

Abstract
We report that B cell–activating factor of the tumor necrosis factor (TNF) family (BAFF) is expressed in the normal human brain at ∼10% of that in lymphatic tissues (tonsils and adenoids) and is produced by astrocytes. BAFF was regularly detected by enzyme-linked immunosorbent assay in brain tissue lysates and in normal spinal fluid, and in astrocytes by double fluorescence microscopy. Cultured human astrocytes secreted functionally active BAFF after stimulation with interferon-γ and TNF-α via a furin-like protease-dependent pathway. BAFF secretion per cell was manifold higher in activated astrocytes than in monocytes and macrophages. We studied brain lesions with B cell components, and found that in multiple sclerosis plaques, BAFF expression was strongly up-regulated to levels observed in lymphatic tissues. BAFF was localized in astrocytes close to BAFF-R–expressing immune cells. BAFF receptors were strongly expressed in situ in primary central nervous system (CNS) lymphomas. This paper identifies astrocytes as a nonimmune source of BAFF. CNS-produced BAFF may support B cell survival in inflammatory diseases and primary B cell lymphoma.
BAFF (B cell–activating factor of the TNF family; also named BLyS) plays a crucial role in B cell development, survival, and production of Ig (1). Studies on BAFF transgenic mice and patients with rheumatoid diseases indicate that overexpression of BAFF is linked to the development of autoimmune pathology (1).
Until now, myeloid cells, namely monocytes, macrophages, dendritic cells (2), and neutrophils (3), as well as unidentified nonbone marrow–derived radioresistant cells (4), have been considered as sources of BAFF.
In different immunopathologies, the brain provides a fostering environment to B cells: primary central nervous system (CNS) lymphomas are usually of B cell origin (5). The cerebrospinal fluid (CSF) of patients with chronic infectious and autoimmune diseases of the CNS typically contains remarkably stable oligoclonal Ig bands (6). In the CNS of multiple sclerosis (MS) patients clonally expanded B cells and plasma cells persist (7–9), ectopic B cell follicles develop in the meninges of patients with secondary progressive MS (10), and B cell differentiation may be recapitulated in the CNS of MS patients (11).
Our paper establishes astrocytes as BAFF producers and suggests that CNS-derived BAFF might foster B cell survival in MS and primary CNS lymphoma.
Results and Discussion
BAFF in the normal CNS
BAFF transcription was analyzed in 17 different normal brain samples from four adults including gray and white matter of the cerebrum and the cerebellum. Similar levels of transcripts were detected in each specimen (Figs. 1 and 2). The level of BAFF mRNA in the normal brain was almost 10% of that detected in tonsils or adenoids (Fig. 2). Similar results were obtained with PPIA (Fig. 2) or with GAPDH (not depicted) as housekeeping gene.
Figure 1.

BAFF transcripts in brain specimens and lymphatic tissue. RT-PCR detected full-length BAFF and the splice variant ΔBAFF in human CNS and lymphatic tissue. Genomic DNA (gDNA) and H2O served as controls.
Figure 2.

BAFF expression in normal CNS, MS lesions, and primary CNS lymphomas. Expression level of individual tissue samples was determined by quantitative PCR, and the median of each group is indicated. Mann-Whitney rank sum test was used for statistical analysis. Note that the BAFF levels in normal CNS and primary CNS lymphoma were not zero, but clearly detectable in each specimen with a mean of 0.034% PPIA.
The transcript level of BAFF was quantified in a series of other tissues. BAFF expression was detectable in heart, skeletal muscle, pancreas, kidney, skin with adipose tissue, and liver at similar levels as in the CNS (i.e., in the range of 0.03–0.1% PPIA). In the lung, the BAFF levels were slightly higher, reaching 0.18% PPIA. Although the present paper identifies astrocytes as BAFF producers in the CNS (Figs. 3 and 4), further studies might establish whether in other tissues resident macrophages (particularly in lung) or parenchymal cells are responsible for local BAFF transcription.
Figure 3.
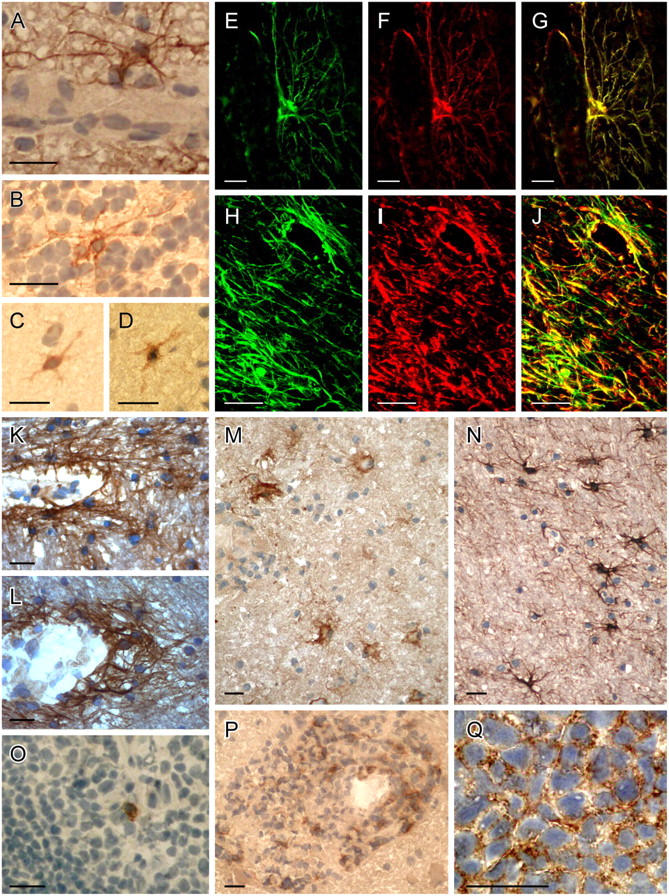
BAFF and BAFF-R–expressing cells in the normal CNS, MS, and primary CNS lymphoma. (A–D) BAFF in the normal CNS. (A) Molecular stratum and (B) granular stratum were from the cerebellum, (C and D) from the cerebrum. BAFF immunolocalized to scattered astrocytes in the perivascular area (A) and in the parenchyma (B–D). Similar staining patterns were observed with the two different BAFF-specific mAbs B1A3.2 (A–C) and A9C9.1 (D). (E–N) BAFF-specific astrocytes in different MS lesions. (E–G and M) Patient 1 (acute lesion); (H–L) patient 2 (chronic lesions); and (N) patient 3 (chronic lesion). BAFF-positive astrocytes are detected in the perivascular area (H–L) and in the parenchyma (H–J, M, and N). Double stainings are shown for GFAP (F and I, red), BAFF (E and H, green), and the overlays (G and J). Confocal microscopy (H–J) shows that BAFF-positive astrocytes form a network in the parenchyma and in the perivascular area. The two different BAFF-specific mAbs give similar staining patterns (K and L). (M) BAFF on astrocytes is demonstrated in the vicinity of infiltrating immune cells. The mAb B1A3.2 (E, G, H, J, K, M, N) and the mAb A9C9.1 (L) were applied. (O) A scattered cell in the germinal center of a tonsil is stained for BAFF (mAb B1A3.2). (P) BAFF-R expression on infiltrating immune cells within an acute MS lesion (patient 1). (Q) BAFF-R on the densely packed tumor cells on a primary CNS lymphoma. Sections were developed with diaminobenzidine that resulted in a brown color; nuclei were stained blue with hematoxylin (A–D and K–Q). Bars, 20 μm.
Figure 4.

BAFF production by cultured astrocytes. (A) RT-PCR identified full-length BAFF and the splice variant ΔBAFF. To detect the constitutive expression of both isoforms and the induction by cytokine stimulation (25 h), two different cycle numbers were used as indicated. (B and C) Regulation by inflammatory cytokines. From the same stimulated astrocytes, RNA and cell culture supernatant were analyzed after 48 h by quantitative PCR (B) and ELISA (C). Error bars represent the SD. (D) Comparison of BAFF secretion by astrocytes, monocytes, and macrophages. After stimulation for 48 h, cell culture supernatants were analyzed by ELISA. The means of two experiments with monocytes, three with macrophages, and two with astrocytes are shown. Error bars represent SEM. Astrocyte stimulation with LPS + IFN-γ was not determined (ND) because a series of experiments had shown that these cultures do not respond to LPS, in agreement with the absence of myeloid cells in these cultures. (E) Functional activity of astrocyte-derived BAFF in a B cell proliferation assay. The concentrated astrocyte supernatant was added at a final dilution of 1:10 to the B cell culture. Medium processed in the same way as the astrocyte supernatant served as negative control and recombinant BAFF served as positive control. BAFF-R:Fc was used to block the BAFF effect. Error bars indicate the SD.
An alternative splice variant lacking exon 3 (ΔBAFF) has been identified in mouse and human myeloid cells (12). We detected ΔBAFF, which was less abundant than the full-length isoform, in all brain specimens as well as in lymphatic tissue (Fig. 1).
BAFF-expressing cells had the morphology of astrocytes. They were found in three normal brains in the parenchyma and in the perivascular area both in the gray and in the white matter (Fig. 3, A–D). BAFF+ cells displayed long processes that extended to the perivascular area (Fig. 3 A).
In tonsils and adenoids, BAFF was detected on a few scattered cells without a clear distribution pattern. The frequency of BAFF-labeled cells differed considerably in different tonsil specimens. The labeled cells appeared in germinal centers (Fig. 3 O), T cell areas, and in some specimens also in the connective tissue (not depicted).
BAFF was detected in brain tissue by ELISA at a concentration of ∼140 pg/mg protein lysate, whereas ∼5 ng/mg was observed in tonsils/adenoids. The amount of BAFF detected in normal brain tissue cannot be explained by blood contamination because in lysates of whole blood the BAFF concentration was ∼10-fold lower than in brain tissue lysates.
10 paired noninflammatory serum/CSF samples were analyzed by ELISA. BAFF concentrations ranged from 153 to 845 pg/ml in CSF (mean ± SD: 483 ± 263 pg/ml) and from 2,257 to 6,197 pg/ml in serum (4,268 ± 1,295 pg/ml). No significant correlation was found between BAFF values in serum and CSF (R = 0.35, P = 0.31). The relatively high level of BAFF in CSF (on average 10% of the serum concentration) and the lack of correlation between serum and CSF content suggest that the BAFF present in the CSF is at least in part produced locally.
The functional relevance of the constitutive BAFF production in the CNS is presently unknown. Although no neurological deficits of BAFF-deficient mice have been described yet, this does not exclude subtle functional changes in the CNS.
BAFF production by cultured human astrocytes
Cultured human astrocytes constitutively contained a low level of BAFF transcripts (Fig. 4 A), which was increased by IFN-γ, to a lesser extent by TNF-α or IL-1β, but not by IL-10 (Fig. 4 B), GM-CSF, or IL-4 (not depicted). The strongest induction was seen with a combination of IFN-γ and TNF-α (Fig. 4 B). The amount of BAFF secreted by astrocytes paralleled the induction of transcription (Fig. 4 C). An inhibitor of furin-like proteases, decanoyl-Arg-Val-Lys-Arg-chloromethylketone, reduced the amount of secreted BAFF by 96% (unpublished data), indicating that astrocytes use furin-like proteases to release BAFF as do granulocytes (3). The biological activity of the astrocyte-produced BAFF was verified in a B cell proliferation assay (Fig. 4 E).
The level of BAFF released per cell after stimulation was much greater in astrocyte cultures than by activated monocytes (50 times) or macrophages (5 times; Fig. 4 D), which released in our study similar BAFF levels as described previously (2). To exclude the possibility that the BAFF levels in monocyte and macrophage cultures were artificially lowered by paracrine BAFF consumption of B cells, PBMCs were depleted of B cells before negatively selecting monocytes. FACS analysis showed that, in the purified monocyte cultures, essentially no BAFF binding cells could be detected.
BAFF and BAFF receptors in MS lesions
The transcript level of BAFF was ∼10-fold higher in MS lesions as compared with normal CNS (P < 0.01), reaching approximately the same level as in lymphatic tissues (Fig. 2).
Immunostaining for BAFF was performed on 11 brain specimens from four MS patients. In each specimen, BAFF was immunolocalized on astrocytes (Fig. 3, E–N). In acute MS lesions, BAFF localized to astrocytes outside of the inflammatory cuffs (Fig. 3 M). In chronic MS lesions, numerous BAFF+ cells with the morphology of astrocytes were detected in both the perivascular area and in the parenchyma (Fig. 3, H–L and N). Double labeling for glial fibrillary acidic protein (GFAP) and BAFF identified BAFF-expressing cells as astrocytes in both acute (Fig. 3, E–G) and chronic MS lesions (Fig. 3, H–J). Confocal analysis revealed a dense network of BAFF+ astrocytes in the perivascular area and in the parenchyma of MS lesions (Fig. 3, H–J). Although the most sensitive staining was observed with the BAFF-specific mAb B1A3.2 (Fig. 3, A–C, E, G, H, J, K, M, and N), a similar astrocytic staining was observed with the BAFF-specific mAb A9C9.1 in normal brain (Fig. 3 D) and MS lesions (Fig. 3 L). BAFF-R was detected on a subset of inflammatory cells in the vicinity of BAFF-expressing astrocytes, mainly in the perivascular area (Fig. 3 P).
Our histological observation of enhanced BAFF expression on astrocytes in MS lesions is in harmony with the in vitro induction of BAFF on astrocytes by inflammatory cytokines. However, it should be noted that the histological detection of BAFF is presumably insensitive. For example, in mice, BAFF staining was unsuccessful (13), and in tonsils we detected BAFF only on a few scattered cells (Fig. 3 O) despite its abundant transcription. This indicates that, by immunohistochemical staining, the number and cell types expressing BAFF might be underestimated. Because macrophages are abundant in MS lesions and produce BAFF in vitro, the elevated BAFF expression in MS lesions might be due to BAFF produced by both astrocytes and macrophages. Our in vitro findings that appropriately stimulated astrocytes produce five times more BAFF per cell than highly activated macrophages might explain why in MS lesions BAFF is immunolocalized preferentially to astrocytes.
Our findings suggest the following scenario: BAFF produced by astrocytes promotes the survival of BAFF-R–expressing B cells in MS brains, allowing the persistence and clonal expansion of B cells in the CNS (7–9, 11). The oligoclonal Ig in the CSF are remarkably stable for many years (6), consistent with the long-term survival of local plasma cells. Recently, it has been shown that BCMA, a receptor for BAFF and APRIL, is critical for the long-term survival of plasma cells (14). Locally produced BAFF might contribute to plasma cell survival in the CNS. Indeed, our quantitative PCR indicated that BCMA was up-regulated in MS brain specimens (Fig. 5). Our conclusions are consistent with two animal studies. First, the normal brain microenvironment supports development of antigen-directed humoral immunity in rodents (15). Second, increased BAFF gene expression was detected in the CNS of animals with experimental autoimmune encephalomyelitis, although in this paper, neither BAFF-producing nor BAFF receptor–expressing cells were described previously (16).
Figure 5.
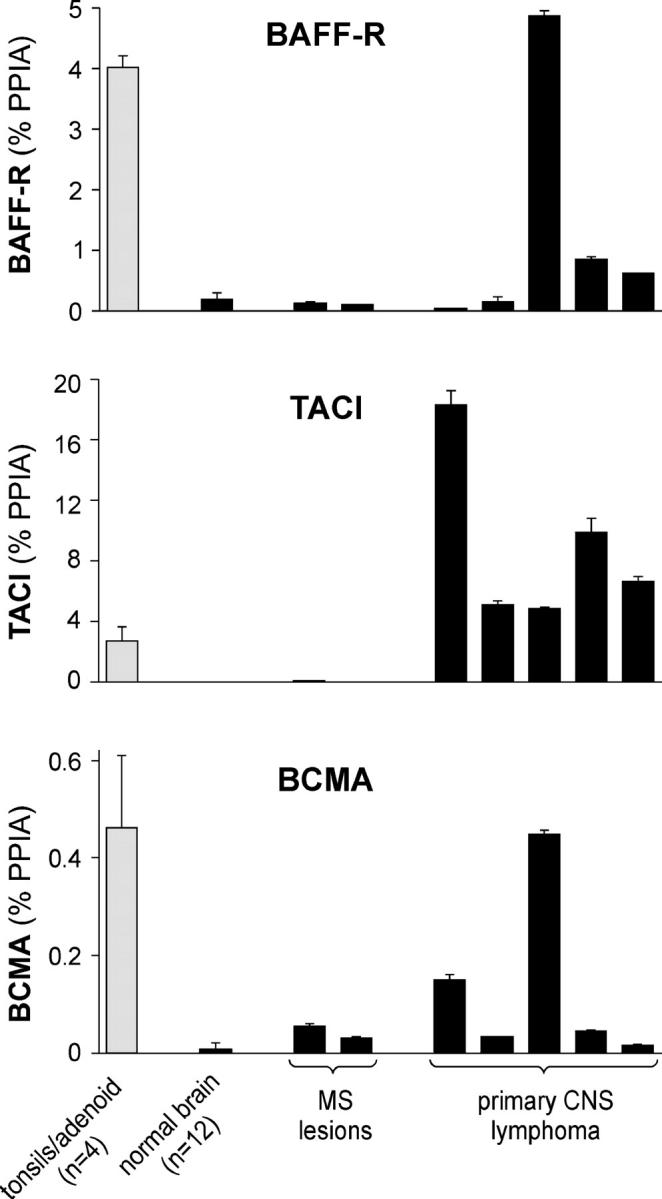
BAFF receptors in normal CNS, MS lesions, and primary CNS lymphomas. The expression level of BAFF-R, TACI, and BCMA was determined by quantitative PCR and is indicated as the percentage of the housekeeping gene PPIA. The mean values of 4 specimens from adenoids/tonsils and of 12 specimens from normal CNS are shown. Individual expression levels are shown for the analyzed pathological tissue specimens from MS lesions or primary CNS lymphomas. Error bars indicate the SD.
Local BAFF production within MS lesions might also promote T cell activation because BAFF has also been recognized as a costimulator for T cells (17). The elevated expression of BAFF in MS lesions was not reflected by an increase of BAFF in the CSF (reference 18 and unpublished data). This resembles observations with CCL-2/MCP-1. This chemokine is constitutively present in the CSF and it is abundantly expressed in MS lesions, but its concentration in the CSF is even reduced in MS (19). Presumably, the ratio of production and consumption by inflammatory cells determines the CSF level.
BAFF and BAFF receptors in primary CNS lymphomas
Primary CNS lymphomas are extranodal lymphomas that are confined to the CNS in the absence of systemic disease. Primary CNS lymphomas are typically of B cell origin for still unknown reasons (5).
Although BAFF receptors had not been analyzed in primary CNS lymphomas before, we report that in these CNS tumors, the transcript levels of all three BAFF receptors greatly exceeded that in normal brain and reached the same level as in lymphatic tissue (Fig. 5). Immunohistochemical analysis detected BAFF-R in primary CNS lymphomas rather homogeneously (Fig. 3 Q). A clear BAFF-R staining was observed on tumor cells in six out of seven specimens of the CNS lymphomas. Expression of the three BAFF receptors in individual tumor samples was heterogeneous (Fig. 5). The PCR data paralleled the histological data. The tumor sample with the lowest BAFF-R transcript level was the only one of the seven specimens without a clear BAFF-R staining. This tumor showed the highest TACI expression and the second highest BCMA level of the specimens analyzed (Fig. 5).
BAFF was transcribed in each of the five analyzed tumor specimens, in three of them at a similar level as in normal CNS, and in two of them at an elevated level (Fig. 2). The CNS lymphoma tissue we investigated in this paper did not contain parenchyma surrounding the tumors. However, based on our findings in normal brain and in inflammatory MS lesions, it seems plausible to assume that BAFF released by resident astrocytes supports the survival of BAFF-receptor–expressing B cell lymphomas, which are otherwise found in lymph nodes where BAFF is abundant. Primary CNS lymphomas reside at least partially behind an intact blood–brain barrier (5), implicating the importance of the CNS-derived BAFF as a survival factor. This local supply of BAFF together with the shielding of the brain microenvironment from the immune system might provide a favorable basis for tumor development. Recent animal studies are consistent with this conclusion; in the absence of TNF-α, BAFF-overexpressing mice develop lymphomas (20), showing that BAFF in combination with a reduced tumor immunosurveillance facilitates lymphoma development. Our findings have therapeutic implications by suggesting that it might be beneficial to inhibit the local availability of BAFF in and around the CNS lymphoma.
In conclusion, we have extended the spectrum of BAFF sources to astrocytes and demonstrate that BAFF is constitutively present in the CNS. In MS lesions, BAFF expression was elevated to levels of lymphatic tissues. Local BAFF production in the CNS might promote B cell survival in MS and primary CNS lymphoma.
Materials and methods
Tissue specimens, CSF, and serum
Control tissue specimens were obtained from autopsy material of persons who had died of acute accidents: normal brain including gray and white matter of the cerebrum and cerebellum (17 tissue blocks from four adult persons), heart, skeletal muscle, lung, liver, skin with adipose tissue, pancreas, and kidney. Tonsils and adenoids were obtained after operations. Eight brain lesions of six MS patients were examined by RT-PCR. One case was published (case 2 in reference 21), the other specimens were obtained from the Institute of Neurology in London (courtesy of L. Cuzner and J. Newcombe). They were classified as acute based on the presence of oil red O positive macrophages and perivascular infiltrates. Frozen brain specimens from 11 blocks of four MS patients were examined immunohistologically: two blocks from an acute lesion (patient 1; case 2 in reference 21), six blocks containing chronic active and chronic inactive lesions (patient 2), a chronic active plaque (patient 3), and two blocks containing chronic active and chronic inactive lesions (patient 4; MS 79 in reference 10). Seven surgical specimens from six primary CNS lymphomas (diffuse large B cell lymphoma, tumor cell content >80%) were analyzed by immunohistochemistry, and five by RT-PCR. Some of these tumors contained an extensive GFAP positive gliosis. Paired CSF and serum samples from 10 patients without inflammatory neurological disease were investigated. The study was approved by the ethics committee of the University of Munich.
Cell culture
Human astrocytes of embryonic origin were devoid of microglial cells or macrophages (22). These cells were stimulated with 10 or 25 ng/ml TNF-α, 100 U/ml IFN-γ (Roche), 50 ng/ml IL-1β, and 100 ng/ml IL-10 (R&D Systems).
For the analysis of the biological activity of secreted BAFF, astrocytes were switched to serum-free medium (Pan-Biotech) and stimulated with IFN-γ + TNF-α for 3 d. The supernatant was concentrated ∼200-fold and a B cell proliferation assay was done essentially as described previously (17).
Monocytes were isolated by negative selection from CD19-depleted PBMCs (both obtained from Miltenyi Biotec), differentiated to macrophages with 20 ng/ml M-CSF for a period of 9–11 d (2), and stimulated in new medium. BAFF-binding cells were identified by myc-BAFF, biotinylated anti-myc mAb, and streptavidin-PE. Cells were stimulated with 300 U/ml IFN-γ + 25 ng/ml TNF-α or 300 ng/ml LPS for 48 h.
PCR
Quantitative PCR and the identification of BAFF isoforms was performed as described in the supplemental material (available at http://www.jem.org/cgi/content/full/jem.20041674/DC1).
BAFF ELISA
The anti–human BAFF mAb clone B4H7.2 was used for coating, the biotin-labeled anti–human BAFF clone A9C9.1 was used for detection, and recombinant human BAFF was used as a standard (all obtained from Biogen Idec Inc.). To determine whether this ELISA can be used to detect BAFF content in tissue lysates, 400 pg/ml BAFF was spiked in a 1:50 and 1:20 dilution of the RIPA lysis buffer; >95% of the spiked BAFF was detected. Protein lysates of brain specimens and whole blood were obtained using RIPA buffer and measured in a dilution of at least 1:40.
Immunohistochemistry
Mabs recognizing BAFF (B1A3.2 and A9C9.1), BAFF-R (9–1; 5–10 μg/ml), and IgG1 control were used. The mAb B1A3.2 revealed a good staining with 20 μg/ml, the strongest BAFF-expressing cells could still be seen with 10 μg/ml. mAb A9C9.1 was used at 35 μg/ml. Stainings were made on PFA or formalin-fixed cryosections with the peroxidase antiperoxidase method or the avidin–biotin complex system (DakoCytomation). The autoptic specimens of patient 4, which had been fixed in PFA before freezing, were processed as described previously (10). For double immunofluorescence staining, mouse anti-BAFF mAb B1A3.2 and rabbit anti-GFAP were applied simultaneously, followed by donkey anti–mouse Alexa Fluor 488 or donkey anti–mouse biotin plus Cy2-conjugated streptavidin, and donkey anti–rabbit Alexa Fluor 594. Confocal images were taken on a TCS NT laser scanning microscope (Leica).
Online supplemental material.
Materials and methods for the quantitative PCR and the identification of BAFF isoforms. Online supplemental material is available at http://www.jem.org/cgi/content/full/jem.20041674/DC1.
Acknowledgments
We thank D. Zech for technical assistance; J. Benson for help in editing the paper; and D. Gong, K. Strauch, and T. Cachero for the BAFF and anti-BAFF reagents. We are grateful to Dr. B. Hemmer for providing CSF samples and Dr. I. Sinicina for valuable support. We thank Drs. R. Voltz and D. Jenne for comments on the paper. Autoptic brain tissue was supplied by the UK Multiple Sclerosis Tissue Bank. We are also grateful to Dr. M. Neumann and Prof. H. Kretzschmar (Brain-Net), Dr. C. Stadelmann and Prof. W. Brück, and Dr. J. Newcombe and Dr. L. Cuzner for providing MS tissue.
This work was supported by the DFG (SFB 571 and GRK 688), the Gemeinnützige Hertie-Foundation, and the Verein zur Therapieforschung für MS Kranke. The Institute for Clinical Neuroimmunology is supported by the Hermann and Lilly Schilling Foundation.
S.L. Kalled and D. Hess are employed by and shareholders of Biogen Idec, Inc. The authors have no other potential conflicting financial interests.
References
- 1.Mackay, F., and S.G. Tangye. 2004. The role of the BAFF/APRIL system in B cell homeostasis and lymphoid cancers. Curr. Opin. Pharmacol. 4:347–354. [DOI] [PubMed] [Google Scholar]
- 2.Nardelli, B., O. Belvedere, V. Roschke, P.A. Moore, H.S. Olsen, T.S. Migone, S. Sosnovtseva, J.A. Carrell, P. Feng, J.G. Giri, and D.M. Hilbert. 2001. Synthesis and release of B-lymphocyte stimulator from myeloid cells. Blood. 97:198–204. [DOI] [PubMed] [Google Scholar]
- 3.Scapini, P., B. Nardelli, G. Nadali, F. Calzetti, G. Pizzolo, C. Montecucco, and M.A. Cassatella. 2003. G-CSF–stimulated neutrophils are a prominent source of functional BLyS. J. Exp. Med. 197:297–302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Gorelik, L., K. Gilbride, M. Dobles, S.L. Kalled, D. Zandman, and M.L. Scott. 2003. Normal B cell homeostasis requires B cell activation factor production by radiation-resistant cells. J. Exp. Med. 198:937–945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.DeAngelis, L.M. 1999. Primary central nervous system lymphoma. J. Neurol. Neurosurg. Psychiatry. 66:699–701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Walsh, M.J., and W.W. Tourtellotte. 1986. Temporal invariance and clonal uniformity of brain and cerebrospinal IgG, IgA, and IgM in multiple sclerosis. J. Exp. Med. 163:41–53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Baranzini, S.E., M.C. Jeong, C. Butunoi, R.S. Murray, C.C. Bernard, and J.R. Oksenberg. 1999. B cell repertoire diversity and clonal expansion in multiple sclerosis brain lesions. J. Immunol. 163:5133–5144. [PubMed] [Google Scholar]
- 8.Qin, Y., P. Duquette, Y. Zhang, P. Talbot, R. Poole, and J. Antel. 1998. Clonal expansion and somatic hypermutation of V(H) genes of B cells from cerebrospinal fluid in multiple sclerosis. J. Clin. Invest. 102:1045–1050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Owens, G.P., A.M. Ritchie, M.P. Burgoon, R.A. Williamson, J.R. Corboy, and D.H. Gilden. 2003. Single-cell repertoire analysis demonstrates that clonal expansion is a prominent feature of the B cell response in multiple sclerosis cerebrospinal fluid. J. Immunol. 171:2725–2733. [DOI] [PubMed] [Google Scholar]
- 10.Serafini, B., B. Rosicarelli, R. Magliozzi, W. Stille, and F. Aloisi. 2004. Detection of ectopic B-cell follicles with germinal centers in the meninges of patients with secondary progressive multiple sclerosis. Brain Pathol. 14:164–174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Corcione, A., S. Casazza, E. Ferretti, D. Giunti, E. Zappia, A. Pistorio, C. Gambini, G.L. Mancardi, A. Uccelli, and V. Pistoia. 2004. Recapitulation of B cell differentiation in the central nervous system of patients with multiple sclerosis. Proc. Natl. Acad. Sci. USA. 101:11064–11069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Gavin, A.L., D. Ait-Azzouzene, C.F. Ware, and D. Nemazee. 2003. delta BAFF, an alternate splice isoform that regulates receptor binding and biopresentation of the B cell survival cytokine, BAFF. J. Biol. Chem. 278:33127–33133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lesley, R., Y. Xu, S.L. Kalled, D.M. Hess, S.R. Schwab, H.B. Shu, and J.G. Cyster. 2004. Reduced competitiveness of autoantigen-engaged B cells due to increased dependence on BAFF. Immunity. 20:441–453. [DOI] [PubMed] [Google Scholar]
- 14.O'Connor, B.P., V.S. Raman, L.D. Erickson, W.J. Cook, L.K. Weaver, C. Ahonen, L.L. Lin, G.T. Mantchev, R.J. Bram, and R.J. Noelle. 2004. BCMA is essential for the survival of long-lived bone marrow plasma cells. J. Exp. Med. 199:91–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Knopf, P.M., C.J. Harling-Berg, H.F. Cserr, D. Basu, E.J. Sirulnick, S.C. Nolan, J.T. Park, G. Keir, E.J. Thompson, and W.F. Hickey. 1998. Antigen-dependent intrathecal antibody synthesis in the normal rat brain: tissue entry and local retention of antigen-specific B cells. J. Immunol. 161:692–701. [PubMed] [Google Scholar]
- 16.Magliozzi, R., S. Columba-Cabezas, B. Serafini, and F. Aloisi. 2004. Intracerebral expression of CXCL13 and BAFF is accompanied by formation of lymphoid follicle-like structures in the meninges of mice with relapsing experimental autoimmune encephalomyelitis. J. Neuroimmunol. 148:11–23. [DOI] [PubMed] [Google Scholar]
- 17.Ng, L.G., A.P. Sutherland, R. Newton, F. Qian, T.G. Cachero, M.L. Scott, J.S. Thompson, J. Wheway, T. Chtanova, J. Groom, et al. 2004. B cell-activating factor belonging to the TNF family (BAFF)-R is the principal BAFF receptor facilitating BAFF costimulation of circulating T and B cells. J. Immunol. 173:807–817. [DOI] [PubMed] [Google Scholar]
- 18.Thangarajh, M., A. Gomes, T. Masterman, J. Hillert, and P. Hjelmstrom. 2004. Expression of B-cell-activating factor of the TNF family (BAFF) and its receptors in multiple sclerosis. J. Neuroimmunol. 152:183–190. [DOI] [PubMed] [Google Scholar]
- 19.Sorensen, T.L., M. Tani, J. Jensen, V. Pierce, C. Lucchinetti, V.A. Folcik, S. Qin, J. Rottman, F. Sellebjerg, R.M. Strieter, et al. 1999. Expression of specific chemokines and chemokine receptors in the central nervous system of multiple sclerosis patients. J. Clin. Invest. 103:807–815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Batten, M., C. Fletcher, L.G. Ng, J. Groom, J. Wheway, Y. Laabi, X. Xin, P. Schneider, J. Tschopp, C.R. Mackay, and F. Mackay. 2004. TNF deficiency fails to protect BAFF transgenic mice against autoimmunity and reveals a predisposition to B cell lymphoma. J. Immunol. 172:812–822. [DOI] [PubMed] [Google Scholar]
- 21.Babbe, H., A. Roers, A. Waisman, H. Lassmann, N. Goebels, R. Hohlfeld, M. Friese, R. Schroder, M. Deckert, S. Schmidt, et al. 2000. Clonal expansions of CD8+ T cells dominate the T cell infiltrate in active multiple sclerosis lesions as shown by micromanipulation and single cell polymerase chain reaction. J. Exp. Med. 192:393–404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Aloisi, F., G. Borsellino, P. Samoggia, U. Testa, C. Chelucci, G. Russo, C. Peschle, and G. Levi. 1992. Astrocyte cultures from human embryonic brain: characterization and modulation of surface molecules by inflammatory cytokines. J. Neurosci. Res. 32:494–506. [DOI] [PubMed] [Google Scholar]