

The crystallization and data collection of topoisomerase IV from S. aureus is described. Phasing by molecular replacement proved difficult owing to the presence of translational NCS and strategies used to overcome this are discussed.
Keywords: topoisomerase IV, DNA cleavage, quinolone binding, translational NCS
Abstract
DNA topoisomerase IV removes undesirable topological features from DNA molecules in order to help maintain chromosome stability. Two constructs of 56 and 59 kDa spanning the DNA-cleavage domain of the A subunit of topoisomerase IV from Staphylococcus aureus (termed GrlA56 and GrlA59) have been crystallized. Crystals were grown at 291 K using the sitting-drop vapour-diffusion technique with PEG 3350 as a precipitant. Preliminary X-ray analysis revealed that GrlA56 crystals belong to space group P21, diffract to a resolution of 2.9 Å and possess unit-cell parameters a = 83.6, b = 171.5, c = 87.8 Å, β = 90.1°, while crystals of GrlA59 belong to space group P21212, with unit-cell parameters a = 41.5, b = 171.89, c = 87.9 Å. These crystals diffract to a resolution of 2.8 Å. This is the first report of the crystallization and preliminary X-ray analysis of the DNA-cleavage domain of a topoisomerase IV from a Gram-positive organism.
1. Introduction
DNA is prone to topological problems owing to its double-helical structure and the intertwining of its complementary strands. For example, during replication the unwinding of the chromosome forces adjacent regions of DNA to overwind and underwind, generating a superhelical strain (Wu et al., 1988 ▶). The removal of this intermolecular strain is vital for the prevention of DNA tangling and knotting, which produces structures that can lead to the formation of double-stranded breaks in the DNA and chromosome instability (Kato et al., 1990 ▶); furthermore, it is essential that circular daughter chromosomes of bacteria are disentangled at cell division. The resolution of such structures is performed by topoisomerase IV (topoIV), a type IIA DNA topoisomerase, which can remove these regions of unusual topology by passing one double-helical segment of DNA through another.
All bacterial type IIA topoisomerases are A 2 B 2 heterotetramers containing three discrete subunit interfaces which open and close sequentially in response to ATP binding and hydrolysis in order to effect DNA transport (Wang, 1998 ▶). One segment of DNA (termed the gate segment) is initially bound by the A subunit, followed by the capture of a second strand (termed the transfer segment) by the B subunits as they bind ATP and dimerize. The dimerization and ensuing interaction with the A subunits triggers the cleavage of the gate segment. Subsequent hydrolysis of ATP causes the transfer segment to be passed through the DNA gate. DNA passage is followed by resealing of the gate segment and release of the transfer segment from the enzyme (Corbett & Berger, 2004 ▶).
Topoisomerases perform such a vital role in the maintenance of genetic stability within the cell that they are ideal drug targets and are the site of action of numerous antibacterial agents (Hooper, 2001 ▶). One class of inhibitor includes the quinolone antibiotics, which have proven to be highly effective against a variety of bacterial species (Oliphant & Green, 2002 ▶). These molecules bind to the A subunit and stabilize the topoisomerase–DNA complex formed during the catalytic turnover of the enzyme. The stalled topoisomerase complex then acts as a physical barrier to DNA replication. Collision of an active replication complex with the topoisomerase is thought to lead to the double-stranded breaks necessary for cell lethality (Drlica & Zhao, 1997 ▶), although the exact mechanism by which this occurs is unknown.
Topoisomerase IV has been shown to be the primary target for quinolones in Gram-positive species (Oliphant & Green, 2002 ▶), but currently structural information is only available for the DNA-cleavage domains of topoisomerases from eukaryotic (PDB code 1bgw; Berger et al., 1996 ▶) and Gram-negative bacterial species (PDB codes 1ab4 and 1zvu; Morais-Cabral et al., 1997 ▶; Corbett et al., 2005 ▶). Here, we describe the crystallization and X-ray analysis of two different constructs of the A subunit of topoisomerase IV from Staphylococcus aureus (GrlA) with a view to gaining further insight into the mode of inhibition of quinolone antibiotics against this enzyme.
2. Materials and methods
2.1. Purification
Full details of the expression and purification are given elsewhere (Carr et al., 2006 ▶). Briefly, the two constructs encompassing the N-terminal domains of S. aureus GrlA (accession code D67075) were generated by PCR from genomic DNA into the expression vector pET15b, such that 23 amino acids derived from the vector (including a hexahistidine tag) are fused to codons 25–491 or 1–491 of GrlA. This leads to the generation of proteins with molecular weights of approximately 56 kDa (GrlA56) and approximately 59 kDa (GrlA59), respectively. Protein expression was performed in Escherichia coli B834 DE3 (plysS) cultures by incubation at 310 K for 3 h after the addition of IPTG to a final concentration of 0.5 mM. Cells were lysed by sonication in 50 mM Tris–HCl pH 7.5, 400 mM KCl, 10%(v/v) ethanediol and debris was removed by centrifugation (12 000g for 30 min). The resulting supernatant was loaded onto a column of HP chelating Sepharose (GE Healthcare), previously charged with nickel, and eluted with a 0–500 mM imidazole gradient. Protein-containing fractions were pooled and diluted with 50 mM Tris–HCl pH 7.5, 10%(v/v) ethanediol to a final KCl concentration of 75 mM. This material was then loaded onto a column of heparin Sepharose (GE Healthcare) pre-equilibrated with 50 mM Tris–HCl pH 7.5, 75 mM KCl, 10%(v/v) ethanediol. Protein was then eluted with a 0–1 M KCl gradient. Samples containing GrlA56 or GrlA59 were identified by absorbance at 280 nm and SDS–PAGE analysis. The purified proteins were extensively dialysed against 50 mM Tris–HCl pH 7.5, 400 mM KCl, 10%(v/v) ethanediol and concentrated to 5 mg ml−1 in an ultrafiltration cell (Millipore, 10 kDa cutoff).
2.2. Crystallization
Initial crystallization conditions were obtained by sparse-matrix screening (Jancarik & Kim, 1991 ▶) using the sitting-drop vapour-diffusion technique. Drops were prepared by mixing 0.5 µl protein solution with 0.5 µl screen solution using an Oryx6 liquid-handling robot (Douglas Instruments) and were equilibrated against a 100 µl reservoir of screen solution at 291 K. Initial crystals of both GrlA56 and GrlA59 appeared after 5 d from Index Screen solution No. 73 [100 mM Tris pH 8.5, 200 mM NaCl and 25%(w/v) PEG 3350; Hampton Research]. Streak-seeding into drops (2 µl protein and 2 µl 100 mM Tris pH 8.3–8.5, 200 mM NaCl and 12–16% PEG 3350) pre-equilibrated for 1–2 h against a 0.5 ml reservoir produced large diffraction-quality crystals for both constructs.
2.3. Data collection
Crystals were transferred into 100 mM Tris pH 8.5, 200 mM NaCl, 20%(w/v) PEG 3350 and 25% ethanediol prior to flash-cooling by immersion in liquid nitrogen. Data collection was performed at 100 K on station ID23.2, ESRF, France with a MAR CCD image-plate detector (MAR Research). Data were collected in multiple batches spanning 30° (0.5° oscillations), each separated by a translation of 50 µm along the long axis of the crystal, in order to minimize radiation damage. Diffraction data were integrated using MOSFLM (Leslie, 1992 ▶), followed by scaling and merging using SCALA (Evans, 1997 ▶). Further data analysis was performed using various programs from the CCP4 program suite (Collaborative Computational Project, Number 4, 1994 ▶).
3. Results and discussion
Crystals of both protein fragments displayed a plate-like morphology and initially grew in tightly packed clusters. Streak-seeding proved the most effective method of producing large single crystals suitable for diffraction analysis (Figs. 1 ▶ a and 1 ▶ b). The diffraction images collected from crystals of GrlA56 were unusual in that they contained strong reflections when the h index was even and weak reflections when h was odd (Fig. 2 ▶).
Figure 1.

(a) A monoclinic crystal of GrlA56; typical dimensions are 400 × 300 × 20 µm. (b) An orthorhombic crystal of GrlA59 mounted in a cryoloop prior to data collection; typical dimensions are 500 × 150 × 20 µm.
Figure 2.

Diffraction pattern observed for GrlA56 crystals, with the differences in intensity between weak h odd and strong h even reflections highlighted.
The GrlA56 crystals belong to space group P21 (Table 1 ▶) and could contain between two and six copies of the construct per asymmetric unit, with four copies corresponding to a solvent content of 56% (Matthews, 1968 ▶). Calculation of a self-rotation function (Fig. 3 ▶) reveals the presence of orthorhombic pseudosymmetry, so the data were rescaled applying the pseudosymmetry (P21212) indicated by inspection of systematic absences. Preliminary attempts at molecular replacement into the pseudo-cell using AMoRe (Navaza, 1994 ▶) were performed using the complete model of the 59 kDa fragment of DNA gyrase from E. coli (PDB code 1ab4; Morais Cabral et al., 1997 ▶). This revealed a dimer within the asymmetric unit with an intermolecular twofold axis coincident with the crystallographic twofold. A slight distortion of the molecules that shifted the molecular twofold away from the crystallographic twofold would result in a lower symmetry monoclinic space group.
Table 1. GrlA56 and GrlA59 X-ray diffraction data-collection statistics.
Values in parentheses are for the highest resolution bin.
| GrlA59 | GrlA56 | |
|---|---|---|
| Space group | P21212 | P21 |
| Unit-cell parameters (Å, °) | a = 41.5, b = 171.89, c = 87.9 | a = 83.6, b = 171.5, c = 87.8, β = 90.1 |
| Wavelength (Å) | 0.873 | 0.873 |
| Resolution limits (Å) | 88–2.8 (2.95–2.8) | 88–2.9 (3.06–2.9) |
| Matthews coefficient (Å3 Da−1) | 2.80 | 2.81 |
| Molecules per ASU | 1 | 4 |
| Unique reflections | 16113 (2308) | 54736 (8090) |
| Completeness (%) | 99.3 (99.3) | 99.0 (99.0) |
| Multiplicity | 3.6 (3.5) | 3.7 (3.7) |
| 〈I/σ(I)〉 | 13.5 (3.1) | 12.7 (2.7) |
| Rmerge† (%) | 8.4 (42.5) | 8.7 (46.5) |
R
merge = 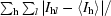
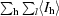 , where I
l is the lth observation of reflection h and 〈I
h〉 is the weighted average intensity for all observations l of reflection h.
, where I
l is the lth observation of reflection h and 〈I
h〉 is the weighted average intensity for all observations l of reflection h.
Figure 3.

Self-rotation function (κ = 180°) calculated for GrlA56 reveals orthorhombic pseudosymmetry within the monoclinic crystal lattice.
Applying a solvent content of 56% to the monoclinic cell suggests the presence of four molecules per asymmetric unit. Knowledge of the packing within the crystal suggests they should be arranged as a pair of dimers related by a direct translation of a/2 and each possessing a noncrystallographic twofold axis almost parallel to c. This packing also accounts for the presence of weak h odd reflections in the diffraction pattern, as the similar orientation of each pair of dimers results in the strong reflections corresponding to a unit cell with a cell edge of a/2 interspersed with weak intensities generated solely by the small differences between the two independent dimers. Further refinement of this model was severely impeded by the translational NCS relating each dimer within the asymmetric unit and this led to the redesign and production of the GrlA59 construct.
Crystals of GrlA59 belong to space group P21212 (Table 1 ▶), contain one molecule per asymmetric unit and have a solvent content of 56%. The unit cell is related to that observed for GrlA56 crystals, but slight differences in packing have increased the order within the crystals, leading to the smaller unit cell and higher symmetry. It is possible that the additional residues at the N-terminus have created extra crystal contacts. It is likely that the orientation of the monomer within the asymmetric unit is such that the intermolecular twofold axis is parallel to c. Molecular replacement performed as previously described for GrlA56 confirm this assertion, generating a model suitable for rebuilding and refinement.
Acknowledgments
We would like to thank Val Sergeant for technical support and David Flot (ID23.2, ESRF) for assistance during data collection. We would also like to thank the Wellcome Trust for funding.
References
- Berger, J. M., Gamblin, S. J., Harrison, S. C. & Wang, J. C. (1996). Nature (London), 379, 225–232. [DOI] [PubMed] [Google Scholar]
- Carr, S. B., Makris, G., Cove, J. H., Phillips, S. E. V. & Thomas, C. D. (2006). In preparation.
- Collaborative Computational Project, Number 4 (1994). Acta Cryst. D50, 760–763. [Google Scholar]
- Corbett, K. D. & Berger, J. M. (2004). Annu. Rev. Biophys. Biomol. Struct.33, 95–118. [DOI] [PubMed] [Google Scholar]
- Corbett, K. D., Schoeffler, A. J., Thomsen, N. D. & Berger, J. M. (2005). J. Mol. Biol.351, 545–561. [DOI] [PubMed] [Google Scholar]
- Drlica, K. & Zhao, X. L. (1997). Microbiol. Mol. Biol. Rev.61, 377–392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans, P. R. (1997). Jnt CCP4/ESF–EACBM Newsl. Protein Crystallogr.33, 22–24.
- Hooper, D. C. (2001). Clin. Infect. Dis.32, Suppl. 1, S9–S15. [DOI] [PubMed] [Google Scholar]
- Jancarik, J. & Kim, S.-H (1991). J. Appl. Cryst.24, 409–411. [Google Scholar]
- Kato, J., Nishimura, Y., Imamura, R., Niki, H., Hirago, S. & Suzuki, H. (1990). Cell, 63, 393–404. [DOI] [PubMed] [Google Scholar]
- Leslie, A. G. W. (1992). Jnt CCP4/ESF–EACBM Newsl. Protein Crystallogr.26
- Matthews, B. W. (1968). J. Mol. Biol.33, 491–497. [DOI] [PubMed] [Google Scholar]
- Morais Cabral, J. H., Jackson, A. P., Smith, C. V., Shikotra, N., Maxwell, A. & Liddington, R. C. (1997). Nature (London), 388, 903–906. [DOI] [PubMed] [Google Scholar]
- Navaza, J. (1994). Acta Cryst. A50, 157–163. [Google Scholar]
- Oliphant, C. M. & Green, G. M. (2002). Am. Fam. Physician, 65, 455–464. [PubMed] [Google Scholar]
- Wang, J. C. (1998). Q. Rev. Biophys.31, 107–144. [DOI] [PubMed] [Google Scholar]
- Wu, H. Y., Shyy, S., Wang, J. C. & Liu, L. F. (1988). Cell, 53, 433–440. [DOI] [PubMed] [Google Scholar]