

Abstract
The outer membrane protein PulD (secretin) of Klebsiella oxytoca is required for transport of pullulanase across this membrane. We have purified a multimeric PulD complex from an Escherichia coli strain expressing all the proteins involved in pullulanase secretion. The outer membrane-anchored lipoprotein PulS was found to copurify with PulD. The molar ratio of the two proteins is close to 1:1, and the size of the complex is ≈1 MDa. Scanning transmission electron and cryo-electron microscopy analyses showed that the purified complex is a cylindrical structure having a central cavity of ≈7.6 nm and peripheral radial spokes. Fusion of proteoliposomes containing the purified complex with a planar lipid bilayer resulted in the appearance of small, voltage-activated, ion-conducting channels. We conclude that the central cavity seen in the electron microscope is part of a large gated channel and propose that the observed current fluctuations correspond to voltage-induced, relatively minor displacements of domains in the purified complex rather than to a complete opening of the secretin channel.
Keywords: protein secretion, secreton, pullulanase
Protein secretion by Gram-negative bacteria provides one of the very few examples of the translocation of folded proteins across a lipid bilayer, in this particular case the outer membrane. Protein translocation across this membrane is presumed to involve a large, proteinaceous gated channel. Secretins, outer membrane components of a wide variety of systems for protein secretion (1–4), pilus assembly (5), and filamentous phage assembly and secretion (6) are thought to be the major subunit of such a channel. This idea originally was based on the observation that secretins form multimers composed of 10–14 subunits that are highly resistant to dissociation in SDS. Much stronger support for this idea came from recent electron microscopic studies showing that secretins form ring-like structures with a large central cavity (2–4, 7). The organization of the secretin monomers in the complex is apparently different from that of porin monomers in the well-characterized bacterial outer membrane porin trimer (8). Nevertheless, the C-terminal half of secretin monomers is predicted to contain several amphipathic β strands similar to the transmembrane segments of porins (3). The N-terminal region of secretins is presumed to face the periplasm and might be involved in substrate recognition (9), interaction with other components of the secretion machinery (10), or gating of the proposed secretin channel.
Secretins are known to interact with other components of their respective secretion machineries. The best-characterized interaction is with the secretin-specific pilot proteins that protect secretins from proteolysis and are essential for their insertion in the outer membrane (1, 2, 5, 11–13). Three pilot proteins have been shown to bind to a short C-terminal segment of their cognate secretins (12–14).
This report concerns PulD, the secretin from the pullulanase-specific step (the secreton) in the general secretory pathway of Klebsiella oxytoca. The aim was to characterize the structure of purified secretin by electron microscopy (EM) and to examine its ability to form channels that might correspond to the conduit through which pullulanase crosses the outer membrane.
MATERIALS AND METHODS
Purification of PulD.
Cells grown overnight in LB medium (15) containing 0.4% maltose were disrupted by passage through a French press (10,000 psi), and outer membranes were isolated by ultracentrifugation (1 min at 165,000 × g). The pellet was washed with 50 mM Tris (pH 7.5), 5 mM EDTA, and outer membranes were pelleted as described above. Outer membranes were solubilized in 50 mM Tris (pH 7.5), 5% 3-(N,N-dimethylmyristylammonio)propanesulfonate (SB3–14), 250 mM NaCl, 1 mM EDTA in the presence of a protease inhibitor mixture (Sigma) at a protein concentration of 10 mg/ml. After 30 min, insoluble material was removed by ultracentrifugation at 100,000 × g for 30 min. The clarified extract was loaded onto a Q-Sepharose anion exchange column (20 ml) equilibrated with buffer A (20 mM Bis-Tris-propane, pH 7.0/0.6% SB3–14/250 mM NaCl/0.1 mM EDTA). The column was rinsed with 8 vol of buffer A and eluted with a gradient of 0.25–0.5 M NaCl. Fractions were analyzed by SDS/PAGE and immunoblotting for the presence of PulD. Fractions containing PulD were pooled, concentrated by ultrafiltration (Amicon XM50), loaded on a Sephacryl S300 HR size exclusion column, and eluted with buffer C (20 mM Bis-Tris-propane, pH 7.0/0.6% SB3–14/200 mM NaCl/0.1 mM EDTA). Peak PulD-containing fractions were pooled, concentrated, and stored at −70°C.
Secreton components PulC, PulD, PulE, PulG, PulL, and PulS were detected by immunoblotting using antibodies raised against purified proteins or against MalE hybrids.
Scanning Transmission EM (STEM) Analysis.
PulD-PulS in buffer C was adsorbed for 1 min to glow-discharged thin carbon films over a thick fenestrated carbon layer on gold-coated copper grids. For mass measurement, the grids were blotted, washed on drops of distilled water, and freeze-dried at −80°C. For structural analysis, the sample on the carbon film was stained with uranyl formate (pH 4.2) for 10 sec and blotted before dehydration in air.
A Vacuum Generators STEM HB-5 interfaced to a modular computer system (Tietz Video and Image Processing Systems, Gauting, Germany) was used to record images. The dark-field micrographs of the unstained samples were recorded at 80 kV, a nominal magnification of ×200,000 (pixel size 0.8 nm), and at typical doses of 300 electrons/nm2. The 512-×-512-pixel digital images were evaluated by using the impsys program package (16).
The 512-×-512-pixel digital images of negatively stained PulD at ×500,000 were recorded at 100 kV (pixel size 0.32 nm) and doses of 2,000 ± 500 electrons/nm2. The semper program package (17) was used to select individual particles as 128-×-128-pixel subframes, which were aligned with respect to an arbitrary reference, averaged, and classified (18). Particles falling into clusters were averaged, and their angular power spectrum was inspected for angular harmonics. This procedure was repeated by using the cluster average exhibiting the clearest features as reference.
Cryo-EM Analysis.
Cryo-EM of PulD-PulS complex in buffer C was performed as described (19). Images of the PulD-PulS complex on a carbon support film were taken at a magnification of ×30,000 by using a Jeol 1200EX microscope equipped with an Oxford Instruments cryo-transfer stage. Negatives were digitized by using a Leafscan 45 scanner at a resolution of 6.67 Å/pixel. A total of 2,600 images of the PulD-PulS complex were interactively selected and sorted into classes by using multivariate statistical analysis in imagic (20). Twelve-fold symmetry was imposed to obtain a preliminary three-dimensional (3D) reconstruction from representative end-view and side-view class-average images. Iterative alignment and reproduction were performed by using spider (21).
Reconstitution of PulD into Proteoliposomes.
Fifty microliters of purified PulD-PulS complex (1 mg/ml) was mixed with 50 μl of soybean l-α-phosphatidylcholine (100 mg/ml) (Sigma) in 2% octyl glucoside. After 10 min on ice, the mixture was rapidly diluted with 4 ml of 10 mM Hepes (pH 7.4) and 400 mM KCl and incubated for 10 min before addition of Bio-Beads SM-2 (Bio-Rad; 200 mg). The suspension was gently shaken for 5 hr, the Bio-Beads were discarded, and the suspension was centrifuged at 250,000 × g in a TLA100 Beckman ultracentrifuge. The pellet was resuspended in 250 μl of 10 mM Hepes (pH 7.4), 400 mM KCl, frozen in dry ice-ethanol, and stored at −70°C. Proteoliposomes were used within 1 week of reconstitution.
Planar Lipid Bilayer Experiments.
Membranes were formed from a solution of soybean l-α-phosphatidylcholine phospholipids (Sigma) in n-decane at 30 mg/ml. Bilayers were formed across a 250-μm diameter hole by presenting a bubble of lipids in decane in front of the hole. After membrane formation, the mechanical and electrical stability was monitored before addition of proteoliposomes to the cis compartment (0.14–0.5 μg/ml final concentration). Fusion with the planar lipid bilayer was induced by imposing a salt gradient between the two chambers (400 mM KCl in the cis compartment versus 100 mM KCl in the trans compartment). The trans compartment was held at virtual ground potential. Recordings were stored on digital audio tape (Biologic DTR 1200 DAT recorder, Grenoble, France). Records were filtered at 1 kHz (−3 dB point) through a four-pole Bessel low-pass filter and digitized off-line at a rate of 2 kHz on a personal computer.
RESULTS
Purification of PulD.
PulD was purified from an Escherichia coli strain lacking the major porins OmpC and OmpF and the maltose-inducible LamB maltoporin, and carrying a plasmid (pCHAP231) encoding PulD and all other secreton proteins needed for pullulanase secretion in E. coli (22). Isolated outer membranes were solubilized with the zwitterionic detergent SB3–14, which is less likely to cause protein denaturation than the SDS used in some previous studies of secretins (3, 4). PulD incorporated into the outer membrane could not be extracted by nonionic detergents like octypolyoxyethylene and octyl glucoside, and PulD precipitated if any of these detergents were substituted for SB3–14 after solubilization. In addition, PulD precipitated at less than 200 mM NaCl.
The solubilized protein was purified by anion-exchange and size-exclusion chromatography. The final preparation contained two proteins, a multimer that could be dissociated only with phenol (to give a 66-kDa monomer) and that reacted with PulD-specific antibodies, and a ca. 12-kDa band that reacted with antiserum against PulS (Fig. 1 and not shown). The multimer detected by SDS/PAGE did not react with PulS antibodies, indicating that PulS is associated with the purified complex but dissociates in SDS. Thus, the PulD-PulS complex is different from the InvG(secretin)-InvH(pilot) complex from the type III secretion machinery encoded by genes in pathogenicity island 1 of Salmonella typhimurium, which apparently dissociates during purification (2). Treatment of the purified complex with 4 M urea caused partial dissociation of the PulD multimers and parallel release of PulS. Treatment with DTT to reduce the disulfide bond in PulS did not cause its dissociation from the complex.
Figure 1.

Purification of PulD. Pooled PulD-containing fractions at each step in the purification were analyzed by SDS/PAGE (4–20% acrylamide), and then stained with Coomassie brilliant blue. The 68-kDa band in the PulD-containing peak from the Q Sepharose column is not PulD. The positions of multimeric and monomeric PulD and PulS are indicated. The sizes of the molecular mass markers (extreme right and left lanes) are (from top to bottom) 212, 158, 116, 97, 66, 56, 43, 36, 26, 20, 14, and 7 kDa.
The size of the PulD-PulS complex in SB3–14, as determined by size-exclusion chromatography (Superose 6 column), was >1,338 kDa (the size of the marker protein thyroglobin). PulS also was found in a second peak on the anion exchange column, separate from the PulD-PulS complex (not shown), suggesting that it is present in excess. The secreton components PulC, PulE, PulL, and PulG were not present in the purified complex.
Estimates of the molar ratio of PulS/PulD, as determined by scanning gels such as that shown in Fig. 1 and by quantitative immunoblotting, were between 1:2 and 1:1. However, PulS sometimes gives diffuse bands when separated by SDS/PAGE and binds with variable efficiency to nitrocellulose membranes (unpublished observation). Thus, the amount of PulS bound to PulD might be underestimated in some of these experiments. For this reason, the ratio of PulS to PulD also was determined by dissociating the purified complex with phenol, then separating the two monomers according to their size on a Superose 6 column. The molar ratio of PulS/PulD determined in this way was 0.8:1. We conclude that PulD and PulS are present in close to equivalent amounts in the purified complex.
Examination of Secretin Complex by STEM.
Unstained samples of purified PulD-PulS in SB3–14 were examined by STEM (Fig. 2a). More than 90% of the complexes had a molecular mass of 1,011 ± 112 kDa (n = 1,864) (Fig. 2b). The remaining complexes were exactly double this size (not shown). Thus, the PulD-PulS complex has a much lower tendency to form higher-ordered multimers than other secretins (2, 23), possibly because of the presence of PulS.
Figure 2.

STEM analysis of purified PulD complex. (a) Dark-field STEM micrograph of unstained PulD complex. (b) Histogram of masses of unstained PulD complex. (c) STEM micrograph of PulD complex stained with uranyl formate. (d) Averaged angular power spectrum from 256 selected top views, illustrating the presence of four, six, nine, and 12 angular harmonics. (e) Averaged image of PulD showing the 12 radial spokes. (Scale bar represents 20 nm.)
Examination of negatively stained samples of the PulD-PulS complex by STEM revealed ring-like structures (external and internal diameters of approximately 11.6 nm and 7.7 nm, respectively) with peripheral radial spokes (Fig. 2c). The total average diameter of the complex, including the peripheral structures, was ± 25.8 nm. A total of 256 particles were first aligned with respect to an arbitrary reference, averaged, and then realigned with respect to this average. Although the radial spokes were disordered, the correlation indicated a 12-fold rotational symmetry. Therefore, the average was 12-fold symmetrized and used as a reference for the final alignment. The angular power spectrum of all particles calculated over a radial interval of 12.8–25.6 nm (corresponding to the radial spokes) were averaged to demonstrate the presence of a distinct 12-fold symmetry (Fig. 2d). Multivariate statistical classification after this realignment yielded clusters whose angular power spectra also was calculated. Clusters containing a total of 168 particles showed a strong 12-fold angular order. The average calculated from these particles reveals 12 distinct spokes and a massive, rather unstructured ring (Fig. 2e). PulD-PulS complexes adsorbed preferentially with their axes perpendicular to the air-glow discharged carbon film. Consequently, side views were rarely found and therefore could not be averaged.
Cryo-EM of Secretin Complexes.
Cryo-EM of unstained, frozen-hydrated protein showed both end and side views of the complex. End views of the PulD-PulS complex displayed a ring-shaped structure (external and internal diameters approximately 13.5 and 7.5 nm, respectively). In addition, poorly ordered structures were visible, extending radially from the ring (Fig. 3a). These structures were less ordered than in the negatively stained STEM images, and it was impossible to determine their number. As in the STEM images, the spokes extend far from the ring, giving a total diameter of ≈24.5 nm. Only approximately 10% of the particles observed by cryo-EM were in a side-on orientation. Averages of these side views (Fig. 3b) show two strong bands of protein density, about 8 nm apart, which could correspond to two stacked rings. Additional density was present to the sides of the two rings, indicating that a substantial part of the protein extends laterally. Density also was observed above and below the stacked rings, suggesting that the cylinder might extend well beyond the membrane surface.
Figure 3.
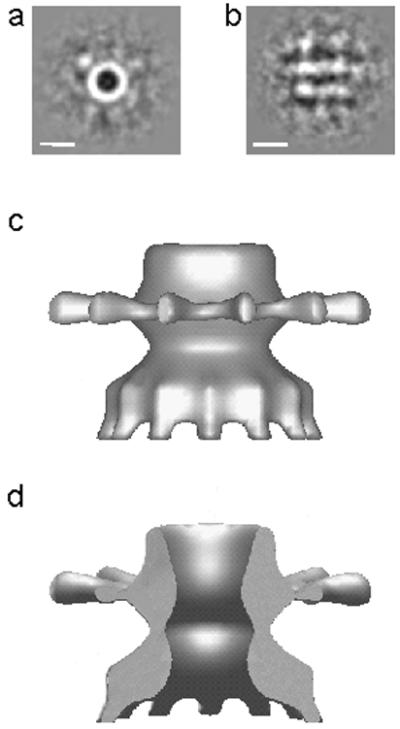
Cryo-EM analysis of unstained preparations of purified PulD-PulS complex. Average end (a) and side (b) views of the PulD-PulS complex. The scale bar in a and b represents 10 nm. A 3D model of the PulD-PulS complex (c) has been sectioned (d), removing the front of the complex so that the channel and profile of the rings can be clearly seen.
The end- and side-view averages (Fig. 3 a and b) were used to make a 3D map based on the 12-fold symmetry observed in the STEM images. This map then was masked to remove noise from around the structure and reprojected, and the reproductions were used as templates for iterative alignment. The resulting cylindrically averaged model (Fig. 3c) shows two rings that correspond to the two bands of density in the side-view average (Fig. 3b). The radial spokes project from one of these two rings, whereas the other ring is radially splayed. A section through the reconstruction (Fig. 3d) shows the central channel and the profile of the rings. The density above and below the plane of the rings in the class average views is not resolved in this preliminary map.
Channel Formation in Planar Lipid Bilayers.
To the best of our knowledge, there are no reports of the formation of channels by secretins and the assumption that they form channels that allow proteins to cross the outer membrane has no experimental foundation other than the fact that the purified protein appears as ring-like structures. The fact that the structure of the purified PulD-PulS complex seemed to be relatively well preserved encouraged us to analyze it for channel activity in planar lipid bilayers. Purified PulD-PulS was reconstituted into proteoliposomes that then were added to a planar lipid bilayer apparatus. Two types of channel activity were detected. The first, observed very infrequently and only with some preparations, displayed the characteristics of porin channels, as described (24). These channels had a high conductance, were maximally open at 0 mV, and closed at positive and negative potential with an asymmetric voltage dependence. The threshold for closure was high (−200 mV) compared with other porins, and we therefore attribute this activity to a contamination of these preparations by a minor, unknown porin.
A second channel activity was consistently observed in all preparations (> 20 independent experiments). Insertion of the channels in the bilayer was revealed at 20 mV as brief, rare openings. However, the application of higher potentials (e.g., 80 mV) progressively increased the size and frequency of the transitions (Fig. 4). After prolonged periods at 80 mV, the channels were irreversibly activated and presented the typical characteristics displayed in Fig. 5. At positive potentials, brief, often aborted, transitions were superimposed on longer openings of various sizes. At low and negative potentials, the activity always presented a more regular pattern with well-defined, correlated transitions over a given period. However, the size of these transitions varied from one bilayer to another, thus precluding further analysis. In asymmetrical media (400 mM KCl in the cis compartment versus 100 mM in the trans compartment), the current reversed around −20 mV, indicating that the channel was weakly selective for cations. This type of electrophysiological activity was never observed in the absence of protein and is unlike that of any other outer membrane protein studied to date. Therefore, we consider it likely that these small, ion-conducting channels are formed by secretin. Pullulanase added to the cis or the trans compartments (12 μg/ml) had no effect on the channel activity (not shown).
Figure 4.

Activation of channel activity at +80 mV. Proteoliposomes reconstituted with PulD-PulS complex were allowed to fuse with a planar lipid bilayer by an osmotic gradient. When channel activity was observed, the membrane potential was clamped at +80 mV, and the channel activity was monitored for 10 min. Solutions in the cis and trans compartments were 10 mM Hepes-KOH, 400 mM KCl, pH 7.4, and 10 mM Hepes-KOH, 100 mM KCl, pH 7.4, respectively.
Figure 5.

Channel activity before and after activation at high membrane potential. Conditions in the cis and trans compartments were as in Fig. 4. (Upper) Trace recordings illustrating the channel activity at different membrane potentials before activation (Left) and after activation at high membrane potential (Right). (Lower) Amplitude histograms (15-sec recordings) at +80 mV and −80 mV, respectively, showing two well-defined transitions at negative potential.
DISCUSSION
The structure of the purified secretin complex composed of PulD and PulS is radically different from that of any previously analyzed secretin. In addition to a ring-shaped structure with a large central cavity, the PulD-PulS complex has radial structures (“spokes”) that extend from the ring. An averaged image of stained specimens examined by STEM indicated that the ring structure has a 12-fold symmetry. This, and a molar ratio for PulD/PulS close to 1:1, leads us to propose that the complex is composed of 12 subunits of PulD that are surrounded by 12 subunits of PulS (see below). The calculated molecular size of such a complex is 956 kDa, slightly less than determined by STEM analysis of unstained preparations (1,011 kDa) and considerably less than determined by gel filtration (>1,338 kDa). However, neither of these size determinations make allowances for the unknown amounts of detergent bound to the complex.
The preliminary 3D reconstruction shown in Fig. 3 is based on the assumption of 12-fold symmetry, as determined by STEM. This map gives only a first tentative view of the secretin structure because it may have been flattened or otherwise distorted by the interaction of the protein with the carbon support film and because unresolved structures above and below the image are not represented. The spokes could be substantially larger than seen in this 3D map because they were also poorly represented in the original images. These spokes are particularly interesting features of the secretin complex because they could correspond to PulS, together with the C-terminal segment of PulD to which it binds (14). Interestingly, radial spokes were not observed when purified preparation of secretins PilQ, XcpQ (3), InvG (2), and YscC (4), all of which were apparently devoid of their cognate pilot proteins (4, 5, 13), were examined by EM. Our previous description of PulS as a chaperone (1, 11) is now obsolete because we have shown that it remains associated to PulD once it is assembled in the outer membrane.
The data presented here suggest that PulD-PulS complexes might form channels in artificial lipid bilayers. The internal diameter of the cavity of the barrel-like structures observed by EM (≈7.6 nm) is sufficient to allow the passage of a molecule with the size of the exoprotein pullulanase. However, the maximum conductance of the channels observed in the planar lipid bilayer experiments was much lower than would be expected for channels of this size and are at variance with the reconstituted image of secretin (Fig. 3d), which shows an open channel. It is worth noting, however, that close inspection of the cryo-EM images reveals the presence of small region of high density in the center of the channel (e.g., Fig. 3a), indicating that it might not, in fact, be open. Moreover, bacteria with open, 7.6-nm diameter channels in the outer membrane would release periplasmic proteins and become more sensitive to normally nonpermeating antibiotics, neither of which occur in bacteria secreting pullulanase (unpublished observations). We consider it likely that the current fluctuations observed in the bilayer experiments correspond to relatively minor, voltage-induced displacements of a small segment of secretin that could form this “plug.”
How, then, is channel gating controlled in vivo? Exoprotein binding or interaction with other components of the secretion machinery might be involved. Such protein-protein contacts might involve the N-terminal half of PulD, which is predicted to face the periplasm. Interestingly, this region of a PulD homologue, the Erwinia chrysanthemi protein OutD, is reported to bind exoproteins secreted by this bacterium (9). Pullulanase had no effect on the channel activity, however. Another possibility is that energy, either from ATP hydrolysis or the proton motive force (25, 26), controls channel opening. This energy coupling might be mimicked by the requirement for a voltage gradient to open ion-conducting secretin channels in vitro, but energy transduction to the channel in vivo probably requires other secreton components. Therefore, future efforts should be directed toward the identification of secreton components that interact with PulD and a more detailed analysis of the roles of the putative periplasmic domain of PulD and of PulS and their contribution to the structures observed by EM.
Acknowledgments
We are grateful to Shirley Müller for her expert help with the STEM mass analysis, Ben de Kruijff for advice regarding the stoichiometry measurements, Odile Possot and Manon Gérard for antibodies against PulC, PulE, and PulL, and all members of the secretion lab at the Institut Pasteur for their interest and encouragement. The work was supported by the European Union (Training and Mobility Research Grant FMRX-CT96–0004) and by a French Research Ministry Programme de Recherche Fondamentale en Microbiology, Maladies Infectieuses et Parasitaries grant.
ABBREVIATIONS
- EM
electron microscopy
- STEM
scanning transmission EM
- 3D
three-dimensional
- SB3–14
3-(N,N-dimethylmyristylammonio)propanesulfonate
References
- 1.Hardie K R, Seydel A, Guilvout I, Pugsley A P. Mol Microbiol. 1996;22:967–976. doi: 10.1046/j.1365-2958.1996.01539.x. [DOI] [PubMed] [Google Scholar]
- 2.Crago A M, Koronakis V. Mol Microbiol. 1998;30:47–56. doi: 10.1046/j.1365-2958.1998.01036.x. [DOI] [PubMed] [Google Scholar]
- 3.Bitter W, Koster M, Latijnhouwers M, de Cock H, Tommassen J. Mol Microbiol. 1998;27:209–219. doi: 10.1046/j.1365-2958.1998.00677.x. [DOI] [PubMed] [Google Scholar]
- 4.Koster M, Bitter W, de Cock H, Allaoui A, Cornelis G, Tommassen J. Mol Microbiol. 1997;26:789–797. doi: 10.1046/j.1365-2958.1997.6141981.x. [DOI] [PubMed] [Google Scholar]
- 5.Drake S L, Sandstedt S A, Koomey M. Mol Microbiol. 1997;23:657–668. doi: 10.1046/j.1365-2958.1997.2511618.x. [DOI] [PubMed] [Google Scholar]
- 6.Kazmierczak B I, Mielke D L, Russel M, Model P. J Mol Biol. 1994;238:187–198. doi: 10.1006/jmbi.1994.1280. [DOI] [PubMed] [Google Scholar]
- 7.Linderoth N A, Simon M N, Russel M. Science. 1997;278:1635–1638. doi: 10.1126/science.278.5343.1635. [DOI] [PubMed] [Google Scholar]
- 8.Cowan S W, Schrimer T, Rummer G, Steiert M, Ghosh R, Pauptit R A, Jansonius J N, Rosenbush J P. Nature (London) 1992;358:727–733. doi: 10.1038/358727a0. [DOI] [PubMed] [Google Scholar]
- 9.Shevchik V E, Robert-Badouy J, Condemine G. EMBO J. 1997;16:3007–3016. doi: 10.1093/emboj/16.11.3007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Daefler S, Russel M, Model P. J Mol Biol. 1997;266:978–992. doi: 10.1006/jmbi.1996.0866. [DOI] [PubMed] [Google Scholar]
- 11.Hardie K R, Lory S, Pugsley A P. EMBO J. 1996;15:978–988. [PMC free article] [PubMed] [Google Scholar]
- 12.Shevchik V E, Condemine G. Microbiology. 1998;144:3219–3228. doi: 10.1099/00221287-144-11-3219. [DOI] [PubMed] [Google Scholar]
- 13.Daefler S, Russel M. Mol Microbiol. 1998;28:1367–1380. doi: 10.1046/j.1365-2958.1998.00908.x. [DOI] [PubMed] [Google Scholar]
- 14.Daefler S, Guilvout I, Hardie K R, Pugsley A P, Russel M. Mol Microbiol. 1997;24:465–475. doi: 10.1046/j.1365-2958.1997.3531727.x. [DOI] [PubMed] [Google Scholar]
- 15.Miller J H. Experiments in Molecular Genetics. Plainview, NY: Cold Spring Harbor Lab. Press; 1972. [Google Scholar]
- 16.Müller S A, Goldie K N, Bürki R, Häring R, Engel A. Ultramicroscopy. 1992;46:317–334. [Google Scholar]
- 17.Saxton W O, Pitt T J, Horner M. Ultramicroscopy. 1979;4:343–354. [Google Scholar]
- 18.Frank J, Bretaudiere J-P, Carazo J-M, Verschoor A, Wagenknecht T. J Microsc. 1987;150:99–115. doi: 10.1111/j.1365-2818.1988.tb04602.x. [DOI] [PubMed] [Google Scholar]
- 19.Chen S, Roseman A M, Hunter A S, Wood S P, Burston S G, Ranson N A, Clarke A R, Saibil H R. Nature (London) 1994;371:261–264. doi: 10.1038/371261a0. [DOI] [PubMed] [Google Scholar]
- 20.van Heel M, Harauz G, Orlova E V. J Struct Biol. 1996;116:17–24. doi: 10.1006/jsbi.1996.0004. [DOI] [PubMed] [Google Scholar]
- 21.Frank J, Radermacher M, Penczek P, Zhu J, Li Y, Ladjadj M, Leith A. J Struct Biol. 1996;116:190–199. doi: 10.1006/jsbi.1996.0030. [DOI] [PubMed] [Google Scholar]
- 22.d’Enfert C, Ryter A, Pugsley A P. EMBO J. 1987;6:3531–3538. doi: 10.1002/j.1460-2075.1987.tb02679.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Linderoth N A, Simon M N, Russel M. Science. 1997;278:1635–1638. doi: 10.1126/science.278.5343.1635. [DOI] [PubMed] [Google Scholar]
- 24.Berrier C, Besnard M, Ghazi A. J Membr Biol. 1997;156:105–115. doi: 10.1007/s002329900193. [DOI] [PubMed] [Google Scholar]
- 25.Possot O, Letellier L, Pugsley A P. Mol Microbiol. 1997;24:457–464. doi: 10.1046/j.1365-2958.1997.3451726.x. [DOI] [PubMed] [Google Scholar]
- 26.Letellier L, Howard S P, Buckley T J. J Biol Chem. 1997;272:11109–11113. doi: 10.1074/jbc.272.17.11109. [DOI] [PubMed] [Google Scholar]