

Abstract
Immunostimulatory DNA and oligodeoxynucleotides containing unmethylated CpG motifs (CpG DNA) are strongly stimulatory for B cells and antigen-presenting cells (APCs). We report here that, as manifested by CD69 and B7-2 upregulation, CpG DNA also induces partial activation of T cells, including naive-phenotype T cells, both in vivo and in vitro. Under in vitro conditions, CpG DNA caused activation of T cells in spleen cell suspensions but failed to stimulate highly purified T cells unless these cells were supplemented with APCs. Three lines of evidence suggested that APC-dependent stimulation of T cells by CpG DNA was mediated by type I interferons (IFN-I). First, T cell activation by CpG DNA was undetectable in IFN-IR−/− mice. Second, in contrast to normal T cells, the failure of purified IFN-IR−/− T cells to respond to CpG DNA could not be overcome by adding normal IFN-IR+ APCs. Third, IFN-I (but not IFN-γ) caused the same pattern of partial T cell activation as CpG DNA. Significantly, T cell activation by IFN-I was APC independent. Thus, CpG DNA appeared to stimulate T cells by inducing APCs to synthesize IFN-I, which then acted directly on T cells via IFN-IR. Functional studies suggested that activation of T cells by IFN-I was inhibitory. Thus, exposing normal (but not IFN-IR−/−) T cells to CpG DNA in vivo led to reduced T proliferative responses after TCR ligation in vitro.
Keywords: type I interferons, CpG DNA, T cell stimulation, adjuvant, antigen-presenting cell stimulation
CpG DNA (nonvertebrate DNA and synthetic oligodeoxynucleotides [ODNs]1 containing unmethylated CpG motifs) has been shown to cause strong activation of B cells, NK cells, macrophages, and dendritic cells (1–7). Although activation of NK cells is thought to be mediated by cytokines produced by activated macrophages, i.e., IL-12 or IL-18 (8), the effect of CpG DNA on both B cells and macrophages appears to be direct (3, 6). Thus, CpG DNA can stimulate highly purified normal B cells and macrophages as well as cell lines derived from these cells (3, 6, and our unpublished data). On the other hand, several recent studies have demonstrated that CpG DNA and ODNs can function as a strong adjuvant for antigen-specific T cell responses (9–14). However, the mechanisms involved in the adjuvant function of CpG DNA are largely unknown.
With one exception (13), most studies have revealed no evidence for direct T cell stimulation by CpG DNA. Nevertheless, injection of CpG DNA in mice results in production of a variety of inflammatory cytokines, including TNF-α, IL-1, IL-6, and type I (α/β) IFNs (IFN-I) (6, 8, 15–17). Of these cytokines, IFN-I has pleomorphic effects on the immune system, including activation of macrophages (18), stimulation of NK cells (19), and proliferation of memory-phenotype (CD44hi) CD8+ cells (20).
In this paper, we present evidence that the activities of CpG DNA and IFN-I are closely connected. Injecting mice with CpG DNA or CpG ODNs causes partial activation (upregulation of CD69 and B7-2) of various cell types, including B cells, APCs, and both naive- and memory-phenotype T cells. Significantly, for T cells such activation is IFN-I dependent and can be mimicked by purified IFN-I.
Materials and Methods
Mice.
C57BL/6J (B6), C57BL/6J-Rag-1tm1Mom (Rag-1−/−), C57BL/6J-bgJ/+ (beige), C57BL/6-Igh-6tm1Cgn (μMT), C3H/ HeJ, and C3H/HeOuJ mice were purchased from The Jackson Laboratory (Bar Harbor, ME). Mice lacking both class I (β2 microglobulin) and class II MHC molecules (MHC I−II− mice) were provided by Dr. C. Surh at The Scripps Research Institute (TSRI). 129/SvEvTacfBR (129) mice were purchased from Taconic Farms (Germantown, NY). 129 background mice defective in IFN-α/β receptor function (IFN-IR−/− mice) (21) were originally purchased from B&K Universal (North Humberside, UK) and were maintained and bred in the animal facility at TSRI.
Antibodies and Other Reagents.
The following mAbs were used: 3.168.8 (anti-CD8); RL172 (anti-CD4); J11d (anti–heat stable antigen); J1j.10 (anti–Thy-1.2; as described in reference 4). Conjugated antibodies (PE–anti-CD8α, PE–anti-CD8β, PE– anti-CD4, PE–anti-B220, FITC–anti-CD69, FITC–anti-B7-2, FITC–anti-CD25, FITC–anti-intercellular adhesion molecule (ICAM)-1, and FITC–anti-Ly-6C) were purchased from PharMingen (San Diego, CA). FITC–anti-Kb mAb was purchased from Caltag Labs. (Burlingame, CA). Biotin–anti-CD44 (pgp-1) mAbs were prepared by standard techniques. Cy-5–anti-CD4 (GK1.5) and Cy-5–anti-CD8 (YTS.169) were prepared using a kit from Amersham Pharmacia Biotech (Arlington Heights, IL). Streptavidin-Red 670 and Streptavidin-FITC was purchased from GIBCO BRL (Gaithersburg, MD). LPS was purchased from CalBiochem (La Jolla, CA). Recombinant mouse IFN-β and IFN-γ were purchased from Access Biomolecules (La Jolla, CA) and R&D Systems, Inc. (Minneapolis, MN), respectively.
DNA and ODNs.
DNA from the Drosophila melanogaster cell line, SC2, was prepared as previously described (4). DNA was used without denaturation. CpG (GCATGACGTTGAGCT) and ZpG (GCATGAZGTTGAGCT, Z = 5′-methyl-C) phosphorothioate ODNs were designed using published sequences (3) with modification. The ODNs were synthesized and purified using HPLC by Research Genetics, Inc. (Huntsville, AL). Residual LPS in DNA preparations was measured using a Limulus Amebocyte Lysate QCL-1000 kit from BioWhittaker (Walkersville, MD). D. melanogaster DNA preparations contained 0–10 pg of LPS/mg of DNA.
Medium.
For proliferation assays, RPMI 1640 (GIBCO BRL) was supplemented with 10% FCS (Irvine Scientific Co., Santa Ana, CA), 5% NCTC 109, 2 mM glutamine, 5 × 10−5 M 2-mercaptoethanol, and antibiotics.
Cell Purification.
T cells were depleted from spleen or LN cell suspensions by incubation with a mixture of mAbs specific for Thy1, CD4, and CD8 plus complement for 45 min at 37°C; B cells were depleted using J11d mAb plus C. CD4+ or CD8+ cells were purified by first depleting B and CD8+ or CD4+ cells by incubation with mAbs plus C followed by positive panning on plates coated with anti-CD4 or anti-CD8 mAbs. To obtain highly purified resting naive CD8+ cells, CD8+ cells purified by positive panning were first stained for CD8β and CD44 and then sorted by FACS® (Becton Dickinson, San Jose, CA). The resulting population was 99.9% pure for CD8β+CD44lo/int cells.
In Vitro Cultures.
For cell surface staining, unseparated spleen cells (2.5 × 106 cells/ml), T-enriched LN (B-depleted), B-enriched LN (T-depleted), or positively purified LN CD4+ or CD8+ cells (106 cells/ml) were cultured in a 24- or 48-well plate overnight in the presence of DNA or rIFNs. In reconstitution cultures, purified CD4+ or CD8+ cells were supplemented with Rag-1−/− spleen cells. For proliferation assays, CD8+ cells were purified from LN by panning after mAb plus C treatment. Transfected Drosophila APCs were prepared as previously described (4). For stimulating proliferation of CD8+ cells from 129 or IFN-IR knockout (KO) mice, purified anti-TCR (1 μg/ml) and anti-CD28 (10 μg/ml) (PharMingen) mAbs were used to coat 96-well plates overnight at 4°C. Cell proliferation was assessed by measuring [3H]thymidine incorporation after adding 1 μCi [3H]thymidine for 8 h to the cultures. The data shown in the figures are means of either duplicate or triplicate cultures.
In Vivo Activation of B and T Cells by DNA.
B6, 129, or IFN-IR KO mice were injected subcutaneously in the lower hind limbs with either PBS, D. melanogaster DNA (100 μg/mouse), or ODNs (50 μg/mouse) overnight. Draining (popliteal) LN and nondraining (mesenteric) LN cells were harvested 18 h later and stained.
Cell Surface Staining and Flow Cytometry.
Cultured cells were first depleted of dead cells and debris by Ficoll-gradient centrifugation (Sigma Chemical Co., St. Louis, MO). Staining for cell surface markers was done essentially as previously described (4). When staining unseparated spleen or LN cells, four-color staining was usually performed, which involved initial staining with Cy-5–anti-CD4 (or CD8), PE–anti-B220, biotin–anti-CD44, and FITC–CD69 (or B7-2, Kb, ICAM-1, Ly-6C, or CD25) followed by washing and staining with streptavidin–Red-670. For two-color staining, propidium iodide was also included at a concentration of 1 μg/ml. Live cells (propidium iodide–negative) were acquired and analyzed on a FACScan® or FACsort® flow cytometer.
Results
Upregulation of CD69 Expression on T Cell Subsets by CpG DNA.
We and others have shown that nonvertebrate DNA and synthetic ODNs containing unmethylated CpG motifs act as a strong B cell mitogen in vitro and elicit cytokine production by APC (1, 3, 4, 6, 8, 15–17). To investigate whether CpG DNA is capable of causing T cell activation, normal B6 mice were injected subcutaneously with insect DNA or synthetic ODNs; the two ODNs used contained a single CpG motif and were identical except that the C in the CpG motif was unmethylated for one ODN (CpG ODN) but methylated for the other (ZpG ODN). For insect DNA, we have shown previously that selective CpG methylation greatly reduced the capacity of DNA to cause B cell proliferation (4), indicating that, as for other nonvertebrate DNA, the mitogenic properties of insect DNA are controlled largely by unmethylated CpG motifs.
To examine T cell activation, draining LN cells from the injected mice were stained for upregulation of CD69 at 18 h after injection. As expected, both insect DNA and CpG (but not ZpG) ODNs induced strong CD69 upregulation on B cells (data not shown). Surprisingly, however, CD69 upregulation was also apparent on LN CD4+ and CD8+ cells, both on naive-phenotype (CD44lo/int) and memory-phenotype (CD44hi) cells (Fig. 1 A). Essentially similar findings applied when spleen cells were cultured overnight with insect DNA or ODNs in vitro (Fig. 1 B).
Figure 1.
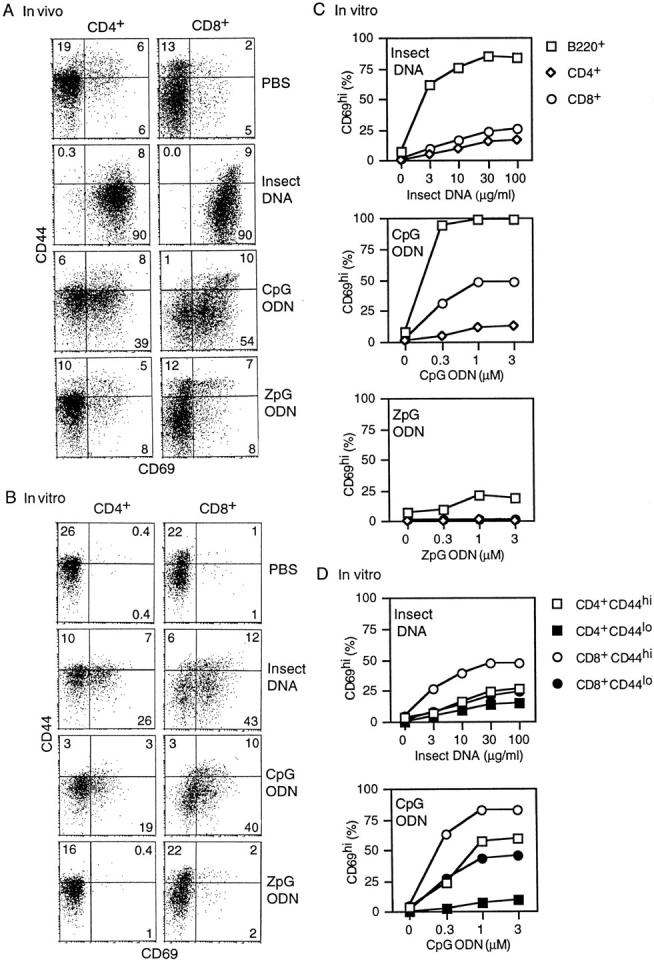
CD69 upregulation on T cells in response to insect DNA and synthetic ODNs. (A) B6 mice were injected subcutaneously with 100 μg/mouse D. melanogaster DNA or 50 μg/mouse ODNs. Draining LN cells were removed 18 h later and double-stained for CD4 or CD8 and also for CD44 and CD69 expression. (B) 5 × 106 unseparated spleen cells/ml were cultured in vitro in the presence of D. melanogaster DNA (100 μg/ml) or ODNs (1 μM) overnight. Cells were then harvested, purified with Ficoll gradients, and stained for CD4 or CD8, CD44, and CD69 expression. For C and D, see B, except that spleen cells were cultured with various concentrations of D. melanogaster DNA or ODNs.
Titrating the concentration of DNA/ODNs added to spleen cell cultures showed that CD69 upregulation was more marked on B (B220+) cells than on T cells and was more apparent on CD8+ cells than on CD4+ cells (Fig. 1, B and C). For both CD4+ and CD8+ cells, CD69 upregulation was most noticeable on memory CD44hi cells, but was clearly apparent on naive CD44lo/int cells (Fig. 1, B and D). Unlike insect DNA and CpG ODNs, ZpG ODNs caused only minimal activation of B cells and no detectable activation of T cells (Fig. 1, B and C).
In previous studies, CD69 upregulation on naive T cells was found after injecting mice with LPS (22), which raised the possibility that the stimulatory properties of DNA and ODNs reflected contamination with LPS. This possibility is unlikely because, with both insect DNA (Fig. 2 A) and CpG ODNs (data not shown), upregulation of CD69 on splenic CD8+ cells in vitro applied equally to LPS-nonresponder C3H/HeJ and LPS-responder C3H/HeOuJ spleen; by contrast, adding LPS to spleen cells induced T cell CD69 upregulation only on C3H/HeOuJ and not C3H/ HeJ spleen cells (Fig. 2 A). DNA-induced CD69 upregulation on T cells was also found for spleen cells taken from B cell–deficient μMT and NK-deficient beige mice (Fig. 2, B and C), suggesting that neither B cells nor NK cells were required for the stimulatory activity of DNA.
Figure 2.

DNA-induced upregulation of CD69 on spleen CD8+ cells from LPS-nonresponder, μMT, and beige mice. As in Fig. 1 B, spleen cells from C3H/HeJ versus C3H/HeOuJ mice (A), B6 versus μMT mice (B), or B6 versus beige mice (C) were cultured overnight with D. melanogaster DNA (100 μg/ml) or LPS (1 μg/ml) and then stained for CD8 and CD69 expression. The data show the percentage of CD8+ cells that were positive for CD69 relative to unstimulated CD8+ cells.
Role of APCs.
The above in vitro data apply to T cells in suspensions of spleen, an organ rich in APC. Significantly, addition of insect DNA or CpG ODNs to highly purified LN T cells in vitro caused only minimal CD69 upregulation both for CD4+ and CD8+ T cells (Fig. 3). However, strong CD69 upregulation on T cells occurred when purified LN T cells were supplemented with T and B cell–deficient APCs, e.g., spleen cells from Rag-1−/− (Rag-1 KO) mice (Fig. 3 A). This effect of APCs did not require T cell recognition of MHC molecules because similar findings occurred with APCs prepared from combined MHC I−/−II−/− (MHC KO) mice (Fig. 3 B). The capacity of APCs to restore DNA-induced upregulation of CD69 on T cells was not restricted to memory-phenotype T cells because similar findings applied when Rag-1−/− APCs were added to highly purified CD44lo/int cells (tested only for CD8+ cells; Fig. 3 C). Thus, the above findings indicate that CpG DNA-induced CD69 upregulation on T cells is APC dependent but MHC independent.
Figure 3.

DNA-induced upregulation of CD69 on T cells requires APCs. Purified CD4+ or CD8+ cells (106 cells/well) were cultured overnight in the presence or absence of D. melanogaster DNA (100 μg/ml), either alone or in the presence of RAG-1−/− spleen cells (A) or B cell– depleted spleen cells from MHC I−/−II−/− mice (panel B; B− MHC KO spl.); spleen APCs were added at 2 × 106 cells/well. In C, purified CD8+ CD44lo/int cells (106 cells/well) ± DNA were cultured overnight with titrated numbers of RAG-1 KO spleen cells. For A–C, cells were stained for CD8 and CD69 expression. The data show the percentage of gated CD8+ cells that were positive for CD69 expression relative to unstimulated CD8+ cells.
Upregulation of Other Cell Surface Molecules on T Cells and the Role of IFN-I.
DNA-induced activation of T cells in the presence of APCs was manifested not only by CD69 upregulation but also by significant upregulation of several other cell surface molecules, including B7-2, class I (Kb), Ly-6C, and ICAM-1; expression of certain other molecules, such as CD25 (IL-2Rα) and B7-1, remained unchanged (Fig. 4 A and data not shown). Under in vivo conditions, these surface marker changes were induced by injection of either insect DNA (Fig. 4 A) or CpG ODNs (data not shown) and affected B as well as T cells. Upregulation of surface markers was prominent for B6 mice (Fig. 4 A) but was also apparent at a lower level for 129 mice (Fig. 4 B; note the difference in scales for A and B). As illustrated for 129 mice, DNA-induced upregulation of multiple surface markers on T cells was also apparent in vitro, e.g., when insect DNA was added to purified T cells (CD8+ cells) supplemented with APC (Fig. 4 C).
Figure 4.

DNA-induced upregulation of various cell surface molecules on lymphoid subsets from normal versus IFN-IR−/− mice. (A and B) Normal B6 mice (A) or 129 versus IFN-IR−/− mice (B) were injected subcutaneously with D. melanogaster DNA (100 μg/mouse); 18 h later, draining LN cells were double-stained for expression of the markers shown and also for B220, CD4, and CD8 expression. The data are expressed as the percentage increase in the mean fluorescence intensity of staining for cells from DNA-injected mice versus PBS-injected mice. In C, purified CD8+ cells from 129 or IFN-IR−/− mice (106 cells/well) were cultured in vitro with or without D. melanogaster DNA (100 μg/ml), either alone or with RAG-1−/− spleen APCs (2 × 106 cells/well), and then stained for CD8 and the markers shown 18 h later. The data are expressed as the mean fluorescence intensity of staining for the marker shown, gating on CD8+ cells.
Of the above cell surface molecules upregulated by DNA, one of the markers, Ly-6C, is known to be upregulated by IFN-I (23). For this reason, it seemed possible that the T cell–activating properties of DNA were mediated largely or partly through production of IFN-I by APC. This possibility was examined by injecting insect DNA into IFN-IR−/− mice on a 129 background versus wild-type 129 mice as controls. The striking finding was that, for all of the cell surface markers studied, upregulation of these markers after DNA injection was virtually undetectable in IFN-IR−/− mice but clearly apparent in the control 129 mice (Fig. 4 B). Except for ICAM-1 (see below), essentially identical findings applied when DNA was added in vitro to purified populations of IFN-IR−/− versus 129 T cells (CD8+ cells) supplemented with normal IFN-IR+/+ APCs, i.e., Rag-1−/− spleen cells (Fig. 4 C). Since these APCs were ineffective with IFN-IR−/− T cells, the data indicate that DNA-induced T cell activation is (a) IFN-I dependent and (b) requires IFN-IR expression at the T cell level. Therefore, the implication is that DNA stimulates APC to produce IFN-I, which then activates T cells via IFN-IR.
T and B Cell Activation by IFN-I.
The notion that DNA causes T cell activation through production of IFN-I by APCs suggests that IFN-I can induce direct activation of T cells without a requirement for APCs. In support of this prediction, culturing purified B cells (Fig. 5, left) or T cells (CD8+ cells; Fig. 5, right) with graded concentrations of IFN-I (IFN-β) overnight caused much the same pattern of cell surface marker upregulation as was seen above with insect DNA; expression of other markers (B7-1, CD25) remained unchanged (data not shown). With addition of IFN-γ, by contrast, upregulation of surface markers was restricted to ICAM-1 on T cells and to B7-2, Kb and ICAM-1 on B cells. It should be noted that only IFN-γ and not IFN-I caused ICAM-1 upregulation on purified B cells, which suggests that the upregulation of ICAM-1 on B cells induced after DNA injection (Fig. 4) may have been mediated by IFN-γ rather than IFN-I; if so, the lack of ICAM-1 upregulation on IFN-IR−/− B cells after DNA injection (Fig. 4 B) implies that IFN-γ production in vivo may be IFN-I dependent. In contrast to B cells, both IFN-γ and IFN-I induced ICAM-1 upregulation on T cells (Fig. 5). Interestingly, for several molecules, notably CD69 and B7-2, IFN-γ potentiated the stimulatory properties of IFN-I (data not shown).
Figure 5.
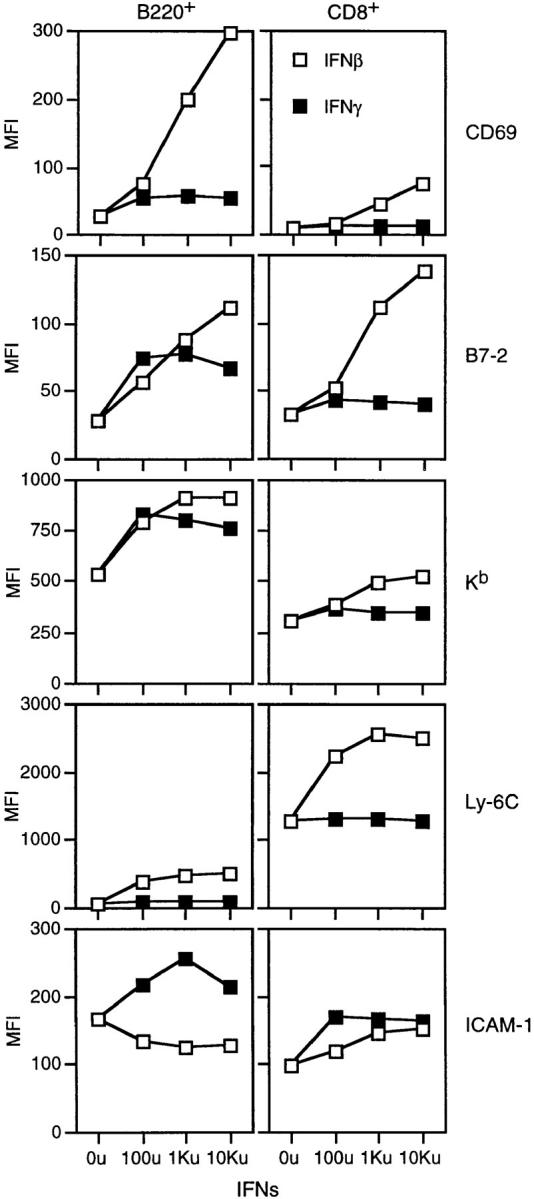
Expression of various cell surface markers on purified B (B220+) and CD8+ cells after incubation in vitro with IFN-I or IFN-γ. Cells (2 × 106/ well) were cultured with titrated concentrations of IFN-I (IFN-β) or IFN-γ overnight and then double-stained for B220 or CD8 and the markers shown.
These data applied to purified T and B cells. Essentially similar findings applied when unseparated spleen cells were cultured overnight in vitro with IFN-I (data not shown). In this situation, upregulation of the above markers (and also B7-1) was apparent on dendritic cells, i.e., on CD11c (N418)+ (24) cells.
Influence of IFN-I on T Proliferative Responses.
As discussed above, exposing T cells to IFN-I induced partial activation of the cells, i.e., upregulation of B7-2 and CD69. Would such activation alter the capacity of T cells to mount T proliferative responses? This question is of interest because it is well established that IFN-I has an antiproliferative effect on many cell types, including T cells (25–28). To examine this issue, we studied the proliferative response of T cells prepared from mice injected with insect DNA.
In initial experiments, normal 129 versus IFN-IR−/− mice were injected with large doses of insect DNA and then killed 1 d later to prepare purified LN CD8+ cells. Confirming earlier results (Fig. 4), after DNA injection CD69 expression was high on 129 CD8+ cells (>50%) but low on IFN-IR−/−CD8+ cells (<5%), indicating that only 129 CD8+ cells were activated after DNA injection. Proliferative responses of purified CD8+ cells after culture for 2 or 3 d in wells coated with a mixture of anti-TCR and anti-CD28 mAbs are shown in Fig. 6 A. As expected, proliferative responses of IFN-IR−/− CD8+ cells to TCR/ CD28 ligation were as high with cells from DNA-injected mice as with cells from PBS-injected controls. By contrast, with CD8+ cells from 129 mice, proliferative responses were considerably reduced (by about twofold) with cells from DNA-injected mice relative to PBS-injected mice.
Figure 6.


Prior exposure to Drosophila DNA in vivo inhibits proliferation of CD8+ cells in vitro. CD8+ cells were purified from the LNs of the indicated mice 18 h after injection of either PBS or D. melanogaster DNA (200 μg/mouse for 2C mice, 500 μg/mouse for 129 or IFN-IR KO mice). (A) Purified CD8+ cells from 129 or IFN-IR KO mice were cultured for 2 or 3 d in 200 μl wells precoated with a mixture of anti-TCR and anti-CD28 mAbs. The data show means and SDs of duplicate cultures. (B) Purified 2C CD8+ cells (2.5 × 104 cells/well) were cultured with Drosophila APCs (Ld/B7.1/ ICAM-1) and a titrated dose of QL9 peptide for the periods shown. Drosophila APCs were used at 2 × 105 cells/well. The data show means and SDs of triplicate cultures; for many of the data points, SDs were too small to be illustrated.
Similar findings applied to the proliferative response of CD8+ 2C TCR transgenic cells. As described previously (4), 2C CD8+ cells on a B6 (Kb) background respond strongly to a specific peptide, QL9, presented by Ld molecules on Ld-transfected Drosophila cells coexpressing B7-1 and ICAM-1 molecules. As illustrated in Fig. 6 B, purified CD8+ 2C cells from control PBS-injected donors gave strong proliferative responses to even low doses of QL9 peptide presented by transfected Drosophila cells as APCs; with limiting doses of peptide peak responses occurred on day 3 of culture. However, when CD8+ 2C cells were prepared from donors injected 1 d before with insect DNA, these CD8+ cells (>50% CD69+) gave poor proliferative responses to QL9 peptide in vitro. Relative to PBS- injected controls, the responses of 2C cells from DNA- injected mice were reduced by three- to fourfold. It should be noted that prior exposure to DNA in vivo did not reduce T cell viability, either before or after culture in vitro.
Discussion
As mentioned earlier, it is now well established that nonvertebrate DNA and synthetic ODNs containing unmethylated CpG motifs are stimulatory for B cells and APCs. The data in this paper extend these observations by showing that CpG DNA/ODNs are also able to stimulate T cells, including resting T cells, as manifested by induction of CD69 expression and upregulation of several surface markers. Significantly, however, stimulation of T cells by CpG DNA/ODNs required the presence of APCs and appeared to be mediated solely by IFN-I. Thus, in contrast to normal T cells, T cells from IFN-IR−/− mice could not be activated by CpG DNA/ODNs even when supplemented with normal (IFN-IR+/+) APCs. This finding suggested that CpG DNA/ODNs caused T cell activation by inducing APCs to release IFN-I, which then acted directly on T cells. In support of this notion, the pattern of T cell activation induced by CpG DNA/ODNs in the presence of APCs could be reproduced by culturing purified T cells with IFN-I in vitro; IFN-γ was ineffective.
As for T cells, stimulation of B cells in vivo by CpG DNA/ODNs appeared to be largely under the control of IFN-I. Thus, in contrast to normal mice, injecting CpG DNA/ODNs into IFN-IR−/− mice caused minimal activation of B cells. Nevertheless, under in vitro conditions we found that culturing unseparated IFN-IR−/− spleen cells with high concentrations of insect DNA did cause low but significant upregulation of CD69 and B7 on B cells and moderately strong proliferation of these cells (our unpublished data). B cell stimulation in this situation might be controlled by low-level IFN-I–independent production of IFN-γ because, like IFN-I, even small amounts of IFN-γ added to B cells in vitro caused upregulation of B7-2 (although not CD69) (Fig. 5). Alternatively, CpG DNA/ ODNs may be directly stimulatory for B cells (3, 5). Studies with purified B cells from combined IFN-IR−/−IFN-γ−/− mice will be needed to assess this possibility.
In the case of T cells, we have found that, as with LPS and Poly I:C (20, 22), injecting mice with CpG DNA/ODNs induces selective proliferation of memory-phenotype CD44hi CD8+ cells (our unpublished data); such proliferation can also be induced by injecting various cytokines, including IFN-γ and IFN-I. For IFN-I, it is important to emphasize that proliferation of memory-phenotype T cells in vivo does not reflect a direct action of IFN-I on T cells but, instead, stimulation of secondary cytokines, perhaps IL-15, by APCs (29). The activity of IFN-I on naive T cells that is described here is quite different. With naive T cells, IFN-I acts directly on the cells, apparently by binding to IFN-IR, and induces a form of partial activation associated with upregulation of CD69 and B7. However, such activation is not associated with entry into cell cycle (29).
It is of interest that the partial activation of T cells induced by IFN-I had an inhibitory effect on T cell function. Thus, in accordance with the evidence that IFN-I has an antiproliferative function (25–28), exposing T cells to CpG DNA for 1 d in vivo before TCR ligation in vitro led to a marked reduction in T proliferative responses; this effect was not seen with IFN-IR−/− T cells, implying a direct inhibitory effect by IFN-I on T cells. Indeed, in more recent studies we have observed that adding IFN-I to purified T cells in vitro has a comparable antiproliferative effect (our unpublished data). It is important to emphasize that this antiproliferative effect of IFN-I is only apparent when T cells are stimulated in the absence of viable APCs. Thus, when normal APCs are present, IFN-I has the opposite effect and considerably augments the T proliferative response, presumably by stimulating APCs to upregulate costimulatory molecules; this effect is not seen with Drosophila APCs, which are nonviable at 37°C and refractory to IFN-I. Hence, under in vivo conditions where APCs are prevalent, the antiproliferative function of IFN-I on T cells is presumably countered by an adjuvant-like effect of IFN-I on APCs. This scenario is supported by the finding that CpG DNA/ODNs (9–14) and IFN-I–inducing agents such as Poly I:C (20) act as powerful adjuvants for antigen-specific responses in vivo.
In summary, the data in this paper document that injecting mice with CpG DNA causes strong upregulation of activation markers on naive T cells. Such activation of T cells is not mediated by CpG DNA per se, however, but by IFN-I released via CpG DNA-induced stimulation of APCs. The functional consequences of partial T cell activation induced by IFN-I are complex and will be the topic of a future publication.
Acknowledgments
We thank Ms. Barbara Marchand for typing the manuscript and Drs. Susan Webb, Charles Surh, and Hidehiro Kishimoto for their comments.
This work was supported by grants CA38355, CA25803, AI21487, AI32068, and AG01743 from the United States Public Health Service. This is publication No. 11809-IMM from the Scripps Research Institute.
Abbreviations used in this paper
- ICAM
intercellular adhesion molecule
- KO
knockout
- ODNs
oligodeoxynucleotides
References
- 1.Yamamoto S, Yamamoto T, Shimada S, Kuramoto E, Yano O, Kataoka T, Tokunaga T. DNA from bacteria, but not from vertebrates, induces interferons, activates natural killer cells and inhibits tumor growth. Microbiol Immunol. 1992;36:983–997. doi: 10.1111/j.1348-0421.1992.tb02102.x. [DOI] [PubMed] [Google Scholar]
- 2.Messina JP, Gilkeson GS, Pisetsky DS. Stimulation of in vitro murine lymphocyte proliferation by bacterial DNA. J Immunol. 1991;147:1759–1764. [PubMed] [Google Scholar]
- 3.Krieg AM, Yi AK, Matson S, Waldschmidt TJ, Bishop GA, Teasdale R, Koretzky GA, Klinman DM. CpG motifs in bacterial DNA trigger direct B-cell activation. Nature. 1995;374:546–549. doi: 10.1038/374546a0. [DOI] [PubMed] [Google Scholar]
- 4.Sun S, Cai Z, Langlade-Demoyen P, Kosaka H, Brunmark A, Jackson MR, Peterson PA, Sprent J. Dual function of Drosophila cells as APC for naive CD8+T cells: implications for tumor immunotherapy. Immunity. 1996;4:555–564. doi: 10.1016/s1074-7613(00)80482-3. [DOI] [PubMed] [Google Scholar]
- 5.Sun S, Beard C, Jaenisch R, Jones P, Sprent J. Mitogenicity of DNA from different organisms for murine B cells. J Immunol. 1997;159:3119–3125. [PubMed] [Google Scholar]
- 6.Stacey KJ, Sweet MJ, Hume DA. Macrophages ingest and are activated by bacterial DNA. J Immunol. 1996;157:2116–2122. [PubMed] [Google Scholar]
- 7.Sparwasser T, Koch ES, Vabulas RM, Heeg K, Lipford GB, Ellwart JW, Wagner H. Bacterial DNA and immunostimulatory CpG oligonucleotides trigger maturation and activation of murine dendritic cells. Eur J Immunol. 1998;28:2045–2054. doi: 10.1002/(SICI)1521-4141(199806)28:06<2045::AID-IMMU2045>3.0.CO;2-8. [DOI] [PubMed] [Google Scholar]
- 8.Halpern MD, Kurlander RJ, Pisetsky DS. Bacterial DNA induces murine interferon-γ production by stimulation of interleukin-12 and tumor necrosis factor-α. Cell Immunol. 1996;167:72–78. doi: 10.1006/cimm.1996.0009. [DOI] [PubMed] [Google Scholar]
- 9.Roman M, Martin-Orozco E, Goodman JS, Nguyen M-D, Sato Y, Ronaghy A, Kornbluth RS, Richman DD, Carson DA, Raz E. Immunostimulatory DNA sequences function as T helper-1-promoting adjuvants. Nat Med. 1997;3:849–854. doi: 10.1038/nm0897-849. [DOI] [PubMed] [Google Scholar]
- 10.Chu RS, Targoni OS, Krieg AM, Lehman PV, Harding CV. CpG oligodeoxynucleotides act as adjuvants that switch on T helper 1 (Th1) immunity. J Exp Med. 1997;186:1623–1631. doi: 10.1084/jem.186.10.1623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Davis HL, Weeranta R, Walsschmidt TJ, Tygrett L, Schorr J, Krieg AM. CpG DNA is a potent enhancer of specific immunity in mice immunized with recombinant Hepatitis B surface antigen. J Immunol. 1998;160:870–876. [PubMed] [Google Scholar]
- 12.Weiner GJ, Liu HM, Wooldridge JE, Dahle CE, Krieg AM. Immunostimulatory oligodeoxynucleotides containing the CpG motif are effective as immune adjuvants in tumor antigen immunization. Proc Natl Acad Sci USA. 1997;94:10833–10837. doi: 10.1073/pnas.94.20.10833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lipford GB, Bauer M, Blank C, Reiter R, Wagner H, Heeg K. CpG-containing synthetic oligonucleotides promote B and cytotoxic T cell responses to protein antigen: a new class of vaccine adjuvants. Eur J Immunol. 1997;27:2340–2344. doi: 10.1002/eji.1830270931. [DOI] [PubMed] [Google Scholar]
- 14.Sun S, Kishimoto H, Sprent J. DNA as an adjuvant: capacity of insect DNA and synthetic oligodeoxynucleotides to augment T cell responses to specific antigen. J Exp Med. 1998;187:1145–1150. doi: 10.1084/jem.187.7.1145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Tokunaga T, Yano O, Kuramoto E, Kimura Y, Yamamoto T, Kataoka T, Yamamoto S. Synthetic oligonucleotides with particular base sequences from the cDNA encoding proteins of mycobacterium bovisBCG induce interferons and activate natural killer cells. Microbiol Immunol. 1992;36:55–66. doi: 10.1111/j.1348-0421.1992.tb01642.x. [DOI] [PubMed] [Google Scholar]
- 16.Sparwasser T, Miethke T, Lipford G, Borschert K, Hacker H, Heeg K, Wagner H. Bacterial DNA causes septic shock. Nature. 1997;386:336–337. doi: 10.1038/386336a0. [DOI] [PubMed] [Google Scholar]
- 17.Klinman DM, Yi A-K, Beaucage SL, Conover J, Krieg AM. CpG motifs present in bacterial DNA rapidly induce lymphocytes to secrete interleukin 6, interleukin 12, and interferon γ. Proc Natl Acad Sci USA. 1996;93:2879–2883. doi: 10.1073/pnas.93.7.2879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Chakrabarti D, Hultgren B, Stewart TA. IFN-α induced autoimmune T cells through the induction of intracellular adhesion molecule-1 and B7.2. J Immunol. 1996;157:522–528. [PubMed] [Google Scholar]
- 19.Herberman, R.B. 1997. Effect of alpha-interferons on immune function. Semin. Oncol. 24(3 Suppl. 9):S9-78–S9-80. [PubMed]
- 20.Tough DF, Borrow P, Sprent J. Induction of bystander T cell proliferation by viruses and type I interferon in vivo. Science. 1996;272:1947–1950. doi: 10.1126/science.272.5270.1947. [DOI] [PubMed] [Google Scholar]
- 21.Muller U, Steinhoff U, Reis LF, Hemmi S, Pavlovic J, Zinkernagel RM, Aguet M. Functional role of type I and type II interferons in antiviral defense. Science. 1994;264:1918–1921. doi: 10.1126/science.8009221. [DOI] [PubMed] [Google Scholar]
- 22.Tough DF, Sun S, Sprent J. T cell stimulation in vivo by lipopolysaccharide (LPS) J Exp Med. 1997;185:2089–2094. doi: 10.1084/jem.185.12.2089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Dumont FJ, Coker LZ. Interferon-α/β enhances the expression of Ly-6 antigens on T cells in vivo and in vitro. Eur J Immunol. 1986;16:735–740. doi: 10.1002/eji.1830160704. [DOI] [PubMed] [Google Scholar]
- 24.Metlay JP, Witmer-Pack MD, Agger R, Crowley MT, Lawless D, Steinman RM. The distinct leukocyte integrins of mouse spleen dendritic cells as identified with new hamster monoclonal antibodies. J Exp Med. 1990;171:1753–1771. doi: 10.1084/jem.171.5.1753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Holan V, Nakamura S, Minowada J. Inhibitory versus stimulatory effects of natural human interferon-alpha on proliferation of lymphocyte subpopulations. Immunology. 1992;75:176–181. [PMC free article] [PubMed] [Google Scholar]
- 26.Noronha A, Toscas A, Jensen MA. Interferon beta decreases T cell activation and interferon gamma production in multiple sclerosis. J Neuroimmunol. 1993;46:145–153. doi: 10.1016/0165-5728(93)90244-s. [DOI] [PubMed] [Google Scholar]
- 27.Krishnaswamy G, Smith JK, Srikanth S, Chi DS, Kalbfleisch JH, Huang SK. Lymphoblastoid interferon-alpha inhibits T cell proliferation and expression of eosinophil-activating cytokines. J Interferon Cytokine Res. 1996;16:819–827. doi: 10.1089/jir.1996.16.819. [DOI] [PubMed] [Google Scholar]
- 28.Petricoin EF, III, Ito S, Williams BL, Audet S, Stancato LF, Gamero A, Clouse K, Grimley P, Weiss A, Beeler J, et al. Antiproliferative action of interferon-α requires components of T-cell-receptor signalling. Nature. 1997;390:629–632. doi: 10.1038/37648. [DOI] [PubMed] [Google Scholar]
- 29.Zhang X, Sun S, Hwang I, Tough DF, Sprent J. Potent and selective stimulation of memory-phenotype CD8+T cells in vivo by IL-15. Immunity. 1998;8:591–599. doi: 10.1016/s1074-7613(00)80564-6. [DOI] [PubMed] [Google Scholar]