

Abstract
Dendritic cells, but not macrophages, efficiently phagocytose apoptotic cells and cross-present viral, tumor, and self-antigens to CD8+ T cells. This in vitro pathway corresponds to the in vivo phenomena of cross-priming and cross-tolerance. Here, we demonstrate that phagocytosis of apoptotic cells is restricted to the immature stage of dendritic cell (DC) development, and that this process is accompanied by the expression of a unique profile of receptors, in particular the αvβ5 integrin and CD36. Upon maturation, these receptors and, in turn, the phagocytic capacity of DCs, are downmodulated. Macrophages engulf apoptotic cells more efficiently than DCs, and although they express many receptors that mediate this uptake, they lack the αvβ5 integrin. Furthermore, in contrast to DCs, macrophages fail to cross-present antigenic material contained within the engulfed apoptotic cells. Thus, DCs use unique pathways for the phagocytosis, processing, and presentation of antigen derived from apoptotic cells on class I major histocompatibility complex. We suggest that the αvβ5 integrin plays a critical role in the trafficking of exogenous antigen by immature DCs in this cross-priming pathway.
Keywords: dendritic cells, phagocytosis, CD36, integrins, cross-presentation
poptosis triggers a distinct sequence of events characterized by the expression of phosphatidylserine (PS)1 on the cell surface (1), DNA fragmentation or laddering (2), and the release of membrane-bound cell fragments called apoptotic blebs and bodies (3, 4). Apoptotic cells and bodies are phagocytosed via various receptors that recognize PS and other undefined ligands unique to the surface of apoptotic material (1). In this way, dying cells, which contain potentially inflammatory factors, are rapidly cleared by neighboring cells, scavenger cells, or macrophages without inducing an inflammatory response (5, 6).
We have recently demonstrated that human dendritic cells (DCs) acquire antigens from apoptotic cells and stimulate antigen-specific class I–restricted CD8+ T cells (7). Apoptotic death is a critical trigger for this pathway, as antigen from necrotic cells is not presented on MHC I (7). This in vitro model for cross-priming may be akin to the in vivo phenomenon in which antigens derived from MHC-mismatched donor cells are cross-presented on host APC's class I MHC (8–10). This event is potentially significant in the maintenance of tolerance to tissue-specific antigens and the induction of immunity to antigens that may not access the endogenous MHC class I pathway of a professional APC.
Here, we investigate the developmental stage and receptors used by DCs for the efficient phagocytosis of apoptotic cells in this exogenous pathway. DCs undergo a differentiation process that includes ‘immature' and ‘mature' stages (11). Monocytes that have been cultured in GM-CSF and IL-4 develop into immature DCs. These cells are analogous to peripheral tissue DCs. Immature DCs are characterized by high endocytic and macropinocytic activity (12–16) and low expression of accessory signals for T cell activation (11, 17). Maturation of DCs, induced by antigen, cytokines, or signaling molecules (e.g., LPS, monocyte-conditioned medium [MCM], ceramide, CD40L, TNF-α, and PGE2 [12, 13, 18– 22]) is associated with the downregulation of antigen uptake, but an enhancement of the T cell stimulatory capacity. By using a modified FACS®-based method for detecting phagocytic uptake, we compared immature DCs, mature DCs, and macrophages for their capacity to phagocytose apoptotic cells and cross-present viral antigens to CTLs (23). We report that immature DCs are specialized in their ability to cross-present antigen and that the phagocytosis of apoptotic cells correlates with the use of a unique receptor profile.
Materials and Methods
Media.
RPMI 1640 supplemented with 20 μg/ml of gentamicin (GIBCO BRL, Gaithersburg, MD), 10 mM Hepes (Mediatech, Herndon, VA), and either 1% human plasma, 5% pooled human serum (c-Six Diagnostics, Mequon, WI), or 5% single donor human serum was used for DC preparation, cell isolation, and culture conditions (18, 24).
Preparation of Cells.
PBMCs, DCs, macrophages, and T cells were prepared as previously described (18, 19, 24). In brief, peripheral blood was obtained from normal donors in heparinized syringes and PBMCs were isolated by sedimentation over Ficoll-Hypaque (Amersham Pharmacia Biotech, Piscataway, NJ). T cell–enriched and T cell–depleted fractions were prepared by rosetting with neuraminidase-treated sheep red blood cells (24). Immature DCs were prepared from the T cell–depleted fraction by culturing cells in the presence of GM-CSF and IL-4 for 7 d. 1,000 U/ml of GM-CSF (Immunex Corp., Seattle, WA) and 500–1,000 U/ml of IL-4 (Schering-Plough Corp., Kenilworth, NJ) were added to the cultures on days 0, 2, and 4. To generate mature DCs, the cultures were transferred to fresh wells on day 7 and MCM was added for an additional 3–4 d (18, 19). At day 7, >95% of the cells were CD14−CD83−HLA-DRlo DCs. On days 10–11, 80–100% of the cells were of the mature CD14−CD83+ HLA-DRhi phenotype. FACSort® (Becton Dickinson, San Jose, CA) was used to generate highly pure populations of immature and mature DCs, based on their CD83− and CD83+ phenotypes, respectively. Macrophages were isolated from T cell–depleted fractions by plastic adherence for 1 h. After 24 h, cells were removed from the plates and placed in Teflon beakers for 3–9 d. T cells were further purified from the T cell–enriched fraction by removing contaminating monocytes, NK cells, and B cells (24).
Antibodies.
Antibodies to the following proteins were used: CD8-PE, CD14-PE, HLA-DR–PE, HLA-DR–biotin (Becton Dickinson), IgG2b (clone 6603001; Coulter Corp., Hialeah, FL), CD8 (CRL 8014; American Type Culture Collection, Rockville, MD), CD83 (clone HB15a; Coulter Corp.), MHC I (W6/ 32, ATCC clone HB95), CD36 (clone FA6; obtained from the fifth international workshop on leukocyte differentiation antigens), αv (clone CLB-706, Chemicon International, Inc., Temecula, CA; clone 69.6.5, Coulter Corp.), β1 (clone 6S6; Chemicon International, Inc.), β3 (clone SZ21, Coulter Corp.; clone RUU-PL 7F12, Becton Dickinson), β5 (clone B5-IVF2; Upstate Biotechnology, Inc., Lake Placid, NY), αvβ3 (clone 23C6; PharMingen, San Diego, CA), αvβ5 (clone P1F6; Chemicon International, Inc.), CD71 (Dako Corp., Carpinteria, CA), mannose receptor (clone 3.2PB1; a gift from A. Lanzavecchia, Basel Institute, Sweden), and influenza nucleoprotein (ATCC clone HB85).
Induction of Apoptotic Death.
Monocytes were infected with influenza virus in serum-free RPMI. These cells undergo viral-induced apoptotic death within 6–8 h. Cell death was confirmed using the Early Apoptosis Detection kit (Kayima Biomedical Co., Seattle, WA [7]). As previously described, cells are stained with Annexin V–FITC (Ann V) and propidium iodide (PI). Early apoptosis is defined by Ann V+/PI− staining as determined by FACScan® (Becton Dickinson). To ensure that we were studying the uptake of early apoptotic cells, the kinetics of death were carefully worked out. 5–8 h after infection, monocytes first externalize PS on the outer leaflet of their cell membrane, as detected with Ann V. By 8–10 h, these cells were TUNEL (Tdt-mediated dUTP- biotin nick-end labeling) positive. It was not until 24–36 h that the majority of the monocyte population included trypan blue into the cytoplasm, an indicator of secondary necrosis (Albert, M.L., and N. Bhardwaj, unpublished data, references 25, 26). HeLa cells were triggered to undergo apoptosis using a 60 UVB lamp (Derma Control Inc.), calibrated to provide 2 mJ/cm2/s. The kinetics of cell death in these cells has been defined previously (4).
Phagocytosis of Apoptotic Cells.
Monocytes and HeLa cells were dyed red using PKH26-GL (Sigma Biosciences, St. Louis, MO), and induced to undergo apoptosis by influenza infection and UVB irradiation, respectively. After 6–8 h, allowing time for the cells to undergo apoptosis, they were cocultured with phagocytic cells that were dyed green using PKH67-GL (Sigma Biosciences), at a ratio of 1:1. Macrophages were used 3–6 d after isolation from peripheral blood; immature DCs were used on days 6–7 of culture; and mature DCs were used on days 10–11. Where direct comparison of cells was needed, cells were prepared from the same donor on different days. In blocking experiments, the immature DCs were preincubated in the presence of 50 μg/ml of various mAbs for 30 min before the establishment of cocultures. After 45–120 min, FACScan® analysis was performed and double positive cells were enumerated.
Phagocytosis of Latex Beads.
Immature DCs were preincubated at 37°C with mAbs specific for αv and αvβ5. 106 cells were then cultured with 5 × 107 red fluorescent microspheres (diameter 1 μm, 2.5% solids, carboxylate-modified latex; Sigma Chemical Co.) for varying periods of time. Alternatively, the cells were maintained at 4°C. At the end of the assay, cells were separated from unengulfed beads by density gradient centrifugation and analyzed by FACScan® analysis (27).
Immunofluorescence.
Cells were adhered to Alcian blue (Sigma Chemical Co.) treated coverslips and fixed using 100% acetone. Cells were stained with antiinfluenza nucleoprotein antibody (HB85; American Type Culture Collection) and Texas red conjugated goat anti–mouse IgG (Jackson ImmunoResearch Labs., Inc., West Grove, PA). This was followed by staining with biotinylated anti–HLA-DR (Becton Dickinson) followed by FITC-conjugated streptavidin (Jackson ImmunoResearch). Cells were visualized using a Zeiss Axioplan2 microscope (Carl Zeiss, Inc., Thornwood, NY).
Assay for Cross-priming of Apoptotic Cells.
Various APC populations were prepared from HLA-A2.1+ donors as described above. Mature DCs were further purified by labeling with the DC-restricted marker CD83, followed by cell sorting on the FACSort®. Immature DCs were CD14− and sorted by FACSort® as a CD83− population. Mature macrophages were generated by culturing an adherent mononuclear cell fraction in a Teflon beaker for 9 d. These APC populations were cocultured with HLA-A2.1− influenza-infected monocytes. After 12 h, the APCs were loaded with Na51CrO4 and used as targets for influenza-specific CTLs in a standard 51Cr-release assay (7, 24). Specific lysis indicates that the APC had cross-presented antigenic material derived from the apoptotic cell, leading to the formation of specific peptide–MHC class I complexes on its surface. Specific lysis = (% killing of APCs cross-presenting influenza-infected monocytes) − (% killing of APCs cross-presenting uninfected monocytes). Background lysis ranged from 0 to 8%. Controls included influenza-infected and uninfected mature DCs, immature DCs, and macrophages. The HLA-A2.1− monocytes used as a source of apoptotic material were also tested as targets to demonstrate the absence of lysis when using a mismatched target. For the control targets, specific lysis = (% killing of influenza-infected APCs) − (% killing of uninfected APCs). Background lysis ranged from 0 to 5%. Maximal influenza-specific killing was determined using T2 cells (a TAP−/−HLA-A2.1+ class II− cell line) pulsed for 1 h with 1 μM of the immunodominant influenza matrix peptide, GILGFVFTL, as targets (28). Responses varied from 25 to 60% as a function of the individual's prior exposure to influenza.
Reverse Transcriptase PCR.
RNA was purified from highly purified sorted cell populations of immature and mature DCs as described above. Messenger RNA for β3, β5, and CD36 were identified using a one-step reverse transcriptase PCR reaction (Titan kit; Boehringer Mannheim, Indianapolis, IN). The forward primer 5′-TGAGAAGTGCCCCTGCCC was used for both β3 and β5. The reverse primers 5′-GTTGGCTGTGTCCCATTTTGCT and 5′-TTGTAGGATTTGTGAACTTG were used for β3 and β5 to obtain 438- and 509-bp products, respectively (primer sequences were provided by S. Silletti, Scripps, San Diego, CA). The forward primer 5′-GGGAATTCATATGAAATCATAAAAGCAACAAACAT and the reverse primer 5′-CGGAATTCTACATTTCACTTCCTCATTTTCTG for CD36 yielded a product of 392 bp (29). The reverse transcriptase reaction was carried out for 30 min at 56°C followed by 30 cycles of amplification. After 30 cycles of PCR the samples were visualized on an agarose gel.
Results
Immature DCs Efficiently Phagocytose Apoptotic Cells.
Based on previous observations that immature DCs are the cells responsible for capturing antigen (11), we predicted that apoptotic cells would be engulfed best by immature DCs. To test this hypothesis, we established a phagocytosis assay that allowed us to visually detect the uptake of apoptotic cells, and compare the phagocytic capacity of immature DCs, mature DCs, and macrophages. In brief, immature DCs were prepared by culturing a T cell–depleted fraction from peripheral blood in the presence of IL-4 and GM-CSF. Mature DCs were generated with the addition of MCM and these cells expressed the cell surface DC-restricted maturation marker CD83 (18, 19, 30). Macrophages were prepared by culturing a plastic adherent cell population in Teflon beakers for 3–9 d. As a source of apoptotic cells, we used influenza-infected monocytes (7); virus infection induces apoptotic death in these cells within 6–10 h (7, 25, 26). Monocytes were first infected with influenza virus as previously described (24), then dyed red using PKH-26 (Sigma Biosciences). After 6–8 h, the various APCs were dyed green using the fluorescent cell linker compound PKH67-GL (Sigma Biosciences) and cocultured with the apoptotic cells at a ratio of 1:1. After 2 h at 37°C, cocultures of cells were analyzed by FACScan® analysis, allowing for quantification of phagocytic uptake as double positive cells. 80% of the macrophages, 50% of the immature DCs, and <10% of the mature DCs engulfed the apoptotic monocytes after 2 h of coculture (Fig. 1 A). The smear of double positive cells (PKH67-labeled APCs that engulfed the PKH26-labeled apoptotic cells) indicates that both apoptotic bodies and whole apoptotic cells served as ‘food' for the phagocytic cell (Fig. 1, iii, vi, and ix). Note that as the forward scatter of the APCs increased and the setting of the FACS® shifted, the dying monocytes were excluded from the established region (Fig. 1, ii, v, and viii). Maximal uptake by all APC populations was achieved within 2–4 h and partially depended upon the source of apoptotic cell used (Fig. 1 B and data not shown). Given this kinetic data, we believe that macrophages and DCs engage and internalize dying cells while still displaying features of early apoptotic cell death. This data also demonstrates that it is the immature DC that preferentially acquires apoptotic material compared with the mature DC. The source of apoptotic cells was not critical, since we obtained similar results with UVB-irradiated HeLa cells (see Fig. 7, and data not shown).
Figure 1.


Immature but not mature DCs efficiently phagocytose apoptotic cells. Freshly isolated blood monocytes were infected with live influenza A, PR/8 (Spafas Inc., Storrs, CT), labeled with the PKH26-GL fluorescent cell linker compound (Sigma Biosciences), and incubated at 37°C for 6–8 h, allowing apoptosis to occur. Macrophages, immature DCs, and mature DCs were dyed with PKH67-GL and added to the culture wells containing the apoptotic monocytes at a ratio of 1:1. Cells were analyzed by FACScan® where double positive cells indicate uptake of the apoptotic cells by the various APCs (iii, vi, and ix). We used the various APCs alone to establish the proper settings (i, iv, and vii). Note that as the forward scatter of the APCs increased, the dying monocytes were excluded from the established region (ii, v, and viii). After 2 h, 80% of the macrophages, 50% of the immature DCs, and <10% of the mature DCs had engulfed the apoptotic monocytes (A). In an independent experiment, macrophages (squares), immature DCs (diamonds), and mature DCs (circles) were prepared, and cocultures with apoptotic monocytes were established as described above. FACS® was performed at various time points. Percentage of phagocytosis was calculated based on the number of double positive cells (B).
Figure 7.


Direct inhibition of phagocytosis by anti-αvβ5 and anti-CD36 antibodies. HeLa cells were labeled with PKH26-GL, followed by irradiation using a 60 UVB lamp (Derma Control Inc.), calibrated to provide 240 mJ cm−2 in 2 min, sufficient for the induction of apoptosis. After 6–8 h, immature DCs dyed with PKH67-GL and pretreated with 50 μg/ml of various mAbs for 30 min were added to the wells containing apoptotic HeLa cells. 45–60 min later, cells were analyzed by FACS® for double positive cells. Phagocytic uptake is reported as a percentage of untreated cells. Maximal phagocytosis ranged from 44 to 52%. Results from three experiments were averaged and means plotted + SD. Similar results (data not shown) were obtained when apoptotic monocytes were used (A). Immature DCs were incubated with red fluorescent latex beads at 37°C (squares), 4°C (diamonds), or 37°C in the presence of 50 μg/ml anti-αvβ5 (circles), or anti-αv (triangles). The results shown indicate percentage of phagocytosis over background (B).
To confirm that this FACS® assay was measuring phagocytosis, we carried out the assay at 4°C and in the presence of inhibitors of phagocytosis. Both low temperature (Fig. 2 A) and cytochalasin D, an inhibitor of cytoskeletal function, blocked uptake (Fig. 2 B). Phagocytosis by immature DCs also requires divalent cations as EDTA was inhibitory (Fig. 2 C). To visually confirm the uptake recorded by FACS®, we prepared cytospins of the dyed cocultures. The frequency of uptake correlated with that measured on FACS® (data not shown). We also performed immunofluorescence on cocultures of immature DCs labeled with anti– HLA-DR (DR) and apoptotic influenza-infected monocytes labeled with antiinfluenza nucleoprotein (NP) (Fig. 3). In the top panel an apoptotic cell is seen just prior to being engulfed by a DC (arrowhead). After phagocytosis, apoptotic cells were found in DR+ vesicles (arrows), but not in the cytoplasm.
Figure 2.

Low temperature, Cytochalasin D, and EDTA block phagocytosis of apoptotic cells by immature DCs. Apoptotic monocytes and immature DCs were prepared as described above. Immature DCs were preincubated at 4°C (A) in the presence of varying concentrations of Cytochalasin D (B) or EDTA (C) for 30 min. Apoptotic monocytes were then added to the DC cultures at 4°C (A) or 37°C (B and C). FACS® analysis was performed after 1–2 h. Data shown are representative of five independent experiments in which influenza- infected monocytes or UVB-irradiated HeLa cells were sources of apoptotic food for the immature DCs. Percentages of inhibition ± SD for these experiments were: 4°C, 85 ± 7%; 10 μM Cytochalasin D, 69 ± 3%; and 2 mM EDTA, 76 ± 14%.
Figure 3.
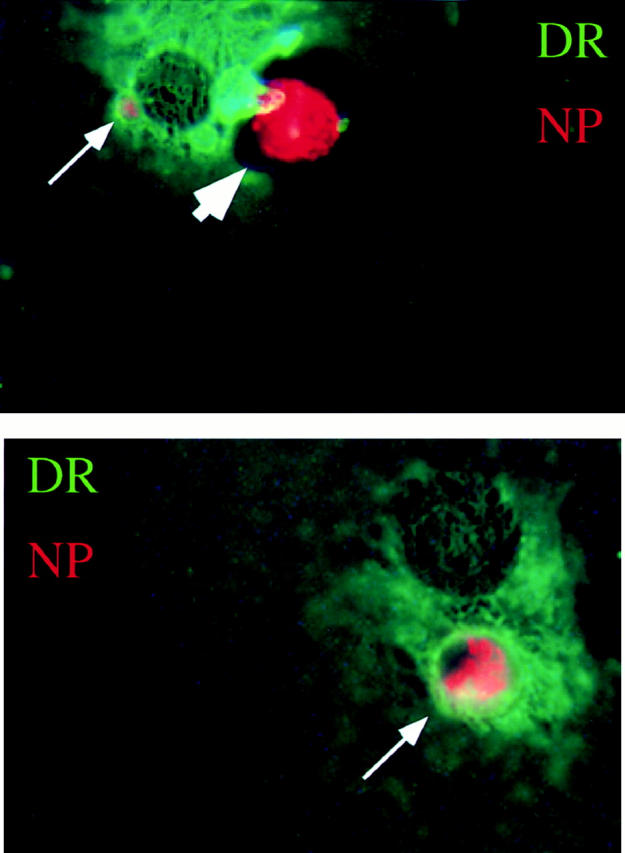
Immature DCs engulf influenza-infected monocytes. Influenza-infected apoptotic monocytes were cocultured with immature DCs for 1 h, after which the cells were adhered to a coverslip and fixed with acetone. Immunofluorescence was performed with antiinfluenza nucleoprotein antibodies (NP) and Texas red–conjugated goat anti–mouse IgG and by biotinylated anti–HLA-DR (DR) followed by FITC-conjugated streptavidin. The large arrowhead indicates apoptotic cell outside the DC before engulfment. Small arrows indicate apoptotic material derived from the influenza-infected monocytes within DR+ vesicles of the DC. These images were not generated on a confocal scope, so the structures of the DC underlying the apoptotic cell can be seen.
Only Immature DCs Cross-present Antigen from the Apoptotic Cell on Class I MHC.
We next correlated the phagocytic capability of macrophages and DCs with their ability to cross-present antigenic material derived from apoptotic cells. The cells were prepared from HLA-A2.1+ donors (18, 19), cocultured with HLA-A2.1− influenza-infected monocytes for 12 h, and then loaded with Na51CrO4 for use as targets for influenza-specific CTLs (7, 24). Specific lysis indicates that the APCs cross-presented antigenic material derived from the apoptotic cell by forming specific peptide–MHC class I complexes on its surface (Fig. 4 A). As a direct comparison with the endogenous pathway for class I MHC presentation, the same APC populations were infected with live influenza virus and used as targets (Fig. 4 B).
Figure 4.

Immature DCs but not mature DCs or macrophages cross-present antigenic material derived from apoptotic cells. Various populations of HLA-A2.1+ APCs were cocultured with HLA-A2.1− influenza-infected monocytes. After 12 h, the APCs were loaded with 51Cr and used as targets for HLA-A2.1+ influenza-reactive CTL lines. Mature DCs were isolated by labeling with the DC-restricted marker CD83, followed by cell sorting on the FACSort®. Immature DCs were CD14− and were sorted by FACSort® as a CD83− population. Mature macrophages were generated by culturing an adherent mononuclear cell fraction in a Teflon beaker for 9 d. Effector/target ratios = 45:1 and 15:1 (A). Controls included infected and uninfected mature DCs, immature DCs, and macrophages. The HLA-A2.1− monocytes used as a source of apoptotic material were also tested as targets to demonstrate the absence of lysis when using a mismatched target. Effector/target ratios = 45:1 and 15:1. Results are representative of three experiments and the values shown represent the mean of triplicate wells (B).
Although mature DCs were efficient targets when infected with influenza, they were unable to cross-present antigens, presumably because they had downregulated the ability to phagocytose the apoptotic monocytes. However, the immature DCs did cross-present antigens from apoptotic cells. Furthermore, if the immature DCs were cocultured with the apoptotic cells in the presence of MCM, a maturation stimulus, they were even better targets. This is possibly due to the upregulation of costimulator and adhesion molecules (13, 31), or to the increased stability of peptide–MHC I complexes. Given that maximal uptake of apoptotic cells by immature DCs occurs between 2 and 4 h (Fig. 1 D), we believe that cross-presentation of apoptotic material reflects the phagocytosis and processing of early apoptotic cells rather than secondary necrotic cells (see Materials and Methods). With respect to this issue, it is important to recognize that the influenza-infected monocytes require 24 h to undergo secondary necrosis (Albert, M.L., and N. Bhardwaj, unpublished data; references 25, 26).
Notably, macrophages that efficiently phagocytose apoptotic cells (Fig. 1 A) did not cross-present antigens to CTLs (Fig. 4 B). Presumably, the engulfed material is degraded, not cross-presented, on MHC I. This profound difference between the DC and macrophage populations is supported by our previous findings that macrophages do not cross-present antigens from apoptotic cells during the induction phase of a class I–restricted antigen-specific T cell response. In fact, when put into culture with DCs in a competition assay, they sequester the apoptotic material and abrogate the CTL response (7).
Immature DCs Can Be Distinguished from Macrophages by Intracellular Expression of CD83 and a Unique Profile of Phagocytic Receptors.
We investigated the possibility that immature DCs might phagocytose apoptotic cells via pathways distinct from macrophages. To clearly distinguish these cells, we characterized them phenotypically. Immature DCs are distinguished by the absence of both CD14, a macrophage restricted marker, and CD83, a maturation marker for DCs (30). We have extended the use of CD83, finding that immature DCs can be distinguished from both macrophages and mature DCs by their intracellular expression of CD83. Macrophages do not express CD83 intra- or extracellularly, whereas mature DCs express CD83 both intra- and extracellularly (Fig. 5).
Figure 5.

Intracellular but not extracellular CD83 expression distinguishes immature DCs from mature DCs and macrophages. Macrophages, immature DCs, and mature DCs were prepared as previously described. Cells were either untreated (Surface) or permeabilized with saponin (Intracellular), and incubated with anti-CD83. Cells were then labeled using a PE-conjugated GAM-Ig (Sigma Biosciences). An isotype-matched antibody was used (IgG2a) as a control.
These APC populations were examined for surface expression of receptors involved in phagocytosing apoptotic material (Table 1). These include: αvβ3 and CD36, which act as coreceptors for engulfment of apoptotic neutrophils and lymphocytes by macrophages (32, 33); and CD14, which has been implicated in the uptake of apoptotic cells by macrophages (34). While studying the immature DC populations, we identified a discrepancy in the expression of the αv and β3 integrin chains and investigated the possibility that αv was binding an alternate β chain. Using antibodies that recognize combined epitopes of the αvβ3 and the αvβ5 heterodimers, we noted the selective expression of αvβ5 on immature DCs (Fig. 6 A). As is true for most receptors involved in antigen uptake (11, 12), the expression of CD36, αvβ5, and mannose receptor on DCs is downregulated with maturation (Fig. 6 B, Table 1).
Table 1.
Receptor Profile of Various APCs
| Immature DCs Day 7 | Mature DCs Day 11 | Monocytes Day 0 | Macrophages Day 3 | Macrophages Day 9 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| n = 7 | n = 6 | n = 4 | n = 5 | n = 5 | ||||||
| CD8* | 0–2 (3 ± 1)§ | 0–3 (4 ± 1) | 0–1 (4 ± 1) | 0–5 (3 ± 1) | 1–2 (3 ± 1) | |||||
| CD14* | 1–10 (4 ± 1) | 0–6 (4 ± 1) | 83–97 (149 ± 90) | 67–98 (61 ± 20) | 49–90 (68 ± 48) | |||||
| CD83‡ | 8–43 (8 ± 4) | 73–99 (114 ± 15) | 0–1 (3 ± 1) | 4–19 (4 ± 2) | 2–14 (4 ± 2) | |||||
| MHC I‡ | 99–100 (311 ± 49) | 96–100 (280 ± 69) | 99–100 (99 ± 38) | 97–99 (164 ± 24) | 95–100 (116 ± 18) | |||||
| HLA-DR* | 88–100 (84 ± 35) | 98–99 (188 ± 40) | 94–99 (63 ± 9) | 97–99 (173 ± 34) | 5–89 (25 ± 27) | |||||
| CD8‡ | 3–9 (4 ± 1) | 2–8 (3 ± 1) | 0–2 (3 ± 1) | 1–4 (3 ± 0) | 1–5 (3 ± 1) | |||||
| IgG2b‡ | 3–7 (3 ± 1) | 2–7 (3 ± 1) | 0–2 (3 ± 0) | 1–4 (3 ± 1) | 1–5 (3 ± 1) | |||||
| CD36‡ | 75–99 (75 ± 31) | 20–50 (9 ± 2) | 89–99 (62 ± 18) | 64–98 (69 ± 45) | 9–55 (15 ± 16) | |||||
| αv (CD51)‡ | 45–76 (14 ± 3) | 11–40 (9 ± 3) | 2–12 (4 ± 1) | 2–35 (6 ± 2) | 18–75 (10 ± 5) | |||||
| β1 (CD29)‡ | 99–100 (287 ± 57) | 84–100 (112 ± 28) | 95–100 (61 ± 33) | 83–100 (72 ± 23) | 68–99 (104 ± 29) | |||||
| β3 (CD61)‡ | 14–34 (7 ± 2) | 10–24 (6 ± 1) | 76–91 (40 ± 16)‖ | 15–85 (14 ± 9)‖ | 19–92 (16 ± 9) | |||||
| β5 ‡ | 78–95 (30 ± 8) | 32–57 (11 ± 2) | 6–8 (5 ± 1) | 8–15 (7 ± 2) | 5–8 (3 ± 1) | |||||
| αvβ3 (VnR)‡ | 22–28 (7 ± 1) | 14–30 (7 ± 2) | 4–8 (4 ± 1) | 7–27 (5 ± 2) | 19–68 (10 ± 4) | |||||
| αvβ5 ‡ | 81–90 (25 ± 5) | 12–53 (9 ± 3) | 1–14 (4 ± 1) | 0–11 (3 ± 1) | 6–15 (4 ± 1) | |||||
| CD71‡ | 75–99 (50 ± 29) | 75–94 (61 ± 27) | 3–7 (4 ± 0) | 14–30 (7 ± 2) | 58–81 (30 ± 7) | |||||
| Mannose receptor‡ | 99–100 (272 ± 68) | 38–80 (26 ± 11) | 2–7 (4 ± 0) | 24–63 (13 ± 8) | 48–95 (34 ± 20) |
PE-conjugated.
Unconjugated antibody.
Results are expressed as range of values, calculated as percentage of positive cells as compared with an isotype-matched control antibody. Values in parentheses indicate the average of geometric mean fluorescence intensities of the various samples ± SD.
The β3 expression in the monocytes and day 3 macrophages does not match the αvβ3 expression due to contaminating platelets that stuck to the cells during isolation, as determined by staining with anti-CD41 (data not shown).
Figure 6.


Protein and mRNA expression of αvβ5 and CD36 are downregulated during DC maturation. Immature DCs (A) and mature DCs (B) were incubated with anti-αvβ3 (clone 23C6; PharMingen), anti-CD36 (clone FA6; obtained from the fifth international workshop on leukocyte differentiation antigens), or anti-αvβ5 (clone P1F6; Chemicon International, Inc.), followed by PE-conjugated GAM-Ig (Sigma Biosciences). All cells were analyzed by FACScan®. (C) RNA was isolated from highly purified sorted cell populations of immature and mature DCs as previously described. Reverse transcription was carried out and after 30 cycles of PCR the distinct bands for β3, β5, and CD36 could be seen in the immature DCs (lane 1). In mature DCs, only a faint band for CD36 and no band for β5 could be visualized (lane 2), indicating minimal mRNA. In contrast, a band was evident for β3 in the mature DCs. Note that the doublet for β3 is an artifact in this particular exposure and does not indicate two unique bands (lane 2). As a positive control, extracts from Bowes melanoma cells, which are known to express β3 and CD36 (29), were run (lane 3). As negative control, the Bowes melanoma cells were run in the absence of a reverse transcriptase (lane 4).
To evaluate whether this downregulation could be observed on the level of mRNA expression, we performed reverse transcriptase PCR using primers specific for β3, β5, and CD36 (Fig. 6 C). Immature DCs (lane 1) showed amplified DNA of the appropriate size for β3, β5, and CD36. In contrast, in mature DCs (lane 2), no β5 and much fewer CD36 sequences were seen, whereas β3 sequences were comparable to those in immature cells. These data, although not quantitative, are consistent with the levels of protein expression observed by FACS® and suggest that phagocytic receptor expression in DCs may be regulated at a transcriptional level as mRNA expression of CD36 and β5 is downregulated during maturation.
αvβ5 and CD36 Mediate Phagocytosis of Apoptotic Cells in Immature DCs.
To demonstrate a direct role for αvβ5 in the recognition of apoptotic cells by immature DCs, we performed the phagocytosis FACS® assay in the presence of antibodies specific for αvβ5 (Fig. 7 A). In addition to the blocking observed using the mAb to αvβ5, blocking was also detected when using mAbs to αv, β5, and CD36. Blocking was not observed when isotype-matched mAbs were specific for β1, β3, or the transferrin receptor CD71. Note that control antibodies chosen recognized surface receptors present on the immature DCs (Fig. 7 A, Table 1). mAbs were tested in doses ranging from 10 to 80 μg/ml (data not shown). Maximal inhibition of phagocytosis of apoptotic cells was seen with mAbs specific for CD36, αv, and β5 at 50 μg/ml. The inhibition of phagocytosis of apoptotic cells by DCs was specific. We were unable to block the uptake of red fluorescent latex beads, a control particle, by DCs in the presence of these mAbs (Fig. 7 B). By histogram analysis, DCs phagocytose 1–6 particles per cell. mAbs to αvβ5 or αv did not alter the profile of these histogram plots (data not shown).
Although some inhibition of phagocytosis was observed when using αvβ3 this may be due in part to transdominance and/or the effect on the pool of free αv (35). For example, anti-αvβ3 antibodies suppress the intracellular signaling of the α5β1 integrin (36). Alternatively, αvβ3 and αvβ5 may be working cooperatively in the immature DCs. We therefore tested combinations of anti-αvβ3 and anti-αvβ5 but did not observe an increase in the inhibition of phagocytosis. The low receptor density of αvβ3 on DCs (average mean fluorescence intensity of 7 ± 2; Table 1) also makes it unlikely that this integrin heterodimer is involved in the engulfment of apoptotic cells by immature DCs.
Our data do not exclude a role for other receptors in the phagocytosis of apoptotic cells, e.g., the putative PS receptor or the lectin receptor (5). In fact, other receptors are probably involved, as blocking observed did not exceed 60% even when combinations of all relevant mAbs were tested (data not shown). CD14 is unlikely to be involved in the engulfment of apoptotic cells by DCs, as DCs do not express this receptor (Table 1). In macrophages, phagocytosis of apoptotic cells was inhibited by antibodies to αv, β3, αvβ3, and CD36 but not by antibodies to β1, β5, or αvβ5 (data not shown). This correlates with published data (6, 33).
Discussion
Cross-presentation of antigens to CTLs appears to have two critical features: (a) it is mediated by DCs, and (b) apoptotic cells are the preferred source of antigen (7). The requisite stage of DC development for the acquisition of apoptotic cells is the immature phase. In fact, immature DCs are four to five times more efficient than mature DCs in phagocytosis, a feature that also correlates with their ability to cross-present antigen. This exogenous pathway for class I MHC loading is highly effective: relatively few apoptotic cells (ratio of 1:10 DCs) are needed to charge the DCs as efficiently as the live replicating virus; exposure of 3–12 h is sufficient for generating a peptide–MHC complex that is capable of activating CTLs; and it is relatively indiscriminate, as the cellular source can be allogeneic or xenogeneic cells (7). We believe our earlier studies with mature DCs are explained by the fact that our cell populations were asynchronous and that only by sorting these cells have the differences become apparent. Based on the findings presented here, we suggest that the peripheral tissue DC, exemplified by the immature DC, has an additional important role. It is responsible for phagocytosing cells within tissues that undergo apoptosis (e.g., secondary to viral infection; during normal cell turnover) and migrating to the draining lymph nodes where appropriate T cells are engaged. This pathway may be used for stimulating or tolerizing CTLs and can account for the in vivo observations of cross-priming of tumor and viral antigens (9, 37) and cross-tolerance of self-proteins (10, 38) in their requirement for a bone marrow–derived APC.
A sharp distinction was also demonstrated between immature DCs and macrophages in the handling of apoptotic material. Although macrophages are more efficient at phagocytosing apoptotic cells than immature DCs, they fail to induce virus-specific CTLs (7). In fact, they cannot even generate effective levels of peptide–MHC I complexes. In a short-term assay, influenza-specific CTLs could not kill macrophages cocultured with apoptotic cells. Therefore, macrophages degrade rather than cross-present the ingested apoptotic cells. Our findings probably do not conflict with the report of Bellone et al. (39), as their ‘macrophages' were prepared from bone marrow–derived precursors by culturing the cells in GM-CSF for 7 d. This method is traditionally used to generate DCs from bone marrow (40). We believe that contaminating DCs account for the cross-presentation observed in their studies.
Additional evidence exists that macrophages process apoptotic cells differently from DCs and prevent an immune response. Two groups have demonstrated that phagocytosis of apoptotic cells suppresses a subsequent inflammatory response to LPS stimulation. The macrophage's cytokine profile is skewed toward the synthesis of IL-10, IL-13, and TGF-β, whereas the production of proinflammatory cytokines such as TNF-α, IL-1β, and IL-12 is downmodulated (41, 42). Therefore, the resolution of inflammation is dependent on at least two pathways for removal of apoptotic cells: via (a) macrophages, which subvert and suppress proinflammatory responses, and (b) DCs, which stimulate T cell responses that clear pathogens responsible for the induction of the apoptotic death.
The αvβ5 integrin receptor may be pivotal in the distinctive handling of apoptotic cells by immature DCs versus macrophages, in that its expression is restricted to the former. We suggest that the unique profile of receptors expressed by immature DCs affects trafficking of phagocytosed apoptotic cells, and consequently facilitates cross-presentation. We have previously shown that NH4Cl inhibits the ability of DCs to process antigen derived from apoptotic cells, suggesting that processing in an acidic vesicle (e.g., CIIVs or MIICs) is required (7). Indeed, class I MHC may interact with processed antigens in such a compartment, as MHC I molecules have been described in association with invariant chain (43) and can recycle from the cell surface to class II vesicles (44). Additionally, there may be as yet undescribed routes whereby antigens within vesicles can enter the classical endogenous pathway as described recently for antigens derived from the endoplasmic reticulum (45).
In contrast to our studies, Rubartelli et al. (46) have shown that immature DCs express high levels of the αvβ3 integrin but lack CD36. Their observations are hard to reconcile with data that indicates that αvβ3 and CD36 are both required for the engulfment of apoptotic cells by macrophages (33, 47). Furthermore, although they demonstrated inhibition of phagocytosis by DCs with anti-αv antibodies, they did not present phagocytosis data relevant to β3 or the αvβ3 heterodimer. However, it is possible that a different process is being studied, as phagocytosis in their hands is dependent on late stage Ann V+/PI+ apoptotic cells. Here, we discuss data relevant to the phagocytosis of early apoptotic cells with intact plasma membranes.
αvβ5 and αvβ3 have both been described as important in angiogenesis, cell adhesion, migration, and now in their ability to phagocytose apoptotic cells. αvβ3 is critical in the phagocytosis of apoptotic cells in macrophages, where it acts in a cooperative way with CD36 and thrombospondin, collectively forming a ‘molecular bridge' (47). Recently, it was reported that αvβ5 but not αvβ3 is critical for the engulfment of rod outer segments by CD36+ retinal pigment epithelial cells (48, 49). This phagocytic system is also inhibited by anti-CD36 antibodies, suggesting that αvβ5, like αvβ3, might cooperate with CD36. Taken together with our observations, thrombospondin, or possibly other soluble factors, may serve to bridge CD36, αvβ5, and the apoptotic cell.
Although similarities in function exist, αvβ5 can be distinguished from αvβ3 in its use of various ligands (e.g., vascular endothelial growth factor [VEGF] vs. basic fibroblast growth factor [bFGF]), by the requirements for activation, and by the intracellular signaling pathways (e.g., indirect activation of protein kinase C) (50). Also significant is the fact that the cytoplasmic domains of the two β chains are the portions that show the most considerable diversity (51). Thus, it is possible that the distinct use of the αvβ5 versus αvβ3 integrin receptors might account for the specialized functions of DCs in the route by which apoptotic material is trafficked and presented. In other words, differential expression of αvβ5 may be responsible for the ability of DCs to cross-present antigenic material derived from apoptotic cells, whereas macrophages scavenge and degrade such material.
Acknowledgments
We thank Drs. R. Steinman and R. Darnell for advice and critical review of the manuscript; M. Levin for assistance with figures; Drs. S. Silletti and D. Cheresh for providing us with the β3 and β5 primer pairs; F. Isdell for assistance with the FACSort®; T. de Lange for use of the microscope; and S. Turley for assistance with the immunofluorescence.
Supported by National Institutes of Health (NIH) grants HL-42540 and EY-10967 (to S.F.A. Pearce and R.L. Silverstein); NIH MSTP grant GM-07793 (to M.L. Albert); NIH grant AI-39516, and grants from the SLE foundation (to N. Bhardwaj).
Abbreviations used in this paper
- Ann V
Annexin V
- DC
dendritic cell
- MCM
monocyte-conditioned medium
- PI
propidium iodide
- PS
phosphatidylserine
References
- 1.Fadok VA, Savill JS, Haslett C, Bratton DL, Doherty DE, Campbell PA, Henson PM. Different populations of macrophages use either the vitronectin receptor or the phosphatidylserine receptor to recognize and remove apoptotic cells. J Immunol. 1992;149:4029–4035. [PubMed] [Google Scholar]
- 2.Wyllie AH. Glucocorticoid-induced thymocyte apoptosis is associated with endogenous endonuclease activation. Nature. 1980;284:555–556. doi: 10.1038/284555a0. [DOI] [PubMed] [Google Scholar]
- 3.Godman GC, Miranda AF, Deitch AD, Tanenbaum SW. Action of cytochalasin D on cells of established lines. Zeiosis and movement at the cell surface. J Cell Biochem. 1975;64:644–647. doi: 10.1083/jcb.64.3.644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Casciola-Rosen L, Anhalt G, Rosen A. Autoantigens targeted in systemic lupus erythematosus are clustered in two populations of surface structures on apoptotic keratinocytes. J Exp Med. 1994;179:1317–1330. doi: 10.1084/jem.179.4.1317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Savill J. Phagocytic docking without shocking. Nature. 1998;392:442–443. doi: 10.1038/33025. [DOI] [PubMed] [Google Scholar]
- 6.Savill J. Recognition and phagocytosis of cells undergoing apoptosis. Br Med Bull. 1997;53:491–508. doi: 10.1093/oxfordjournals.bmb.a011626. [DOI] [PubMed] [Google Scholar]
- 7.Albert ML, Sauter B, Bhardwaj N. Dendritic cells acquire antigen from apoptotic cells and induce class I–restricted CTLs. Nature. 1998;392:86–89. doi: 10.1038/32183. [DOI] [PubMed] [Google Scholar]
- 8.Bevan MJ. Cross-priming for a secondary cytotoxic response to minor H antigens with H-2 congenic cells which do not cross-react in the cytotoxic assay. J Exp Med. 1976;143:1283–1288. doi: 10.1084/jem.143.5.1283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Huang AYC, Golumbek P, Ahmadzadeh M, Jaffee E, Pardoll D, Levitsky H. Role of bone marrow-derived cells in presenting MHC class I–restricted tumor antigens. Science. 1994;264:961–965. doi: 10.1126/science.7513904. [DOI] [PubMed] [Google Scholar]
- 10.Kurts C, Heath WR, Carbone FR, Allison J, Miller JFAP, Kosaka H. Constitutive class I–restricted exogenous presentation of self antigens in vivo. J Exp Med. 1996;184:923–930. doi: 10.1084/jem.184.3.923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Banchereau J, Steinman RM. Dendritic cells and the control of immunity. Nature. 1998;392:245–252. doi: 10.1038/32588. [DOI] [PubMed] [Google Scholar]
- 12.Sallusto F, Lanzavecchia A. Dendritic cells use macropinocytosis and the mannose receptor to concentrate antigen in the MHC class II compartment. Downregulation by cytokines and bacterial products. J Exp Med. 1995;182:389–400. doi: 10.1084/jem.182.2.389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Sallusto F, Lanzavecchia A. Efficient presentation of soluble antigen by cultured human dendritic cells is maintained by granulocyte/macrophage colony-stimulating factor plus interleukin 4 and downregulated by tumor necrosis factor α. J Exp Med. 1994;179:1109–1118. doi: 10.1084/jem.179.4.1109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Pierre P, Turley SJ, Gatti E, Hull M, Meltzer J, Mirza A, Inaba K, Steinman RM, Mellman I. Developmental regulation of MHC class II transport in mouse dendritic cells. Nature. 1997;388:787–792. doi: 10.1038/42039. [DOI] [PubMed] [Google Scholar]
- 15.Inaba K, Inaba M, Naito M, Steinman RM. Dendritic cell progenitors phagocytose particulates, including Bacillus Calmette-Guerin organisms, and sensitize mice to mycobacterial antigens in vivo. J Exp Med. 1993;178:479–488. doi: 10.1084/jem.178.2.479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Reis e Sousa C, Stahl PD, Austyn JM. Phagocytosis of antigens by Langerhans cells in vitro. J Exp Med. 1993;178:509–519. doi: 10.1084/jem.178.2.509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Inaba K, Witmer-Pack M, Inaba M, Hathcock KS, Sakuta H, Azuma M, Yagita H, Okumura K, Linsley PS, Ikehara S, et al. The tissue distribution of the B7-2 costimulator in mice: abundant expression on dendritic cells in situ and during maturation in vitro. J Exp Med. 1994;180:1849–1860. doi: 10.1084/jem.180.5.1849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Bender A, Sapp M, Schuler G, Steinman RM, Bhardwaj N. Improved methods for the generation of dendritic cells from nonproliferating progenitors in human blood. J Immunol Methods. 1996;196:121–135. doi: 10.1016/0022-1759(96)00079-8. [DOI] [PubMed] [Google Scholar]
- 19.Romani N, Reider D, Heuer M, Ebner S, Eibl B, Niederwieser D, Schuler G. Generation of mature dendritic cells from human blood: an improved method with special regard to clinical applicability. J Immunol Methods. 1996;196:137–151. doi: 10.1016/0022-1759(96)00078-6. [DOI] [PubMed] [Google Scholar]
- 20.Sallusto F, Nicolo C, De Maria R, Corinti S, Testi R. Ceramide inhibits antigen uptake and presentation by dendritic cells. J Exp Med. 1996;184:2411–2416. doi: 10.1084/jem.184.6.2411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Caux C, Massacrier C, Vanbervliet B, Dubois B, Van Kooten C, Durand I, Banchereau J. Activation of human dendritic cells through CD40 cross-linking. J Exp Med. 1994;180:1263–1272. doi: 10.1084/jem.180.4.1263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Rieser C, Bock G, Klocker H, Bartsch G, Thurnher M. Prostaglandin E2 and tumor necrosis factor α cooperate to activate human dendritic cells: synergistic activation of interleukin 12 production. J Exp Med. 1997;186:1603–1608. doi: 10.1084/jem.186.9.1603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hess KL, Babcock GF, Askew DS, Cook-Mills JM. A novel flow cytometric method for quantifying phagocytosis of apoptotic cells. Cytometry. 1998;27:145–152. [PMC free article] [PubMed] [Google Scholar]
- 24.Bhardwaj N, Bender A, Gonzalez N, Bui LK, Garrett MC, Steinman RM. Influenza virus–infected dendritic cells stimulate strong proliferative and cytolytic responses from human CD8+T cells. J Clin Invest. 1994;94:797–807. doi: 10.1172/JCI117399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Fesq H, Bacher M, Nain M, Gemsa D. Programmed cell death (apoptosis) in human monocytes infected by influenza virus. Immunobiology. 1994;190:175–182. doi: 10.1016/S0171-2985(11)80292-5. [DOI] [PubMed] [Google Scholar]
- 26.Hofmann P, Sprenger H, Kaufmann A, Bender A, Hasse C, Nain M, Gemsa D. Susceptibility of mononuclear phagocytes to influenza A virus infection and possible role in the antiviral response. J Leukocyte Biol. 1997;612:408–414. doi: 10.1002/jlb.61.4.408. [DOI] [PubMed] [Google Scholar]
- 27.Steinkamp JA, Wilson JS, Saunders GC, Stewart CC. Phagocytosis: flow cytometric quantitation with fluorescent microspheres. Science. 1981;215:64–66. doi: 10.1126/science.7053559. [DOI] [PubMed] [Google Scholar]
- 28.Gotch F, Rothbard J, Howland K, Townsend A, McMichael A. Cytotoxic T lymphocytes recognize a fragment of influenza virus matrix protein in association with HLA-A2. Nature. 1987;326:881–882. doi: 10.1038/326881a0. [DOI] [PubMed] [Google Scholar]
- 29.Silverstein RL, Baird M, Lo S, Yesner LM. Sense and antisense cDNA transfection of CD36 in melanoma cells. Role of CD36 as a thrombospondin receptor. J Biol Chem. 1992;267:16607–16612. [PubMed] [Google Scholar]
- 30.Zhou L-J, Tedder TF. Human blood dendritic cells selectively express CD83, a member of the immunoglobulin superfamily. J Immunol. 1995;154:3821–3835. [PubMed] [Google Scholar]
- 31.De Togni P, Goellner J, Ruddle NH, Streeter PR, Fick A, Mariathasan S, Smith SC, Carlson R, Shornick LP, Strauss-Schoenberger J, et al. Abnormal development of peripheral lymphoid organs in mice deficient in lymphotoxin. Science. 1994;264:703–707. doi: 10.1126/science.8171322. [DOI] [PubMed] [Google Scholar]
- 32.Savill JS, Dransfield I, Hogg N, Haslett C. Vitronectin receptor-mediated phagocytosis of cells undergoing apoptosis. Nature. 1990;343:170–173. doi: 10.1038/343170a0. [DOI] [PubMed] [Google Scholar]
- 33.Ren Y, Silverstein RL, Allen J, Savill J. CD36 gene transfer confers capacity for phagocytosis of cells undergoing apoptosis. J Exp Med. 1995;181:1857. doi: 10.1084/jem.181.5.1857. (Abstr.) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Devitt A, Moffatt OD, Raykundalia C, Capra JD, Simmons DL, Gregory CD. Human CD14 mediates recognition and phagocytosis of apoptopic cells. Nature. 1998;392:505–509. doi: 10.1038/33169. [DOI] [PubMed] [Google Scholar]
- 35.Diaz-Gonzalez F, Forsyth J, Steiner B, Ginsberg MH. Trans-dominant inhibition of integrin function. Mol Biol Cell. 1996;7:1939–1951. doi: 10.1091/mbc.7.12.1939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Blystone SD, Graham IL, Lindberg FP, Brown EJ. Integrin alpha v beta 3 differentially regulates adhesive and phagocytic function of the fibronectin receptor alpha 5 beta 1. J Cell Biol. 1994;127:1129–1137. doi: 10.1083/jcb.127.4.1129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Carbone FR, Moore MW, Sheil JM, Bevan MJ. Induction of cytotoxic T lymphocytes by primary in vitro stimulation with peptides. J Exp Med. 1988;167:1767–1779. doi: 10.1084/jem.167.6.1767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Heath WR, Kurts C, Miller JFAP, Carbone F. Cross-tolerance: a pathway for inducing tolerance to peripheral tissue antigens. J Exp Med. 1998;187:1549–1553. doi: 10.1084/jem.187.10.1549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Bellone M, Iezzi G, Rovere P, Galati G, Ronchetti A, Protti MP, Davoust J, Rugarli C, Manfredi AA. Processing of engulfed apoptotic bodies yields T cell epitopes. J Immunol. 1997;159:5391–5399. [PubMed] [Google Scholar]
- 40.Inaba K, Inaba M, Romani N, Aya H, Deguchi M, Ikehara S, Muramatsu S, Steinman RM. Generation of large numbers of dendritic cells from mouse bone marrow cultures supplemented with granulocyte/macrophage colony-stimulating factor. J Exp Med. 1992;176:1693–1702. doi: 10.1084/jem.176.6.1693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Voll RE, Herrmann M, Roth EA, Stach C, Kalden JR. Immunosuppressive effects of apoptotic cells. Nature. 1997;390:350–351. doi: 10.1038/37022. [DOI] [PubMed] [Google Scholar]
- 42.Fadok VA, Bratton DL, Konowal A, Freed PW, Westcott JY, Henson PM. Macrophages that have ingested apoptosis cells in vitro inhibit proinflammatory cytokine production through autocrine/paracrine mechanisms involving TGF-β, PGE2, and PAF. J Clin Invest. 1998;101:890–898. doi: 10.1172/JCI1112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Sugita M, Brenner MB. Association of the invariant chain with major histocompatibility complex class I molecules directs trafficking to endocytic compartments. J Biol Chem. 1995;270:1443–1448. doi: 10.1074/jbc.270.3.1443. [DOI] [PubMed] [Google Scholar]
- 44.Jondal M, Schirmbeck R, Reimann J. MHC class-I restricted CTL responses to exogenous antigens. Immunity. 1996;5:295–302. doi: 10.1016/s1074-7613(00)80255-1. [DOI] [PubMed] [Google Scholar]
- 45.Snyder HL, Bacik I, Bennink JR, Kearns G, Behrens TW, Bachi T, Orlowski M, Yewdell JW. Two novel routes of transporter associated with antigen processing (TAP)-independent major histocompatibility complex class I antigen processing. J Exp Med. 1997;186:1087–1098. doi: 10.1084/jem.186.7.1087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Rubartelli A, Poggi A, Zocchi MR. The selective engulfment of apoptotic bodies by dendritic cells is mediated by the αvβ3 integrin and requires intracellular and extracellular calcium. Eur J Immunol. 1997;27:1893–1900. doi: 10.1002/eji.1830270812. [DOI] [PubMed] [Google Scholar]
- 47.Savill J, Hogg N, Ren Y, Hasslet C. Thrombospondin cooperates with CD36 and the vitronectin receptor in macrophage recognition of neutrophils undergoing apoptosis. J Clin Invest. 1992;90:1513–1522. doi: 10.1172/JCI116019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Ryeom SW, Sparrow JR, Silverstein RL. CD36 participates in the phagocytosis of rod outer segments by retinal pigment epithelium. J Cell Sci. 1996;109:387–395. doi: 10.1242/jcs.109.2.387. [DOI] [PubMed] [Google Scholar]
- 49.Finnemann SC, Bonilha VL, Marmorstein AD, Rodriguez-Boulan E. Phagocytosis of rod outer segments by retinal pigment epithelial cells requires alpha(v) beta5 integrin for binding but not for internalization. Proc Natl Acad Sci USA. 1997;94:12932–12937. doi: 10.1073/pnas.94.24.12932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Friedlander M, Brooks PC, Shaffer RW, Kincaid CM, Varner JA, Cheresh DA. Definition of two angiogenic pathways by distinct αvintegrins. Science. 1995;270:1500–1502. doi: 10.1126/science.270.5241.1500. [DOI] [PubMed] [Google Scholar]
- 51.McClean JW, Vestal DJ, Cheresh DA, Bodary SC. cDNA sequence of the human integrin β5subunit. J Biol Chem. 1990;265:17126–17131. [PubMed] [Google Scholar]