

Abstract
Long-lasting expansion of Vδ2neg γδ T cells is a hallmark of cytomegalovirus (CMV) infection in kidney transplant recipients. The ligands of these cells and their role remain elusive. To better understand their immune function, we generated γδ T cell clones from several transplanted patients. Numerous patient Vδ1+, Vδ3+, and Vδ5+ γδ T cell clones expressing diverse Vγ chains, but not control Vγ9Vδ2+ T clones, displayed strong reactivity against CMV-infected cells, as shown by their production of tumor necrosis factor-α. Vδ2neg γδ T lymphocytes could also kill CMV-infected targets and limit CMV propagation in vitro. Their anti-CMV reactivity was specific for this virus among herpesviridae and required T cell receptor engagement, but did not involve major histocompatibility complex class I molecules or NKG2D. Vδ2neg γδ T lymphocytes expressed receptors essential for intestinal homing and were strongly activated by intestinal tumor, but not normal, epithelial cell lines. High frequencies of CMV- and tumor-specific Vδ2neg γδ T lymphocytes were found among patients' γδ T cells. In conclusion, Vδ2neg γδ T cells may play a role in protecting against CMV and tumors, probably through mucosal surveillance of cellular stress, and represent a population that is largely functionally distinct from Vγ9Vδ2+ T cells.
Although γδ T cells represent a minor population at the periphery (<5% of T cells), they are normal residents in several tissues, notably epithelia where they reach 10–50% of T cells. In humans, 70% of γδ T cells in the peripheral blood express Vδ2 in association with Vγ9 chains. Vγ9Vδ2 T cells recognize small nonpeptidic phosphorylated antigens derived, for example, from Mycobacteria (1, 2), some biphosphonates (3), alkylamines (4), and the association of membrane F1-ATPase and apolipoprotein A-I (5). These compounds are ubiquitous components of microorganisms, as well as of self-antigens. In contrast, most tissue γδ T cells (located mainly in spleen and epithelia) use the Vδ1 chain associated with any Vγ chain to form their TCR. Their antigen specificity has been explored less, but their propensity toward self-reactivity has been demonstrated, as it has been for Vγ9/Vδ2 T cells. MHC class I–related chain A (MICA; a stress-induced protein expressed on epithelial cells) and CD1c are indeed able to stimulate Vδ1+ γδ T cell lines and to interact with their Vδ1 TCR (6, 7). An important feature of γδ T cells is that they do not require classical MHC-presenting molecules in the antigen recognition process.
It is now accepted that γδ T cells play specific and nonredundant functions with those of αβ T cells, but the exact role and features of these cells are still a matter of debate (for review see reference 8). The antigenic specificity and tissue distribution of γδ T cells make them important effectors of antitumor immunity. Vγ9Vδ2 T cells can kill myeloma and lymphoma cell lines in vitro (9) and injection of their cognate ligands was associated with improvement in patients with non-Hodgkin lymphoma or multiple myeloma (10). Vδ1 γδ T cells are found expanded in epithelial tumors, and they exert an in vitro antitumor activity by recognizing antigens that are overexpressed on tumor cells (11, 12). In addition, the importance of γδ T cells has been suspected in several antiinfectious immune responses essentially on the basis of the selective expansion of these cells in the peripheral blood of patients. As far as is concerned in viral infections, increase of γδ T cells at the periphery has been documented in HIV (13), EBV (14), and HSV (15) infections. Although the results gathered so far suggest their participation in these contexts, their protective functions remain to be ascertained yet.
A few years ago, we demonstrated the involvement of γδ T cells in human CMV infection. CMV is a widespread β herpesvirus that infects 60–100% of the populations around the world and induces lifelong viral persistence without detectable symptoms in immunocompetent individuals. In contrast, CMV infection in immunocompromised patients usually leads to severe disease. Consequently, numerous studies have been devoted to the understanding of the mechanisms evolved by the immune system to control this pathogen. CD8+ αβ cytotoxic T cells are usually considered as the principal actors of the anti-CMV cellular immune response, as they can kill CMV-infected cells in vitro, and adoptive transfer of CMV-specific CD8+ T cells efficiently restricts CMV infection in recipients of allogeneic bone marrow (16). The marked increase of circulating γδ T cells (from <5 up to 50% of total T cells) we reported in kidney-transplanted patients with active CMV infection (17) strongly indicates an involvement of γδ T cells in anti-CMV immune responses. This cellular amplification occurred concomitant to the resolution of viremia, suggesting a protective antiviral role for γδ T cells (18). Interestingly, Vδ1, Vδ3 as well as Vδ5-expressing γδ T cells were involved, whereas Vγ9Vδ2 T cells never expanded, and they display a restricted repertoire evocative of an antigenic selection in vivo (17). More recent studies describe an increase of γδ T cell number in the salivary glands of infected mice (19) and demonstrate a protective role for liver Vγ1+ γδ T cells against murine CMV by depleting γδ T cells in vivo (20). These data lend further support to the conclusion drawn from CMV-infected patients whereby Vδ2neg γδ T cell subsets most likely expand or are redistributed from the tissues where they reside normally and which are preferential sites of CMV multiplication (e.g., intestinal epithelia, liver, or salivary glands).
The goal of the present study is to define the immune functions of these γδ T cell subpopulations in the response of the host to human CMV. In vitro studies were performed to characterize their reactivity toward CMV-infected cells and ultimately led us to show their reactivity against tumor epithelial cells.
Results
Vδ2neg γδ T cells specifically recognize CMV-infected fibroblasts
To determine whether the Vδ2neg γδ T cells expanded in the context of CMV infection were able to recognize CMV-infected cells, we have sorted these populations from the peripheral blood of different renal- or lung-transplanted patients with a CMV infection and derived cell lines under polyclonal stimulation (in the absence of CMV) before cloning by limiting dilution. These Vδ2neg γδ T cell clones were seeded onto monolayers of the MRC5 fibroblastic cell line, which were previously infected or not with CMV for different periods of time. Several Vδ2neg γδ T cell clones were found to produce high amounts of TNFα in culture supernatants when cocultured with MRC5 monolayers previously infected for 4 d with CMV (Fig. 1 A). Neither infected MRC5 alone, nor γδ T cells cultured with uninfected MRC5 produced TNFα. That TNFα produced in coculture supernatants originated from γδ T cells themselves was indicated by intracytoplasmic staining of this cytokine, which was only positive in Vδ2neg γδ T cell clones incubated on MRC5 cells infected with CMV or treated with PMA and ionomycin, but not when cultured with uninfected fibroblastic cells (Fig. 1 B). U373 (an astrocytoma cell line) and primary cultures of foreskin fibroblasts (FSFs) were also able to stimulate Vδ2neg γδ T cells after infection with CMV (unpublished data). CMV-reactive Vδ2neg γδ T cell clones expressed diverse γδ TCR chain associations comprising Vγ2,3, Vγ4, Vγ8, or Vγ9 chains associated with Vδ1, Vδ3, or Vδ5 chains (Table I). In contrast, CMV-infected MRC5 cells were always found unable to activate control Vγ9Vδ2 T cell clones (Table I). Reactivity of Vδ2neg γδ T cells was specifically directed toward CMV because they did not produce TNFα when challenged with HSV or varicella-zoster virus (VZV)-infected cells (Fig. 1 C), whereas MRC5 infected with any of the CMV strains tested, either clinical (TB40/E) or laboratory adapted (AD169 and Towne), induced the production of TNFα by Vδ2neg γδ T cells.
Figure 1.
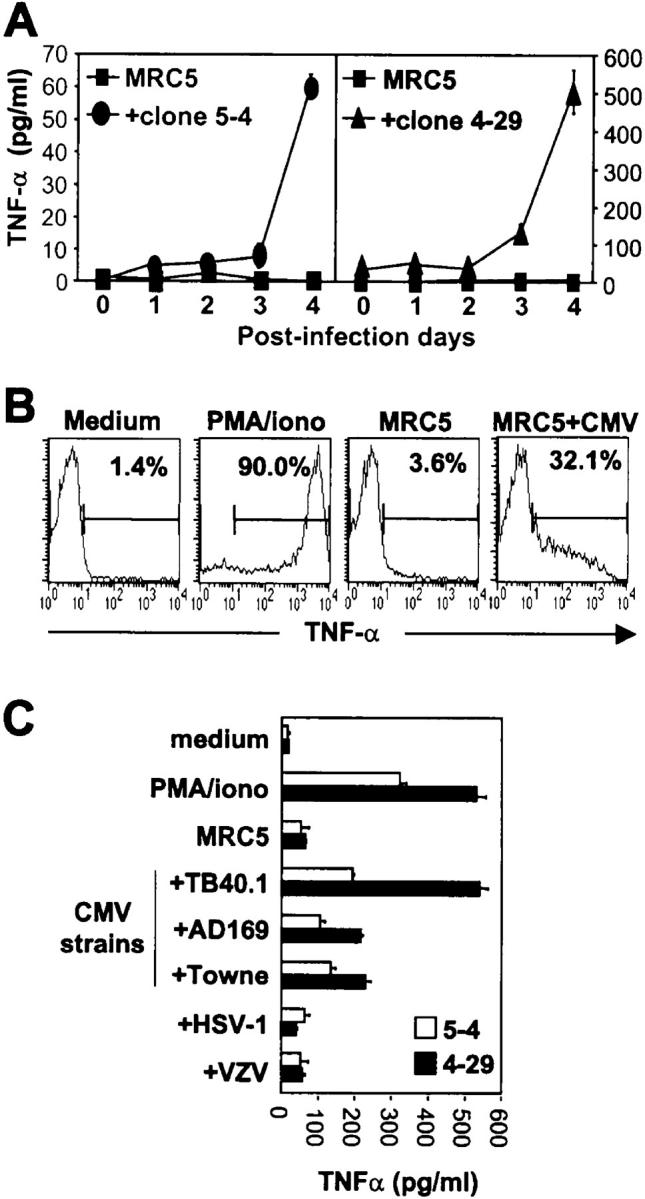
Vδ2neg γδ T cells produce TNFα when cocultured with CMV-infected MRC5 cells. (A) MRC5 cells were infected for 1–4 d with CMV or left uninfected, and cocultured with two representative Vδ2neg γδ T clones (5–4 and 4–29) or medium alone for TNFα release quantification by ELISA. Results are presented as mean ±SD of culture triplicates. (B) Vδ2neg γδ T cells were cultured in medium alone, with PMA and ionomycin, or with uninfected or CMV-infected MRC5 cells. Cells were labeled with anti-Cδ mAb and anti-TNFα mAb and analyzed by flow cytometry. Only γδ T cells are shown in the histograms and the percentages of TNFα- expressing cells are indicated. (C) Anti-CMV reactive 5-4 and 4–29 clones have been cultured for 4 h in medium alone, with PMA and ionomycin, or on MRC5 cells uninfected or infected with TB40/E, AD169, or Towne strains of CMV, or with clinical isolates of VZV or HSV-1. Results are mean ± SD of TNFα secreted in supernatant of culture triplicates. All results are representative of at least three independent experiments.
Table I.
Anti-CMV reactivity of Vδ2neg γδ T cell clones is independent of the TCR V chain usage
| TNFα production (pg/ml)c
|
|||||
|---|---|---|---|---|---|
| Patient-clone numbera |
V segment usageb |
Medium | PMA + iono | MRC5 | MRC5 + CMV |
| 1-12 | Vγ2,3 Vδ1 | 20 | 462 | <5 | 63 |
| 2-6 | Vγ2,3 Vδ1 | <5 | 232 | 12 | 81 |
| 3-1 | Vγ2,3 Vδ1 | <5 | 470 | <5 | 94 |
| 4-7 | Vγ4 Vδ5 | <5 | 208 | <5 | 117 |
| 4-9 | Vγ4 Vδ5 | <5 | 84 | <5 | 114 |
| 4-29 | Vγ4 Vδ5 | <5 | 169 | <5 | 508 |
| 5-4 | Vγ9 Vδ1 | <5 | 233 | <5 | 373 |
| 6-1 | Vγ8 Vδ1 | <5 | 273 | <5 | 46 |
| 7-2
|
Vγ2,3 Vδ3 | <5 | 59 | <5 | 32 |
| TH1 | Vγ9 Vδ2 | <5 | 335 | <5 | <5 |
| G12 | Vγ9 Vδ2 | <5 | 425 | <5 | <5 |
Vδ2neg γδ T cell clones were generated from seven different patients (1–7). Two control Vγ9Vδ2 T cell clones were also tested (TH1 and G12).
V regions were identified by flow cytometry analysis using specific mAbs. Vγ2 and Vγ3 regions cannot be distinguished because they are recognized by the same antibody. Vδ-Jδ sequences of all these clones are presented in Table S1 (available at http://www.jem.org/cgi/content/full/jem.20041851/DC1).
3 × 104 cells/well of each γδ T cell clone were cultured for 4 h in medium alone, or with PMA and ionomycin, or with MRC5 cells uninfected or infected for 4 d with CMV. TNFα concentration was measured in culture supernatants by ELISA. Results are the mean of cultures triplicates and SD was always <15% of the mean value (unpublished data).
Vδ2neg γδ T cells are able to kill CMV-infected cells and to inhibit CMV propagation in vitro
To gain insight into mechanisms of γδ T cell antiviral response, next we examined their ability to lyse CMV-infected cells. Vδ2neg γδ T cell clones were cultured for 4 h with CMV-infected or uninfected FSFs labeled with 51Cr. CMV-infected FSFs were efficiently lysed by Vδ2neg γδ T cell clones (Fig. 2 A), whereas we observed no specific cytotoxicity against uninfected FSFs. γδ T cells were previously shown to kill their targets using either the granzyme/perforin system, or TNFα- and FasL-mediated pathways (21). Most cells within Vδ2neg γδ T cell clones were found to express spontaneously high levels of granzyme B and perforin (unpublished data). In addition, lysis of CMV-infected fibroblasts was not inhibited by blocking mAbs directed against Fas or TNF-α, whereas these mAbs efficiently abrogated Faspos Jurkat cell lysis by Fas-Lpos 1A12 cells and the TNFα-induced lysis of WEHI cells, respectively (Fig. 2 B). Moreover, pretreatment of Vδ2neg γδ T cells with strontium ions, which induce exhaustive granule secretion and render perforin and granzyme-secreting T cells temporarily incapable of killing target cells, inhibited their cytotoxic activity toward HT29 and CMV-infected FSFs (Fig. 2 B). These results indicate that perforin–granzyme B is the main effector pathway used by cytotoxic CMV-reactive Vδ2neg γδ T cells.
Figure 2.

Vδ2neg γδ T cells can lyse CMV-infected cells and limit CMV replication in vitro. (A) FSF uninfected (dashed lines) or infected with CMV (solid lines) were labeled with 51Cr and incubated with four different anti-CMV γδ T cell clones. Results are the mean of specific lysis of culture triplicates. SD was always <15% of the mean value (not depicted). (B) As in A, but blocking mAbs directed against TNFα (20 μg/ml) or Fas (1 μg/ml) or 20 μg/ml control mAb were added at the start of cell contact. When indicated, γδ T cell clone was pretreated with SrCl or NaCl at 20 mM before contact with FSF. Viability of T cells after this treatment was >90%. Effector/target ratio was 10/1. As positive control for mAb efficacy, WEHI or Jurkat cells were labeled with 51Cr and incubated with 50 pg/ml TNFα or Fas-L+ 1A12 cells (10/1 effector/target ratio), respectively. Same results were obtained with the 5–4 clone. (C) Uninfected or infected FSF were labeled as in A and mixed with the same number of either cold uninfected FSF or cold infected FSF, and incubated with the 4–29 anti-CMV γδ T cell clone at a 10/1 effector/target ratio. (D) FSF were incubated for 2 h with CMV, washed, and incubated in medium alone, with IFNγ (100, 10, and 1 U/ml), or with the anti-CMV 4-29 or unrelated T clones (Vδ1 or a Vγ9Vδ2 T cells). Anti-IFNγ blocking mAb or an unrelated control mAb (20 and 2 μg/ml) were added to 4-29 T cell cocultures. After 4 d, infectious CMV was titered in culture supernatants by plaque assay on MRC5 cells. Results are mean ± SD of culture triplicates and are representative of three independent experiments.
To assess whether Vδ2neg γδ T cells could display a bystander cytolytic activity toward neighboring uninfected cells within CMV-infected tissue, a mixture of 51Cr-labeled uninfected FSFs and cold CMV-infected FSFs (or vice versa) were incubated with Vδ2neg γδ T clones. Chromium release could be observed only in conditions where CMV-infected FSFs, but not uninfected FSFs, were labeled with 51Cr, demonstrating the high specificity and direct contact requirement of Vδ2neg γδ T cell killing activity (Fig. 2 C).
We next asked whether the effector functions of Vδ2neg γδ T cells could limit CMV propagation in vitro. Fibroblast monolayers were incubated with CMV for 2 h, washed, and further cultured in the presence of CMV-reactive Vδ2neg γδ T cell clones. After 4 d, the number of infectious particles released into the supernatant were greatly reduced in the presence of the anti-CMV Vδ2neg γδ T cell clone as compared with medium alone and to unrelated Vδ1pos or Vγ9Vδ2 T cell clones (Fig. 2 D). The antiviral action of 5 × 104 Vδ2neg γδ T cell clones was as potent as that of 1 U/ml IFNγ used as a positive control for the inhibition of viral replication. Accordingly, the production of IFNγ by Vδ2neg γδ T cells could explain the inhibition of CMV replication because an anti-IFNγ blocking mAb nearly completely abrogated the antiviral effect of the Vδ2neg γδ T cells (Fig. 2 D). Thus, the recognition of infected MRC5 by Vδ2neg γδ T cells resulted in production of IFNγ in sufficient amount to control CMV replication in vitro.
Recognition of CMV-infected cells by Vδ2neg γδ T cells is mediated through the γδ TCR independently of MHC class I presentation
To assess whether activation of Vδ2neg γδ T cells by CMV-infected cells is mediated by antigen/TCR recognition, cocultures of T cell clones on CMV-infected or uninfected FSF monolayers were performed in the presence of increasing doses of anti-CD3 (OKT3) or anti-Cδ blocking mAbs to prevent TCR engagement. Addition of these mAbs inhibited the release of TNFα by T cells in a dose-dependent manner (Fig. 3 A). The slight increase of TNFα release observed with nonsaturating doses of the mAbs in cocultures performed with uninfected FSFs is likely to be related to mAb binding on FcR faintly expressed on FSFs and to the subsequent triggering of γδ TCR signaling. To further establish this γδ TCR involvement, we monitored the TCR down-modulation at the surface of Vδ2neg γδ T cell clones after TCR engagement by cognate ligand (22). An anti-CMV Vδ2neg and a control Vγ9Vδ2 T cell clone were incubated with uninfected or CMV-infected MRC5 cells and their TCR expression was measured by flow cytometry every 2 h. CMV-infected MRC5 cells, but not uninfected cells, strongly down-modulated TCR expression only on the surface of the anti-CMV γδ T cell clone; those cells do this to an extent comparable to that induced by an agonistic anti-CD3 mAb (UCHT1).
Figure 3.

Vδ2neg γδ TCR recognition of CMV-infected cells is TCR dependent but MHC class I independent. (A) FSF were infected or not with CMV and incubated with the 4–29 anti-CMV Vδ2neg γδ T cell clone in the presence or absence of increasing doses of anti-CD3 or anti-Cδ blocking mAbs or control mAb (ctl). Results are mean ± SD of TNFα secretion in culture duplicates and are representative of three independent experiments alternatively performed with the 4–29 or 5–4 clone. (B) MRC5 cells were infected (open triangles) or not (closed squares) with CMV and incubated with an anti-CMV 4-29 or a Vγ9Vδ2 T cell clone. T cells alone were also incubated with an agonistic anti-CD3 mAb (10 μg/ml, UCHT1, closed circles). Every 2 h, cells were harvested and stained with an anti-Cδ mAb. Results are presented as the MFI of cells and are representative of two independent experiments. (C) CMV-reactive 4-29 clone was cultured on MRC5 or TAP-deficient BRE cells either uninfected (white bars) or infected with intact (gray bars) or heat-inactivated CMV (black bars). Results are mean ± SD of TNFα secretion in culture triplicates and are representative of three independent experiments performed with 4–29 or 5–4 clones.
We investigated next whether the interaction of Vδ2neg γδ TCR with CMV-infected cells required antigenic presentation by MHC class I molecules, which have been shown to be dispensable for cognate ligand recognition by γδ TCR (8). For this purpose, we used an SV40-transformed fibroblastic cell line (BRE-169) genetically deficient for the transport-associated protein-1 (TAP-1), a peptide transporter that is critical for peptide loading on and subsequent surface expression of MHC class I molecules (23). Preliminary experiments demonstrated that this cell line was permissive for CMV infection (unpublished data). Despite their lack of MHC class I expression, 4 d after their infection with CMV, BRE-169 cells were as potent as MRC5 in triggering the production of TNFα by Vδ2neg γδ T cells (Fig. 3 C). As with MRC5, no activation of γδ T cell clones was observed when BRE cells were incubated with heat-inactivated CMV.
Vδ2neg and Vδ2pos γδ T cells express different patterns of homing receptors
Vδ2neg γδ T cells are thought to be mainly located within tissues under physiological conditions, and their expansion observed in the peripheral blood of transplanted patients during CMV infection could be a consequence of their vigorous activation in infected sites. Identification of these tissues would be instrumental to further understanding the antiviral functions of Vδ2 γδ T cells, but obvious ethical reasons limit in situ studies in transplanted patients. Hence, we determined the pattern of expression of homing receptors by Vδ2pos and Vδ2neg γδ T cells through ex vivo staining of circulating lymphocytes from 10 CMV-infected transplanted patients. CCR5, α4β7 integrin, CXCR4, CCR6, and to a lesser extent CXCR3 and CCR9 were expressed on variable fractions of Vδ2pos γδ T cells (Fig. 4 A). In contrast, Vδ2neg γδ T cells were negative for 8 out of the 10 chemokine receptors tested. Only CXCR4 and intestinal homing receptor α4β7 integrin expression were detected in a significant fraction of cells in a majority of patients. Interestingly, expression of CCR9, the receptor for thymus-expressed chemokine (TECK)/CCL25 and marker of small intestine homing capacity of lymphocytes (24), was found in a fraction of cells ranging from 11 to 30% in six patients. Of note, a population of CCR9high Vδ2neg γδ T cells was clearly identified in some patients (unpublished data). Thus, although chemokine receptor expression by Vδ2neg γδ T cells was limited to CXCR4 and CCR9, that of Vδ2pos γδ T cells was much more diverse with especially high percentages of CCR5 positive cells. Expression of CCR9 and β7 integrin was also found on the Vδ2neg γδ T cell clones displaying a reactivity against CMV-infected fibroblasts (Fig. 4 B). CCR9 expression on 5–4 and 4–13 clones was weak, but sufficient to transduce a signal because incubation of these clones with TECK/CCL25 induced cytoskeleton reorganization as detected by F-actin polymerization assay (Fig. 4 C). All these findings suggest an Ag-driven expansion of Vδ2neg γδ T cells within intestinal mucosa, which is also a frequent site of CMV multiplication (25), before their passage in the periphery.
Figure 4.

Pattern of chemokine and homing receptor expression by Vδ2neg and Vδ2pos T cells. (A) PBMCs from 10 CMV-infected transplanted patients were labeled with an anti-Cδ mAb, an anti-Vδ2 mAb, and each specific mAb directed against chemokine or homing receptors, before being analyzed by flow cytometry. For one patient, the percentage of positive cells for each receptor was evaluated among both Vδ2neg γδ T cells and Vδ2pos γδ T cells. Results are presented as the percentage of positive cells in each patient depicted by a specific symbol. The mean expression for each receptor appears as a short line. (B) Three different CMV-specific Vδ2neg γδ T cell clones were stained with anti–β7-integrin or anti-CCR9 mAbs (thin line) or with a control mAb (bold line), and analyzed by flow cytometry. (C) Polymerization of F-actin in response to TECK/CCL25 was tested on two Vδ2neg γδ T clones by flow cytometry as described in Materials and methods.
Anti–CMV-reactive Vδ2neg γδ T cells also recognize intestinal tumor epithelial cells
The aforementioned results were consistent with the well-established association of Vδ1 γδ T cells with intestinal epithelia (26). These cells have been studied particularly for their increased frequencies in various epithelial tumors (11, 27) and their ability in vitro to kill tumor intestinal epithelial cells (6). Because one hallmark of γδ T cells is their recognition of broadly distributed antigens (8), we tested whether these cells could recognize both CMV-infected cells and tumor intestinal epithelial cell lines. As shown in Fig. 5 and Table II, anti-CMV Vδ2neg γδ T cell clones were strongly activated when cocultured with the colon tumor epithelial cell lines Caco2 and HT29. Latent CMV infection of these cell lines was ruled out as CMV DNA could not be detected by PCR (unpublished data). Under these culture conditions, Vδ2neg γδ T cells produced TNFα and IFNγ (Fig. 5 A), and displayed a FasL-, TNFα-independent and perforin-dependent cytotoxic activity toward the target cells (Fig. 5 B). In contrast, Vδ2neg γδ T cell clones did not respond to other transformed cell lines from diverse origins (i.e., Daudi [Burkitt EBV] or THP1 [monocytoid leukemia]; Fig. 5 A). All anti-CMV Vδ2neg γδ T cell clones tested that express distinct Vγ or Vδ chains, but not Vγ9Vδ2 T cell clones, were stimulated by HT29 (Fig. 5 C). This stimulating effect increased with the number of HT29 cells added in the coculture and was prevented by semi-permeable membrane separation between HT29 and T cells (Fig. S1, A and B, available at http://www.jem.org/cgi/content/full/jem.20041851/DC1). As with CMV-infected cells, the Vδ2neg γδ T cell response evoked by HT29 involved TCR engagement because it was associated with a down-modulation of TCR surface expression (Fig. 5 D) and was abrogated by an anti-CD3 or anti-Cδ–blocking mAbs (Fig. S1 C). To test if this reactivity was an attribute of transformed epithelial cells, the activation of Vδ2neg γδ T cells by tumor versus normal epithelial cells was compared. As shown in Fig. 5 E, normal epithelial cells isolated from fetal colon (FHC) or small intestine (HIEC) clearly failed to stimulate TNFα secretion by Vδ2neg γδ T cells in sharp contrast with their tumoral counterparts HT29, CaCo2, or HeLa tumor cells. The latter data strongly suggested that Vδ2neg γδ T cells recognized a self-antigen whose expression on target cells is either linked to CMV infection or malignant transformation.
Figure 5.

Anti–CMV-reactive Vδ2neg γδ T cells are also reactive against intestinal tumor epithelial cells. (A) Vδ2neg γδ T cell clones were cultured in medium alone, with U373 cells uninfected or infected with CMV, with PMA and ionomycin, with HT29 or Caco2 monolayers, or with Daudi or THP-1 cells. All cells were harvested, permeabilized, and labeled with anti-IFNγ and anti-TNFα mAbs. Percentages correspond to cells that expressed TNFα and/or IFNγ. Results shown are representative of experiments performed with four different clones. (B) As in Fig. 2 B with HT29 cells instead of FSF. The same results were obtained with the 5–4 clone. (C) Two Vδ2neg γδ clones (4–29 and 5–4) or one control Vγ9Vδ2 T cell clone were incubated in medium alone (white bars), on HT29 cells (gray bars), or with PMA and ionomycin (black bars). (D) The 4–29 anti-CMV γδ T cell clone was incubated in medium alone, with an agonistic anti-CD3 mAb (10 μg/ml, UCHT1) or with HT29 cells. At indicated times, cells were harvested and stained with an anti-Cδ mAb. Results are presented as the MFI ± SD of culture triplicates and are representative of two independent experiments. (E) The 4–29 T cell clone was cultured in medium alone, with PMA and ionomycin or on normal (HIEC and FHC) or tumor (HeLa, HT29, CaCo-2) intestinal epithelial cell lines. Epithelial cells were also cultured in medium alone. After 6 h of incubation, TNFα secretion was quantified in culture supernatant.
Table II.
Reactivity of Vδ2neg γδ T cells against tumor epithelial cells is independent of MICA expression
| TNFα production (pg/ml)a
|
||||||||
|---|---|---|---|---|---|---|---|---|
| MICAneg cell lines
|
MICApos cell lines
|
|||||||
| Clones | V segment usage | SK-CO1 | HUH-7 | HUTU-80 | SK-MES | HeLa | HT-29 | Caco-2 |
| 2-6 | Vγ2,3 Vδ1 | <5 | 70 | 414 | 7 | 40 | 98 | 103 |
| 4-13 | Vγ4 Vδ5 | <5 | <5 | <5 | <5 | 15 | 220 | 333 |
| 4-29 | Vγ4 Vδ5 | 70 | 91 | <5 | <5 | 281 | 546 | 615 |
| 5-4 | Vγ9 Vδ1 | 105 | 34 | 30 | <5 | 388 | 428 | 543 |
| 5-15 | Vγ8 Vδ1 | 8 | 109 | 324 | 223 | 344 | 54 | 36 |
3 × 104 cells/well of five different Vδ2neg γδ T cell clones were cultured for 4 h on monolayers of seven tumor epithelial cell lines expressing MICA or not. TNFα concentration was measured in culture supernatant by ELISA. Results are the mean of culture triplicates and SD was always <15% of the mean value (unpublished data).
Recognition by Vδ2neg γδ T cells of CMV-infected cells or tumor epithelial cell lines is not mediated by NKG2D ligands
Vδ1 γδ T cells isolated from epithelial tumors have been shown to recognize the stress protein MICA, which is overexpressed on the surface of tumor epithelial cells (11). The receptor for this molecule on intraepithelial lymphocytes is NKG2D, which also interacts with UL16-binding proteins (ULBP1, 2, and 3). We thus wondered whether MICA/B or ULBP could be ligands for the CMV- and epithelial cell–reactive Vδ2neg γδ T cells through recognition of NKG2D that we found constantly expressed on Vδ2neg γδ T cells (unpublished data). An anti-NKG2D–blocking mAb or the NKG2D-Fc molecule added to cocultures of Vδ2neg γδ T cell clones, along with HT29 tumor cells or CMV-infected MRC5 cells, did not affect target cell killing (Fig. 6), while significantly decreasing lysis of MICA+ K562 cells by NKG2D+ NKL; inhibition of NKL-mediated K562 lysis was partial probably because other NKRs than NKG2D might be involved in activation of NKL cytotoxicity by K562. To confirm the aforementioned results, several epithelial tumor cell lines differing by their expression level of NKG2D ligands were tested for their ability to activate Vδ2neg γδ T cell clones: HeLa (cervical carcinoma); HT29, CaCo2, and SK-CO1 (all colon adenocarcinoma); HUTU-80 (duodenal carcinoma); SK-MES (lung carcinoma); HUH-7 (hepatocellular carcinoma); and CMV-infected FSFs. All cells were negative for ULBP1, positive for ULBP2 except CMV-infected FSFs, negative for ULBP3 except HT29 cells, and positive for MICA except HUH-7, SK-CO1, and CMV-infected FSFs (Fig. S2, available at http://www.jem.org/cgi/content/full/jem.20041851/DC1). Two MICA, ULBP1, 3-negative epithelial cell lines (SK-CO1 and HUH-7) stimulated at least two of the five Vδ2neg γδ T cell clones tested (Table II). Moreover, whereas all clones tested produced TNFα when cultured with HT29, most of them were not stimulated when cultured with HUTU-80 or SK-MES that clearly expressed MICA and ULBP2. Finally, CMV-infected FSFs did not express any of the NKG2D ligands tested, whereas they activated Vδ2neg γδ T cell clones well (Table I). These results established that NKG2D ligands were neither necessary, nor sufficient to activate anti-CMV Vδ2neg γδ T cells, and they support our hypothesis that alternative recognition processes of epithelial tumors by Vδ1 T cells must exist (11).
Figure 6.

NKG2D is not involved in the activation of Vδ2neg γδ T cells by CMV-infected fibroblasts or HT29. Vδ2neg γδ T cell clones were incubated with 51Cr-labeled CMV-infected or uninfected FSF or HT29, in medium alone or with a control or a blocking anti-NKG2D mAb (20 μg/ml), or with the NKG2D-Fc or control ICAM2-Fc chimeric molecules (both at 20 μg/ml). Effector/target ratio was 10/1. As positive control of blocking reagent efficiency, 51Cr-labeled MICA+ K562 cells were incubated with NKG2D+ NKL cells at a 10/1 effector/target ratio. Results are expressed as mean ± SD of specific lysis in culture triplicates and are representative of three independent experiments performed with 4–29 and 5–4 T cell clones.
High frequency of CMV- and tumor epithelial cell-reactive Vδ2neg γδ T cells among patients' γδ T cells
To analyze the frequency of Vδ2neg γδ T cells reactive against tumor and CMV-infected cells among peripheral T cells of CMV-infected transplanted patients, polyclonal γδ T cell lines from five CMV-infected transplanted patients were generated with PHA, IL-2, and allogeneic PBMCs. Such polyclonal activation has previously been shown to maintain the initial repertoire of T cells (28). The reactivity of these polyclonal γδ T cell lines against HT29 or CMV-infected FSFs was quantified by detecting the induced expression of membrane CD107a, a highly sensitive marker of cytotoxic activity (29). When incubated for 6 h on HT29 or CMV-infected FSF monolayers, a considerable proportion of Vδ2neg γδ T cells within the polyclonal cell lines displayed a specific cytolytic activity as evidenced by their strong CD107a surface expression (Fig. 7). Uninfected FSFs only induced a minimal expression of CD107a on T cells. Of note, the Vδ2pos T cells contained in the cell lines did not express CD107a in the presence of HT29 or CMV-infected FSFs, highlighting the discrete specificity of Vδ2neg T cell activation. The mean ± SD percentages of CMV/HT29-reactive Vδ2neg γδ T cells for the five tested patients were 16.9 ± 13.1% and 11.3 ± 13.7%, respectively (Fig. 7 B, top). For comparison, the frequency of these cells in a naive CMV repertoire of γδ T cells was studied in blood donors with a negative serology for CMV. The low number of Vδ1pos and Vδ3pos γδ T cells present in the peripheral blood of donors were sorted from 108 PBMCs and expanded in the same conditions as described previously for obtaining patients' Vδ2neg polyclonal cell lines. As shown in Fig. 7 B (bottom), CMV/HT29-reactive Vδ2neg γδ T cells were virtually absent from the three CMV seronegative blood donors analyzed (mean ± SD of CD107apos cells: 2.1 ± 1.8% and 2.1 ± 1.6%, respectively). These results establish the in vivo relevance of the Vδ2neg γδ T cell reactivity against CMV and tumor cells, and the important selection of these cells under viral pressure in allograft recipients.
Figure 7.

Anti-CMV and antitumor reactivity of polyclonal γδ T cell lines isolated from CMV+ transplanted patients and CMV− blood donors. Polyclonal γδ T cell lines were generated from five different CMV-infected transplanted patients (TP 1–5) and polyclonal Vδ2neg γδ T cell lines were generated from three different blood donors (BD 1–3). These lines were incubated in medium alone or on monolayers of HT29 or of CMV-infected or uninfected FSF in the presence of an anti-CD107a mAb. After 6 h, T cells were stained with anti-Cδ and anti-Vδ2 mAbs and analyzed by flow cytometry. (A) Staining profiles obtained with the TP1 γδ T cell line. Indicated are percentages of CD107a+ cells among the Vδ2neg (top left quadrant) or Vδ2pos (top right quadrant) populations. (B) Percentages of CD107a+ cells among Vδ2neg γδ T cells for each tested transplanted patient or blood donor.
Discussion
Because the knowledge on the functions, antigenic specificity, and precise localization of Vδ2neg γδ T cells is currently limited, the marked expansion of these cells in the peripheral blood of transplant patients undergoing active CMV infection provides a unique opportunity to examine their role within the immune system.
In this study, we showed that fibroblasts infected with CMV triggered the production of large amounts of TNFα by Vδ2neg γδ T cells through a mechanism involving TCR engagement but not MHC presentation. In contrast, we could never observe activation of Vγ9Vδ2 T cells in the presence of CMV-infected cells. This result agrees with the absence of Vγ9Vδ2 T cell expansion in the peripheral blood of patients suffering from CMV infection (17). Anti–CMV-reactive Vδ2neg γδ T cell clones expressed diverse Vδ or Vγ chains, such as MIC and CD1c-specific Vδ1 T cells, which express diverse Vγ chains and nevertheless recognize the same antigen (6, 7). Together, these observations suggest common functions for different subsets of the Vδ2neg γδ T cell population (Vδ1, Vδ3, Vδ5) that clearly differ from that of Vγ9Vδ2 T cells.
Vδ1 T cells are the main subset of γδ T cells found in tissues, particularly in epithelial tissue and spleen. The localization of other Vδ2neg γδ T cell subsets remains more elusive and they are virtually undetectable in the peripheral blood under normal conditions. The observation of a major expansion of rare subsets, such as Vδ3 or Vδ5 T cells, in the peripheral blood of CMV-infected transplant patients indicates that these cells probably originate from tissues, where they are activated after CMV infection and redistributed thereafter into the blood. Accordingly, the only tissue-specific homing receptors significantly detected on Vδ2neg γδ T cells in the peripheral blood of CMV-infected transplant patients are α4β7 and CCR9, two homing receptors defining gut tropism of effector lymphocytes, including γδ T cells (24, 30). Indeed, marked diminution of γδ T cells in gut epithelia has been found in mice lacking CCR9 (31). It is noteworthy that the panel of chemokine receptors expressed by Vδ2neg γδ T cells is much more restricted than that expressed by Vγ9Vδ2 T lymphocytes, underlying intestinal origin of these cells.
The antiviral activity of Vδ2neg γδ T cells, which can kill CMV-infected cells, is at least in part mediated by a perforin–granzyme-dependent lytic activity. Notably, these T cells also limit the propagation of CMV in cell cultures through their secretion of IFNγ, a renown inhibitor of viral replication (32). Because Vδ2neg γδ T cell expansion in transplant patients is concomitant to the resolution of CMV infection (18), it is tempting to speculate that their ability to kill CMV-infected cells and to limit CMV propagation in vitro is probably involved in a protective antiviral response, although no data thus far directly supports this hypothesis in vivo. Because intestinal epithelium is a frequent site of CMV replication (25), Vδ2neg γδ T cells might well be activated and expand locally in contact with CMV-infected cells and participate to an efficient, coordinated mucosal host defense. This assumption is supported by a study performed in γδ T cell–deficient mice demonstrating a protective role of liver γδ T cells in host defense against murine CMV (20).
Homing of γδ T cells to mucosal effector sites has been related to their widely illustrated antitumoral activity in vivo and in vitro (9). Vδ1pos T cells were particularly shown to infiltrate carcinoma and to kill tumor epithelial cells (11, 27). Accordingly, we report in the present study that anti-CMV reactive Vδ2neg γδ T cells were activated by several epithelial cell lines in a TCR-dependent fashion, demonstrating an antiviral and antitumor cross-reactivity of these cells. Noteworthy, Vδ2neg γδ T cells did not recognize normal intestinal epithelial cells, indicating their specificity toward transformed cells and suggesting their role in antitumor immunity, although in vivo studies are required to support this hypothesis.
In agreement with the absence of Vδ2neg γδ T cell expansion in transplant patients suffering from VZV, HSV, or EBV infections (17), no reactivity of Vδ2neg γδ T cells was observed in vitro against these herpesviruses. This suggests that the ligand recognized by Vδ2neg γδ T cells is either a specific CMV structure or a cellular antigen selectively induced in CMV-infected cells. Self-recognition of a stress molecule is consistent with both previous descriptions of CD1c and MIC as ligands for Vδ1 T cell lines and the response of Vδ2neg γδ T cells to both CMV-infected and tumor epithelial cells. CD1c involvement in Vδ2neg γδ T cell activation can be excluded because neither infected fibroblasts nor target epithelial cell lines expressed this antigen (Fig. S2, available at http://www.jem.org/cgi/content/full/jem.20041851/DC1), which is exclusively found on antigen-presenting cells (7). The involvement of MICA was excluded using blocking mAbs and showing reactivity of Vδ2neg γδ T cell clones toward CMV-infected FSFs or some epithelial cell lines that do not express MICA. Conceivably, the danger signal recognized by Vδ2neg γδ T cells may be a common self-antigen induced on epithelial cell surfaces by different types of stresses, such as CMV infection or cell transformation. However, a molecular mimicry between a CMV antigen and a structure present on tumor epithelial cell surface cannot be excluded as CMV evolved with its host by incorporating numerous cellular genes in its own genome (33).
Co-stimulation molecules involved in anti-CMV and antitumor-reactive Vδ2neg γδ T cell activation are still to be identified. NKG2D-mediated recognition of intestinal epithelial cells by intraepithelial lymphocytes has been reported in several previous studies (11, 34, 35). NKG2D has also been shown to costimulate CMV-specific CD8 αβ T cells (36). In sharp contrast, cytolytic activity of Vδ2neg γδ T cells against CMV-infected or tumor cells did not involve NKG2D, at least in vitro. Nevertheless, we still cannot exclude that primary activation of naive Vδ2neg γδ T cells in vivo might require NKG2D engagement, but that subsequent induction of committed cell effector functions by target cells might depend exclusively on TCR or on other coreceptors.
The anti-CMV and antitumor reactivity of a significant proportion of Vδ2neg γδ T cells within polyclonal γδ T cell lines isolated from patients highlighted the relevance of this population in vivo. The expansion of these cells related to an acute CMV infection, the association of this expansion with the resolution of CMV infection, and the ability of Vδ2neg γδ T cells to kill CMV-infected cells and limit CMV replication in vitro further support a direct implication of γδ T cells in the anti-CMV immune response. Recognition of their ligand independent of MHC presentation (a) most probably make those cells suitable antiviral immune effectors to neutralize a pathogen such as CMV that down-modulates MHC class I molecule expression, and (b) encourage their use in the development of future immunotherapy protocols for CMV infections once the molecular target recognized by Vδ2neg γδ T cells has been identified.
Materials and Methods
Patients
After giving their informed consent, kidney and lung allograft recipients receiving standard immunosuppressive regimen were included in the present study approved by the local medical ethics commitee. They developed a CMV infection associated with an expansion of Vδ2neg γδ T cells and diagnosed based on CMV pp65 antigen positivity (>1 cell per 2 × 105 leukocytes) in peripheral blood leukocytes (detection kit from Argene Biosoft). Blood samples were taken either during the CMV infection or soon after its resolution.
Antibodies (Abs) and cell lines
The following mAbs were purchased from Beckman Coulter: anti-Cδ (IMMU 510), anti-Vδ1 (R9.12), anti-Vδ2 (IMMU 389), anti-Vδ3 (P11.5B), anti-Vγ9 (IMMU 360), anti-Vγ8 (R4.5), and anti-Vγ2/3/4 (23D12). Anti-Vγ4 mAb (94) was provided by M. Bonneville (INSERM U463, Nantes, France). Anti–TNF-α (Mab11), anti–IFN-γ (25723.11), antiperforin (δG9), and anti-CD107a (H4A3) were purchased from BD Biosciences and anti–granzyme B (2C5) was obtained from Santa Cruz Biotechnology, Inc. Neutralizing anti-CD1c mAb (F10/21A3) was provided by S. Porcelli (Yeshiva University, New York, NY). The anti–HLA class I mAb (W6/32) was purchased from DakoCytomation. The anti-IFNγ (4S.B3) blocking mAb was purchased from BD Biosciences, the anti-TNFα (Infliximab) blocking mAb was obtained from Schering-Plough Laboratory, the anti-NKG2D (149810) blocking mAb, the NKG2D-Fc, and ICAM2-Fc molecules were obtained from R&D Systems, and the anti-Fas (ZB4) blocking mAb was obtained from Beckman Coulter. The anti-MICA mAb (SR104; reference 34) was a gift from S. Caillat-Zuckman (Hôpital Necker, Paris, France). The anti-CD3 neutralizing mAb (OKT3) was provided by Janssen-Cilag and the anti-CD3 (UCHT1) agonist was a gift from C. Hivroz (Institut Curie, Paris, France). Anti-CCR1, 3, 5, 6, 7, 9, and anti-CXCR3, 4, 5, 6 mAbs were obtained from R&D Systems; anti-CLA and anti-integrin β7 was obtained from BD Biosciences; and anti-integrin α4 was obtained from Beckman-Coulter. IFNγ was purchased from Bender MedSystem and IL2 was given by Chiron. Primary human FSFs were a gift from C. Pain (INSERM EMI0217, Bordeaux, France). TAP1−/− BRE-169 cells is a TAP-deficient skin fibroblast cell line (23) immortalized by transformation with a vector expressing SV40 T and t antigens and human telomerase. Normal epithelial cell lines were isolated from human fetal colon (FHC; no. CRL-1831; American Type Culture Collection) and small intestine (HIEC; reference 37) was a gift from J.F. Beaulieu (University of Sherbrook, Quebec, Canada). The WR19L cells transfected with the cDNA coding for the Fas-L (1A12 cells; reference 38) was a gift from S. Nagata (Osaka University Medical School, Osaka, Japan). All cell lines are repeatedly screened for mycoplasma contamination and found to be negative.
Generation of γδ polyclonal cell lines and clones
γδ T cell subpopulations were sorted from patients' or healthy donors' PBMCs using anti-Cδ, anti-Vδ1, or anti-Vδ3 mAbs and magnetic beads coated with sheep anti–mouse Ig (Dynal). Bead-adherent cells were either seeded at 0.5 cell/well for direct cloning or expanded in bulk cultures to generate polyclonal γδ cell lines with 1,000 U/ml rIL-2, 1 μg/ml PHA-L (Sigma-Aldrich), and irradiated allogeneic PBMCs (35 Gy). After 14 d of culture, T cell clones or lines were phenotyped by immunofluorescent staining and analyzed by flow cytometry on a FACSCalibur apparatus (BD Biosciences) as described elsewhere (17).
Infection of MRC5 and FSF
CMV suspensions were usually produced with the clinical strain TB40/E (a gift from C. Sinzger, University of Tubingen, Tubingen, Germany) as previously described (17). When specified, the laboratory strains AD169 and Towne were also used. HSV and VZV suspensions were provided by S. Michelson (Institut Pasteur, Paris, France) and B.M. Imbert (CHU Nantes, Nantes, France), respectively. When necessary, virus suspensions were inactivated by heating at 56°C for 30 min. Subconfluent monolayers of MRC5, U373, primary FSF, or TAP1−/− BRE-169 cells were incubated with CMV suspension at a multiplicity of infection (MOI) of 1 for 2 h, washed, and cultured for 4 d at 37°C. Infected cell layers were washed before being used for coculture experiments with γδ T cell clones. Control for viral infection were performed with CMV (mAb E13), VZV (mAb 2013), and HSV (mAb CHA-437)-specific detection kits according to manufacturer's instructions (Argene Biosoft).
Detection of cytokine production
γδ T cell clones (3 × 104/well) were incubated with monolayers of epithelial cell lines or virus-infected or noninfected MRC5, FSF, U373, or TAP−/− BRE-169 cells for 6 h at 37°C. As a positive control for activation, γδ T cell clones were incubated with 25 ng/ml PMA (Sigma-Aldrich) and 1 μg/ml ionomycin (Sigma-Aldrich). TNF-α released into the supernatant was quantified by ELISA according to the manufacturer's recommendations (Bender Medsystems). For intracellular staining of TNFα and IFNγ, 10 μg/ml brefeldin A (Sigma-Aldrich) was added to the coculture for the last 5 h. Cells were harvested, washed, surface labeled with the anti-Cδ mAb, fixed with FACS lysing solution, and permeabilized in FACS permeabilizing solution (BD Biosciences) followed by staining with anti-TNFα and/or anti-IFNγ mAbs and analysis by flow cytometry.
Cell cytotoxicity assays
Cytolytic activity of γδ T cells was measured by a standard 51Cr-release assay. In brief, epithelial cells and CMV-infected or noninfected fibroblasts were labeled with 51Cr (1.85 MBq/106 cells), for 1 h at 37°C, then 3,000 cells/well were incubated with Vδ2neg γδ T cell clones at the indicated effector/target ratios. In control experiments, WEHI and Jurkat cells were labeled with 51Cr and incubated with 50 pg/ml TNFα or the Fas-ligand+ WR19L cells (1A12 cells), respectively. After 4 h at 37°C, 51Cr released in supernatant was measured and the percent of specific 51Cr release was calculated as previously described (39). The spontaneous 51Cr release from target cells in medium alone was always <15% of the control maximal 51Cr release obtained with 1% Triton X-100. For degranulation experiments, the effector T cells were pretreated with 20 mM strontium chloride (Sigma-Aldrich) or sodium chloride as control at 37°C overnight and washed, and cell viability was evaluated by eosin dye exclusion.
Cytotoxic potential of γδ T cell lines and clones was also measured by the flow cytometric CD107a assay (as described previously (29). In brief, 6-h cocultures of γδ T cells with target cells were performed as for cytokine quantification, except that anti-CD107a mAb was added at the onset of the culture. Brefeldin A was added in the coculture for the last 5 h, and cells were harvested, surface stained with anti-Cδ and anti-Vδ2 mAbs, and analyzed by flow cytometry.
TCR down-modulation experiments
The assay for down-modulation of TCR–CD3 was performed as described previously (22). In brief, T cell clones (105 cells/well) were cultured on HT29 or MRC5 monolayers infected or not with CMV, or with a soluble anti-CD3 agonist mAb at 10 μg/ml for 0, 2, 4, and 6 h. T cells were collected, double stained with anti-Cδ mAb and propidium iodide (Sigma-Aldrich), and analyzed by flow cytometry. The down-modulation of TCR–CD3 complexes at T cell surfaces was expressed as the mean fluorescence intensity (MFI) of the TCR γδ staining as a function of time.
CMV multiplication assay
Subconfluent FSF cell monolayers were cultured for 2 h with a CMV suspension at a low MOI of 0.1. Cells were washed twice to remove free virus and cultured with γδ T cell clones (5 × 104 cells/well in 100 μl) or IFN-γ at indicated concentrations. After 4 d of culture, infectious CMV particles released were quantified in culture supernatants by a standard plaque assay (40).
F-actin polymerization assay
Intracellular filamentous (F)-actin polymerization was tested as previously described (41). In brief, T cells (8 × 106/ml) were incubated at 37°C with or without 300 ng/ml TECK/CCL25, and added at indicated times to 400 μl of the assay buffer containing 4 × 10−7 mol/l FITC-labeled phalloidin, 0.5 mg/ml L-α-lysophosphatidylcholine (both obtained from Sigm-Aldrich), and 4.5% formaldehyde in PBS. Fixed cells were analyzed by flow cytometry, and percentage change in MFI was determined for each sample.
Online supplemental material.
Fig. S1 further details the dose-dependent (SIA), contact-dependent (SIB), and TCR-dependent (SIC) activation of CMV-reactive γδ T cell clones by the HT29 cells. Fig. S2 depicts the expression of HLA-1, MICA, UCBPs, and CD1 by all target cells used in the present study. Table S1 displays the Vδ-Jδ junctional sequences of all Vδ2neg γδ T cell clones. Online supplemental material is available at http://www.jem.org/cgi/content/full/jem.20041851/DC1.
Acknowledgments
We thank S. Michelson for critical reading of the text, the Etablissement Français du Sang d'Aquitaine for blood samples, the nursing staffs of kidney and lung transplant departments, I. Garrigue and M.-E. Lafon for virological analyzes, and D. Roumanes for technical assistance.
This work was supported in part by institutional grants from CNRS and Ministère de l'Enseignement Supérieur et de la Recherche, and by specific grants from Roche Organ Transplantation Research Foundation (ROTRF, grant no. 465708914), Ligue Nationale contre le Cancer (Comité Départemental de la Dordogne), Association pour la Recherche contre le Cancer, Etablissement Français des Greffes, and INSERM (ATC Biothérapie Immunité anti-infectieuse). D. Dlubek was supported by INSERM (poste vert).
The authors have no conflicting financial interests.
Abbreviations used: FSF, foreskin fibroblast; MFI, mean fluorescence intensity; MICA, MHC class I–related chain A, MOI, multiplicity of infection; TAP, transport-associated protein; TECK, thymus-expressed chemokine; ULBP, UL16- binding protein; VZV, varicella zoster virus.
F. Halary and V. Pitard contributed equally to this work.
References
- 1.Constant, P., F. Davodeau, M.A. Peyrat, Y. Poquet, G. Puzo, M. Bonneville, and J.J. Fournié. 1994. Stimulation of human γδ T cells by nonpeptidic mycobacterial ligands. Science. 264:267–270. [DOI] [PubMed] [Google Scholar]
- 2.Tanaka, Y., C.T. Morita, E. Nieves, M.B. Brenner, and B.R. Bloom. 1995. Natural and synthetic non-peptide antigens recognized by human γδ T cells. Nature. 375:155–158. [DOI] [PubMed] [Google Scholar]
- 3.Kunzmann, V., E. Bauer, J. Feurle, F. Weissinger, H.P. Tony, and M. Wilhelm. 2000. Stimulation of gammadelta T cells by aminobisphosphonates and induction of antiplasma cell activity in multiple myeloma. Blood. 96:384–392. [PubMed] [Google Scholar]
- 4.Bukowski, J.F., C.T. Morita, and M.B. Brenner. 1999. Human gamma delta T cells recognize alkylamines derived from microbes, edible plants, and tea: implications for innate immunity. Immunity. 11:57–65. [DOI] [PubMed] [Google Scholar]
- 5.Scotet, E., L.O. Martinez, E. Grant, R. Barbaras, P. Jeno, M. Guiraud, B. Monsarrat, X. Saulquin, S. Maillet, J.P. Esteve, et al. 2005. Tumor recognition following Vgamma9Vdelta2 T cell receptor interactions with a surface F1-ATPase-related structure and apolipoprotein A-I. Immunity. 22:71–80. [DOI] [PubMed] [Google Scholar]
- 6.Groh, V., A. Steinle, S. Bauer, and T. Spies. 1998. Recognition of stress-induced MHC molecules by intestinal epithelial γδ T cells. Science. 279:1737–1740. [DOI] [PubMed] [Google Scholar]
- 7.Spada, F., E. Grant, P. Peters, M. Sugita, A. Melian, D. Leslie, H. Lee, E. van Donselaar, D. Hanson, A. Krensky, et al. 1999. Self-recognition of CD1 by γδ T cells: implications for innate immunity. J. Exp. Med. 191:937–948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Carding, S.R., and P.J. Egan. 2002. Gammadelta T cells: functional plasticity and heterogeneity. Nat. Rev. Immunol. 2:336–345. [DOI] [PubMed] [Google Scholar]
- 9.Ferrarini, M., E. Ferrero, L. Dagna, A. Poggi, and M.R. Zocchi. 2002. Human gammadelta T cells: a nonredundant system in the immune-surveillance against cancer. Trends Immunol. 23:14–18. [DOI] [PubMed] [Google Scholar]
- 10.Wilhelm, M., V. Kunzmann, S. Eckstein, P. Reimer, F. Weissinger, T. Ruediger, and H.P. Tony. 2003. Gammadelta T cells for immune therapy of patients with lymphoid malignancies. Blood. 102:200–206. [DOI] [PubMed] [Google Scholar]
- 11.Groh, V., R. Rhinehart, H. Secrist, S. Bauer, K.H. Grabstein, and T. Spies. 1999. Broad tumor-associated expression and recognition by tumor-derived gamma delta T cells of MICA and MICB. Proc. Natl. Acad. Sci. USA. 96:6879–6884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Maeurer, M.J., D. Martin, W. Walter, K. Liu, L. Zitvogel, K. Halusczcak, H. Rabinowich, R. Duquesnoy, W. Storkus, and M.T. Lotze. 1996. Human intestinal Vδ1+ lymphocytes recognize tumor cells of epithelial origin. J. Exp. Med. 183:1681–1696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Autran, M., F. Triebel, C. Katlama, W. Rozenbaum, T. Hercend, and P. Debré. 1989. T cell receptor γδ lymphocyte subsets during HIV infection. Clin. Exp. Immunol. 75:206–210. [PMC free article] [PubMed] [Google Scholar]
- 14.De Paoli, P., D. Gennari, P. Martelli, V. Cavarzerani, R. Comoretto, and G. Santini. 1990. γδ T cell receptor-bearing lymphocytes during Epstein-Barr virus infection. J. Infect. Dis. 161:1013–1016. [DOI] [PubMed] [Google Scholar]
- 15.Maccario, R., P. Comoli, E. Percivalle, D. Montagna, F. Locatelli, and G. Gerna. 1995. Herpes simplex virus-specific human cytotoxic T-cell colonies expressing either γδ or αβ T-cell receptor: role of accessory molecules on HLA-unrestricted killing of virus-infected targets. Immunology. 85:49–56. [PMC free article] [PubMed] [Google Scholar]
- 16.Reddehase, M.J. 2000. The immunogenicity of human and murine cytomegaloviruses. Curr. Opin. Immunol. 12:390–396. [DOI] [PubMed] [Google Scholar]
- 17.Déchanet, J., P. Merville, A. Lim, C. Retiere, V. Pitard, X. Lafarge, S. Michelson, C. Meric, M.M. Hallet, P. Kourilsky, et al. 1999. Implication of gammadelta T cells in the human immune response to cytomegalovirus. J. Clin. Invest. 103:1437–1449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lafarge, X., P. Merville, M.C. Cazin, F. Berge, L. Potaux, J.F. Moreau, and J. Dechanet-Merville. 2001. Cytomegalovirus infection in transplant recipients resolves when circulating gammadelta T lymphocytes expand, suggesting a protective antiviral role. J. Infect. Dis. 184:533–541. [DOI] [PubMed] [Google Scholar]
- 19.Cavanaugh, V.J., Y. Deng, M.P. Birkenbach, J.S. Slater, and A.E. Campbell. 2003. Vigorous innate and virus-specific cytotoxic T-lymphocyte responses to murine cytomegalovirus in the submaxillary salivary gland. J. Virol. 77:1703–1717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ninomiya, T., H. Takimoto, G. Matsuzaki, S. Hamano, H. Yoshida, Y. Yoshikai, G. Kimura, and K. Nomoto. 2000. Vgamma1+ gammadelta T cells play protective roles at an early phase of murine cytomegalovirus infection through production of interferon-gamma. Immunology. 99:187–194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kabelitz, D. 1999. Effector functions and control of human gammadelta T-cell activation. Microbes Infect. 1:255–261. [DOI] [PubMed] [Google Scholar]
- 22.Liu, H., M. Rhodes, D.L. Wiest, and D.A. Vignali. 2000. On the dynamics of TCR:CD3 complex cell surface expression and downmodulation. Immunity. 13:665–675. [DOI] [PubMed] [Google Scholar]
- 23.de la Salle, H., J. Zimmer, D. Fricker, C. Angenieux, J.P. Cazenave, M. Okubo, H. Maeda, A. Plebani, M.M. Tongio, A. Dormoy, and D. Hanau. 1999. HLA class I deficiencies due to mutations in subunit 1 of the peptide transporter TAP1. J. Clin. Invest. 103:R9–R13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Uehara, S., A. Grinberg, J.M. Farber, and P.E. Love. 2002. A role for CCR9 in T lymphocyte development and migration. J. Immunol. 168:2811–2819. [DOI] [PubMed] [Google Scholar]
- 25.Patra, S., S.C. Samal, A. Chacko, V.I. Mathan, and M.M. Mathan. 1999. Cytomegalovirus infection of the human gastrointestinal tract. J. Gastroenterol. Hepatol. 14:973–976. [DOI] [PubMed] [Google Scholar]
- 26.Chowers, Y., W. Holtmeier, J. Harwood, E. Morzycka-Wroblewska, and M.F. Kagnoff. 1994. The Vδ1 T cell receptor repertoire in human small intestine and colon. J. Exp. Med. 180:183–190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Choudhary, A., F. Davodeu, A. Moreau, M.A. Peyrat, M. Bonneville, and F. Jotereau. 1995. Selective lysis of autologous tumor cells by recurrent γδ tumor-infiltrating lymphocytes from renal carcinoma. J. Immunol. 154:3932–3940. [PubMed] [Google Scholar]
- 28.David-Ameline, J., A. Lim, F. Davodeau, M.A. Peyrat, J.M. Berthelot, G. Semana, C. Pannetier, J. Gaschet, H. Vie, J. Even, and M. Bonneville. 1996. Selection of T cells reactive against autologous B lymphoblastoid cells during chronic rheumatoid arthritis. J. Immunol. 157:4697–4706. [PubMed] [Google Scholar]
- 29.Betts, M.R., J.M. Brenchley, D.A. Price, S.C. De Rosa, D.C. Douek, M. Roederer, and R.A. Koup. 2003. Sensitive and viable identification of antigen-specific CD8+ T cells by a flow cytometric assay for degranulation. J. Immunol. Methods. 281:65–78. [DOI] [PubMed] [Google Scholar]
- 30.Lefrancois, L., C.M. Parker, S. Olson, W. Muller, N. Wagner, M.P. Schon, and L. Puddington. 1999. The role of β7 integrins in CD8 T cell trafficking during an antiviral immune response. J. Exp. Med. 189:1631–1638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Wurbel, M.A., M. Malissen, D. Guy-Grand, E. Meffre, M.C. Nussenzweig, M. Richelme, A. Carrier, and B. Malissen. 2001. Mice lacking the CCR9 CC-chemokine receptor show a mild impairment of early T- and B-cell development and a reduction in T-cell receptor gammadelta(+) gut intraepithelial lymphocytes. Blood. 98:2626–2632. [DOI] [PubMed] [Google Scholar]
- 32.Bodaghi, B., O. Goureau, D. Zipeto, L. Laurent, J.L. Virelizier, and S. Michelson. 1999. Role of IFN-gamma-induced indoleamine 2,3 dioxygenase and inducible nitric oxide synthase in the replication of human cytomegalovirus in retinal pigment epithelial cells. J. Immunol. 162:957–964. [PubMed] [Google Scholar]
- 33.Michelson, S. 2004. Consequences of human cytomegalovirus mimicry. Hum. Immunol. 65:465–475. [DOI] [PubMed] [Google Scholar]
- 34.Hue, S., J.J. Mention, R.C. Monteiro, S. Zhang, C. Cellier, J. Schmitz, V. Verkarre, N. Fodil, S. Bahram, N. Cerf-Bensussan, and S. Caillat-Zucman. 2004. A direct role for NKG2D/MICA interaction in villous atrophy during celiac disease. Immunity. 21:367–377. [DOI] [PubMed] [Google Scholar]
- 35.Meresse, B., Z. Chen, C. Ciszewski, M. Tretiakova, G. Bhagat, T.N. Krausz, D.H. Raulet, L.L. Lanier, V. Groh, T. Spies, et al. 2004. Coordinated induction by IL15 of a TCR-independent NKG2D signaling pathway converts CTL into lymphokine-activated killer cells in celiac disease. Immunity. 21:357–366. [DOI] [PubMed] [Google Scholar]
- 36.Groh, V., R. Rhinehart, J. Randolph-Habecker, M.S. Topp, S.R. Riddell, and T. Spies. 2001. Costimulation of CD8alphabeta T cells by NKG2D via engagement by MIC induced on virus-infected cells. Nat. Immunol. 2:255–260. [DOI] [PubMed] [Google Scholar]
- 37.Perreault, N., and J.F. Beaulieu. 1998. Primary cultures of fully differentiated and pure human intestinal epithelial cells. Exp. Cell Res. 245:34–42. [DOI] [PubMed] [Google Scholar]
- 38.Tanaka, M., T. Suda, K. Haze, N. Nakamura, K. Sato, F. Kimura, K. Motoyoshi, M. Mizuki, S. Tagawa, S. Ohga, et al. 1996. Fas ligand in human serum. Nat. Med. 2:317–322. [DOI] [PubMed] [Google Scholar]
- 39.Davodeau, F., M.A. Peyrat, M.M. Hallet, J. Gaschet, I. Houde, R. Vivien, H. Vie, and M. Bonneville. 1993. Close correlation between Daudi and mycobacterial antigen recognition by human gamma delta T cells and expression of V9JPC1 gamma/V2DJC delta-encoded T cell receptors. J. Immunol. 151:1214–1223. [PubMed] [Google Scholar]
- 40.Halary, F., A. Amara, H. Lortat-Jacob, M. Messerle, T. Delaunay, C. Houles, F. Fieschi, F. Arenzana-Seisdedos, J.F. Moreau, and J. Dechanet-Merville. 2002. Human cytomegalovirus binding to DC-SIGN is required for dendritic cell infection and target cell trans-infection. Immunity. 17:653–664. [DOI] [PubMed] [Google Scholar]
- 41.Krzysiek, R., E.A. Lefevre, J. Bernard, A. Foussat, P. Galanaud, F. Louache, and Y. Richard. 2000. Regulation of CCR6 chemokine receptor expression and responsiveness to macrophage inflammatory protein-3alpha/CCL20 in human B cells. Blood. 96:2338–2345. [PubMed] [Google Scholar]