

Abstract
To investigate modulation of the activation of cGMP-gated ion channels in cone photoreceptors, we measured currents in membrane patches detached from the outer segments of single cones isolated from striped bass retina. The sensitivity of these channels to activation by cGMP depends on the history of exposure to divalent cations of the membrane's cytoplasmic surface. In patches maintained in 20 μM Ca++ and 100 μM Mg++ after excision, the current amplitude dependence on cGMP is well described by a Hill equation with average values of K 1/2, the concentration necessary to activate half the maximal current, of 86 μM and a cooperativity index, n, of 2.57. Exposing the patch to a solution free of divalent cations irreversibly increases the cGMP sensitivity; the average value of K 1/2 shifts to 58.8 μM and n shifts to 1.8. Changes in cGMP sensitivity do not affect other functional parameters of the ion channels, such as the interaction and permeation of mono- and divalent cations. Modulation of cGMP activation depends on the action of an endogenous factor that progressively dissociates from the channel as Ca++ concentration is lowered below 1 μM. The activity of the endogenous modulator is not well mimicked by exogenously added calmodulin, although this protein competes with the endogenous modulator for a common binding site. Thus, the modulation of cGMP affinity in cones depends on the activity of an unidentified molecule that may not be calmodulin.
Keywords: retina, phototransduction, calmodulin, rod photoreceptor, fish
introduction
Rod and cone photoreceptors and olfactory sensory neurons respond to their appropriate stimuli with changes in membrane conductance that reflect the activity of cyclic nucleotide–gated ion channels (Fesenko et al., 1985; Nakamura and Gold, 1987; Haynes and Yau, 1990). In these cells, the stimuli activate enzymatic cascades that ultimately change the cytoplasmic concentration of either cGMP (photoreceptors) (reviewed by Hurley, 1992; Pugh and Lamb, 1993; Baylor, 1996) or cAMP (olfactory neurons) (reviewed by Reed, 1992). Because of similarities in primary structure, the cyclic nucleotide– gated channels in the sensory cells have been recognized to be members of the same gene family (for review see Zagotta and Siegelbaum, 1996; Finn et al., 1996; Yau and Chen, 1995). Despite their general structural similarities, the chemical specificity and affinity of the ligand binding sites differ among channels of the various sensory cell types (Scott et al., 1996). For example, the channels in retinal photoreceptors have a 20- to 50-fold higher affinity for cGMP than cAMP, whereas native channels of olfactory neurons have nearly the same affinity for both nucleotides (see review in Zagotta and Siegelbaum, 1996). Even in the same receptor cell type, the nucleotide affinity of the channels varies from species to species (reviewed in Zagotta and Siegelbaum, 1996), presumably optimized to match the nucleotide cytoplasmic concentration in the cell expressing that specific channel. Furthermore, recent experiments have shown that the ligand affinity of the channels is not static, but changes as a function of Ca++ concentration (reviewed in Molday, 1996). The physiological significance of this Ca++ dependence is not fully understood. However, it is likely to play an important role in the adaptation of the cell's response to changing background levels of stimulation (Kurahashi and Menini, 1997).
The extent to which Ca++ modulates the cGMP activation differs markedly between olfactory and rod photoreceptor channels. In rat olfactory neurons, for example, affinity for cAMP decreases over 100-fold when Ca++ is elevated to 200 μM (Chen and Yau, 1994; Liu et al., 1994; Balasubramanian et al., 1996). Affinity is measured by the value of K 1/2, the concentration necessary to activate half the maximum ligand-gated current. In rod photoreceptors, the Ca++ modulation of K 1/2 has been studied in isolated membrane vesicles (Bauer, 1996), detached membrane patches (Gordon et al., 1995), and truncated outer segments (Nakatani et al., 1995; Sagoo and Lagnado, 1996). In all these preparations, the K 1/2 for cGMP activation shifts ∼1.5-fold with a Ca++ dependence that is half maximal at ∼50 nM. Because this modulation is irreversibly lost in patches and truncated rods after exposure to solutions free of divalent cations, it has been presumed to arise from the action of a soluble, endogenous modulator (Gordon et al., 1995; Nakatani et al., 1995; Sagoo and Lagnado, 1996; Bauer, 1996). The molecular identity of this endogenous modulator in rods is not fully resolved, but the possibility that it is calmodulin has been addressed in a number of recent experiments.
Hsu and Molday (1993) first demonstrated that calmodulin can cause a Ca++-dependent modulation of K 1/2 in rods. In a manner similar to the endogenous modulator, the maximum Ca++/calmodulin-dependent shift in K 1/2 is ∼1.5-fold (Gordon et al., 1995; Kosolapov and Bobkov, 1996; Bauer, 1996). The Ca++ dependence of the shift in K 1/2 in the presence of added calmodulin has a half-maximum value of ∼50 nM in some studies (Hsu and Molday, 1993; Bauer, 1996) and 450 nM in others (Kosolapov and Bobkov, 1996). These values, however, cannot be compared with those characteristic of the endogenous modulator because they change with the calmodulin concentration used in the experiments (Bauer, 1996). Gordon et al. (1995) and Bauer (1996) have directly investigated whether the endogenous factor and calmodulin are one and the same. In membrane vesicles, there are no recognizable differences in the function of the two molecules (Bauer, 1996). However, in membrane patches, the function of both molecules was found to differ in several respects, suggesting that the modulator in rods may differ from calmodulin (Gordon et al., 1995). In truncated rods, pharmacological blockers of calmodulin do not alter the Ca++-dependent modulation of K 1/2, nor does added calmodulin confer modulation (Sagoo and Lagnado, 1996). Finally, Haynes and Stotz (1997) have found that added calmodulin can modulate K 1/2 in rod membrane patches, but not in those of cones from the same species.
The cyclic nucleotide–gated channels of cone outer segments are structurally homologous to those of rods (Bonigk et al., 1993; Weyand et al., 1994), yet their functional properties differ in subtle but important ways. These functional differences may contribute to explain the differences in transduction between the two receptor types (Korenbrot, 1995). The K 1/2 of cGMP binding is higher in cones (Haynes and Yau, 1990; Picones and Korenbrot, 1992) than in rods (Karpen et al., 1988; Zimmerman and Baylor, 1992; Haynes and Stotz, 1997); the energy of interaction of cations such as Na+ and Li+ with the channel is higher in cones (Picones and Korenbrot, 1992; Haynes, 1995) than in rods (Menini, 1990; Furman and Tanaka, 1990; Zimmerman and Baylor, 1992), and the blocking effect of l-cis-diltiazem is also different (Haynes, 1992). Of singular functional significance is the fact that the permeability and interaction of Ca++ with the channels differs between the two photoreceptor types (Korenbrot, 1995). In particular, the permeability of Ca++ relative to Na+ is higher in cone than in rod channels (Picones and Korenbrot, 1995). This difference is also observed in recombinant channels formed by cone or rod α subunits alone (Frings et al., 1995). Whether there exists a calcium-dependent modulation of ligand binding in cone channels and what features this modulation might have has not been previously investigated. We report here that in membrane patches of cone outer segments, as in those of rods, there exists a Ca++-dependent modulation of cGMP activation. This modulation depends on the activity of an endogenous modulator. We further contrast the properties of rod and cone channels with respect to the modulatory effect of added calmodulin. In cones, the endogenous modulator is not well mimicked by calmodulin.
materials and methods
Materials
We obtained striped bass (Morone saxitilis) from Professional Aquaculture Services (Chico, CA) and tiger salamanders (Ambystoma tigrinum) from Charles Sullivan (Memphis, TN). We received cGMP, 8-Br-cGMP, and calmodulin (bovine brain phosphodiesterase activator) from Sigma Chemical Co. (St. Louis, MO).
Photoreceptor Isolation
Methods of cell isolation are described in detail elsewhere (Miller and Korenbrot, 1993b , 1994). Animals were dark adapted and retinas were isolated under infrared illumination. Single cones were obtained by mechanical dissociation of striped bass retinas maintained in a Ringer's solution consisting of (mM): 143 NaCl, 2.5 KCl, 5 NaHCO3, 1 Na2HPO4, 1 CaCl2, 1 MgCl2, 5 pyruvate, 10 HEPES, pH 7.5, osmotic pressure 309 mOsM. Rods were isolated by mechanical dissociation of tiger salamander retinas maintained in a Ringer's solution composed of (mM): 100 NaCl, 2 KCl, 5 NaHCO3, 1 Na2HPO4, 1 CaCl2, 1 MgCl2, 5 pyruvate, 10 HEPES, pH 7.4, osmotic pressure 227 mOsM.
Solitary photoreceptors were firmly attached to a glass coverslip derivatized with wheat germ agglutinin (Picones and Korenbrot, 1992). The coverslip formed the bottom of a recording chamber held on the stage of an upright microscope equipped with DIC optics and operated under visible light. A suspension of photoreceptors in pyruvate-Ringer's was placed on the coverslip and the cells were allowed to settle down and attach for 5 min. The bath solution was then exchanged with a Ringer's solution of the same composition, but in which pyruvate was isosmotically replaced with glucose.
The recording chamber consisted of two side-by-side compartments. Cells were held in one compartment that was continuously perfused with Ringer's. The second, smaller compartment was continuous with the first one, but a movable barrier could be used to separate them (Picones and Korenbrot, 1992). We used tight-seal electrodes to obtain inside-out membrane fragments detached from the side of the outer segments of either cones or rods. After forming a giga-seal and detaching the membrane fragment, we moved the electrode under the solution surface from the compartment containing the intact cells to the smaller compartment. The two compartments were then separated by the movable barrier and the tip of the electrode was placed within 100 μm of the opening of a 300-μm diameter glass capillary. We used this capillary and a rotary valve to deliver selected test solutions onto the cytoplasmic (outside) surface of the membrane patch.
The speed of change in membrane current in response to changes in test solutions varied from patch to patch. This variability likely reflects differences among patches in the accessibility of their cytoplasmic surface to the bath solution. In our experiments, after each solution change, we monitored membrane current by repeated presentation of voltage steps and waited for the current to reach a stationary amplitude. We present and analyze here only currents in this stationary condition (except for Fig. 5).
Figure 5.

Ca++ dependence of current amplitude in the presence of the endogenous modulator in a cone outer segment membrane patch. Currents, activated by changing membrane voltage from 0 to −40 mV, were repeatedly measured in the same patch in the presence of 60 μM cGMP, 20 μM Ca++, and 100 μM Mg++ after 60-s exposure to solutions containing progressively lower Ca++ concentrations. Data points were normalized by dividing current amplitude measured after each Ca++ concentration tested (I) by the amplitude measured after exposure to a Ca++ free solution (EDTA/EGTA solution) (I0). The data point on the far left indicates I0. This is the maximum possible current since repeated washes in EDTA/EGTA solution did not result in any additional current enhancement.
Ionic Solutions
In studies of both cone and rod membranes, we filled the tight-seal electrodes with the same solution (mM): 157 NaCl, 5 EGTA, 5 EDTA, 10 Hepes, adjusted with NaOH to pH 7.5, osmotic pressure 305 mOsM. Free Ca++ concentration in this solution was ⩽10−10 M and total Na+ concentration was 167 mM. In all studies, the initial composition of the solution in the small compartment into which we moved the electrode was 157 mM NaCl, 10 mM Hepes, adjusted with NaOH to pH 7.5, and 20 μM free Ca++ (total 7.637), 100 μM free Mg++ (total 2) with 10 mM HEDTA. We elected to use these free divalent cation concentrations because they are sufficiently low to not block the cGMP-gated conductance (Picones and Korenbrot, 1995), yet sufficiently high to saturate the Ca++ dependence of the phenomena studied here.
We used a titration method to produce solutions with free calcium concentrations between 10 nM and 20 μM (Williams and Fay, 1990). This method yields accurate free calcium concentrations without the need to weigh with extreme precision chemicals of known purity. This is particularly important because 100% pure Ca++ buffering agents cannot be obtained commercially (Miller and Smith, 1984). Briefly, a solution containing (mM) 157 NaCl, 10 HEPES, 10 HEDTA, adjusted with NaOH to pH 7.5, was divided into two parts. One part (solution A) had no added calcium. Into the other part (solution B), CaCl2 was added to 9 mM and the pH readjusted to 7.8. A small sample of this solution was then titrated with 100 mM CaCl2 while monitoring its pH. The amount of CaCl2 necessary to fully titrate HEDTA, that is, to obtain pure CaHEDTA at pH 7.5, was determined from this titration. The appropriate amount of CaCl2 was then added to the remaining solution B. Solutions A and B were mixed to obtain different free calcium concentrations (Table I). Volumes were calculated (EqCal; BioSoft, Cambridge, UK) using published values for HEDTA binding constants (Martell and Smith, 1974). The pH was adjusted to 7.5 and solutions stored at 4°C in plastic containers for up to 1 wk. Solutions containing both calcium and magnesium (at concentrations of 20 and 100 μM free, respectively) were made by adding to solution A appropriate amounts of stocks of CaCl2 and MgCl2. The concentration of divalent cations in these stocks were calibrated by measuring their osmolality.
Table I.
Solutions of Defined Free Ca++ Concentration Using HEDTA
| Free calcium | Solution A | Solution B | ||
|---|---|---|---|---|
| 20 μM | 0.0586 | 0.9414 | ||
| 5 μM | 0.1993 | 0.8007 | ||
| 1 μM | 0.5545 | 0.4455 | ||
| 500 nM | 0.7134 | 0.2866 | ||
| 300 nM | 0.8059 | 0.1942 | ||
| 100 nM | 0.9256 | 0.0744 | ||
| 50 nM | 0.9614 | 0.0386 | ||
| 30 nM | 0.9765 | 0.0235 | ||
| 10 nM | 0.9920 | 0.0080 |
In many of the experiments reported here, we tested the electrical properties of the same membrane patch before and after removing divalent cations. Divalent cations were removed by exposing the cytoplasmic (outside) surface of the patch to a solution composed of (mM): 157 NaCl, 10 HEPES, 5 EDTA, 5 EGTA, adjusted with NaOH to pH 7.5. We refer to this solution in the following text simply as the EDTA/EGTA solution.
Electrical Recordings
Tight-seal electrodes were made from aluminosilicate glass (1724; Corning Glass Works, Corning, NY) (1.5 × 1.0 mm, o.d. × i.d.). We measured membrane currents under voltage clamp at room temperature (19–21°C) with a patch-clamp amplifier (#8900; Dagan Instruments, Minneapolis, MN). Analog signals were low pass filtered below 2.5 kHz with an eight pole Bessel filter and were digitized on line at 6 kHz (FastLab; Indec, Capitola, CA). Membrane voltage was normally held at 0 mV and membrane currents were activated either by 110-ms long step changes to −40 mV or with a continuous voltage ramp that swept between −70 and +70 mV at a rate of 228 mV/s. In measurements with voltage ramps, the voltage was stepped from 0 to −70 mV and held at that value for 200 ms before applying the voltage ramp. This was necessary because in the presence of divalent cations there is a time-dependent change in membrane current amplitude on switching from 0 to −70 mV. This time-dependent change is due to the well characterized voltage-dependent channel block by divalent cations (in rods, Zimmerman and Baylor, 1992; in cones, Picones and Korenbrot, 1995). To generate current–voltage (I-V)1 curves, the voltage ramp was swept in four successive trials and the currents were signal averaged. Between voltage ramp trials, membrane voltage was held at 0 mV for 1.2 s. As usual, outward currents are positive and the extracellular membrane surface is defined as ground.
We began every experiment by determining the current amplitude before and after adding saturating cGMP concentrations. We frequently found (>50%) that membrane patches did not respond initially to the ligand. In many instances, these patches became responsive after rapidly moving the electrode tip across the air–water interface. It is likely, therefore, that unresponsive patches had formed closed vesicles that opened upon crossing the air–water interface (Horn and Patlak, 1980). We only analyzed data measured in patches in which the amplitude of the current generated with saturating cGMP concentrations and the leakage current measured in the absence of cGMP did not change by >10% over the course of the entire experiment. Functions were fit to experimental data by least square minimization algorithms (Origin; MicroCal Software, Northampton, MA). Experimental errors are presented as standard deviations.
results
The K1/2 of the cGMP-gated Currents in Cones Is Modulated by Divalent Cations
We investigated cGMP-dependent currents in inside-out patches detached from the plasma membrane of retinal cone outer segments. These patches contain only cGMP-gated ion channels (Miller and Korenbrot, 1993a ). The dependence of current amplitude on cGMP concentration is well described by the Hill equation (Fig. 1):
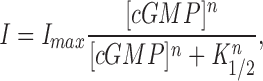 |
1 |
Figure 1.

Modulation of cGMP-dependent currents in detached membrane patches. A and B illustrate currents measured at room temperature in a patch detached from a cone outer segment. Currents were activated with voltage steps from 0 to −40 mV in the presence of 20 μM Ca++ and 100 μM Mg++ and various cGMP concentrations (shown are currents at 0, 40, 60 [arrow], 100 μM, and 1 mM). Currents shown in A were measured shortly after patch excision, while those in B were measured in the same membrane after exposure to the EDTA/EGTA solution. The lower panels illustrate the dependence of normalized current amplitude on cGMP concentration at −40 mV before and after exposure to the EDTA/EGTA solution. Data points were normalized by dividing the amplitude at each cGMP concentration tested (I) by the maximum current (Imax). C illustrates data measured in a patch detached from a cone outer segment. The continuous curve is the best fit to the data of the Hill equation (Eq. 1) with K 1/2 = 88.4 μM and n = 2.73 before exposure to EDTA/EGTA, and K 1/2 = 61.4 μM and n = 1.59 afterwards. D illustrates data measured in rod membrane patches, where the continuous curve is the Hill equation with K 1/2 = 34.2 μM and n = 2.3 before exposure to the EDTA/EGTA solution, and K 1/2 = 17.9 μM and n = 1.6 afterwards.
where I is the amplitude of the cGMP-dependent membrane current, I max is its maximum value, [cGMP] is the concentration of cGMP, K 1/2 is that concentration necessary to reach one half the value of I max, and n is a parameter that reflects the cooperative interaction of cGMP molecules in activating the membrane current.
The cGMP dependence of the membrane current depends on the history of exposure of the membrane patch to divalent cations. In Fig. 1, we present cGMP-dependent currents measured in a cone membrane patch in response to −40 mV voltage steps and in the presence of 20 μM Ca++ and 100 μM Mg++. Currents were measured in the same patch both before and after exposure to the EDTA/EGTA solution (Fig. 1, A and B). Also illustrated is the dependence of current amplitude on cGMP concentration (Fig. 1 C). The data points measured both before and after exposing the patch to EDTA/EGTA were well fit by Eq. 1, but the values of K 1/2 and n are different. The average value of K 1/2 shifted from 86.1 ± 18 to 58.8 ± 19 μM upon exposure to EDTA/EGTA. The average value of n shifted from 2.57 ± 0.34 to 1.80 ± 0.23 (Table II). Experimental data were included in these averages only if cGMP activation was measured in the same patch before and after exposure to the EDTA/EGTA solution. The shift in K 1/2 and n reflect a divalent cation-dependent mechanism that modulates the channel's sensitivity to the cyclic nucleotide.
Table II.
Modulation of Cyclic Nucleotide-gated Nucleotide-gated Currents in Membrane Patches of Rods and Cone * ‡
| Cone | Rod | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| K 1/2 | n | N | K 1/2 | n | N | |||||||
| cGMP | μM | μM | ||||||||||
| Before exposure to EDTA/EGTA solution | 86.1 ± 18 | 2.57 ± 0.34 | 8 | 41.1 ± 7 | 2.58 ± 0.43 | 9 | ||||||
| After exposure to EDTA/EGTA solution | 58.85 ± 19.3 | 1.8 ± 0.23 | 8 | 27.5 ± 6.2 | 1.97 ± 0.28 | 9 | ||||||
| With added calmodulin (200 nM) | 70.4 ± 17.5 | 2.1 ± 0.25 | 8 | 37.3 ± 8.8 | 2.44 ± 0.48 | 9 | ||||||
| 8-Br-cGMP | ||||||||||||
| Before exposure to EDTA/EGTA solution | 22.1 ± 3.1 | 2.29 ± 0.39 | 6 | |||||||||
| After exposure to EDTA/EGTA solution | 16.4 ± 3.8 | 1.54 ± 0.35 | 6 | |||||||||
Under all test conditions, currents were measured in the presence of 20 μM Ca++ and 100 μM Mg++.
The means under all conditions tested are statistically different from each other, with P < 0.02 as determined by a one-tail Student t test. The data for cGMP activation under the three experimental conditions specified were all measured in the same set of eight membrane patches.
To compare the functional properties of rod and cone cGMP-gated channels, we also investigated the Ca++-dependent modulation in patches from the rod outer segments of tiger salamanders (Fig. 1 D). In these membranes, in the presence of 20 μM Ca++ and 100 μM Mg++, K 1/2 for cGMP was 41.1 ± 7 μM and n was 2.58 ± 0.43 before exposure to EDTA/EGTA, and K 1/2 was 27.5 ± 6.2 μM and n to 1.97 ± 0.28 afterwards (Table II). Thus, in tiger salamander rods K 1/2 shifts by ∼1.5-fold, similar to findings in frog rods (Gordon et al., 1995). The fractional change in K 1/2 is thus similar in rod and cone membrane patches.
Modulation of K1/2 Is Not an Artifact Due to Phosphodiesterase Activity in the Membrane Patch
Detached outer segment patches can be structurally complex and may include not only plasma membrane but fragments of disc membranes containing phosphodiesterase (PDE) activity (Ertel, 1990). This may be a particular concern in cone outer segments, where disc and plasma membranes are continuous. The presence of PDE in the patch can lead to the artifactual appearance of variable cGMP titration curves (Ertel, 1990). We tested whether this might be a possible source of experimental artifacts in cone membrane patches by comparing cGMP activation curves in the presence and absence of 0.5 mM IBMX (3-isobutyl-1-methylxanthine), a saturating concentration of this effective cone PDE inhibitor (Gillespie and Beavo, 1989). IBMX had no effect whatsoever on the titration curves (data not shown).
We confirmed further that the modulation of K 1/2 is not an artifact due to the effect of PDE by measuring the properties of membrane currents activated by 8-Br-cGMP. This cGMP analog activates the channels but is not efficiently hydrolyzed by the photoreceptor PDE (Zimmerman et al., 1985). Fig. 2 illustrates currents activated by 8-Br-cGMP and measured in the same patch in the presence of 20 μM Ca++ and 100 μM Mg++ before and after exposure to EDTA/EGTA. As with cGMP activation, the dependence of current amplitude on 8-Br-cGMP was well described by Eq. 1 and removal of divalent cations shifted K 1/2 and n to lower values (Table I). Thus, modulation of the cGMP-gated currents does not reflect the activity of PDE.
Figure 2.

Modulation of 8-Br-cGMP-dependent activation of current in membrane patches detached from cone outer segments. (top) Currents measured at room temperature in the same membrane patch before (A) and after (B) exposure to the EDTA/ EGTA solution. Currents were activated with a voltage step from 0 to −40 mV in the presence of 20 μM Ca++ and 100 μM Mg++ and 8-Br-cGMP at 0, 5, 10 (arrow), 20, and 500 μM. C illustrates the dependence of current amplitude on 8-Br-cGMP before and after exposure to the EDTA/ EGTA solution. The continuous line is the best fit to the data of the Hill equation (Eq. 1). K 1/2 = 22 μM and n = 2.55 before exposure to the EDTA/EGTA solution, and K 1/2 = 15.5 μM and n = 1.66 afterwards.
Ion Permeation and Selectivity Are Not Different in the Two States of Ligand Affinity
We investigated whether the state of modulation affects other functional properties of the cGMP-gated currents. The shape of the I-V curve reflects, according to Eyring rate theory, the energy of interaction between permeant cations and their binding sites within the channel (Alvarez et al., 1992). The success of this theoretical analysis is measured by the ability to predict the shape of the I-V curves measured under various ionic conditions. The I-V curves of the cGMP-gated channels of both cones (Picones and Korenbrot, 1992; Haynes, 1995) and rods (Zimmerman and Baylor, 1992) can be predicted using Eyring rate theory, assuming the energy profile across the channels includes one binding site asymmetrically located within the membrane. If the interaction between permeant cations and the channel is affected by the state of modulation, then the shape of the I-V curve should change.
We compared in detail I-V curves measured in the same cone membrane patch before and after exposure to EDTA/EGTA. Fig. 3 illustrates membrane currents measured under symmetrical NaCl solutions with 20 μM Ca++ and 100 μM Mg++ on the cytoplasmic membrane surface and no divalent cations on the extracellular surface. Currents were measured in the presence of various cGMP concentrations in the range between 40 μM and 1 mM, and activated with a continuous voltage ramp between −70 and +70 mV. The I-V curves were nonlinear under all cGMP concentrations tested (Fig. 3). The nonlinearity is generated both by the voltage dependence of cGMP binding and the voltage dependence of divalent block (reviewed in Yau and Baylor, 1989). To analyze the I-V curves, we determined the voltage dependence of the binding curve for cGMP by fitting Eq. 1 to the current measured at every voltage between −70 and +70 mV. We found n not to change significantly with voltage either before or after exposure to EDTA/EGTA. K 1/2, on the other hand, was voltage dependent, but the form of the voltage dependency was quantitatively the same before and after exposure to EDTA/EGTA (Fig. 3). Thus, the I-V curves are not affected by the state of modulation. We obtained similar results in five other cone patches and four rod patches. Hence, the energy of interaction between cations and the channels, in both rods and cones, does not appear to be affected by the state of channel modulation.
Figure 3.

I-V curves measured in the same membrane patch detached from cone outer segment before (A) and after (B) exposure to the EDTA/EGTA solution. Currents were measured in the presence of 20 μM Ca++, 100 μM Mg++, and cGMP at 0, 40, 60 (arrow), 100, and 200 μM, and 1 mM on the cytoplasmic surface of the membrane. Currents were activated by a continuous voltage ramp between −70 and +70 mV. Each trace is the average of four current ramps from which the average ramp current measured at 0 cGMP has been subtracted. The leak conductance measured at 0 cGMP between 0 and −20 mV was 115 pS. (bottom) The voltage dependence of the K 1/2 (C) and n (D) terms in the Hill equation (Eq. 1) before (thick trace) and after (thin trace) exposure to the EDTA/EGTA solution.
We analyzed the relative Ca++ to Na+ permeabilities of the channels (PCa/PNa) in the two states of ligand affinity. This is a physiologically important feature since PCa/PNa differs in channels of rods and cones (Picones and Korenbrot, 1995; Frings et al., 1995). To determine the value of PCa/PNa, we measured I-V curves (with voltage ramps) under saturating cGMP concentrations (1 mM). The concentration of NaCl was symmetric across the membrane, and we imposed CaCl2 concentration gradients. The extracellular membrane surface was free of Ca++ and the cytoplasmic membrane surface was exposed to concentrations of 5 or 10 mM. The same patch was then exposed to the EDTA/ EGTA solution, and the current measurement repeated. Typical results are illustrated in Fig. 4. The reversal potential shifted towards negative values as the cytoplasmic Ca++ concentration increased, indicating that the channels are more permeable to Ca++ than to Na+. The magnitude of the shift was the same before and after exposure to EDTA/EGTA. Thus, the state of modulation does not affect the ion selectivity of the cone channels. We obtained the same results in three other membrane patches. The values of PCa/PNa, calculated from the shift in reversal voltage (Lewis, 1979), are similar to those we have previously reported (Picones and Korenbrot, 1995).
Figure 4.

I-V curves measured in the same membrane patch detached from a cone outer segment before (continuous trace) and after (○) exposure to the EDTA/EGTA solution. Currents were measured in the presence of 1 mM cGMP and symmetric Na+ (167 mM) solutions. The extracellular membrane surface was free of divalent cations, while the cytoplasmic surface was bathed with solutions containing 0, 5, or 10 mM Ca++. Currents were activated by a continuous voltage ramp between −70 and +70 mV. Each trace is the average of four current ramps, from which the average ramp current measured at 0 cGMP has been subtracted. The leak conductance measured at 0 cGMP between 0 and −20 mV was 70 pS. The value of membrane voltage at zero current reveals that the channels are more permeable to Ca++ than to Na+, but this selectivity is unaffected by the presence or absence of the endogenous modulator. For the data shown, PCa/PNa = 8.6 calculated at 5 mM assuming a constant field and using ionic concentrations, not activities (Lewis, 1979).
Effects of Ca++ on the Endogenous Modulation of cGMP Affinity
The removal of both Ca++ and Mg++ results in an irreversible shift of cGMP affinity. We investigated whether this shift was specific for either cation by testing whether the cGMP affinity shifted when only one of the two cations was removed. Removal of Mg++ alone did not shift the ligand affinity. Removal of Ca++ alone shifted the cGMP activation curve just as when both cations were removed, but the rate of this shift was slower than that caused by the simultaneous removal of both cations.
We explored the effects of varying Ca++ concentration on the shift in cGMP affinity. We measured the current activated by 60 μM cGMP in the presence of 20 μM Ca++ and 100 μM Mg++, exposed the patch for 60 s to solutions free of cGMP with the same Mg++ but progressively lower Ca++ concentrations, and then repeated the current measurement in the 20 μM Ca++, 100 μM Mg++ solution with cGMP. The current was unaffected at concentrations as low as 1 μM Ca++. Below this concentration, current amplitude increased as Ca++ concentration declined (Fig. 5). The increase in current amplitude at a fixed, nonsaturating cGMP concentration reflects shifts in K 1/2 and n to lower values. The current continued to increase, but even at 100 nM Ca++ the current increase was not maximal. Addition of EDTA/EGTA achieved the maximum current enhancement and additional washes in EDTA/EGTA caused no further current enhancement (Fig. 5). We made the same observations in six other patches. Because the shift in K 1/2 is irreversible, the experimental data do not reflect stationary conditions and, therefore, cannot be quantitatively analyzed as equilibrium dose– response data. Nonetheless, the results indicate that in detached patches, cGMP affinity is specifically modulated by Ca++ over a concentration range limited by ∼1 μM at its upper end.
The Effects of Exogenous Calmodulin
In rods, the Ca++-dependent modulation of ligand affinity has been attributed to the action of an endogenous factor similar, and perhaps identical, to calmodulin. Calmodulin confers Ca++ dependence to the cGMP activation of the channels with features that are almost indistinguishable from those of the endogenous modulator (Hsu and Molday, 1993; Gordon et al., 1995; Bauer, 1996). We explored the potential role of calmodulin in the modulation of channels in cone outer segments. In these experiments, we measured the cGMP concentration dependence of currents in the presence of 20 μM Ca++ and 100 μM Mg++. The membrane patch was tested before and after exposure to EDTA/EGTA, and then again in the continuous presence of calmodulin at a concentration of 200 nM. This concentration is effective in rod membrane patches (Gordon et al., 1995, Haynes and Stotz, 1997) and is well above the concentration that saturates modulation in rod membrane vesicles (Hsu and Molday, 1993; Bauer, 1996). In any event, the concentration of calmodulin used when testing its pharmacological action affects the Ca++ dependence of the phenomenon under study, but not its features at saturating Ca++ concentrations (see Bauer, 1996, for detailed mathematical analysis).
Fig. 6 illustrates typical results obtained in both rods and cones. In patches from both photoreceptor types, as expected, exposure to EDTA/EGTA lowered the values of K 1/2 and n in the cGMP titration curves. The addition of calmodulin in the presence of Ca++ shifted K 1/2 and n back towards their initial values. Whereas K 1/2 and n essentially reverted to their starting values in rods, the shift was never fully reversed in cones (Table II). Thus, channels of cones and rods differ in the effectiveness with which calmodulin in high Ca++ shifts their sensitivity to activation by cGMP. In cones, then, the Ca++-dependent regulation of cGMP activation is also likely to reflect the activity of an endogenous modulator. The modulator, however, may not be calmodulin, since this protein does not fully mimic the endogenous function.
Figure 6.

Effect of calmodulin on modulation of cGMP-dependent currents in detached membrane patches. In cone or rod patches, currents were activated with voltage steps from 0 to −40 mV in the presence of 20 μM Ca++ and 100 μM Mg++ and varying concentrations of cGMP. Measurements were repeated in the same patch before and after exposure to the EDTA/EGTA solution and in the continuous presence of 200 nM calmodulin. Data points were normalized by dividing current amplitude at each cGMP concentration (I) by the maximum current measured (Imax). The continuous line is the best fit to the data of the Hill equation (Eq. 1). In the cone patch, K 1/2 = 101 μM, n = 3.15 before exposure to the EDTA/EGTA solution, K 1/2 = 78 μM, n = 2.0 afterwards, and K 1/2 = 87 μM, n = 2.55 in the presence of calmodulin. In the rod patch, K 1/2 = 41.2 μM, n = 2.78 before exposure to the EDTA/EGTA solution, K 1/2 = 24.6 μM, n = 1.98 afterwards, and K 1/2 = 39.4 μM, n = 2.47 in the presence of calmodulin.
Calmodulin Competes with the Endogenous Modulator for Binding to the Channels
To test whether the endogenous modulator in cones and calmodulin share structural features, we tested whether the two compete in their binding to the channel. We investigated the effectiveness of added calmodulin to shift K 1/2 or n in the presence and absence of the endogenous modulator. If both molecules bind to the same site, then calmodulin should be without effect when added to the membrane in the presence of the endogenous modulator. That is, for calmodulin to be effective, the endogenous modulator must first be removed from its binding site.
We measured membrane currents with voltage steps between 0 and −40 mV in 20 μM Ca++ and 100 μM Mg++ in the presence of 60 μM cGMP. At this cGMP concentration, lowering K 1/2 and n will result in an increase in current amplitude despite an unchanging agonist concentration (see Fig. 1). In the same patch, we measured currents before and after adding 200 nM calmodulin. Calmodulin was without effect on current amplitude in membrane patches maintained in high Ca++ and Mg++ (Fig. 7 A). As expected, removal of the endogenous modulator by brief exposure to EDTA/ EGTA shifted K 1/2 and n and therefore increased the current amplitude (Fig. 7 B). After exposure to EDTA/ EGTA, added calmodulin was now able to shift K 1/2 towards its starting value (Fig. 7 B). We obtained the same results in every patch tested with this protocol (n = 6). Thus, while calmodulin and the endogenous modulator may not be the same, they appear to compete for a common site on the channels.
Figure 7.

Competition between calmodulin and the endogenous modulator in a cone outer segment membrane patch. Currents were activated with voltage steps from 0 to −40 mV in the presence of 20 μM Ca++ and 100 μM Mg++. Shown are difference currents measured by subtracting from currents measured at 60 μM cGMP those measured in the absence of the cyclic nucleotide. The leak conductance, measured in 0 cGMP at −40 mV was 325 pS. Currents in A were measured shortly after excision in the absence (thick trace) or continuous presence (thin trace) of 200 nM calmodulin. Currents in B were measured after exposing the same membrane patch to the EDTA/EGTA solution. Again, currents were measured in the absence (thick trace) or continuous presence (thin trace) of 200 nM calmodulin. Calmodulin is completely ineffective before the endogenous modulator is removed. If the endogenous modulator is first removed by exposure to the EDTA/EGTA solution, calmodulin then causes a shift in membrane current similar in direction, but smaller in extent than that caused by the endogenous modulator.
Ca++ Dependence of the Calmodulin-mediated Modulation
We investigated whether the channels of rods and cones differ in their interaction with Ca++/calmodulin by testing the Ca++ dependence of cGMP activation in the presence of 200 nM calmodulin. This Ca++ dependence is itself a function of the calmodulin concentration (Bauer, 1996) and therefore it does not inform us as to what the Ca++ dependence of modulation might be in the intact cell, if calmodulin were the modulator. However, it will reflect differences in the energetics of the modulator's binding site between the channels of the two cell types.
We measured the Ca++ dependence of currents in membrane patches of rods and cones in the presence of fixed concentrations of calmodulin and cGMP. In each experiment, membrane patches were first exposed to the EDTA/EGTA solution to remove the endogenous modulator. In patches from single cones, we measured currents generated by 110-ms voltage steps to −40 mV in the presence of 60 μM cGMP and varying Ca++ concentration between 0 and 20 μM. In the same patch, we measured the effects of Ca++ first in the absence and then in the presence of 200 nM calmodulin (Fig. 8). In cone membranes, Ca++ in the absence of calmodulin had a small but reproducible effect on current amplitude. The maximum change in current amplitude between 0 and 20 μM Ca++ in the absence of calmodulin was ∼5%. In the data shown, we subtracted this effect to obtain the effect of added calmodulin alone. We studied rod channels with the same protocols, except that 20 μM cGMP was used to activate the channel. Fig. 8 illustrates the Ca++ dependence of current amplitude in the presence of calmodulin in typical patches from both rod and cone channels.
Figure 8.

Ca++ dependence of the effect of calmodulin on rod and cone membrane patches. Patches were first exposed to the EDTA/ EGTA solution and then to one containing 200 nM calmodulin and various Ca++ concentrations. Currents were activated with voltage steps from 0 to −40 mV in the presence of 60 μM cGMP for cones and 20 μM cGMP for rods. For the cone, currents measured in the presence of cGMP and 0 and 300 nM, and 20 μM Ca++ are shown. For the rod, shown are currents measured in the presence of cGMP and 0 and 500 nM, and 1 and 20 μM Ca++. The current tracings also include the current measured in the absence of cGMP (nearly noiseless tracing). Data points are normalized cGMP-dependent current amplitudes calculated by dividing current amplitude at each Ca++ concentration (I) by the amplitude measured in the absence of any Ca++ (Izero). The continuous curve is the best fit to the data of Eq. 2. For cones, K i = 357 nM, n = 1.6, and I Θ/I zero = 0.73. For rods, K i = 632 nM, n = 2.4, and I Θ/I zero = 0.53.
The experimental data were well fit by the function:
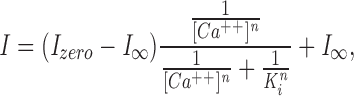 |
2 |
where I is the current, I zero is the current in the absence of Ca++, IΘ is the current in the presence of a saturating Ca++ concentration, [Ca++] is the Ca++ concentration, K i is the Ca++ concentration at which the current is inhibited by one half and n is a parameter that reflects cooperative interaction of cation binding. This is a modified Hill equation that indicates Ca++ interacts cooperatively with calmodulin to block the current amplitude. For cones, K i = 366 ± 131 nM, n = 1.6 ± 0.47, and I Θ/ I zero = 0.72 ± 0.16 (N = 9), while for rods, K i = 679 ± 187 nM, n = 1.81 ± 0.44, and I Θ/I zero = 0.53 ± 0.13 (N = 11). These values suggest that Ca++/calmodulin interacts with the cGMP-gated channel of both rods and cones, but the quantitative features of this interaction differ between the two receptor types.
discussion
The cGMP-gated ion channels in detached patches from cone outer segments exhibit a Ca++-dependent modulation of their affinity for the cyclic nucleotide. The dependence of current amplitude on cyclic nucleotide concentration is described by the Hill equation (Eq. 1) and modulation is manifested as a decrease in the apparent binding affinity (K 1/2) and cooperativity (n) as the Ca++ concentration is lowered. We will refer to this as the endogenous modulation of the channel. Endogenous modulation has been previously reported for the cGMP-gated channels of rods in bovine membrane vesicles (Bauer, 1996), detached frog outer segment patches (Gordon et al., 1995), and truncated outer segments from frogs and tiger salamanders (Nakatani et al., 1995; Sagoo and Lagnado, 1996). In general, the features of channel modulation in the intact cell should not be extrapolated using data from patches alone. In the case of rods, studies in nearly intact truncated outer segments have demonstrated Ca++-dependent modulation of K 1/2 that is quantitatively the same as that measured in detached membrane patches (Table III). The caveat has been introduced, however, that the modulation in truncated rods, which is measured under stationary conditions, may underestimate the extent of modulation present in the truly intact cell (Sagoo and Lagnado, 1996). In the case of cones, modulation measured under stationary conditions in the nearly intact outer segment is larger in extent than that observed in detached membrane patches, the K 1/2 shifts ∼4- rather than ∼1.5-fold (Rebrik and Korenbrot, 1997).
Table III.
Modulation of cGMP-activated Current in Rod Outer Segments
| Membrane patches | Truncated outer segments | |||||||
|---|---|---|---|---|---|---|---|---|
| K 1/2 | n | K 1/2 | n | |||||
| μM | μM | |||||||
| Endogenous modulator | 41.1 ± 7 | 2.58 ± 0.43* | 40 | 2.4‡ | ||||
| 37 ± 8 | 2.3 ± 0.6§ | |||||||
| Free of modulator | 27.5 ± 6.2 | 1.97 ± 0.28* | 27 | 2‡ | ||||
| 28 ± 6 | 2.2 ± 0.6§ | |||||||
Tiger salamander, this report;
Bullfrog, Nakatani et al., 1995;
Tiger salamander, Sagoo and Lagnado, 1996.
The endogenous modulation is Ca++ dependent, but the quantitative features of this dependence cannot be fully studied in membrane patches because modulation is irreversibly lost as Ca++ concentration is reduced and, therefore, equilibrium dose–response curves cannot be determined. In cone patches, the shift in K 1/2 is observed at concentrations starting at and below 1 μM Ca++ in the presence of 100 μM Mg++. This differs from data reported for modulation of channels in rods. In rod membrane patches, under experimental conditions similar to those we have reported for cones, modulation occurs starting at and below 22 nM (Gordon et al., 1995). This difference is significant and may suggest that, in cones, channel modulation may play a role in dim light, when only small changes in Ca++ concentration are expected, whereas in rods it may play a relevant role only in signals generated by relatively bright light (see Bauer, 1996). It is important to recognize that the Ca++ dependence of modulation reported for other rod preparation—for example, washed bovine membrane vesicles (Bauer, 1996) or amphibian truncated rods (Nakatani et al., 1995, Sagoo and Lagnado, 1996)—may differ from data in detached patches because in each preparation the conditions of equilibrium between the modulator and the channel may be different. In the detached membrane patch the effective concentration of unbound modulator is essentially zero. Therefore, the initial condition (high Ca++) is not at equilibrium and the modulator is kinetically “locked” onto the channel. The only information that can be reliably established is the Ca++ concentration at which the modulator becomes “unlocked” over a reasonably short time course (60 s).
The features of the interaction between the modulator and the channel in the intact cone photoreceptor and its functional role in transduction and/or adaptation are yet to be specified in detail. Although the magnitude of the modulation, a shift of ∼1.5-fold in K 1/2 may appear modest, the effect of this modulation on current amplitude can be expected to be large, particularly at low cGMP concentrations. From our experimental results, the change in current expected when Ca++ changes from 1 μM to 10 nM is given by:
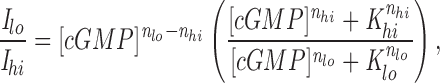 |
3 |
where I lo and I hi are the currents at 10 nM and 20 μM Ca++, respectively, [cGMP] is the ligand concentration, K lo and K hi are the values for K 1/2 and n lo and n hi are the values for n at 10 nM and 20 μM Ca++, respectively. Fig. 9 plots Eq. 3 with [cGMP] in units of K hi. Also shown are data points for currents activated by various cGMP concentrations and measured in a single cone patch before and after exposure to the EDTA/EGTA solution. The concentration of cGMP in the dark can be expected to be between 0.16× and 0.31× K hi since only 1–5% of the channels are open in darkness (Cobbs et al., 1985). Thus, if the modulation of the channel in the intact cell is similar to that in the patch, the amplitude of the light-sensitive current could change by as much as 5- to 10-fold in response to changes in cytoplasmic Ca++ concentration. As Eq. 3 indicates, and Fig. 9 illustrates, the effectiveness of Ca++ as a modulator increases dramatically as the cGMP concentration is lowered. Under steady illumination, when cGMP concentration is expected to be lower than in the dark, the physiological role of Ca++ modulation is likely to be especially significant.
Figure 9.

The effect of Ca++ modulation of ligand affinity on current amplitude as a function of cGMP concentration. Data points are the ratio of current amplitudes activated by cGMP at −40 mV in the same cone membrane patch before (Ihi) and after (Ilo) exposure to the EDTA/EGTA solution. cGMP concentration is expressed in units of K hi, the value of K 1/2 measured before exposure to the EDTA/EGTA solution. The continuous curve is the function presented in Eq. 3. The data shown are a single set selected because their values are similar to the statistical mean of our experimental sample. For the data shown, K hi = 88.4 μM, n hi = 2.73, and K lo = 61.4 μM, n lo = 1.59.
The endogenous modulation of K 1/2 in photoreceptor membrane patches is irreversibly lost upon removal of divalent cations, but can be restored, even if to a limited extent, by calmodulin. These results suggest that endogenous modulation arises from the activity of a “calmodulin-like” protein, since calmodulin not only restores modulation but also competes with the endogenous modulator for the same binding site. As has been argued in reports of similar phenomena in rod patches, it is unlikely that Ca++ modulation arises from phosphorylation since test solutions lacked nucleotides, or from phosphatase activity, because these enzymes are inhibited by lowering Ca++ (Gordon et al., 1995). Thus, modulation of both rod and cone channels likely arises from the activity of an endogenous factor that, like calmodulin, interacts with the channel in a Ca++-dependent manner: it is bound to the channels at high Ca++, dissociates from them in the absence of Ca++, and is then lost to the solution.
The molecular identity of the endogenous modulator is under investigation. Calmodulin exists at high concentration in the rod outer segment (Kohnken et al., 1981; Bauer, 1996). Hsu and Molday (1993) first reported that calmodulin causes a Ca++-dependent shift in K 1/2 in rod channels of thoroughly washed bovine outer segment membrane vesicles (see also Hsu and Molday, 1994; Bauer, 1996). Similarly, Ca++/calmodulin shifts the K 1/2 of channels in rod outer segment membrane patches (Gordon et al., 1995; Kosolapov and Bobkov, 1996; Haynes and Stotz, 1997). Gordon et al. (1995) compared the functional properties of the interaction of the endogenous modulator and calmodulin with the channels in frog rod outer segment membrane patches. Several features of this interaction were quantitatively different between calmodulin and the endogenous modulator, and the authors were not convinced the two molecules were the same. In contrast, Bauer (1996), in studies of bovine rod membrane vesicles, did not find quantitative differences between the endogenous modulator and added calmodulin. Thus, the endogenous modulator in rods is calmodulin-like, but it is not possible to confirm that it is calmodulin itself.
In contrast to the relative similarity between calmodulin and the endogenous modulator in studies of isolated rod membranes, added calmodulin was found ineffective in inducing a Ca++-dependent shift of K 1/2 in truncated rods of tiger salamander, from which the endogenous modulator was first removed (Sagoo and Lagnado, 1996). This finding is surprising because calmodulin, independently of whether it is the endogenous modulator, should have had an effect on the truncated outer segment current since it modulates the channel in membrane patches detached from the same cells (Fig. 6), as well as in patches from frog rods (Gordon et al., 1995; Kosolopov and Bobkov, 1996). We do not have a simple explanation for this experimental puzzle, but it is possible that calmodulin cannot efficiently gain access to the channels within the truncated outer segment.
Data are not available on the concentration of calmodulin in cone outer segments, nor its association with their cGMP-gated channels. In our direct comparison, we found that calmodulin did not quantitatively mimic the activity of the endogenous factor: calmodulin shifted K 1/2 and n to a lesser extent than did the endogenous factor. Indeed, Haynes and Stotz (1997) have reported that in patches of catfish cone outer segments, calmodulin entirely failed to modulate K 1/2. They did not, however, explore the properties of a possible endogenous modulator: in their experiments, the initial condition was to hold the membrane patch in a solution containing EDTA and EGTA to remove any endogenous modulator. Thus, while calmodulin does not appear to modulate K 1/2 in catfish cones, whether any modulation occurs at all is unclear. While it might be surprising that cones in striped bass exhibit modulation while those in catfish do not, it is possible. This issue should be addressed experimentally.
The failure of bovine calmodulin to fully mimic the action of the endogenous modulator in striped bass cones could be due to sequence differences between bovine and fish calmodulin, rather than the nonidentity between calmodulin and the endogenous modulator. This is not likely, however, since the amino acid sequence of calmodulin is nearly 100% identical among vertebrates (Friedberg, 1990). While the endogenous modulator may not be calmodulin in cones, the two molecules are likely to have structural features in common. Calmodulin is a member of a large family of proteins that contain EF hands, a Ca++-binding structural motif consisting of a select sequence of ∼30 amino acids that fold into a helix-loop-helix pattern (Kretsinger, 1979; Klee and Vanaman, 1982). Since the endogenous modulator binds Ca++ and competes with calmodulin for binding to the channels, it is probably also a member of this family of Ca++ binding proteins. We have found, however, that other EF hand-containing proteins expressed in photoreceptors, such as GCAP1 and GCAP2 (reviewed in Polans et al., 1996) do not restore modulation to the channels. Furthermore, these proteins do not compete with calmodulin for binding to the channels (Hackos, Palczewski, Baehr, and Korenbrot, unpublished observations). Knowledge of the specificity of interaction with the channel of different members of the EF hand protein family will be important for the eventual identification of the endogenous modulator.
The state of channel modulation does not affect other functional properties of the channel. In both rods and cones, the interaction and permeation of mono- and divalent cations with the channel are unaffected by the state of modulation, as is the voltage dependence of cGMP binding. Since these functional properties likely reflect the structure of the “pore” region of the channel, the interaction between the endogenous modulator and the channel probably occurs in a structural domain distant from the pore.
The interaction between calmodulin, Ca++, and the target protein is complex. The binding affinity of the three elements for each other changes depending on the identity of the target protein. For example, calmodulin in solution binds four Ca++ ions with affinities in the micromolar range, but in the presence of a target protein the affinity for Ca++ can be elevated by several orders of magnitude (reviewed in Klee, 1988; Gnegy, 1995). Moreover, the Ca++ dependence of a calmodulin-mediated effect on a given target protein changes with the mole ratio of calmodulin to target protein (Bauer, 1996). Therefore, the Ca++ sensitivity of the modulation by calmodulin observed in membrane patches of rods or cones cannot be assumed to be the same as in the intact cell unless the mole ratio of channel to calmodulin were the same. The differences in Ca++ dependence of K 1/2 modulation in rod and cone channels in the presence of calmodulin must reflect differences in the molecular details of the interaction of calmodulin with the channels.
Acknowledgments
We thank A. Picones for his patient instruction and continuing interest. We also thank C. Hackos, A. Olson, and T. Rebrik for their valuable comments and discussion.
Footnotes
D.H. Hackos is a Howard Hughes Medical Institute Predoctoral Fellow.
Abbreviations used in this paper: I-V, current–voltage; PDE, phosphodiesterase.
references
- Alvarez O, Villaroel A, Eisenman G. Calculation of ion currents from energy profiles and energy profiles from ion currents in a multibarrier, multisite, multioccupancy channel model. Methods Enzymol. 1992;207:816–854. doi: 10.1016/0076-6879(92)07058-v. [DOI] [PubMed] [Google Scholar]
- Balasubramanian S, Lynch JW, Barry PH. Calcium-dependent modulation of the agonist affinity of the mammalian olfactory cyclic nucleotide–gated channel by calmodulin and a novel endogenous factor. J Membr Biol. 1996;152:13–23. doi: 10.1007/s002329900081. [DOI] [PubMed] [Google Scholar]
- Bauer PJ. Cyclic GMP-gated channels of bovine rod photoreceptors: affinity, density and stoichiometry of Ca(2+)-calmodulin binding sites. J Physiol (Camb) 1996;494:675–685. doi: 10.1113/jphysiol.1996.sp021523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baylor D. How photons start vision. Proc Natl Acad Sci USA. 1996;93:560–565. doi: 10.1073/pnas.93.2.560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonigk W, Altenhofen W, Muller F, Dose A, Illing M, Molday RS, Kaupp UB. Rod and cone photoreceptor cells express distinct genes for cGMP-gated channels. Neuron. 1993;10:865–877. doi: 10.1016/0896-6273(93)90202-3. [DOI] [PubMed] [Google Scholar]
- Chen TY, Yau KW. Direct modulation by Ca(2+)- calmodulin of cyclic nucleotide-activated channel of rat olfactory receptor neurons. Nature (Lond) 1994;368:545–548. doi: 10.1038/368545a0. [DOI] [PubMed] [Google Scholar]
- Cobbs WH, Barkdoll AE, III, Pugh EN., Jr Cyclic GMP increases photocurrent and light sensitivity of retinal cones. Nature (Lond) 1985;317:64–66. doi: 10.1038/317064a0. [DOI] [PubMed] [Google Scholar]
- Ertel EA. Excised patches of plasma membrane from vertebrate rod outer segments retain a functional phototransduction enzymatic cascade. Proc Natl Acad Sci USA. 1990;87:4226–4230. doi: 10.1073/pnas.87.11.4226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fesenko EE, Kolesnikov SS, Lyubarski AL. Induction by cyclic GMP of cationic conductance in plasma membrane of retinal rod outer segments. Nature (Lond) 1985;313:310–313. doi: 10.1038/313310a0. [DOI] [PubMed] [Google Scholar]
- Finn JT, Grunwald ME, Yau KW. Cyclic nucleotide-gated ion channels: an extended family with diverse functions. Annu Rev Physiol. 1996;58:395–426. doi: 10.1146/annurev.ph.58.030196.002143. [DOI] [PubMed] [Google Scholar]
- Friedberg F. Species comparison of calmodulin sequence. Prot Seq Data Anal. 1990;3:335–337. [PubMed] [Google Scholar]
- Frings S, Seifert R, Godde M, Kaupp UB. Profoundly different calcium permeation and blockage determine the specific function of distinct cyclic nucleotide-gated channels. Neuron. 1995;15:169–179. doi: 10.1016/0896-6273(95)90074-8. [DOI] [PubMed] [Google Scholar]
- Furman RE, Tanaka JC. Monovalent selectivity of the cyclic guanosine monophosphate-activated ion channel. J Gen Physiol. 1990;96:57–82. doi: 10.1085/jgp.96.1.57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gillespie PG, Beavo JA. Inhibition and stimulation of photoreceptors phosphodiesterase by dipyridamole and M&B 22,948. Mol Pharmacol. 1989;36:773–781. [PubMed] [Google Scholar]
- Gnegy ME. Calmodulin: effects of cell stimuli and drugs on cellular activation. Prog Drug Res. 1995;45:33–65. doi: 10.1007/978-3-0348-7164-8_2. [DOI] [PubMed] [Google Scholar]
- Gordon SE, Downing-Park J, Zimmerman AL. Modulation of the cGMP-gated ion channel in frog rods by calmodulin and an endogenous inhibitory factor. J Physiol (Camb) 1995;486:533–546. doi: 10.1113/jphysiol.1995.sp020832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haynes LW. Block of the cyclic GMP-gated channel of vertebrate rod and cone photoreceptors by l-cis-diltiazem. J Gen Physiol. 1992;100:783–801. doi: 10.1085/jgp.100.5.783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haynes LW. Permeation of internal and external monovalent cations through the catfish cone photoreceptor cGMP-gated channel. J Gen Physiol. 1995;106:485–505. doi: 10.1085/jgp.106.3.485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haynes LW, Stotz SC. Modulation of rod, but not cone, cGMP-gated photoreceptor channel by calcium-calmodulin. Vis Neurosci. 1997;14:233–239. doi: 10.1017/s0952523800011378. [DOI] [PubMed] [Google Scholar]
- Haynes LW, Yau K-W. Single channel measurement from the cGMP-activated conductance of catfish retinal cones. J Physiol (Camb) 1990;429:451–481. doi: 10.1113/jphysiol.1990.sp018267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horn R, Patlak J. Single channel currents from excised patches of muscle membrane. Proc Natl Acad Sci USA. 1980;77:6930–6934. doi: 10.1073/pnas.77.11.6930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsu YT, Molday RS. Modulation of the cGMP-gated channel of rod photoreceptor cells by calmodulin . Nature (Lond) 1993;361:76–79. doi: 10.1038/361076a0. [DOI] [PubMed] [Google Scholar]
- Hsu YT, Molday RS. Interaction of calmodulin with the cyclic GMP-gated channel of rod photoreceptor cells. Modulation of activity, affinity purification, and localization. J Biol Chem. 1994;269:29765–29770. [PubMed] [Google Scholar]
- Hurley JB. Signal transduction enzymes of vertebrate photoreceptors. J Bioenerg Biomembr. 1992;24:219–226. doi: 10.1007/BF00762680. [DOI] [PubMed] [Google Scholar]
- Karpen JW, Zimmerman A, Stryer L, Baylor DA. Molecular mechanics of the cGMP-activated channel of retinal rods. Cold Spring Harbor Symp Quant Biol. 1988;53:325–332. doi: 10.1101/sqb.1988.053.01.039. [DOI] [PubMed] [Google Scholar]
- Klee, C.B. 1988. Calmodulin. In Molecular Aspects of Cellular Regulation. P. Cohen and C.B. Klee, editors. Elsevier, Amsterdam, Netherlands. pp. 35–62.
- Klee CB, Vanaman TC. Calmodulin. Adv Prot Chem. 1982;35:213–321. doi: 10.1016/s0065-3233(08)60470-2. [DOI] [PubMed] [Google Scholar]
- Kohnken RE, Chafouleas JG, Eadie DM, Means AR, Mcconnell DG. Calmodulin in bovine rod outer segments. J Biol Chem. 1981;256:12517–12522. [PubMed] [Google Scholar]
- Korenbrot JI. Ca2+ flux in retinal rod and cone outer segments: differences in Ca2+ selectivity of the cGMP-gated ion channels and Ca2+ clearance rates. Cell Calcium. 1995;18:285–300. doi: 10.1016/0143-4160(95)90025-x. [DOI] [PubMed] [Google Scholar]
- Kosolapov AA, Bobkov Y. Modulation of the cGMP- activated conductance of the plasma membrane of photoreceptor cells by calmodulin. Biochem Mol Biol Int. 1996;38:871–877. [PubMed] [Google Scholar]
- Kretsinger RH. The informational role of calcium in the cytosol. Adv Cyclic Nucleotide Res. 1979;11:1–26. [PubMed] [Google Scholar]
- Kurahashi T, Menini A. Mechanism of odorant adaptation in the olfactory receptor cell. Nature (Lond) 1997;385:725–729. doi: 10.1038/385725a0. [DOI] [PubMed] [Google Scholar]
- Lewis CA. Ion-concentration dependence of the reversal potential and the single channel conductance of ion channels at the frog neuromuscular junction. J Physiol (Camb) 1979;286:417–445. doi: 10.1113/jphysiol.1979.sp012629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu M, Chen TY, Ahamed B, Li J, Yau KW. Calcium-calmodulin modulation of the olfactory cyclic nucleotide-gated cation channel. Science (Wash DC) 1994;266:1348–1354. doi: 10.1126/science.266.5189.1348. [DOI] [PubMed] [Google Scholar]
- Martell, A.E., and R.M. Smith. 1974. Critical Stability Constants. Vol. 1. Plenum Press, New York.
- Menini A. Currents carried by monovalent cations through cyclic GMP-activated channels in excised patches from salamander rods. J Physiol (Camb) 1990;424:167–185. doi: 10.1113/jphysiol.1990.sp018061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller DJ, Smith GL. EGTA purity and the buffering of calcium ions in physiological solutions. Am J Physiol. 1984;246:C160–C166. doi: 10.1152/ajpcell.1984.246.1.C160. [DOI] [PubMed] [Google Scholar]
- Miller JL, Korenbrot JI. Differences in calcium homeostasis between retinal rod and cone photoreceptors revealed by the effects of voltage on the cGMP-gated conductance in intact cells. J Gen Physiol. 1994;104:909–940. doi: 10.1085/jgp.104.5.909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller JL, Korenbrot JI. In retinal cones, membrane depolarization in darkness activates the cGMP-dependent conductance. A model of Ca homeostasis and the regulation of guanylate cyclase. J Gen Physiol. 1993a;101:933–960. doi: 10.1085/jgp.101.6.933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller JL, Korenbrot JI. Phototransduction and adaptation in rods, single cones, and twin cones of the striped bass retina: a comparative study. Vis Neurosci. 1993b;10:653–667. doi: 10.1017/s0952523800005356. [DOI] [PubMed] [Google Scholar]
- Molday RS. Calmodulin regulation of cyclic-nucleotide-gated channels. Curr Opin Neurobiol. 1996;6:445–452. doi: 10.1016/s0959-4388(96)80048-1. [DOI] [PubMed] [Google Scholar]
- Nakamura T, Gold GH. A cyclic nucleotide-gated conductance in olfactory receptor cilia. Nature (Lond) 1987;325:442–444. doi: 10.1038/325442a0. [DOI] [PubMed] [Google Scholar]
- Nakatani K, Koutalos Y, Yau K-W. Ca2+ modulation of the cGMP-gated channel of bullfrog retinal rod photoreceptor. J Physiol (Camb) 1995;484:69–76. doi: 10.1113/jphysiol.1995.sp020648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Picones A, Korenbrot JI. Permeability and interaction of Ca2+ with cGMP-gated ion channels differ in retinal rod and cone photoreceptors. Biophys J. 1995;69:120–127. doi: 10.1016/S0006-3495(95)79881-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Picones A, Korenbrot JI. Permeation and interaction of monovalent cations with the cGMP-gated channel of cone photoreceptors. J Gen Physiol. 1992;100:647–673. doi: 10.1085/jgp.100.4.647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polans A, Baehr W, Palczewski K. Turned on by Ca2+! The physiology and pathology of Ca(2+)-binding proteins in the retina. Trends Neurosci. 1996;19:547–554. doi: 10.1016/s0166-2236(96)10059-x. [DOI] [PubMed] [Google Scholar]
- Pugh EN, Jr, Lamb TD. Amplification and kinetics of the activation steps in phototransduction. Biochim Biophys Acta. 1993;1141:111–149. doi: 10.1016/0005-2728(93)90038-h. [DOI] [PubMed] [Google Scholar]
- Rebrik T, Korenbrot JI. cGMP-gated conductance in intact outer segments of cone photoreceptors with an electropermeabilized inner segment. Investig Ophthalmol Vis Sci. 1997;38:S722. . (Abstr.) [Google Scholar]
- Reed RR. Signaling pathways in odorant detection. Neuron. 1992;8:205–209. doi: 10.1016/0896-6273(92)90287-n. [DOI] [PubMed] [Google Scholar]
- Sagoo MS, Lagnado L. The action of cytoplasmic calcium on the cGMP-activated channel in salamander rod photoreceptors. J Physiol (Camb) 1996;497:309–319. doi: 10.1113/jphysiol.1996.sp021770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scott SP, Harrison RW, Weber IT, Tanaka JC. Predicted ligand interactions of 3′5′-cyclic nucleotide–gated channel binding sites: comparison of retina and olfactory binding site models. Protein Eng. 1996;9:333–344. doi: 10.1093/protein/9.4.333. [DOI] [PubMed] [Google Scholar]
- Weyand I, Godde M, Frings S, Weiner J, Muller F, Altenhofen W, Hatt H, Kaupp UB. Cloning and functional expression of a cyclic-nucleotide–gated channel from mammalian sperm. Nature (Lond) 1994;368:859–863. doi: 10.1038/368859a0. [DOI] [PubMed] [Google Scholar]
- Williams DA, Fay FS. Intracellular calibration of the fluorescent calcium indicator Fura-2. Cell Calcium. 1990;11:75–83. doi: 10.1016/0143-4160(90)90061-x. [DOI] [PubMed] [Google Scholar]
- Yau KW, Baylor DA. Cyclic GMP-activated conductance of retinal photoreceptor cells. Annu Rev Neurosci. 1989;12:289–327. doi: 10.1146/annurev.ne.12.030189.001445. [DOI] [PubMed] [Google Scholar]
- Yau, K.-W., and T.-Y. Chen. 1995. Cyclic nucleotide-gated channels. In Ligand- and Voltage-gated Channels. R. Alan North, Editor. CRC Press, Boca Raton, FL. pg. 307–337.
- Zagotta WN, Siegelbaum SA. Structure and function of cyclic nucleotide-gated channels. Annu Rev Neurosci. 1996;19:235–263. doi: 10.1146/annurev.ne.19.030196.001315. [DOI] [PubMed] [Google Scholar]
- Zimmerman AL, Baylor DA. Cation interactions within the cyclic GMP-activated channel of retinal rods from the tiger salamander. J Physiol (Camb) 1992;449:759–783. doi: 10.1113/jphysiol.1992.sp019112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zimmerman AL, Yamanaka G, Eckstein F, Baylor DA, Stryer L. Interaction of hydrolysis-resistant analogs of cyclic GMP with the phosphodiesterase and light-sensitive channel of retinal rod outer segments. Proc Natl Acad Sci USA. 1985;82:8813–8817. doi: 10.1073/pnas.82.24.8813. [DOI] [PMC free article] [PubMed] [Google Scholar]