9 cells failed to do so, except a weak and transient response at 3 and 6 h to IL-5 and anti-CD3, respectively. (b) Regulation of ST2L expression in T cells by serum. X9 (Th1, lane 1), X4 (Th1, lane 2), and D10 (Th2, lane 3) were cultured for 48 h in medium without serum or IL-2, and then incubated with medium containing 20% FCS. Cells were harvested at that time (0 h) or 6 h after addition of FCS, and ST2L message was analyzed by reverse transcriptase PCR Southern blot. Serum failed to induce ST2L in Th1s. Results are representative of four Th1 and Th2 clones.
Flow Cytometric Analysis of Cell Surface Expression of ST2L.
To study the location and function of the molecule, a polyclonal rabbit antibody corresponding to the hydrophilic region of the extracellular domain of ST2L was raised. Th2 clones, but not Th1 clones, were positively labeled with this antibody. The staining was clearly detected on the cell surface (Fig. 3 a). Single cell three-color flow cytometric analysis demonstrated that ST2L coexpressed with IL-4, but not with IFN-γ in cloned T cell lines (Fig. 3 b). We then determined whether this staining pattern also occurs in cells derived from naive T cells. Splenic CD4+ T cells from the OVA TCR-α/β transgenic mice (D011.10) were driven to the Th1 or the Th2 lineage in vitro in a 6-d culture and stimulated with PMA and ionomycin for 4 h (17). Staining for ST2L and intracellular IFN-γ and IL-4 demonstrated that the Th1 line expressed no ST2L or IL-4, but showed intracellular staining for IFN-γ. In contrast, the Th2 line expressed strong ST2L and IL-4 staining, but produced no detectable IFN-γ (Fig. 3 c).
Figure 3.

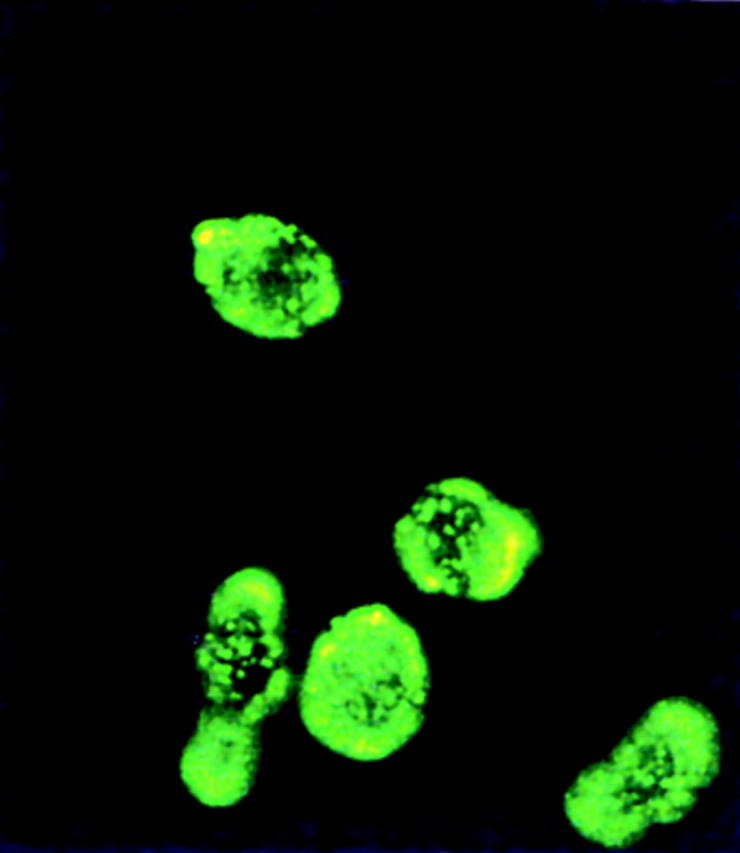


FACS® analysis of cell surface expression of ST2L. (a) Th1 (X4) and Th2 (X12) cells were stained with rabbit anti-ST2L antibody (green) or preimmune rabbit IgG (white) (40 μg/ml) and developed with an FITC-conjugated donkey anti–rabbit IgG. Similar results were obtained for three other Th1 and Th2 clones (not shown). The staining of Th2s (X12) with anti-ST2L antibody can also be visualized by fluorescent microscopy (×400). Staining can be blocked by the ST2L peptide (100 μg/ml) and no staining was detected on Th1s (data not shown). (b) Three-color flow cytometric analysis of Th1 (X4) and Th2 (X12) clones. Th2 (X12) and Th1 (X4) clones were stained for cell surface ST2L (with PerCP), followed by intracellular staining with anti–IL-4 (with PE) and anti–IFN-γ (with FITC). All cells shown in the boxes were activated with PMA/ionomycin. Cells analyzed in boxes (i), (iii), (v) and (vii) were stained with normal rabbit IgG and isotype controls for anti–IFN-γ and anti–IL-4, whereas cells in boxes (ii), (iv), (vi), and (viii) were stained with rabbit anti-ST2L, anti–IFN-γ, and anti–IL-4 antibodies. Cells in boxes (i), (ii), (v), and (vi) have been gated for positive expression of ST2L when stained with anti-ST2L antibody (R3, black) relative to negative staining with control rabbit IgG (R2, gray). Colocalization of R2 and R3 depicts negative expression of ST2L. Cytokines in the culture supernatants (48 h) of T cell clones (106/ml) stimulated with specific antigen and irradiated antigen-presenting cells were (IFN-γ versus IL-4, ng/ml): X4, 5.2 versus <0.02; X12, <0.01 versus 0.25. (c) Three-color flow cytometric analysis of Th1s and Th2s derived from naive T cells. CD4+ splenic T cells from D011.10 TCR-α/β transgenic mice were driven to Th1 or Th2 subsets in a 6-d culture. They were then stimulated with PMA/ionomycin, stained, and analyzed as in b. Cytokines in the culture supernatants (day 3) were (IFN-γ versus IL-5, ng/ml): Th1 line, 2.6 versus <0.02; Th2 line, <0.01 versus 0.52. Results are representative of five experiments.
Anti-ST2L Antibody and Complement Selectively Lysed Th2s.
We next examined the effect of anti-ST2L antibody on Th1 and Th2 clones in vitro. A panel of Th1 and Th2 clones were incubated in vitro with graded concentrations of anti-ST2L or normal rabbit IgG, and then with or without complement. In the presence of complement, anti-ST2L antibody strongly lysed the Th2 clones, but had only a modest effect on Th1 clones (Table 1).
Table 1.
Effect of Anti-ST2L Antibody on T Cells In Vitro
| Treatment of cells | Th1 (X4) percent viability | Th2 (X12) percent viability | ||
|---|---|---|---|---|
| Anti-ST2L + C′ | 85.2 ± 2.1 | 9.7 ± 0.5* | ||
| Anti-ST2L alone | 94.9 ± 3.5 | 95.2 ± 4.6 | ||
| Normal IgG + C′ | 95.9 ± 2.7 | 94.6 ± 3.3 | ||
| Normal IgG alone | 95.0 ± 2.9 | 97.2 ± 5.8 | ||
| Medium alone | 96.2 ± 3.5 | 95.8 ± 3.9 |
Th1s and Th2s were incubated with 400 μg/ml of antibody and then with or without complement (C′). The cell viability was determined microscopically. At this concentration, in the absence of complement, anti-ST2L antibody, but not normal IgG, also partially suppressed the proliferation and cytokine production by Th2s in vitro in response to specific antigen and antigen-presenting cells. The antibody did not affect the proliferation of Th1s (data not shown). Data are mean ± SEM, n = 3.
P <0.001 compared with medium control. Similar results were obtained for three other Th1 and Th2 clones.
Effect of anti-ST2L Antibody on Leishmania Infection.
We next investigated the effect of anti-ST2L antibody in vivo. BALB/c mice are genetically highly susceptible to infection by the protozoan parasite, L. major. The disease progression can be modified and resistance induced by a variety of immunological manipulations (for review see reference 23). Disease progression and resistance are associated with the preferential development of Th2s or Th1s, respectively (for review see reference 24). Fig. 4 demonstrates that BALB/c mice infected with L. major and treated with anti-ST2L antibody developed significantly smaller lesions (Fig. 4 a) and three orders of magnitude less parasite load (Fig. 4 b) compared with mice treated with control rabbit IgG. Draining lymph node cells from the anti-ST2L– treated mice produced significantly more IFN-γ, but less IL-4 and IL-5 compared with cells from control IgG-treated mice (Fig. 4 c). IL-12 levels were not affected.
Figure 4.

Effect of anti-ST2L antibody on L. major infection. BALB/c mice were infected in the footpad with the parasite and injected intraperitoneally with anti-ST2L antibody or normal rabbit IgG. Antibody-treated mice developed significantly smaller lesions (a) and parasite loads (b) compared with controls, n = 10. Experiments were terminated when control mice developed necrotic lesions as required by the United Kingdom Home Office guidelines. (c) Draining lymph node cells were pooled (n = 10) and cultured with leishmanial antigen, and supernatants from triplicate cultures assayed for cytokines. There was no significant difference in the T cell proliferative response between the two groups of mice (data not shown). Results are representative of two experiments.
Effect of Anti-ST2L Antibody on Collagen-induced Arthritis.
Collagen-induced arthritis (CIA) in susceptible DBA/1 mice is a predominantly Th1-mediated disease (25–27), the immunopathogenesis of which closely resembles clinical rheumatoid arthritis (28, 29). We therefore examined whether administration of anti-ST2L antibody could modify CIA. Mice injected intradermally with type II collagen in Freund's complete adjuvant developed arthritis when challenged intraperitoneally 21 d later with collagen. Incidence and severity of disease development were markedly enhanced in mice that received daily intraperitoneal injections of anti-ST2L antibody from day 22, compared with controls that received normal IgG (Fig. 5, a–c). On stimulation with collagen in vitro, spleen cells from antibody-treated mice also produced significantly stronger proliferative response and more IFN-γ and IL-6 compared with cells from control mice (Fig. 5 d).
Figure 5.

Anti-ST2L antibody exacerbated collagen-induced arthritis in DBA/1 mice. Mice were primed (day −21) and challenged (day 0) with collagen and injected with anti-ST2L antibody or normal rabbit IgG daily from day 1. (a) Incidence rate (percent), (b) clinical score, and (c) number of arthritic paws. Data are mean ± SEM, n = 15. Mice (n = 4) were killed 48 h after antibody treatment, spleen cells were pooled and cultured with collagen (50 μg/ml) for 48– 96 h, and culture supernatants were assayed for cytokines by ELISA. *P <0.05, **P <0.01 (two sample t test). IL-4 and IL-5 were not detected (<20 pg/ml). T cell proliferation was expressed as [3H]thymidine uptake in triplicate cultures (mean cpm × 103 ± SEM): antibody treated, 52.8 ± 6.8; control, 27.8 ± 1.1 (P <0.05).
Discussion
Data presented here demonstrate that ST2L is a stable cell surface marker expressed strongly on Th2s, but not Th1s. Furthermore, antibody against ST2L selectively lysed Th2s in vitro and enhanced Th1 response in vivo. Thus, anti-ST2L antibody increased resistance to L. major infection in the highly susceptible BALB/c mice. It also exacerbated CIA in DBA/1 mice. These effects were accompanied by enhanced IFN-γ production and diminished IL-4/ IL-5 synthesis. It is likely that the antibody exerted its effect through direct depletion of Th2s in vivo and hence the regulation of Th1s by Th2s (1–3), such that the balance is shifted towards Th1s. This is consistent with the observation that treatment with anti-ST2L antibody in the L. major–infected mice did not significantly alter the production of IL-12, a cytokine principally secreted by macrophages and a major mediator of Th1 differentiation (30). Such a mechanism would also explain the immediate effect of the antibody seen in the CIA model (Fig. 5).
ST2L is not detectable on B cells, a macrophage cell line (J774), or a CD8+ cell line (CTLL) (data not shown). The developmental stage of Th2s at which ST2L expression occurs is currently unclear. However, the expression of ST2L is clearly associated with the production of IL-4 and IL-5. It has been reported that the transcription factor GATA-3 is necessary and sufficient for Th2 cytokine gene expression in CD4+ T cells (31). Furthermore, the promoter region of ST2L contains the GATA consensus binding motif (32). It is therefore possible that GATA-3 influences Th2 functions by directing the transcription of ST2L. The ontogeny of Th2s in relation to ST2L and the sequence of events leading to the induction of ST2L and Th2 cytokine production remain to be explored.
It is important to note that a soluble form (ST2, mol wt: 37-kD) without the trans-membrane domain (10–13, 33) is also transcribed from the same gene by differential splicing, determined by the use of alternate promoters (13, 32). We have found that all Th2s expressing ST2L gene also express ST2 message (data not shown). The receptor-like nature of ST2L suggests the existence of a ligand that is yet to be identified. It may be that the membrane bound form (ST2L) and the soluble form (ST2) act as regulators of Th2 functions. Furthermore, they should serve as important biologically relevant tools for investigating gene regulation for the expression of a trans-membrane versus a secreted form of a protein.
The in vivo effect of anti-ST2L antibody in mice suggests that selective modulation of ST2L expression could have considerable therapeutic potential. The close association of ST2L expression with IL-4/IL-5 production suggests that the membrane-bound ST2L molecule could be a potentially important therapeutic target for allergic responses such as asthma. ST2L is present in humans (34) and its relevance in human T cell functions is currently being addressed.
In conclusion, we have identified a stable cell surface marker distinguishing Th2s from Th1s. Furthermore, an antibody against this molecule has a range of biological effects in vivo including infection and inflammation. Thus, this marker will not only facilitate functional analysis of Th1 and Th2 interaction, it may also be a potential target for therapeutic intervention for some of the important diseases.
Footnotes
This work was supported by the Wellcome Trust, the Medical Research Council of the United Kingdom, The Joint Research Board of The Barts National Health Service Trust.
We thank Dr. Wanda Niedbala for some of the tissue culture, Dr. Alisdair Gracie for purifying the anti-ST2L antibody, Dr. Jim Cohen for use of fluorescent microscope, Dr. Nick Golding for advice on FACS® graphic, and Dr. Ken Murphy for the D011.10 transgenic mice.
Address correspondence to F.Y. Liew, Department of Immunology, University of Glasgow, Glasgow G11 6NT, UK. Phone: 44-141-211-2695; Fax: 44-141-337-3217; E-mail: f.y.liew@clinmed.gla.ac.uk
Abbreviations used in this paper: CIA, collagen-induced arthritis; DD-PCR, differential display PCR; mRNA, messenger RNA.
References
- 1.Sher A, Coffman RL. Regulation of immunity to parasites by T cells and T cell–derived cytokines. Annu Rev Immunol. 1992;10:385–409. doi: 10.1146/annurev.iy.10.040192.002125. [DOI] [PubMed] [Google Scholar]
- 2.Abbas AK, Murphy KM, Sher A. Functional diversity of helper T lymphocytes. Nature. 1996;383:787–793. doi: 10.1038/383787a0. [DOI] [PubMed] [Google Scholar]
- 3.Mosmann TR, Coffman RL. Th1 and Th2 cells: different patterns of lymphokine secretion lead to different functional properties. Annu Rev Immunol. 1989;7:145–173. doi: 10.1146/annurev.iy.07.040189.001045. [DOI] [PubMed] [Google Scholar]
- 4.Delprete G, Defcarli M, Almerigogna F, Daniel CK, Delios MM, Zancuoghi G, Vinante F, Pizzolo G, Romagnani S. Preferential expression of CD30 by human CD4+ T cells producing Th2-type cytokines. FASEB J. 1995;9:81–86. [PubMed] [Google Scholar]
- 5.Szabo SJ, Dighe AS, Gubler U, Murphy KM. Regulation of the interleukin (IL)-12R β2 subunit expression in developing T helper 1 (Th1) and Th2 cells. J Exp Med. 1997;185:817–824. doi: 10.1084/jem.185.5.817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Rogge L, Barberis-Maino L, Biffi M, Passini N, Presky DH, Gubler U, Sinigaglia F. Selective expression of an interleukin-12 receptor component by human T helper 1 cells. J Exp Med. 1997;185:825–831. doi: 10.1084/jem.185.5.825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Austrup F, Vestweber D, Borges E, Löhning M, Bräuer R, Herz U, Renz H, Hallmann R, Scheffold A, Radbruch A, Hamann A. P- and E-selectin mediate recruitment of T-helper-1 but not T-helper-2 cells into inflamed tissues. Nature. 1997;385:81–83. doi: 10.1038/385081a0. [DOI] [PubMed] [Google Scholar]
- 8.Sallusto F, Mackay CR, Lanzavecchia A. Selective expression of the eotaxin receptor CCR3 by human T helper 2 cells. Science. 1997;277:2005–2007. doi: 10.1126/science.277.5334.2005. [DOI] [PubMed] [Google Scholar]
- 9.Liang P, Pardee AB. Differential display of eukaryotic messenger RNA by means of the polymerase chain reaction. Science. 1992;257:967–971. doi: 10.1126/science.1354393. [DOI] [PubMed] [Google Scholar]
- 10.Tominaga S. A putative protein of a growth specific cDNA from BALB/c-3T3 cells is highly similar to the extracellular portion of mouse interleukin 1 receptor. FEBS Lett. 1989;258:301–304. doi: 10.1016/0014-5793(89)81679-5. [DOI] [PubMed] [Google Scholar]
- 11.Werenskiold AK, Hoffmann S, Klemenz R. Induction of a mitogen-responsive gene after expression of the Ha-ras oncogene in NIH 3T3 fibroblasts. Mol Cell Biol. 1989;9:5207–5214. doi: 10.1128/mcb.9.11.5207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lanahan A, Williams JB, Sanders LK, Nathans D. Growth factor-induced delayed early response gene. Mol Cell Biol. 1992;12:3919–3929. doi: 10.1128/mcb.12.9.3919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bergers G, Reikerstorfer A, Braselmann S, Graninger P, Busslinger M. Alternative promoter usage of the Fos-responsive gene Fit-1generates mRNA isoforms coding for either secreted or membrane-bound proteins related to the IL-1 receptor. EMBO (Eur Mol Biol Organ) J. 1994;13:1176–1188. doi: 10.1002/j.1460-2075.1994.tb06367.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Robler U, Thomassen E, Hultner L, Baier S, Danescu J, Werenskiold AK. Secreted and membrane-bound isoforms of T1, an orphan receptor related to IL-1 binding proteins are differentially expressed in vivo. . Dev Biol. 1995;168:86–97. doi: 10.1006/dbio.1995.1063. [DOI] [PubMed] [Google Scholar]
- 15.Rossiter BA, Alfonso C, Kehoe MA, Robinson JH. Processing of viable group A streptococci leads to MHC class II presentation of T cell epitopes from the major protective antigen. Eur J Immunol. 1994;24:1244–1247. doi: 10.1002/eji.1830240537. [DOI] [PubMed] [Google Scholar]
- 16.Murphy KM, Heimberger AB, Loh DY. Induction by antigen of intrathymic apoptosis of CD4+CD8+ TCR (lo) thymocytes in vivo. . Science. 1990;250:1720–1723. doi: 10.1126/science.2125367. [DOI] [PubMed] [Google Scholar]
- 17.Openshaw P, Murphy EE, Hosken NA, Maino V, Davis K, Murphy KM, O'Garra A. Heterogeneity of intracellular cytokine synthesis at the single-cell level in polarized T helper 1 and T helper 2 populations. J Exp Med. 1995;182:1357–1367. doi: 10.1084/jem.182.5.1357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Zhang H, Zhang R, Liang P. Differential screening of gene expression difference enriched by differential display. Nucleic Acids Res. 1996;24:2454–2456. doi: 10.1093/nar/24.12.2454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Howard JG, Hale C, Liew FY. Immunological regulation of experimental cutaneous leishmaniasis III. Nature and significance of specific suppression of cell-mediated immunity in mice highly susceptible to Leishmania tropica. . J Exp Med. 1980;152:594–607. doi: 10.1084/jem.152.3.594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.McSorley SJ, Xu D, Liew FY. Vaccine efficacy of Salmonellastrains expressing glycoprotein 63 with different promoters. Infect Immun. 1997;65:171–178. doi: 10.1128/iai.65.1.171-178.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Van Roon JAG, Van Roy J, GmeligMeyling FHJ, Lafeber F, Bijlsma JWJ. Prevention and reversal of cartilage degradation in rheumatoid arthritis by interleukin-10 and interleukin-4. Arthritis Rheum. 1996;39:829–835. doi: 10.1002/art.1780390516. [DOI] [PubMed] [Google Scholar]
- 22.Joosten LAB, Lubberts E, Durez P, Helsen MMA, Jacobs MJM, Goldman M, Van Den Berg WB. Role of interleukin-4 and interleukin-10 in murine collagen-induced arthritis: protective effect of interleukin-4 and interleukin-10 treatment on cartilage destruction. Arthritis Rheum. 1997;40:249–260. doi: 10.1002/art.1780400209. [DOI] [PubMed] [Google Scholar]
- 23.Liew FY, O'Donnell CA. Immunology of Leishmania. . Adv Parasitol. 1993;32:161–259. doi: 10.1016/s0065-308x(08)60208-0. [DOI] [PubMed] [Google Scholar]
- 24.Reiner SL, Locksley RM. The regulation of immunity to Leishmania major. . Annu Rev Immunol. 1995;13:151–177. doi: 10.1146/annurev.iy.13.040195.001055. [DOI] [PubMed] [Google Scholar]
- 25.Simon AK, Seipelt E, Sieper J. Divergent T-cell cytokine patterns in inflammatory arthritis. Proc Natl Acad Sci USA. 1994;91:8562–8566. doi: 10.1073/pnas.91.18.8562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Mauri C, Williams RO, Walmsley M, Feldmann M. Relationship between Th1/Th2 cytokine pattern and the arthritogenic response in collagen-induced arthritis. Eur J Immunol. 1996;26:1511–1518. doi: 10.1002/eji.1830260716. [DOI] [PubMed] [Google Scholar]
- 27.Mussener A, Litton MJ, Lindroos E, Kareskog L. Cytokine production in synovial tissue of mice with collagen-induced arthritis (CIA) Clin Exp Immunol. 1997;107:485–493. doi: 10.1046/j.1365-2249.1997.3181214.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Trentham DE, Townes AS, Kang AH. Autoimmunity to type II collagen: an experimental model of arthritis. J Exp Med. 1977;146:857–868. doi: 10.1084/jem.146.3.857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Courtenay JS, Dallam MJ, Dayan AB, Martin A, Mosedale B. Immunisation against heterologous type II collagen-induced arthritis in mice. Nature. 1980;283:666–668. doi: 10.1038/283666a0. [DOI] [PubMed] [Google Scholar]
- 30.Trinchieri G. Interleukin-12 and its role in the generation of Th1 cells. Immunol Today. 1993;14:335–338. doi: 10.1016/0167-5699(93)90230-I. [DOI] [PubMed] [Google Scholar]
- 31.Zheng WP, Flavell RA. The transcription factor GATA-3 is necessary and sufficient for Th2 cytokine gene expression in CD4 T cells. Cell. 1997;89:587–596. doi: 10.1016/s0092-8674(00)80240-8. [DOI] [PubMed] [Google Scholar]
- 32.Gachter T, Werenskiold AK, Klemenz R. Transcription of the interleukin-1 receptor–related T1 gene is initiated at different promoters in mast cells and fibroblasts. J Biol Chem. 1996;271:124–129. doi: 10.1074/jbc.271.1.124. [DOI] [PubMed] [Google Scholar]
- 33.Yanagisawa K, Takagi T, Tsukamoto T, Tetsuka T, Tominaga S. Presence of a novel primary response gene ST2L, encoding a product highly similar to the interleukin 1 receptor type 1. FEBS Lett. 1993;318:83–87. doi: 10.1016/0014-5793(93)81333-u. [DOI] [PubMed] [Google Scholar]
- 34.Tominaga S, Yokota T, Yanagisawa K, Tsukamoto T, Takagi T, Tetsuka T. Nucleotide sequence of a complementary DNA for human ST2. Biochim Biophys Acta. 1992;1171:215–218. doi: 10.1016/0167-4781(92)90125-j. [DOI] [PubMed] [Google Scholar]