

Abstract
Upon reaching the mature heat stable antigen (HSA)low thymic developmental stage, CD1d-restricted Vα14-Jα18 thymocytes undergo a well-characterized sequence of expansion and differentiation steps that lead to the peripheral interleukin-4/interferon-γ–producing NKT phenotype. However, their more immature HSAhigh precursors have remained elusive, and it has been difficult to determine unambiguously whether NKT cells originate from a CD4+CD8+ double-positive (DP) stage, and when the CD4+ and CD4−CD8− double-negative (DN) NKT subsets are formed. Here, we have used a CD1d tetramer-based enrichment strategy to physically identify HSAhigh precursors in thymuses of newborn mice, including an elusive DPlow stage and a CD4+ stage, which were present at a frequency of ∼10−6. These HSAhigh DP and CD4+ stages appeared to be nondividing, and already exhibited the same Vβ8 bias that characterizes mature NKT cells. This implied that the massive expansion of NKT cells is separated temporally from positive selection, but faithfully amplifies the selected TCR repertoire. Furthermore, we found that, unlike the DN γδ T cells, the DN NKT cells did not originate from a pTα-independent pathway bypassing the DP stage, but instead were produced during a short window of time from the conversion of a fraction of HSAlow NK1.1neg CD4 cells. These findings identify the HSAhigh CD4+ stage as a potential branchpoint between NKT and conventional T lineages and between the CD4 and DN NKT sublineages.
Mouse Vα14-Jα18/Vβ8,Vβ7,Vβ2 and human Vα24-Jα18/Vβ11 NKT cells are a conserved CD1d-restricted innate-like lymphocyte lineage that is involved in various infectious, allergic, autoimmune, and tumor conditions through the recognition of conserved endogenous and exogenous glycolipids (1–5). These glycolipids include the self-antigen, iGb3, generated in lysosomal compartments (4); microbial α-glycuronosylceramides found in the cell wall of Gram− LPS− bacteria (3, 5); and mycobacterial phosphatidylinositolmannosides (6).
The development of this unusual lineage has remained largely elusive. Two main models have been proposed: a precommitment model (7), which suggests that NKT cells originate from a committed precursor before TCR expression, and an instructive model (8) whereby mainstream thymocytes expressing the rare canonical CD1d-reactive TCR are instructed—by virtue of their interaction with CD1d-expressing cells—to differentiate into NKT cells. In strong support of the instructive model, transgenic expression of the canonical NKT cell receptor was sufficient to induce NKT cell differentiation (9). In addition, staining with CD1d tetramers identified developmental intermediates after positive selection, but preceding the acquisition of NK differentiation (10, 11). The earliest, well-characterized precursor identified was a rare, mature heat stable antigen (HSA)low CD44low thymocyte with a CD4+ or double-negative (DN) phenotype. Unlike other mature thymocytes, this cell seemed to be engaged in cell cycle, and gave rise to the CD44high memory-type stage. Cycling CD44high cells migrated to the periphery where they induced an “NK program”—including the expression of NK1.1, Ly49, and CD94/NKG2A—over the course of several weeks. In parallel with these phenotypic events, NKT cells acquired the ability to produce IL-4, then also IFN-γ upon TCR stimulation.
The conspicuous activation and expansion undergone by mature HSAlow NKT thymocytes suggest that they must have received unusual signals. Unlike mainstream T cells, NKT development depends on interaction with antigen expressed on the surface of bone marrow–derived cells, including double-positive (DP) thymocytes, rather than epithelial cells (12–16). Furthermore, recognition of the endogenous glycosphingolipid iGb3, an agonist antigen for mouse and human NKT cells, also is likely to be involved in the unusual signaling that leads to NKT cell expansion and differentiation (4). Consistent with the hypothesis of differential signaling, NKT development is unaffected by dominant negative forms of Ras or Mek-1 (17), but is abrogated completely in Fyn null mice (18, 19). The recent finding that signaling lymphocyte activation molecule (SLAM)–associated protein (SAP) connects Fyn to SLAM family members expressed on the cell surface (20–22) suggests that, in conjunction with TCR signaling, key signals may emanate from homotypic interaction between SLAM family members expressed by thymocytes (i.e., between the developing NKT precursors and neighboring CD1d-expressing DP thymocytes; references 23–26).
Integration of these recent advances in the general scheme of NKT cell development requires precise characterization of the immature HSAhigh stages; however, their very low frequency (estimated at less than 1/106) has precluded physical identification. For example, it is unclear whether NKT cells transit through a DP stage. The existence of very rare tetramer-positive DP thymocytes is controversial (11, 27, 28). PCR-based studies have revealed rare Vα14-Jα18 joints among DP thymocytes; however, it has not been determined whether the corresponding cells are enriched in or deprived of NKT cell precursors because the coexpression of Vβ chains (Vβ8, Vβ7, Vβ2) allowing endogenous ligand recognition has not been determined (27). In one study, purified DP thymocytes generated rare mature NKT cells upon intrathymic transfer; however, the inordinately high number of cells injected (107) may have included enough immature DN cell contaminants to account for NKT cell generation independent of the DP stage (27).
The origin of the DN sublineage of NKT cells, which is found in mouse and human, also is elusive. Because DN and CD4 cells are represented at the first developmental stage identified, the HSAlow CD44low cell, it remains unclear whether DN cells originate early from precursors that bypass the DP stage (e.g., as is the case for DN γδ T cells), or later from DP or even from CD4 T cells. Although the CD4 and CD8 coreceptors are dispensable for the development of the CD4 and DN NKT sublineages, the absence of CD8 has a measurable impact on the Vβ repertoire of CD4 and DN cells (12). This suggests that earlier expression of CD8 influences positive or negative selection, perhaps through interaction with CD1d. Transgenic expression of CD8αβ—under a CD2 promoter—drastically reduced the thymic NKT cell population; this also supports the hypothesis that CD8 may interact with CD1d (12). Finally, in mice transgenically expressing the Vα14-Jα18 TCR α chain, CD8 cells were depleted of the Vβ8 and Vβ7 chains that confer reactivity to CD1d (9). These findings suggested that the conspicuous absence of CD8+ NKT cells in WT mice may reflect negative selection or, as previously observed in a TCR transgenic system with agonist ligand signaling (29), the direct transformation of DP into DN or CD8αα cells.
In this study, we used a CD1d tetramer-based strategy to enrich NKT cell precursors from the thymus of WT newborn mice. Comparison with Jα18−/− and CD1d−/− NKT deficient controls allowed the identification of rare, noncycling HSAhigh DPlow and HSAhigh CD4+ precursors; intrathymic cell transfers demonstrated that the DN subset originated later from cycling HSAlow CD4 cells. These findings that identify the early stages of NKT cells in the thymus provide significant additions to the developmental map of NKT cells at the cellular level, and have implications on the molecular mechanisms that underlie this enigmatic pathway of T cell development.
RESULTS
Identification of HSAhigh Vα14 NKT cells in newborn thymus
Because of their massive intrathymic expansion, NKT cells are readily detectable among adult mature HSAlow thymocytes by specific tetramer staining. In contrast, as shown in Fig. 1, their frequency among the immature HSAhigh population is very low and comparable to that of Jα18-deficient thymuses where the canonical NKT TCR is absent or of CD1d-deficient thymus where NKT cells are not selected. This indicates that HSAhigh NKT lineage cells are too rare to be detected over a “nonspecific” staining background of 0.02%. However, upon magnetic-activated cell sorting (MACS)-enrichment with the same tetramers, bona fide, predominantly HSAhigh tetramer-positive cells were recovered from WT 3-d-old thymuses, but not Jα18-deficient or CD1d-deficient controls (Fig. 2). This result indicates that the cells that were stained “nonspecifically” by FACS were not recovered after MACS enrichment, perhaps because of their lower affinity interactions with the tetramers. Using this strategy, we were able to enrich the frequency of NKT lineage cells >1,000 fold; however, we estimate that under MACS conditions, only one third of the NKT lineage cells were recovered from these early thymuses. Therefore, the rarity of these NKT lineage cells in newborns required large-scale experiments; typically, we pooled 40–60 newborn thymuses to enrich and examine ∼200–300 “authentic” tet+ cells, because the procedure recovered approximately one NKT cell per 106 thymocytes at this age (Table I). In these 3-d-old mice, ∼70% of the tet+ cells, on average, were composed of HSAhigh cells (Fig. 2 and Table I), whereas the average proportion decreased to 57% at day 4, 13% at day 5, and 2% at day 8 (Fig. 3 and Table I). Nearly all of the tet+HSAhigh cells found in 3-d-old mice were CD4+ (Fig. 2). Rare other cells included DPlow cells that could not be distinguished readily from the background staining of control Jα18−/− thymuses. In older newborn mice, the HSAhigh cells were largely of the CD4+ phenotype, although minor proportions of DN cells could be detected (Fig. 3). In contrast, in the same tetramer-enriched population, the tet+HSAlow cells were composed of nearly equal proportions of CD4+ and DN cells as early as day 3 of age (Figs. 2 and 3). As shown in Figs. 2 and 3, HSAhigh CD4+ cells were absent from the thymus of CD1d-deficient mice, which supports the notion that they are the product of positive selection.
Figure 1.

Detection of HSAhigh NKT lineage cells in total thymocyte preparations. Total thymocytes from 2-wk-old WT, Jα18−/−, and CD1d−/− mice were stained with HSA and CD1d-tetramers. Gates identifying the tet+HSAlow and tet+HSAhigh cells are shown with corresponding frequencies. Note that the level of background staining in the HSAhigh gates (as measured in Jα18−/− and CD1d−/− mice) precludes the identification of HSAhigh NKT lineage cells in wild type mice. Data representative of >10 individual WT or mutant mice.
Figure 2.

Improved detection of HSAhigh NKT lineage cells in MACS-enriched thymocyte preparations. Thymocytes from 3-d-old WT, Jα18−/−, and CD1d−/− mice were MACS-enriched for tet+ cells and stained for HSA, CD4, and CD8. The left column shows CD1d-tetramer/HSA staining of MACS-enriched preparations, the other columns show CD4/CD8 staining of whole thymocytes, or MACS-enriched thymocytes in the R1 (HSAlow) and R2 (HSAhigh) gates, as indicated. The numbers on the HSA/CD1d-tetramers dot plots represent the percentages of HSAlow and HSAhigh cells among the tet+ population. The numbers on the R1 and R2 gated CD4/CD8 FACS dot plots represent the percentage of CD4+ and DN populations among HSAlow and HSAhigh NKT lineage cells. Note that in the tet+ MACS-enriched panels, each dot represents a cell: there were 3 and 5 tet+ dots in the MACS-enriched preparations of Jα18−/− and CD1d−/− thymuses, respectively, and 224 tet+ dots in the WT. Data are representative of at least three individual experiments with pooled thymuses from 40 to 60 3-d-old newborns.
Table I.
Ontogeny of NKT lineage thymocytes
| Frequency among total thymocytesc |
Frequency among tet+ cells |
||||
|---|---|---|---|---|---|
| Day | Total thymocyte no.a |
tet+ cells recovered/thymusb |
HSAhigh | HSAlow | HSAhigh |
|
|
×106
|
|
×10−7
|
%
|
|
| 3 | 12 ± 3 | 8 ± 3 | 3.5 ± 1.5 | 1.5 ± 1 | 70.5 ± 4.5 |
| 4 | 18 ± 2 | 10 ± 1 | 2.5 ± 0.5 | 2 ± 0.5 | 57.5 ± 7.5 |
| 5 | 25 ± 4 | 38 ± 5 | 2.8 ± 1 | 16 ± 2.5 | 13 ± 3 |
| 8 | 37 ± 2 | 3,729 ± 70 | 16 ± 4 | 982 ± 52 | 1.5 ± 0.5 |
Means ± SD are calculated from at least three separate experiments using pools of up to 40–60 newborn thymuses.
tet+ cells were counted after MACS enrichment. Only approximately one third of the tet+ cells were recovered by this method in 8-d-old thymuses.
Calculated based on tet+ cells recovered after MACS.
Figure 3.
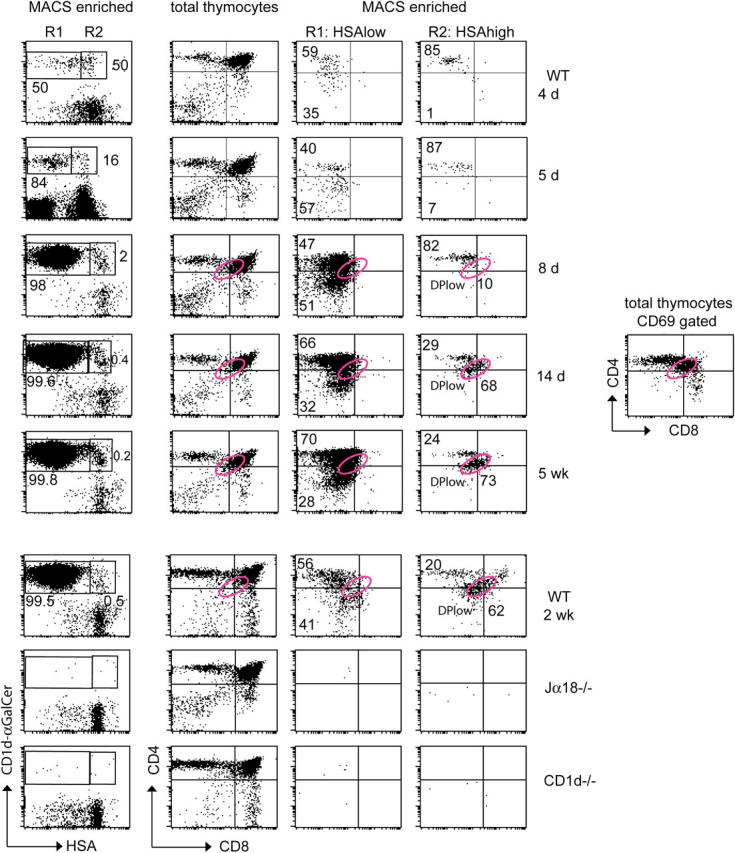
Ontogeny of HSAhigh and HSAlow NKT lineage thymocytes. Thymocytes of WT mice at different ages were enriched and analyzed as in Fig. 2. The total numbers of MACS-enriched tet+ cells at 4 and 5 d was 208 and 265, respectively (note that all dots are displayed and each dot represents a cell). Tet+ DPlow cells are in the pink elliptic gate and have the same DPlow phenotype as the CD69+ DP cells in the total thymocyte population shown in the rightmost panel of fourth row. Note that for consistency of CD4/CD8 staining, thymuses at 8 d, 14 d, and 5 weeks were analyzed side by side in the same experiment (rows 3–5), with doublet and propidium iodide uptake exclusion. The 2-wk-old thymocytes also were analyzed side by side with Jα18−/− and CD1d−/− thymuses in the same experiment (bottom three rows). Data are representative of at least three individual experiments where 5 to 60 thymuses were pooled.
Given the extreme rarity of the HSAhigh CD4+ cell, it is not surprising that we could not detect earlier stages, such as putative DP precursors. The lower TCR level and lower frequency of these precursors likely renders them undetectable. A population of DPlow cells belonging to the NKT lineage became visible at 2 wk of age (Fig. 3). A similar phenotype had been reported previously (27, 28), but was not confirmed in a subsequent study (11). Therefore, it was not clear whether these rare, unusual cells were bona fide NKT lineage cells or a staining artifact. The absence of DPlow cells in Jα18−/− mice (Fig. 3) indicates that they represent an authentic NKT lineage cell type, and their appearance at 2 weeks may help to explain seemingly contradictory reports. Their similar frequency to HSAhigh CD4+ cells suggests that they are a product of positive selection, as confirmed by their absence in the CD1d−/− thymus. In addition, the DPlow phenotype was identical to that of CD69+ DP thymocytes previously associated with positive selection (30; see the position of the pink gate in Figs. 3 and 4). The absence of this phenotype in younger mice may reflect different kinetics in the down-regulation of the CD4 and CD8 coreceptors and regulation of TCR following positive selection.
Figure 4.

Phenotypic characterization of HSAhigh NKT cells. Thymocytes were MACS-enriched for tet+ cells and (A) analyzed for size (forward scatter, FSC) and granularity (side scatter, SSC) or (B) stained for HSA, CD44, NK1.1, Vβ8, or CD69. Line graphs display the frequency of CD44+, NK1.1+, Vβ8+, and CD69+ cells among tet+HSAlow or tet+HSAhigh cells gated as indicated. Data are representative of three individual experiments using pools of 7-d-old or 2-wk-old mice as indicated.
HSAhigh Vα14 NKT cells are not cycling and express a biased Vβ8 usage
Mature HSAlow NKT cells are large, cycling cells that characteristically undergo multiple rounds of cell division that lead to massive intrathymic expansion (10). We found that the size of the HSAhigh cells, both DPlow and CD4, was comparable to that of the majority of thymocytes, and much smaller than the cycling HSAlow cell (Fig. 4 A). Although the rarity of these cell types precludes BrdU incorporation studies, their small size likely reflects the lack of cell cycling. This is consistent with the lack of expansion of this population after birth, compared with the explosive growth of HSAlow cells between days 5 and 8 (Table I). Thus, cell division is a late event that can be separated from the stages of development. The HSAhigh DP and HSAhigh CD4 cells expressed a CD44low NK1.1− phenotype similar to that of the following HSAlow stage (Fig. 4 B). In addition, they expressed CD69, as reported for cells having recently undergone positive selection (30).
Mature HSAlow NKT cells express a highly biased repertoire overusing Vβ8, Vβ7, and Vβ2; however, it is unclear whether this Vβ hierarchy results from preferential expansion at mature stages or from preferential selection at immature stages. Although the CD1d-α-galactosylceramide (CD1d-α-GalCer) tetramers may bias the analysis to cells expressing these three Vβ chains, Fig. 4 B shows that the HSAhigh cells exhibited the same frequency of Vβ8 chain usage as found in the HSAlow stages. This result suggests that the hierarchy of Vβ chain usage, at least among the three Vβ families, is already established after positive selection and is not modified substantially later by preferential cellular expansion.
DN cells arise from CD4 cells at the HSAlow NK1.1− stage
The exclusive CD4+ phenotype of the HSAhigh stage and the appearance of DN cells at the cycling HSAlow CD44low stage suggested that DN cells might arise at this latter stage from CD4+ precursors. We sorted the CD4+ subset of thymic HSAlow NK1.1− tet+ cells from WT mice and injected it into the thymus of Jα18−/− (NKT-deficient) recipients. In this transfer assay, the transferred NK1.1− cells matured to NK1.1+ within 5–7 d, as previously shown. In addition, the progeny of the CD4 subset included CD4+ and DN cells (Fig. 5 A), which demonstrated a precursor product relationship between CD4 and DN cells. A similar relationship was suggested previously (11). In contrast, the mature NK1.1+ cells conserved their CD4 or DN phenotype in the same intrathymic transfer conditions (Fig. 5 A). To rule out potential artifacts caused by tetramer staining, we sorted CD4+ CD44highNK1.1− cells from a preparation of HSAlowCD8neg thymocytes, and transferred them intrathymically. The progeny of the CD4+ cells contained CD4+ and DN tet+ NKT cells (Fig. 5 B). Together, these results demonstrate that DN cells arise from the conversion of CD4+ cells during a short developmental window.
Figure 5.

DN NKT cells arise from CD4+ precursors. NKT thymocyte subsets were isolated and FACS-sorted from 2–4-wk-old mice for intrathymic injection into NKT-deficient Jα18−/− recipients. The subsets injected included (A) tet+NK1.1−CD4+, tet+NK1.1+CD4+, and tet+NK1.1+CD4− (as gated) or (B) CD4+CD44highNK1.1− cells (as gated) obtained from a preparation of HSAlowCD8-negative IAβb−/− thymocytes without tetramer staining. CD4/NK1.1 profiles of sorted cells and their progeny 7 d after intrathymic injection are shown, as indicated. Numbers represent the percentages of each subpopulation in indicated gate or quadrant. Data are representative of three separate experiments.
DN NKT cells do not arise through a pTα-independent “γδ-like” pathway
It was hypothesized that a proportion of DN NKT cells might originate through a distinct pTα-independent “γδ-like” pathway that is characterized by the absence of induction of CD4 or CD8 coreceptor. Although pTα-deficient mice have normal γδ T cell compartments, they were reported to lack NKT cells, a finding that apparently is inconsistent with the hypothesis (31). However, it is recognized that the NKT cell defect in these mice might not be cell autonomous, but could be secondary to the profound depletion of thymocytes acting as CD1d-presenting cells for NKT cell selection. Therefore, we generated mixed, congenically marked pTα−/−:WT into irradiated Jα18−/− recipient bone marrow radiation chimeras. Fig. 6 demonstrates that, in contrast with WT NKT cells, which developed normally, there was a profound, cell autonomous NKT cell defect in the CD45.1+ pTα−/− compartment of these mixed chimeras.
Figure 6.

Cell-autonomous requirement for pTα in NKT thymocyte development. Mature NKT cells in WT, pTα−/−, and pTα−/−:WT bone marrow → Jα18−/− radiation chimeras. In the mixed chimera, the WT compartment is identified by CD45.2 gating and the pTα−/− compartment by CD45.1 as indicated. Data are representative of at least four mice in each group.
DISCUSSION
By enriching NKT precursors from pools of up to 60 newborn thymuses with CD1d-α-GalCer tetramers and MACS sorting, we have been able to identify and characterize very rare (10−6) HSAhigh cells that represent the earliest reported stages of NKT cell development (10, 11, 27, 28). The physical identification of the HSAhigh DPlow and HSAhigh CD4+ stages essentially completes the map of this enigmatic developmental pathway (Fig. 7). The nearly exclusive CD4+ phenotype of the rare HSAhigh precursor in 3-d-old mice, and the demonstration by cell transfer experiments that the CD4+ cells could give rise to a substantial fraction of DN cells clearly support a model whereby all DN cells arise late in development, after down-regulation of CD4 by a fraction of CD4+ cells. However, the presence of rare DN cells among the HSAhigh subset of older mice and the appearance of a population of DPlow cells in 2-wk-old mice suggest that the respective kinetics of CD4/CD8 coreceptor down-modulation may vary with age, and that commitment to the CD4 versus DN sublineage already may be imparted at a DP stage. The existence of DPlow cells has been controversial (11, 27, 28). Our results differ markedly from those of Gapin et al. (27) and Gadue et al. (28) who found relatively high frequencies of DPlow cells among total tet+ thymocytes. Instead, we found that authentic DPlow cells were very rare, and were restricted to the previously unrecognized HSAhigh stage of development.
Figure 7.

New model outlining the cellular developmental stages leading to mature CD4 and DN NKT cells. The dashed arrows indicate hypothetical precursor–product relationship.
This study could not detect tet+ DP thymocytes preceding positive selection in CD1d−/− mice. However, the unambiguous presence of DPlow cells after positive selection is important because it provides critical and direct evidence for the existence of such a DP precursor stage. Other experimental results support this conclusion. For example, the cell-autonomous requirement for pTα, as demonstrated in the present study, is highly suggestive of such a stage because pTα signaling induces CD4 and CD8 expression. The genetic role of CD8 in shaping the Vβ repertoire of CD4 and DN NKT cells also suggests their previous expression of CD8, and its interaction with CD1d (12). Finally, it was shown recently in a TCR transgenic model of agonist-mediated selection that immature DP cells could give rise to mature DN or CD8αα cells, a direct demonstration of the down-regulation of CD4 and CD8β (29). This example is particularly relevant to the development of NKT cells, because their thymic ligand, iGb3, also is an agonist that is capable of full activation of mature cells.
Given the rarity of the HSAhigh DPlow and HSAhigh CD4+ cell, it may not be surprising that, for technical reasons, preselection DP precursors would not be detected. The rarity of these precursors also supports the notion that canonical Vα14-Jα18 rearrangements occur stochastically at very low frequency.
It remains unclear what determines the CD4 versus DN fate of NKT cells. Their near 1:1 ratio might suggest a stochastic decision; however, an alternative model is that CD8 could be forcibly down-regulated from DP cells undergoing high levels of signaling. The CD4 versus DN choice also might be instructed by different TCR affinities for ligand.
The newly identified HSAhigh DPlow and HSAhigh CD4+ cells have a small size and express CD69. These stages, which precede the previously identified HSAlow CD4+ at which cells blast and undergo intense cell division, are analogous to the HSAhigh CD69+ stage following positive selection of mainstream MHC-restricted T cells (30). These positively selected MHC-restricted T cells will not engage in cell cycle. Thus, the HSAhigh CD4+ cell represents the likely branchpoint in the development of mainstream T versus NKT cells. An unresolved issue is whether commitment to cell division already is imparted at this stage, or whether it occurs later at the HSAlow stage (e.g., upon second antigenic stimulation by medullary dendritic cells). Although there is no example, to our knowledge, of mitogenic stimuli imparted at the immature HSAhigh stage, some arguments suggest that this may be the case. First, entry into the cell cycle is delayed by 1 or 2 d following TCR stimulation, corresponding to the lag time between HSAhigh and HSAlow CD4 T cells. Second, recent experiments have demonstrated that restricted expression of CD1d on DP thymocytes was sufficient to drive the major expansion observed at the medullary HSAlow stage (32).
These different lines of evidence raise the surprising possibility that a single set of signaling interactions between HSAhigh NKT cell precursors and neighboring cortical thymocytes—involving TCR and perhaps SLAM family members as coreceptors—may represent the defining event of the NKT lineage imparting the subsequent NKT expansion and differentiation program.
MATERIALS AND METHODS
Mice.
C57BL/6, C57BL/6.CD45.2, and C57BL/6.I-Aβb−/− mice were obtained from Taconic Farms. CD1d−/− mice were used after 12 backcrosses to C57BL/6 (33). Jα18−/− (34) and pTα−/− (35) mice on C57BL/6 backgrounds were gifts from M. Taniguchi (RIKEN, Yokohama, Japan) and I. Aifantis (University of Chicago, Chicago, IL), respectively. All mice were maintained under specific pathogen-free conditions at the University of Chicago, and experimental studies were in accordance with the Institutional Animal Care and Use Guidelines.
Flow cytometry.
FITC-conjugated mAbs against CD8, Vβ8.1/8.2, CD45.2, CD4, CD24 (HSA), CD44, and CD69; PE-conjugated mAbs against CD69, HSA, CD44, and NK1.1; PE-Cy7–conjugated mAb against CD4; cychrome-conjugated mAbs against CD24, CD4, and CD8; and allophycocyanin-conjugated mAbs against CD4, CD44, CD8, and NK1.1 were obtained from BD Biosciences. Propidium iodide was from Sigma-Aldrich. CD1d-α-GalCer tetramers were produced with streptavidin-PE or -allophycocyanin and used for staining as described previously (36). Samples were analyzed using a four-color FACSort equipped with argon and 635-nm diode lasers (Becton Dickinson) and CELLQuest software. For five-color staining, samples were analyzed using BD LSR II equipped with 488 sapphire and 633 HeNe lasers (Becton Dickinson) and FACSDiva software. In some experiments, doublet and propidium iodide exclusion was applied to remove irrelevant dots.
Enrichment of CD1d-αGalCer tetramer+ cells with paramagnetic microbeads.
Thymuses were pooled from batches of 40 to 60 newborns (day of birth considered as day 1), and CD1d tetramer+ thymocytes were enriched using anti-PE or -allophycocyanin microbeads and the AutoMACs system (Miltenyi Biotec) at ice cold temperature following the manufacturer's instructions.
FACS sorting of NKT cells.
Pooled thymuses from batches of 25 to 50 C57BL/6.IAβb−/− or C57BL/6 mice (2–4-wk-old) were depleted of CD8+ cells using anti-CD8 microbeads and the AutoMACs system (Miltenyi Biotec); stained with CD1d-αGalCer streptavidin-PE along with anti-CD4 and -NK1.1; and the NKT cells were sorted into CD4+NK1.1−, CD4+NK1.1+, and CD4−NK1.1+ subsets. Alternatively, to avoid TCR cross-linking by tetramer staining, CD8-negative thymocyte preparations were stained with HSA, CD4, CD44, and NK1.1 for sorting of HSAlowCD4+CD44highNK1.1− cells.
Intrathymic injection.
Sorted tetramer-positive CD4+NK1.1−, CD4+ NK1.1+, and CD4−NK1.1+ subsets or sorted CD4+CD44highNK1.1− cells (4–8 × 105) were injected intrathymically as described, and their progeny were analyzed 5 to 7 d later after MACS enrichment of CD1d-tetramer positive cells.
Bone marrow radiation chimeras.
C57BL/6.Jα18−/− mice received whole body γ-irradiation (1,000 Rad) with a cesium source (Gammacell 40), and were reconstituted 6 h later with one i.v. injection of 5–10 × 106 bone marrow cells from various adult donors.
Acknowledgments
We thank R. Duggan, J. Marvin, and B. Eisfelder (Flow Cytometry Facility) for cell sorting, and all Bendelac lab members for expert advice and discussion.
This work is supported by a special fellowship from the Leukemia and Lymphoma Society (to K. Benlagha) and by National Institutes of Health grant no. RO1 AI 38339 (to A. Bendelac).
The authors have no conflicting financial interests.
Abbreviations used: CD1d-αGalCer; CD1d-α-galactosylceramide; DP, double-positive; DN, double-negative; HSA, heat stable antigen; MACS, magnetic-activated cell sorting; SAP, SLAM-associated protein; SLAM, signalling lymphocyte activation molecule.
References
- 1.Park, S.H., and A. Bendelac. 2000. CD1-restricted T-cell responses and microbial infection. Nature. 406:788–792. [DOI] [PubMed] [Google Scholar]
- 2.Godfrey, D.I., K.J. Hammond, L.D. Poulton, M.J. Smyth, and A.G. Baxter. 2000. NKT cells: facts, functions and fallacies. Immunol. Today. 21:573–583. [DOI] [PubMed] [Google Scholar]
- 3.Kinjo, Y., D. Wu, G. Kim, G.W. Xing, M.A. Poles, D.D. Ho, M. Tsuji, K. Kawahara, C.H. Wong, and M. Kronenberg. 2005. Recognition of bacterial glycosphingolipids by natural killer T cells. Nature. 434:520–525. [DOI] [PubMed] [Google Scholar]
- 4.Zhou, D., J. Mattner, C. Cantu III, N. Schrantz, N. Yin, Y. Gao, Y. Sagiv, K. Hudspeth, Y.P. Wu, T. Yamashita, et al. 2004. Lysosomal glycosphingolipid recognition by NKT cells. Science. 306:1786–1789. [DOI] [PubMed] [Google Scholar]
- 5.Mattner, J., K.L. DeBord, N. Ismail, R.D. Goff, C. Cantu III, D. Zhou, P. Saint-Mezard, V. Wang, Y. Gao, N. Yin, et al. 2005. Both exogenous and endogenous glycolipid antigens activate NKT cells during microbial infections. Nature. 434:525–529. [DOI] [PubMed] [Google Scholar]
- 6.Fischer, K., E. Scotet, M. Niemeyer, H. Koebernick, J. Zerrahn, S. Maillet, R. Hurwitz, M. Kursar, M. Bonneville, S.H. Kaufmann, and U.E. Schaible. 2004. Mycobacterial phosphatidylinositol mannoside is a natural antigen for CD1d-restricted T cells. Proc. Natl. Acad. Sci. USA. 101:10685–10690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Makino, Y., R. Kanno, H. Koseki, and M. Taniguchi. 1996. Development of Valpha4+ NK T cells in the early stages of embryogenesis. Proc. Natl. Acad. Sci. USA. 93:6516–6520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bendelac, A., M.N. Rivera, S.-H. Park, and J.H. Roark. 1997. Mouse CD1-specific NK1 T cells. Development, specificity, and function. Annu. Rev. Immunol. 15:535–562. [DOI] [PubMed] [Google Scholar]
- 9.Bendelac, A., R.D. Hunziker, and O. Lantz. 1996. Increased interleukin 4 and immunoglobulin E production in transgenic mice overexpressing NK1 T cells. J. Exp. Med. 184:1285–1293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Benlagha, K., T. Kyin, A. Beavis, L. Teyton, and A. Bendelac. 2002. A thymic precursor to the NKT cell lineage. Science. 296:553–555. [DOI] [PubMed] [Google Scholar]
- 11.Pellicci, D.G., K.J. Hammond, A.P. Uldrich, A.G. Baxter, M.J. Smyth, and D.I. Godfrey. 2002. A natural killer T (NKT) cell developmental pathway involving a thymus-dependent NK1.1(-)CD4(+) CD1d-dependent precursor stage. J. Exp. Med. 195:835–844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Bendelac, A., N. Killeen, D. Littman, and R.H. Schwartz. 1994. A subset of CD4+ thymocytes selected by MHC class I molecules. Science. 263:1774–1778. [DOI] [PubMed] [Google Scholar]
- 13.Bix, M., M. Coles, and D. Raulet. 1993. Positive selection of Vb8+ CD4-8- thymocytes by class I molecules expressed by hematopoietic cells. J. Exp. Med. 178:901–908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Coles, M.C., and D.H. Raulet. 2000. NK1.1+ T cells in the liver arise in the thymus and are selected by interactions with class I molecules on CD4+CD8+ cells. J. Immunol. 164:2412–2418. [DOI] [PubMed] [Google Scholar]
- 15.Bendelac, A. 1995. Positive selection of mouse NK1+ T cells by CD1-expressing cortical thymocytes. J. Exp. Med. 182:2091–2096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ohteki, T., and H.R. MacDonald. 1994. Major histocompatibility complex class I related molecules control the development of CD4+8- and CD4-8- subsets of natural killer 1.1+ T cell receptor-a/b+ cells in the liver of mice. J. Exp. Med. 180:699–704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Alberola-Ila, J., K.A. Hogquist, K.A. Swan, M.J. Bevan, and R.M. Perlemutter. 1996. Positive and negative selection invoke distinct signaling pathways. J. Exp. Med. 184:9–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gadue, P., N. Morton, and P.L. Stein. 1999. The Src family tyrosine kinase Fyn regulates natural killer T cell development. J. Exp. Med. 190:1189–1196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Eberl, G., B. Lowin-Kropf, and H.R. MacDonald. 1999. Cutting edge: NKT cell development is selectively impaired in Fyn-deficient mice. J. Immunol. 163:4091–4094. [PubMed] [Google Scholar]
- 20.Sayos, J., C. Wu, M. Morra, N. Wang, X. Zhang, D. Allen, S. van Schaik, L. Notarangelo, R. Geha, M.G. Roncarolo, et al. 1998. The X-linked lymphoproliferative-disease gene product SAP regulates signals induced through the co-receptor SLAM. Nature. 395:462–469. [DOI] [PubMed] [Google Scholar]
- 21.Chan, B., A. Lanyi, H.K. Song, J. Griesbach, M. Simarro-Grande, F. Poy, D. Howie, J. Sumegi, C. Terhorst, and M.J. Eck. 2003. SAP couples Fyn to SLAM immune receptors. Nat. Cell Biol. 5:155–160. [DOI] [PubMed] [Google Scholar]
- 22.Latour, S., R. Roncagalli, R. Chen, M. Bakinowski, X. Shi, P.L. Schwartzberg, D. Davidson, and A. Veillette. 2003. Binding of SAP SH2 domain to FynT SH3 domain reveals a novel mechanism of receptor signalling in immune regulation. Nat. Cell Biol. 5:149–154. [DOI] [PubMed] [Google Scholar]
- 23.Chung, B., A. Aoukaty, J. Dutz, C. Terhorst, and R. Tan. 2005. Signaling lymphocytic activation molecule-associated protein controls NKT cell functions. J. Immunol. 174:3153–3157. [DOI] [PubMed] [Google Scholar]
- 24.Nichols, K.E., J. Hom, S.Y. Gong, A. Ganguly, C.S. Ma, J.L. Cannons, S.G. Tangye, P.L. Schwartzberg, G.A. Koretzky, and P.L. Stein. 2005. Regulation of NKT cell development by SAP, the protein defective in XLP. Nat. Med. 11:340–345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Pasquier, B., L. Yin, M.C. Fondaneche, F. Relouzat, C. Bloch-Queyrat, N. Lambert, A. Fischer, G. de Saint-Basile, and S. Latour. 2005. Defective NKT cell development in mice and humans lacking the adapter SAP, the X-linked lymphoproliferative syndrome gene product. J. Exp. Med. 201:695–701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Borowski, C., and A. Bendelac. 2005. Signaling for NKT cell development: the SAP-Fyn connection. J. Exp. Med. 201:833–836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gapin, L., J.L. Matsuda, C.D. Surh, and M. Kronenberg. 2001. NKT cells derive from double-positive thymocytes that are positively selected by CD1d. Nat. Immunol. 2:971–978. [DOI] [PubMed] [Google Scholar]
- 28.Gadue, P., and P.L. Stein. 2002. NK T cell precursors exhibit differential cytokine regulation and require Itk for efficient maturation. J. Immunol. 169:2397–2406. [DOI] [PubMed] [Google Scholar]
- 29.Yamagata, T., D. Mathis, and C. Benoist. 2004. Self-reactivity in thymic double-positives commits cells to a CD8αα lineage with characteristics of innate immune cells. Nat. Immunol. 5:597–605. [DOI] [PubMed] [Google Scholar]
- 30.Bendelac, A., P. Matzinger, R.A. Seder, W.E. Paul, and R.H. Schwartz. 1992. Activation events during thymic selection. J. Exp. Med. 175:731–742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Eberl, G., H.J. Fehling, H. von Boehmer, and H.R. MacDonald. 1999. Absolute requirement for the pre-T cell receptor alpha chain during NK1.1+ TCRalphabeta cell development. Eur. J. Immunol. 29:1966–1971. [DOI] [PubMed] [Google Scholar]
- 32.Wei, D.G., H. Lee, S.H. Park, L. Beaun, L. Teyton, A. Lehuen, and A. Bendelac. 2005. Expansion and long range differentiation of the NKT lineage in mice expressing CD1d exclusively on cortical thymocytes. J. Exp. Med. 202:239–248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Park, S.H., D. Guy-Grand, F.A. Lemonnier, C.R. Wang, A. Bendelac, and B. Jabri. 1999. Selection and expansion of CD8alpha/alpha(1) T cell receptor alpha/beta(1) intestinal intraepithelial lymphocytes in the absence of both classical major histocompatibility complex class I and nonclassical CD1 molecules. J. Exp. Med. 190:885–890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Cui, J., T. Shin, T. Kawano, H. Sato, E. Kondo, I. Toura, Y. Kaneko, H. Koseki, M. Kanno, and M. Taniguchi. 1997. Requirement for Valpha14 NKT cells in IL-12-mediated rejection of tumors. Science. 278:1623–1626. [DOI] [PubMed] [Google Scholar]
- 35.Fehling, H.J., A. Krotkova, C. Saint-Ruf, and H. von Boehmer. 1995. Crucial role of the pre-T-cell receptor alpha gene in development of alpha beta but not gamma delta T cells. Nature. 375:795–798. [DOI] [PubMed] [Google Scholar]
- 36.Benlagha, K., A. Weiss, A. Beavis, L. Teyton, and A. Bendelac. 2000. In vivo identification of glycolipid antigen specific T cells using fluorescent CD1d tetramers. J. Exp. Med. 191:1895–1903. [DOI] [PMC free article] [PubMed] [Google Scholar]