

Abstract
Cells expressing the hemagglutinin protein of influenza virus were fused to planar bilayer membranes containing the fluorescent lipid probes octadecylrhodamine (R18) or indocarbocyanine (DiI) to investigate whether spontaneous curvature of each monolayer of a target membrane affects the growth of fusion pores. R18 and DiI lowered the transition temperatures for formation of an inverted hexagonal phase, indicating that these probes facilitate the formation of negative curvature structures. The probes are known to translocate from one monolayer of a bilayer membrane to the other in a voltage-dependent manner. The spontaneous curvature of the cis monolayer (facing the cells) or the trans monolayer could therefore be made more negative through control of the polarity of voltage across the planar membrane. Electrical admittance measurements showed that the open times of flickering fusion pores were shorter when probes were in trans monolayers and longer when in cis monolayers compared with times when probe was symmetrically distributed. Open times were the same for probe symmetrically distributed as when probes were not present. Thus, open times were a function of the asymmetry of the spontaneous curvature between the trans and cis monolayers. Enriching the cis monolayer with a negative curvature probe reduced the probability that a small pore would fully enlarge, whereas enriching the trans monolayer promoted enlargement. Lysophosphatidylcholine has positive spontaneous curvature and does not translocate. When lysophosphatidylcholine was placed in trans leaflets of planar membranes, closing of fusion pores was rare. The effects of the negative and positive spontaneous curvature probes do not support the hypothesis that a flickering pore closes from an open state within a hemifusion diaphragm (essentially a “flat” structure). Rather, such effects support the hypothesis that the membrane surrounding the open pore forms a three-dimensional hourglass shape from which the pore flickers shut.
Keywords: membrane fusion, influenza hemagglutinin, membrane asymmetry, lipid flip-flop, planar bilayer membranes
introduction
Membrane fusion permits encapsulated material to move from one previously enclosed aqueous compartment to another. This is the dominant function of fusion in such cases as exocytotic secretion, fertilization of egg by sperm, and viral infection of cells (White, 1992). Movement of aqueous material occurs through fusion pores that connect the formerly distinct membranes. Does the ability of membranes to bend affect this movement? The work required to bend a membrane depends on the spontaneous curvature of that membrane. Membrane spontaneous curvature can be considered either by viewing the membrane as a single elastic sheet (Helfrich, 1973) or as composed of two monolayers, each with their own spontaneous curvature. Considerable effort has been devoted to determining how spontaneous curvature of contacting monolayers affects the establishment of fusion (Chernomordik et al., 1995a , 1997) and some work has examined how spontaneous curvature of noncontacting leaflets affects fusion (Chernomordik et al., 1995b , 1998; Melikyan et al., 1997a ). But until this study, no experimental effort has been devoted to determining the effect of spontaneous curvature of either of these leaflets upon fusion pores once they have been formed.
Initially, fusion pores have small lumens (Spruce et al., 1991; Lollike et al., 1995). Enlargement of these pores greatly facilitates release of vesicular contents in exocytosis (Alvarez de Toledo et al., 1993; Zhou et al., 1996), and, in the case of viral infection of cells (Hernandez et al., 1996; Kielian, 1995), it is essential that a fusion pore enlarge sufficiently to allow passage of the viral nucleocapsid into cytosol (Zimmerberg et al., 1994; Blumenthal et al., 1996; Melikyan et al., 1997b ). Not all fusion pores grow appreciably: some pores even close, a phenomenon known as flickering (Fernandez et al., 1984; Zimmerberg et al., 1987; Spruce et al., 1991). The factors that determine whether a pore closes or fully opens are not yet known. The lumen of a pore is contained by the membrane that forms the pore wall. The pore wall will evolve in a manner so as to minimize the total energy of the pore, contributed by protein and lipid. Because the membrane's energy must, at least in part, be a function of the deviation of its geometric curvature from spontaneous curvature, it is expected that spontaneous curvature should affect pore growth. But how, to what extent, and whether or not spontaneous curvature is critical to pore growth has yet to be investigated.
A systematic approach for measuring the effect of spontaneous curvature on pore growth would begin with a membrane in which the chemical compositions of the two monolayers are identical. In such a membrane, the two monolayers would bend in opposite directions to equal extents if they were not required to remain adhered to each other. Thus, the sum total of the membrane's spontaneous curvature is zero. In general, the spontaneous curvature of a bilayer is a sum total of the curvatures of its constituent monolayers. If the composition of each of the two monolayers could be alternately changed in identical fashions, membranes would in turn be produced that bend to the same extent, but in opposite directions. By measuring pore enlargement and contraction for each altered membrane, the influence of spontaneous curvature on pore evolution could be determined.
We carried out this strategy by fusing cells that express the fusion protein hemagglutinin (HA)1 of influenza virus to planar bilayer membranes. The lipid composition of the monolayers that comprise the bilayer can be controlled, thus also controlling spontaneous bilayer curvature. An amphipathic molecule with a large polar portion (in terms of cross-sectional area) relative to its hydrophobic portion will displace the headgroup region of a monolayer more than it will displace the acyl chain region. Amphiphiles of this shape are said to confer a positive spontaneous monolayer curvature. Inversely, an amphiphile with a hydrophobic portion large relative to its polar portion confers negative spontaneous monolayer curvature.
We showed that the amphiphiles octadecylrhodamine (R18) and indocarbocyanine (DiI) promote formation of inverted hexagonal (HII) phases in homogeneous lipid suspensions: this strongly suggests that they confer negative spontaneous curvature to the monolayer in which they reside. Both these cationic probes flip-flop from one leaflet to the other in a voltage-dependent manner (Melikyan et al., 1996). By altering polarity of voltage, we were able to change the spontaneous curvature of each monolayer of a voltage-clamped planar membrane containing either R18 or DiI by accumulating these molecules into either monolayer of the bilayer. We were also able to make the spontaneous monolayer curvature of either monolayer more positive by incorporating lysophosphatidylcholine in one or the other monolayer of the planar membrane. We thus were able to alternately make opposite spontaneous membrane curvatures of equal magnitude out of a symmetric planar membrane. After setting the spontaneous bilayer curvature, we triggered fusion. We found that the ability of small pores to close or fully open was strongly dependent on spontaneous membrane curvature: negative spontaneous bilayer curvature inhibits full pore enlargement and promotes an increased amount of flickering, whereas positive spontaneous curvature promotes the opposite effect.
materials and methods
Differential Scanning Calorimetry
Dielaidoylphosphatidylethanolamine (DEPE; Avanti Polar Lipids, Alabaster, AL) dissolved in chloroform/methanol (2:1, vol/vol) was mixed with varying amounts of a methanolic solution of one of the fluorescent probes. The solvent was evaporated with a stream of nitrogen and last traces removed in a vacuum desiccator for 2 h. Dried films were stored under argon at −20°C until used. The lipid films were then hydrated with 2 ml of 20 mM PIPES, 0.15 M NaCl, 1 mM EDTA, 20 mg/liter NaN3, pH 7.4 buffer. The mixture was heated to 45°C while vortexing vigorously to obtain a homogeneous suspension. The samples were made to a final DEPE concentration of 10 mg/ml and contained between 0 and 2% probe. The lipid suspension and buffer were loaded into the sample and reference cell, respectively, of a differential scanning calorimetry instrument. An MC-2 high sensitivity scanning calorimeter (Microcal Co., Amherst, MA) was employed at a heating scan rate of 37°K/h. The sample chamber held 1.4 ml. Continuous heating scans were run from ∼25 to ∼80°C. A single van't Hoff component was used to fit the recorded bilayer to hexagonal phase transition temperature; the reported transition temperature is that for the fitted curve.
Cell Culture and Treatment
NIH-3T3 fibroblasts stably transfected with HA from the A/Japan/305/57 strain of influenza virus, HAb2 cells (Doxey et al., 1985; Sambrook et al., 1985), were provided by J.M. White (University of Virginia, Charlottesville, VA). These cells were maintained and prepared for fusion experiments as described (Melikyan et al., 1995a , 1995b ).
Planar Membrane Formation and Cell Bilayer Fusion
Solvent-free horizontal lipid bilayers were formed from a solution of dioleoylphosphatidylcholine (DOPC)/dioleoylphosphatidylethanolamine (DOPE) (Avanti Polar Lipids, Inc.) 2:1 (wt/wt) and 5 mol% of ganglioside GD1a (Sigma Chemical Co., St. Louis, MO) with (or without) addition of 5 mol% octadecylrhodamine B chloride (R18; Molecular Probes, Inc., Eugene, OR) or 3 mol% 1,1′-dioctadecyl-3,3,3′3′-tetramethyl indocarbocyanine iodide (DiI; Molecular Probes, Inc.) dissolved in squalene (Aldrich Chemical Co., Milwaukee, WI) that had been passed through an activated alumina column. The ganglioside GD1a functioned as binding receptor for HA (Suzuki et al., 1986). Planar membrane were formed in a 150-μm diameter hole in a black Teflon® film with a brush technique and were bathed in symmetrical 140 mM NaCl, 2.5 mM KCl, 5 mM MgCl2, 2 mM CaCl2, 1 mM PIPES, pH 7.0, maintained at 35–37°C. Oleoyl lysophosphatidylcholine (LPC, Avanti Polar Lipids, Inc.) was included in either monolayer of the planar membranes by adding a stock solution (stock solution: 1 mg/ml lysophosphatidylcholine (LPC) in bathing solution with 10% methanol) to the appropriate aqueous solution bathing the bilayer, up to 16 μg/ml LPC. Membrane formation and cell contact with the bilayer were observed with an inverted microscope (Nikon Diaphot, Garden City, NY). A small amount (∼10 μl) of a concentrated cell suspension (∼107 cells/ml) in PBS was added to the solution above the planar membrane with a micropipette. The cells spontaneously sedimented and 15–25 cells rested upon the membrane within 30–50 s. The remainder of the cells came to rest on the Teflon® partition and never achieved contact with the bilayer. 4 min after the cells established contact with the lipid membrane, fusion was triggered by lowering the pH of the top, cell-containing solution to 4.9 by injecting 25 μl of concentrated isotonic succinate buffer directly over the bilayer.
Time-resolved Admittance Measurements
Cell-bilayer fusion was monitored by changes in the frequency-dependent bilayer admittance. For each membrane, we waited at least 6 min after acidification for a pore to form and, after a pore formed, at least 6 min for full pore enlargement. The bilayer was voltage clamped and a sine wave, superimposed on a holding potential of either +20, −20, or 0 mV, was applied to the trans, bottom solution bathing the membrane. The sign of voltage refers to the trans solution; the top, cis solution was grounded. When the holding potential was 0 mV, at least two sine waves of different frequencies were simultaneously applied so that pore conductances could be calculated. The in-phase (Y0), out-of-phase (Y90), and DC (YDC) components of admittance were calculated on-line with a software-based phase detector (Ratinov et al., 1998) and used to calculate pore conductance (Melikyan et al., 1995b ).
Sign Convention of Spontaneous Monolayer and Membrane Curvature
One can refer to the spontaneous curvatures of each monolayer or to the spontaneous curvature of the membrane. In accord with standard convention (Kozlov et al., 1989), we define the spontaneous bilayer (or membrane) curvature as positive if it naturally bends so as to bulge into the cis solution (i.e., the bilayer bends in a direction opposite the curvature of the cells at their point of contact). This direction of bending occurs if positive spontaneous monolayer curvature agents (e.g., lysophosphatidylcholine) are included in the cis leaflet or negative spontaneous curvature molecules are added to only the trans leaflet of an otherwise symmetrical bilayer. We define the bilayer curvature as negative if it bulges into the trans solution. This is accomplished either by adding negative spontaneous monolayer curvature agents within the cis leaflets or positive spontaneous monolayer curvature agents in the trans leaflets.
Data Analysis
Several characteristics of fusion activity were obtained to characterize the consequences of altering spontaneous monolayer curvature (see Fig. 2). (a) The lag time between triggering fusion by acidification and the opening of the first fusion pore (t lag) was measured. The probability P(t) that a pore had not formed within the planar membrane by time t was plotted as P(t) = N(t)/ N(0) where N(t) is the number of experiments for which pores have not yet formed by time t, and N(0) is the total number of experiments with pores. (b) The open times (t o) of flickering pores were characterized. (c) Pores that fully opened are referred to as successful. The time that a successful pore was open before its full enlargement (as determined by noting when the conductance increased in a stepwise fashion to conductances >60 nS), t S, was measured. (d) We also obtained the total time all pores were open, t o + t S, before the successful pore fully opened. (Experiments in which full pore enlargement was not observed, either because the membrane broke or only flickering occurred, were excluded from this analysis.)
Figure 2.
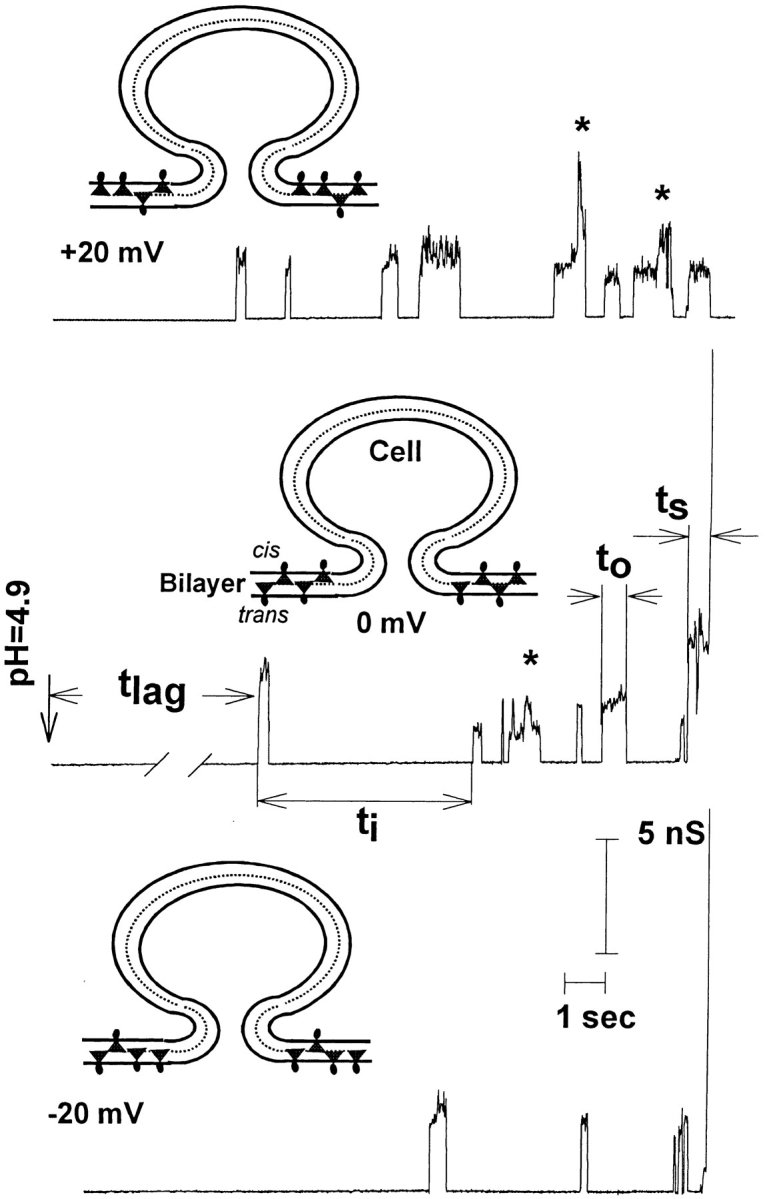
Schematic for pore evolution as a function of placement of R18 and DiI. Cells were adhered to planar membranes containing the cationic probes R18 or DiI. Probe was preferentially placed in the cis leaflet with 20 mV applied to the trans solution (top), symmetrically placed with 0 mV (middle), and preferentially placed in the trans leaflet with −20 mV (bottom). Typical fusion pore conductance records are shown for each case. When these negative-curvature probes (illustrated as molecules with their positively charged headgroup small compared with their membrane-imbedded portion) were situated in the cis leaflet, flickering occurred for extended periods and often a pore never fully enlarged. Fusion pores connecting a cell and planar bilayer are drawn as toroidal rather than as a pore within a hemifusion diaphragm. As is shown and elaborated in the discussion, open time distributions suggest a toroidal rather than a hemifusion structure at the pore flickering stage. Shown are the times from triggering fusion by acidification to pH 4.9 until the appearance of the first fusion pore, t lag, the time a flickering pore remained open, t o, the time between the opening of consecutive flickering pores, t i, and the time a successful pore had been open before full enlargement, t S. Approximately 20% of flickering pores were complex (*) and were excluded from open time distributions shown in other figures (Figs. 4–6). Conclusions for effects on open times by probe did not depend on whether or not complex pores were included for analysis. Long vertical line (middle and bottom right) shows abrupt full pore enlargement.
The conductance through an individual open pore was not constant in time, as it is for a classical ion-permeable channel. In the majority of cases, the variations in conductance were continuous, as expected of individual fusion pores (see Fig. 2). We refer to these pores as “simple.” For ∼20% of flickering pores, abrupt, discontinuous increases and decreases in conductance were superimposed on the continuous variations (e.g., pores marked by asterisks in Fig. 2). We refer to pores with discrete conductance jumps >1 nS as “complex” pores. These abrupt changes in pore conductance could be due to contractions and enlargements of a single pore between substates, or due to opening and closing of one pore while one or more others were still open, or to a combination of both. Because the true basis for a complex pore is unknown, open time distributions might have been corrupted if these pores had been included. We therefore present t o distributions for simple pores only. (But, in fact, when complex pores were treated as single entities, the effects of probes on open times of flickering pores proved to be independent of whether simple pores were segregated [as shown] or all pores were pooled.) Statistical differences for placement of probes within monolayers were, however, somewhat greater when complex pores were excluded from analysis.
Our criterion that distributions were indistinguishable was that P > 0.05 where P is the attained level of significance as evaluated by the nonparametric two-sample Kolmogorov-Smirnov test.
results
R18 and DiI Promote Negative Curvatures
R18 and DiI are positively charged amphiphiles with one or two C18-saturated hydrocarbon chains, respectively, causing them to partition into membranes. The manner by which R18 and DiI affect monolayer curvature was assessed by measuring the direction and extent by which they shift the bilayer to hexagonal phase transition temperature (TH) of a model phosphatidylethanolamine membrane. A plot of TH of DEPE as a function of the mole fraction of dye added to the membrane is shown in Fig. 1. Both probes exhibited a linear relationship between the lowering of TH and the mole fraction of probe added to the membrane. This indicates that both probes facilitated the formation of inverted phases with negative curvature. At a mole fraction of 0.01, DiI lowered the TH of DEPE by 3.6 ± 0.3°C, while R18 lowered this transition temperature by 1.4 ± 0.1°C. That is, a mole fraction of 0.01 R18 and 0.005 DiI lowered TH by approximately the same amount. For the fusion studies, we used 5 mol% R18 and 3 mol% DiI in the planar membrane. Thus, at these concentrations, the two probes should have altered spontaneous curvatures to roughly the same extent.
Figure 1.

The bilayer to hexagonal phase transition of DEPE was measured by differential scanning calorimetry as a function of the mole fraction of R18 and DiI added to the phospholipid. Both probes facilitated the formation of the negative curvature inverted phase. A linear relationship was exhibited between the amount of probe added and the observed transition temperature. Linear regression analysis gave correlation coefficients of 0.98 and 0.99 for R18 and DiI, respectively. A mole fraction of 0.01 R18 and 0.005 DiI lowered this transition temperature to the same extent.
The Kinetics of Fusion Are Dependent on the Presence of R18 or DiI, but Independent of Their Distribution between Monolayers
In our experimental arrangement, HA-expressing cells rested upon a horizontal planar bilayer. The top monolayer (or leaflet) of the planar membrane faced the cells and is referred to as the cis monolayer (Fig. 2). The bottom monolayer of the planar bilayer is referred to as trans. After settling cells on planar bilayers, fusion was triggered by acidifying the cis solution, thereby reconfiguring HA. Fusion pores appeared, typically within a few minutes. After opening, most pores closed shut (referred to as a “flickering” pore). At the present time, there is no experimental means of distinguishing whether repetitive flickering is due to successive opening and closing of distinct pores or of the same pore. For our analysis, we treated flickering pores as independent stochastic events. We refer (a posteriori) to a pore that remains open before fully enlarging as “successful.” With the DOPE/DOPC/GD1a membranes used in this study, in the absence of probe, a successful pore resulted for almost every experiment (Table I).
Table I.
Effect of Probe Asymmetry on Full Pore Enlargement
| Percentage of full pore enlargement | Median time from first pore opening until full enlargement | Number of flickers before full pore enlargement | Median open time of successful pore t s | |||||
|---|---|---|---|---|---|---|---|---|
| % | s | s | ||||||
| Without probes (−20 mV) | 93 | 35 | 4.1 | 0.7 | ||||
| Without probes (+20 mV) | 91 | 35 | 4.5 | 0.8 | ||||
| R18 cis | 57 | 60 | 10.0 | 0.7 | ||||
| R18 trans | 97 | 10 | 3.2 | 0.5 | ||||
| R18 symmetric | 100 | 25 | 4.9 | 0.8 | ||||
| DiI cis | 55 | 80 | 8.1 | 0.6 | ||||
| DiI trans | 91 | 15 | 3.6 | 0.4 | ||||
| DiI symmetric | 90 | 30 | 4.0 | 0.7 | ||||
| LPC trans | 100 | 10 | 0.4 | 1.5 |
The probes R18 and DiI flip-flop between monolayers and, as they are cationic, do so in a voltage-dependent manner, reaching steady state distributions of concentration within about a second (Melikyan et al., 1996). By including R18 or DiI in a planar membrane, the composition, and hence the spontaneous curvature of each monolayer, can be altered by changing the polarity of voltage. We applied 20 mV to the trans solution to drive the cationic probes to the cis monolayer and −20 mV to preferentially accumulate them in the trans monolayer (the cis solution was grounded). The probes were equally distributed between the two monolayers when 0 mV was applied (Fig. 2).
The distributions of times, t lag, from acidification until the occurrence of the first pore (regardless of whether it closed or fully opened) were generated to characterize the kinetics of fusion. The presence of R18 or DiI shortened these “waiting times” (Fig. 3, left), but which monolayer the probe preferentially resided in was not of consequence (right). As a control, we also measured waiting times for pore formation when rhodamine-phosphatidylethanolamine (rho-PE) was included in the bilayer. This probe is negatively charged and does not flip-flop (Melikyan et al., 1996). (The rhodamine of rho-PE is a lissamine rhodamine. It contains a sulfonyl group with localized charge. This rhodamine is chemically distinct from that of R18.) rho-PE raises the transition temperature for formation of HII phases by 144°C/mol fraction of probe (data not shown), from which we infer the probe promotes positive spontaneous monolayer curvature. That is, rho-PE induces an opposite curvature to that induced by R18 or DiI, and it has an opposite charge. But, as did R18 and DiI, its presence somewhat shortened the waiting time distributions for pore formation (Fig. 3, left). Similarly, LPC, well known to have large positive spontaneous curvature (Epand, 1985; Tilcock and Cullis, 1987; Kumar et al., 1989), placed within the trans leaflet, also led to shorter lag times (Fig. 3, right).
Figure 3.

Waiting time distributions of time lags from triggering fusion until the first fusion pore appeared in the presence of various probes. The addition of R18, DiI, or rho-PE to DOPC/ DOPE/GD1a bilayers led to shorter lag times, but lag times were independent of the partitioning of probes between leaflets. Similarly, LPC trans shortened lag times.
The time between the appearance of flickering pores, referred to as an interflicker interval (t i), provides another measure of fusion kinetics. For both R18 and DiI, the t i distributions were statistically the same in the absence and presence of probe, regardless of whether probe was preferentially situated in the cis or trans monolayers or symmetrically distributed (data not shown). In short, the kinetics of pore formation was independent of spontaneous curvature over the range studied for R18, DiI, and rho-PE.
Asymmetry of Probe Distribution Affects Properties of Open Pores
The asymmetry of probe location between the two monolayers comprising the planar membrane did, however, strongly affect the likelihood that an HA- induced pore would close or one would fully enlarge. If either DiI or R18 was enriched in the cis monolayer, successful pores did not always occur: a pore fully enlarged in only half of the experiments (Table I). For experiments in which no pore fully enlarged, frequently pores would open and then close throughout the full 6-min period between pore formation and termination of the experiment. If probe was enriched in the trans leaflet or symmetrically placed, a successful pore almost always occurred. In short, the negative curvature agents inhibited full pore enlargement if preferentially located in the cis monolayer of a bilayer. Even when a successful pore formed with probe enriched in the cis monolayer, the time that pores remained in a stage of repetitive flickering was greatly increased above that obtained when the concentration of probe was the same in both monolayers, which in turn was longer than when probe was preferentially placed in the trans monolayer (Table I, median time from first pore, usually a flickering one, until a pore fully enlarged). In accord with the duration of the flickering stages, a greater number of flickering pores occurred before formation of a successful pore when the probes were preferentially placed in the cis leaflet as opposed to the trans leaflet. In contrast, initial pore conductances were independent of the presence and location of probe (data not shown).
For 20 mV, the excess of probe was not large (Melikyan et al., 1996) (∼70% of the R18 or DiI was in the enriched leaflet, 30% in the depleted one), emphasizing how strong an effect the probe indeed had on pore enlargement. We would have preferred to apply larger holding potentials to exaggerate the asymmetry of probe within the two monolayers, but bilayer stability was reduced by cells and fusion; it was thus not practical to maintain larger potentials. The polarity of the bilayer holding potential per se did not affect the probability of full pore enlargement: without any probe, successful pores almost always formed for both +20 and −20 mV. In fact, the distribution of all times we have measured (times from acidification until the appearance of a pore, open times for flickering pores, total time period over which flickering pores were observed, interflicker intervals, open times for successful pores until full enlargement, and the sum total open times for all pores, both flickering and successful, before the moment of full pore opening) were independent of voltage polarity when probes were not included or when probes that could not flip-flop (i.e., rho-PE) were included in the membrane. Thus, properties we report were due to the disposition of probe, not to the voltage used to situate them.
Open Times of Flickering Fusion Pores Are Longer When Either R18 or DiI are Preferentially Present in the Cis Monolayer than When They Are Located in the Trans Monolayer
Open times of HA-induced flickering pores were strongly affected by the placement of probe. Open times were significantly longer for either R18 or DiI in the cis monolayer and shorter if in the trans monolayer, compared with open times for probe at the same concentration in each monolayer (Fig. 4). The same distributions of open times were obtained in the absence of probes as when probes were symmetrically placed (Fig. 4, bottom). That is, the presence of probe per se did not affect open times. Rather, asymmetric placement of the probe between the two monolayers of the bilayer caused the changes in open time distributions. As we emphasize in the discussion, it is significant that open times in the absence of any probe were the same as for probe divided equally between the two monolayers.
Figure 4.
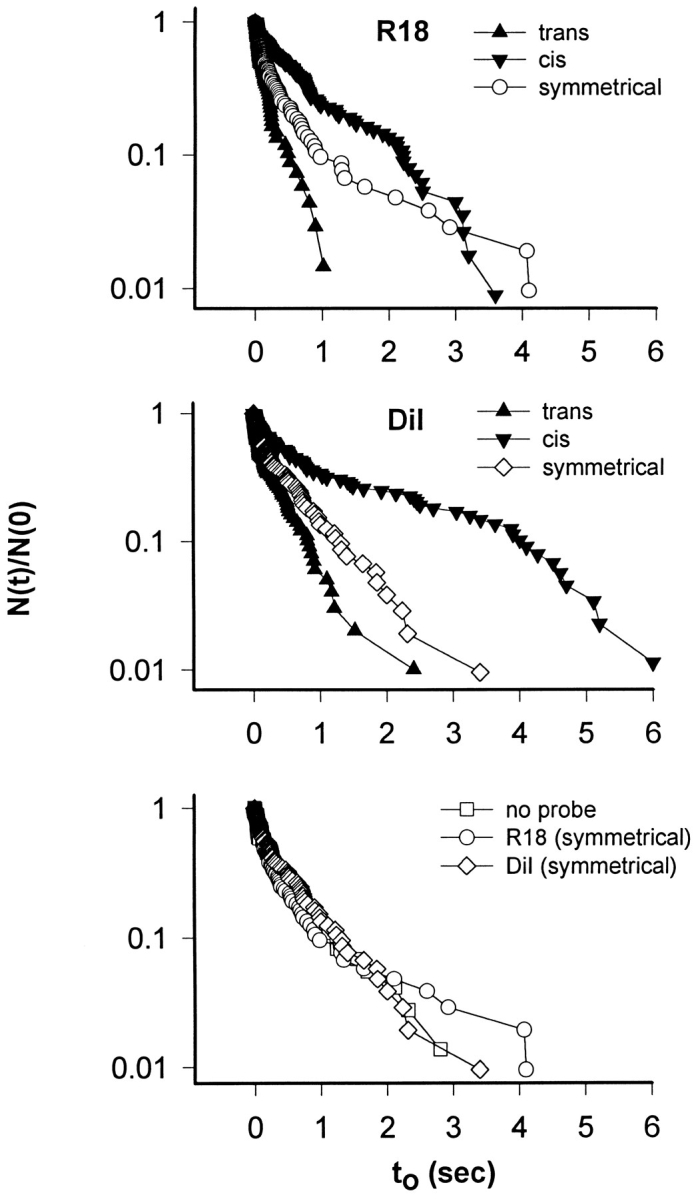
The effect of R18 (top) and DiI (middle) on distributions of open times (t o) for flickering pores. Open times were longer when probe was located in the cis leaflet than in the trans leaflet and intermediate when probes were symmetrically placed. Importantly, open times were the same for all symmetrical membranes (bottom), regardless of whether probe was present
Probe Affects Open Times While Pores Flicker, but Not before Pore Formation
The placement of probe could affect open times of flickering pores by its presence in the membrane comprising the pore wall while the pore is open. Alternatively, there may be more than one population of pore types, and probe in the membrane could be influencing the selection of pore structure leading to the formation of a specific type of pore. That is, some pores may intrinsically have shorter open times than others, and probe placement could be affecting the formation of a particular type. We sought to distinguish between the two possibilities by starting with probe in the trans monolayer, switching voltage after the first flickering pore closed (thus moving probe to cis monolayer), and then determining whether new pores followed with short or long open times. For DiI, open times of the first new pore after moving probe to the cis leaflet were long (Fig. 5, right, □), as were open times of all pores that formed subsequent to switching voltage (not shown). Further, they were statistically indistinguishable from open times of pores that formed when probe was originally placed cis and never flipped (Fig. 5, ▾, replotted from Fig. 4 for convenience of comparison). Also, when probe was originally in the trans leaflet, after flipping of probe, pores tended to flicker repetitively for extended times (Fig. 5, top) and in many cases no pores fully opened, the same phenomena observed when DiI was always cis. For R18, all the above phenomena were observed with one exception: while open times of the first new pore after flipping the probe to the cis leaflet were noticeably shifted visually to longer times (Fig. 5, left, □ vs. ▴), the differences were not statistically significant. Since only the first pore after switching voltage is counted for this analysis, it is possible that the small sample size is obscuring a true lengthening of open times. Support for open time lengthening is shown by the fact that the open times of the second new pore after switching voltage was statistically longer than when R18 was always trans (Fig. 5, ⋄). In short, pore properties did differ according to the location of probe after pore formation, and therefore it is likely that probe exerts its effect as a lipid constituent of the pore wall.
Figure 5.

Open times of flickering pores can be switched during flickering. (top) R18 or DiI were preferentially located in the trans monolayer (−20 mV). After the first flickering pore closed, voltage polarity was changed to 20 mV to move probe to the cis monolayer and open times of pores that followed were obtained. For DiI (right), the first new pore that formed after probe was moved (□) had the same open times as pores that occurred when probe was always cis (▾, replotted from Fig. 4). For R18 (left), the first new pore after switching voltage polarity had shifted to longer times, but open times had not statistically lengthened until the second new pore formed.
The Location of Probe Affects Total Open Times of Pores Before Final Full Enlargement
The time from opening of a successful pore until its full enlargement, t S, was independent of whether R18 or DiI was included in the planar membrane and was independent of asymmetries of probe location (Table I). In distinct contrast, the total times flickering pores and the successful pore were open before a full enlargement (the sum of all t o plus t S) did depend on which leaflet was enriched by probe (Fig. 6): the greater number of flickers and longer open times when probe was predominantly in the cis leaflet (▾) led to total open times that were significantly larger than when probe was in the trans leaflet (▴). The total open times were the same in the absence of probe (□) as when probe was symmetrically placed by maintaining a 0-mV holding potential (○).
Figure 6.

The sum total of times all flickering pores plus the subsequent successful pore were in open states before full pore enlargement. Total open times were the same for all symmetrical membranes, regardless of whether probe was present. For both R18 (left) and DiI (right), total open times were the greatest for probe accumulated in the cis monolayer and least for probe accumulated in the trans monolayer.
LPC in Cis Leaflets Inhibits Fusion, LPC in Trans Leaflets Promotes It
The time it takes for LPC to flip-flop is much longer than the time scale of our experiments (Mohandas et al., 1982). We added LPC, which has positive spontaneous curvature, in separate experiments to the cis or trans solution so that it would incorporate into the respective monolayer. With LPC (16 μg/ml) in the cis solution, fusion was largely if not completely abolished. We did, however, observe small isolated conductance fluctuations. These were probably due to the lytic effects of LPC on bilayers (Chernomordik et al., 1985), rather than to the presence of small flickering fusion pores because the changes in admittance were relatively uniform in time and smaller than the changes observed when pores flickered in the absence of LPC.
There were not many flickers with LPC in the trans leaflet: the first pore usually fully enlarged after a relatively long t S (Table I). When flickering did occur, on average only one flicker every other experiment (Table I), the openings had extremely short duration (47 ± 17 ms, n = 8). In other words, when LPC resided within the trans leaflet, the total open time represents a single successful pore.
discussion
Flickering of fusion pores occurs in both viral and exocytotic membrane fusion. The present study shows that pore closure, which defines flickering, is strongly regulated by lipid asymmetry. The regulation of pore closure by this asymmetry can be understood on the basis of spontaneous bilayer curvature. The lipid compositions of inner and outer monolayers of cell membranes are quite different, with amino phospholipids and phosphoinositides in inner leaflets and choline-containing phospholipids (PC and sphingomyelin) enriched in outer leaflets (Devaux, 1993). Pore flickering in all protein-mediated fusion systems should therefore be affected by membrane asymmetry. Experimentally, R18 and DiI are commonly used to detect membrane continuity for many fusion systems (Cohen and Melikyan, 1998). These cationic probes should accumulate in inner leaflets of cell membranes (Sheetz and Singer, 1974). In any case, as their presence must have some effect on pore growth, the manner and extent to which these fluorescent probes affect pores is of practical consequence for the accurate interpretation of data.
The Ability of a Lipid to Flip-Flop Is Consistent with Its Ability to Confer Negative Spontaneous Monolayer Curvature
Differential scanning calorimetry shows that R18 and DiI lower the transition temperature at which DEPE forms HII phases (TH), strongly suggesting that the probes promote lipid–water interfaces with negative spontaneous curvature. Shifts in TH caused by other amphiphiles are quantitatively similar for DEPE and 1-palmitoyl-2-oleoylphosphatidylethanolamine (Epand, 1985) as well as dipalmitoylphosphatidylethanolamine (R.M. Epand, unpublished observations). That is, the shifts in TH for these three lipids are affected by the added amphiphiles, rather than by the host lipid. If this should prove to be generally the case, R18 and DiI would lower TH for the DOPC/DOPE lipid mixture used for our fusion pore studies. However, we did not verify that the probes lowered TH for the DOPC/DOPE mixture because the mixture's lamellar to HII transition temperature was not experimentally accessible.
It is perhaps surprising that R18 has negative spontaneous curvature—its large headgroup and single acyl chain could lead one to assume that its “molecular shape” would confer a positive spontaneous curvature. But the very fact that R18 flip-flops shows that the headgroup of R18 must readily translocate across the low dielectric of the interior of a bilayer (Ganong and Bell, 1984), indicating that the headgroup is relatively hydrophobic. Further, the voltage dependence of charge movement shows that the charge of R18 responds to only ∼85% of the applied field (Melikyan et al., 1996), strongly indicating the headgroup is somewhat buried within the hydrophobic core of the lipid bilayer. Therefore, the headgroup of R18 does not appear to reside entirely in the aqueous phase. The polar portion of the rhodamine moiety is probably small, with much of the headgroup contributing, in actuality, to the bulk of the hydrophobic tail of the molecule, the molecular shape envisioned for a negative spontaneous curvature substance. Since flip-flop and negative spontaneous curvature both require that a lipid have relatively low polarity, it may be that lipids that flip-flop, in general, also promote negative spontaneous curvature (Bröring et al., 1989; Epand et al., 1991).
Short Flicker Times Correlate with the Propensity of Pores to Fully Enlarge
Proteins control membrane fusion. However, membrane merger and, therefore, the extent and kinetics of fusion can be modulated by adding high concentrations of amphiphiles to cis leaflets. The amphiphiles probably act by altering spontaneous curvatures (Chernomordik et al., 1995a ). For the comparatively low concentrations of R18, DiI, and rho-PE used in this study, any changes in the kinetics of fusion pore formation (as measured by either t lag or t i) did not depend on spontaneous curvature of the bilayer. The addition of probes to either leaflet always reduced t lag, regardless of the sign of spontaneous curvature or electrical charge, and t i was never affected. Clearly then, changes in spontaneous curvature are not responsible for shortening of lag times. It may be that the addition of any lipid component promotes formation of fusion pores by allowing a more favorable combination of lipids to assemble into the configuration of a pore.
We previously observed that when surfaces of HAb2 cells were treated with varied proteases that do not affect HA (e.g., collagenase and hyaluronidase), the shorter the open flicker times, the more likely a pore would fully enlarge (Melikyan et al., 1995a ). The present study employs the same HAb2 cells and the same correlation has been observed, although the means of altering conditions were quite different from the first study. Thus, short flickers and full pore enlargement may be connected, possibly controlled by the same energetic factors.
Pore Evolution Is Regulated by Spontaneous Membrane Curvature
The addition of lipid probes into a membrane can perturb many physical properties, such as hydration energy, surface tension, line tension, and spontaneous curvatures. Of course, these parameters are not independent—changes in one are often associated with changes in the others (Kozlov et al., 1994). Symmetrical bilayers (each monolayer has the same lipid composition) with and without probe have the same overall spontaneous membrane curvature, zero. In contrast, other physical parameters will, in general, differ amongst symmetrical bilayers that do not have identical compositions. All characteristics of open pores were found to be the same for symmetrical bilayers regardless of whether R18, DiI, or rho-PE were present. This lends support to the view that these probes affected pore behavior, once the pore has formed, through changes in spontaneous membrane curvatures alone, regardless of changes in other physical parameters. Further, the invariance of pore behavior for all symmetrical membranes demonstrates that the effects of probes can be discussed not only in terms of the spontaneous curvature of each individual monolayer, but also in terms of spontaneous membrane curvature.
We note that we were not able to study pore behavior when LPC was symmetrically placed in the bilayer since LPC within the cis leaflet prevents fusion. Therefore, we cannot rule out the possibility that LPC in the trans leaflet alters pore properties by means other than through changes in spontaneous curvature. In fact, LPC significantly lowered membrane tension (Chernomordik et al., 1985); its addition caused the bilayer to readily bulge (our unpublished observations). In contrast, the presence of R18 or DiI caused no bulging, independent of its location. Therefore, it may be that R18 and DiI affect pore properties through curvature effects, while LPC exerts its action through changes not only in spontaneous curvature but also in membrane tension.
In fact, while the actions of R18 and DiI on pore flickering can be understood on the basis of their spontaneous monolayer curvatures and placement within monolayers, full pore opening cannot be explained by membrane asymmetry. R18 and DiI within the cis leaflet should have the same effect on full pore enlargement as LPC within the trans leaflet, based on their spontaneous monolayer curvatures. Instead, the effects of R18 and DiI were opposite that of LPC. For R18 and DiI to have the same effect as LPC trans (promotion of pore enlargement), they too had to be in the trans leaflet. Thus, while all probes strongly affected pore enlargement, alterations in spontaneous curvature cannot be responsible in every case: all probes may be contributing to full pore enlargement through changes in spontaneous curvature, but, for some, alterations in another parameter may be the dominant factor.
The act of probe flip-flop itself could have an effect on membrane properties. In a closed system that contains a constant number of lipid molecules, such as vesicles or cells, flipping of probes would cause mismatches in the total area of the cis (outer) monolayer relative to the trans (inner) monolayer. Area mismatches cause energy changes that directly result in shape changes (Sheetz and Singer, 1974; Steck, 1989; Döbereiner et al., 1997), independent of spontaneous curvature. Because the number of lipids is not fixed within the planar bilayer (there is a reservoir of lipid from the supporting Gibbs-Plateau border), lipid flip-flop in planar bilayers does not lead to area mismatches. Flip-flop of R18 and DiI would not lead to a static redistribution of probe between leaflets of the bilayer, but rather to a continual movement of probe via the Gibbs-Plateau border. We cannot exclude the possibility that such lipid flow, if it occurs, would have some effect on pore properties, with the sign dependent on the direction of flow. But for this study, we assume that such effects are negligible or nonexistent.
Probes Exert Their Effects while They Are Constituents of the Pore Wall
If, once a pore formed, probe could be induced to instantaneously flip from one monolayer to another, it could be unambiguously determined whether the open times depended on the probe distribution only while a pore was open, or depended on probe distribution before opening. Because flip-flop times of R18 and DiI at 20 mV are comparable to mean open times, this experiment could not be performed—pores would close before probe had reached its new steady state distribution. It was practical, however, to flip the probe after one pore had already opened and closed, and then obtain the open times of all subsequent pores. The time from triggering fusion until the first pore opened, t lag (Fig. 3), was much longer than times between the opening of successive pores, t i. Thus, if the distribution of probe were to influence the opening of one type of pore over another by affecting slow processes (such as the association between HA trimers) that occur during the long lag before initial pore formation, the open times of pores would be independent of whether probes were flipped after the lag. In the case of DiI, the first pore after flip-flop already exhibited open times that were characteristic of times one would see if probe had always been in the new location (Fig. 5). For R18, the second pore after flip-flop exhibited such characteristics. It remains formally possible that probe location affects a process immediately before pore opening, thereby selecting a flickering pore with an intrinsically short or long open time. We favor the view that probes affect pore growth while incorporated in the membrane that comprises a pore wall while that pore is open.
A Flickering Pore Has the Properties of a Three-Dimensional Structure Rather Than Those of an Opening in a Two-Dimensional Hemifusion Diaphragm
It is often conjectured that fusion proceeds through an intermediate stage known as hemifusion (Kemble et al., 1994; Melikyan et al., 1995c; Bagai and Lamb, 1996; Nüssler et al., 1997; Chernomordik et al., 1998; Cleverley and Lenard, 1998). Hemifusion is defined as the merger of cis leaflets of two membranes, during which aqueous contents remain separated by a single bilayer membrane that has formed from the two unperturbed trans leaflets. Fusion would be completed if a pore formed within this new membrane (known as a hemifusion diaphragm).
Our data indicates that even a pore still small enough to close is already a three-dimensional structure (i.e., a structure with length greater than the thickness of a bilayer; Fig. 7, bottom left), a “tunnel” connecting aqueous phases rather than simply an opening within a hemifusion diaphragm. We infer this based on the following considerations: within a hemifusion diaphragm, only lipids from trans leaflets could contribute to pore structure (Fig. 7, top left). Probes located in cis (contacting) monolayers probably have no effect on open time distributions of flickering pores; t o distributions should be accounted for entirely by the concentration of probe within the trans monolayers. The greater the amount of probe within the trans leaflet of the bilayer (the bilayer leaflet contributing to the hemifusion diaphragm), the greater should be the effect on the t o distribution; the sequence of order for open times should follow the sequence of concentration of probe within the trans leaflet. In fact, probe asymmetry determined t o distributions: without probe, the same t o distribution was observed as when probe was equally divided between monolayers. In other words, open times were the same for symmetrical membranes, whatever the concentration of probe. Therefore, we conclude that a pore wall is composed of a fully integrated membrane, with both inner and outer monolayers. Such a structure would be roughly hourglass shaped, referred to as “toroidal” (Fig. 7, bottom left) and would, of necessity, have some length. If fusion proceeds through hemifusion, it is likely that the initial pore forms while the extent of hemifusion remains local. If so, it would already be roughly toroidal. Alternatively, it is possible that the fusion pore does originate within an extended hemifusion diaphragm and the diaphragm quickly retracts into the membrane wall before pore closure.
Figure 7.

A toroidal pore (bottom left) but not a pore within a hemifusion diaphragm (top left) accounts for the dependence of open times on bilayer asymmetry. For a pore within a hemifusion diaphragm, only lipids from trans leaflets contribute to the pore wall. For a toroidal pore, the pore wall is a bona fide synthesis of the cell and bilayer membrane. A pore has two geometric curvatures: parallel (1/ R2) and meridian (1/R1). The negative meridian curvature of a toroidal pore favorably accommodates a negative spontaneous monolayer curvature probe within the cis leaflet but not within the trans leaflet (bottom left). Thus, the energy of a small pore is reduced by negative spontaneous monolayer curvature probe within the cis leaflet, but increased by probe within the trans leaflet (right). A small pore with lowered energy is stabilized: it neither closes nor fully opens as readily as a higher energy, destabilized one.
The Energy of a Toroidal Flickering Pore Is Controlled More Strongly by Its Negative Than Positive Curvature
The membrane of a toroidal pore has two orthogonal curvatures (Fig. 7, left): the negative meridian (longitudinal) curvature (1/R1) and the positive parallel (latitudinal) curvature (1/R2) (Struik, 1961; Kozlov et al., 1989; Chernomordik et al., 1995b ). For the meridian curvature, molecules that promote negative spontaneous monolayer curvature would be favorably accommodated by the cis leaflet, but disfavored by the trans leaflet (Fig. 7). In contrast, for the positive parallel membrane curvature, agents that promote negative spontaneous monolayer curvature would be disfavored by the cis leaflet, but would be well situated in the trans monolayer. Clearly, the preferences would be exactly opposite for a molecule that promotes positive spontaneous monolayer curvature.
The observed t o distributions are consistent with a meridian curvature that is energetically more important for stabilization of the small pore. If the negative meridian curvature is dominant, it follows that the energy of bending into the shape of a small fusion pore would be less when R18 or DiI were enriched in the cis monolayer, whereas the energy would be greater when they accumulated in the trans monolayer. A lowering of bending energy would stabilize the pore because a larger energy would have to be surmounted for the pore to close (Fig. 7, right). This would account for the longer open times observed when these probes were enriched in the cis monolayer. Similarly, the raised energy of a small pore when probes were enriched in the trans monolayer would account for the observed shorter t o. When probe is symmetrically distributed among the two monolayers or is absent, clearly the energy of bending should be intermediate between the two asymmetric cases. The open times should, accordingly, fall in between the open times for probe enriched in cis or trans monolayers; this is as observed. If the destabilization of small pores by R18 and DiI in trans leaflets is due to curvature effects, increasing the positive spontaneous monolayer curvature of trans leaflets should stabilize these pores. This too was experimentally observed: LPC placed in trans leaflets virtually abolished pore flickering (Table I), showing that the pore was stabilized against closures. Thus, the outcomes of the experiments all confirm that the energy required to bend a membrane into a toroidal fusion pore is affected more by accommodating the negative meridian curvature than the positive parallel one.
The Effects of Probe on Pore Closure and Full Enlargement Suggest that Flickering and Successful Pores Form from a Common Small Pore Structure
The energy necessary for a small open pore, O, in an energy well to either surmount a barrier against closure, C, or against full opening, F (the energy profile of Fig. 7, right), can be described by the kinetic scheme C ↔ O → F. The rate constant k c for pore closure, O → C, is equal to 1/t o. The rate constant for full pore enlargement, k F, is less clear. There are two principal possibilities. It may be that as soon as a pore forms, it is destined to either flicker shut or to fully open. If this is the case, k F = 1/t S. Because t S distributions have been experimentally shown to be independent of the location of probe (Table I), the energy difference between the well depth and barrier height would not be affected by probe. But as the location of probe did affect the likelihood of enlargement, this interpretation runs counter to experiment.
In the alternative possibility, flickering and successful pores would be fundamentally the same, with probabilities of either closing or fully opening. There is prior evidence that this is the case: the conductance trajectory of a pore at times early after opening is independent of whether the pore flickers closed or fully opens (Melikyan et al., 1995b ). If flickering and successful pores are initially the same, k F = 1/(Σt o + t S). With R18 or DiI preferentially in the cis leaflet, k F would be smallest; when preferentially placed in the trans leaflet, k F would be largest; when probes are symmetrically placed, the value of k F would be intermediate (Fig. 6). When R18 or DiI were preferentially located in the cis leaflet, successful pores were observed in only half the experiments, in accord with small values of k F. On the other hand, as Σt o + t S was not altered (data not shown), although t S was lengthened (Table I), the LPC trans results did not support the view that flickering and successful pores were initially the same. The reason may be that the O → F description of full pore enlargement, qualitatively accounted for by Σt o + t S, is oversimplified. LPC is an agent often used to promote positive curvature (Chernomordik et al., 1995b ); it probably causes a greater increase in positive spontaneous curvature than the negative increases caused by R18 or DiI. If so, LPC within the trans leaflet could have caused the relative absence of flickering by lengthening open times to the point that pores rarely closed. Pore conductance tends to continually increase if closure does not occur and abrupt pore enlargement becomes more probable as pore conductance increases (Melikyan et al., 1995b ). Thus, one pore that is continually open should be more likely to exhibit full pore enlargement in a given time than if the open times are spread among several pores that close. This would account for the fact that the Σt o + t S did not increase when LPC was placed in the trans leaflet even if flickering and successful pores were initially the same. Alternatively, it is possible that the large changes in membrane tension caused by LPC could have affected pore properties. We do not yet know whether spontaneous curvature has an effect on full pore enlargement; theory predicts that it should (Nanavati et al., 1992; Chizmadzhev et al., 1995), but our observations are not in complete accord with this theory. Specifically, the effects of R18, DiI, and LPC upon full pore enlargement cannot be accounted for solely by spontaneous bilayer curvature; properties other than spontaneous bilayer curvature must be involved.
In summary, the role of spontaneous monolayer and bilayer curvature in membrane fusion is becoming increasingly apparent. Not only are they factors in merger of contacting leaflets and formation of small pores, it is now clear that spontaneous bilayer curvature also regulates the flickering of small pores. On balance, the evidence to date supports the interpretation that flickering and successful pores are initially the same, with their eventual outcome stochastically determined and that closure of pores is strongly regulated by spontaneous curvatures. If flickering and successful pores are initially indistinguishable, they evolve out of a common small pore. The small pore quickly, perhaps even at formation, assumes a toroidal shape.
Acknowledgments
We thank Sofya Brenner for excellent technical assistance and Dr. Yuri Chizmadzhev for critical comments upon an earlier version of the manuscript.
Supported by National Institutes of Health grants GM-27367 (F.S. Cohen) and GM-54787 (G.B. Melikyan) and the Medical Research Council of Canada grant MA-7654 (R.M. Epand).
Abbreviations used in this paper
- HA
hemagglutinin of influenza virus
- HII
inverted hexagonal
- LPC
lysophosphatidylcholine
- rho-PE
rhodamine-phosphatidylethanolamine
references
- Alvarez de Toledo G, Fernandez-Chacon R, Fernandez JM. Release of secretory products during transient vesicle fusion. Nature. 1993;363:554–558. doi: 10.1038/363554a0. [DOI] [PubMed] [Google Scholar]
- Bagai S, Lamb RA. Truncation of the COOH-terminal region of the paramixovirus SV5 fusion protein leads to hemifusion but not complete fusion. J Cell Biol. 1996;135:73–84. doi: 10.1083/jcb.135.1.73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blumenthal R, Sarkar DP, Durell S, Howard DE, Morris SJ. Dilation of the influenza hemagglutinin fusion pore revealed by the kinetics of individual cell–cell fusion events. J Cell Biol. 1996;135:63–71. doi: 10.1083/jcb.135.1.63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bröring K, Haest CWM, Deuticke B. Translocation of oleic acid across the erythrocyte membrane. Evidence for a fast process. Biochim Biophys Acta. 1989;986:321–331. doi: 10.1016/0005-2736(89)90484-7. [DOI] [PubMed] [Google Scholar]
- Chernomordik LV, Kozlov MM, Zimmerberg J. Lipids in biological membrane fusion. J Membr Biol. 1995a;146:1–14. doi: 10.1007/BF00232676. [DOI] [PubMed] [Google Scholar]
- Chernomordik LV, Chanturia A, Green J, Zimmerberg J. The hemifusion intermediate and its conversion to complete fusion: regulation by membrane composition. Biophys J. 1995b;69:922–929. doi: 10.1016/S0006-3495(95)79966-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chernomordik LV, Kozlov MM, Melikyan GB, Abidor IG, Markin VS, Yu, Chizmadzhev A. The shape of lipid molecules and monolayer membrane fusion. Biochim Biophys Acta. 1985;812:643–655. [Google Scholar]
- Chernomordik LV, Leikina E, Frolov V, Bronk P, Zimmerberg J. An early stage of membrane fusion mediated by the low pH conformation of influenza hemagglutinin depends upon membrane lipids. J Cell Biol. 1997;136:81–93. doi: 10.1083/jcb.136.1.81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chernomordik LV, Frolov V, Leikina E, Bronk P, Zimmerberg J. The pathway of membrane fusion catalyzed by influenza hemagglutinin: restriction of lipids, hemifusion, and lipidic fusion pore formation. J Cell Biol. 1998;140:1369–1382. doi: 10.1083/jcb.140.6.1369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ChizmadzhevYu.A., F.S. Cohen, A. Shcherbakov, and J. Zimmerberg. Membrane mechanics can account for fusion pore dilation in stages. Biophys J. 1995;69:2489–2500. doi: 10.1016/S0006-3495(95)80119-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohen, F.S., and G.B. Melikyan. 1998. Methodologies in the study of cell-cell fusion. Methods (Orlando). In press. [DOI] [PubMed]
- Cleverley DZ, Lenard J. The transmembrane domain in viral fusion: essential role for a conserved glycine residue in vesicular stomatitis virus G protein. Proc Natl Acad Sci USA. 1998;95:3425–3430. doi: 10.1073/pnas.95.7.3425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Devaux PF. Lipid transmembrane asymmetry and flip-flop in biological membranes and in lipid bilayers. Curr Opin Struct Biol. 1993;3:489–494. [Google Scholar]
- Döbereiner H-G, Evans E, Kraus M, Seifert U, Wortis M. Mapping vesicle shapes into the phase diagram: a comparison of experiment and theory. Phys Rev E. 1997;55:4458–4474. [Google Scholar]
- Doxsey SJ, Sambrook J, Helenius A, White JM. An efficient method for introducing macromolecules into living cells. J Cell Biol. 1985;101:19–27. doi: 10.1083/jcb.101.1.19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Epand RM. Diacylglycerols, lysolecithin, or hydrocarbons markedly alter the bilayer to hexagonal phase transition temperature of phosphatidylethanolamines. Biochemistry. 1985;24:7092–7095. doi: 10.1021/bi00346a011. [DOI] [PubMed] [Google Scholar]
- Epand RM, Epand RF, Ahmed N, Chen R. Promotion of hexagonal phase formation and lipid mixing by fatty acids with varying degrees of unsaturation. Chem Phys Lipids. 1991;57:75–80. doi: 10.1016/0009-3084(91)90051-c. [DOI] [PubMed] [Google Scholar]
- Fernandez JM, Neher E, Gomperts BD. Capacitance measurements reveal stepwise fusion events in degranulating mast cells. Nature. 1984;312:453–455. doi: 10.1038/312453a0. [DOI] [PubMed] [Google Scholar]
- Ganong BR, Bell RM. Transmembrane movement of phosphatidylglycerol and diacylglycerol sulfhydryl analogues. Biochemistry. 1984;23:4977–4983. doi: 10.1021/bi00316a023. [DOI] [PubMed] [Google Scholar]
- Helfrich W. Elastic properties of lipid bilayers: theory and possible experiments. Z Naturforsch Sect C Biosci. 1973;28:693–703. doi: 10.1515/znc-1973-11-1209. [DOI] [PubMed] [Google Scholar]
- Hernandez LD, Hoffman LR, Wolfberg TG, White JM. Virus–cell and cell–cell fusion. Annu Rev Cell Dev Biol. 1996;12:627–661. doi: 10.1146/annurev.cellbio.12.1.627. [DOI] [PubMed] [Google Scholar]
- Kemble GW, Danieli T, White JM. Lipid-anchored influenza hemagglutinin promotes hemifusion, not complete fusion. Cell. 1994;76:383–391. doi: 10.1016/0092-8674(94)90344-1. [DOI] [PubMed] [Google Scholar]
- Kielian M. Membrane fusion and the alphavirus life cycle. Adv Vir Res. 1995;45:113–151. doi: 10.1016/s0065-3527(08)60059-7. [DOI] [PubMed] [Google Scholar]
- Kozlov MM, Leikin SL, Chernomordik LV, Markin VS, Yu, Chizmadzhev A. Stalk mechanism of membrane fusion. Eur Biophys J. 1989;17:121–129. doi: 10.1007/BF00254765. [DOI] [PubMed] [Google Scholar]
- Kozlov MM, Leikin SL, Rand RP. Bending, hydration and interstitial energies quantitatively account for the hexagonal-lamellar-hexagonal reentrant phase transition in dioleoylphosphatidylethanolamine. Biophys J. 1994;67:1603–1611. doi: 10.1016/S0006-3495(94)80633-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar VV, Malewicz B, Baumann WJ. Lysophosphatidylcholine stabilizes small unilamellar phosphatidylcholine vesicles. Phosphorus-31 NMR evidence for the wedge effect. Biophys J. 1989;55:789–792. doi: 10.1016/S0006-3495(89)82877-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lollike K, Borregaard N, Lindau M. The exocytotic fusion pore of small granules has a conductance similar to an ion channel. J Cell Biol. 1995;129:99–104. doi: 10.1083/jcb.129.1.99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melikyan GB, Niles WD, Cohen FS. The fusion kinetics of influenza hemagglutinin expressing cells to planar bilayer membranes is affected by HA density and host cell surface. J Gen Physiol. 1995a;106:783–802. doi: 10.1085/jgp.106.5.783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melikyan GB, Niles WD, Ratinov V, Karhanek M, Zimmerberg J, Cohen FS. Comparison of transient and successful fusion pores connecting influenza hemagglutinin expressing cells to planar membranes. J Gen Physiol. 1995b;106:803–819. doi: 10.1085/jgp.106.5.803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melikyan GB, White JM, Cohen FS. GPI-anchored influenza hemagglutinin induces hemifusion to both red blood cell and planar bilayer membranes. J Cell Biol. 1995c;131:679–691. doi: 10.1083/jcb.131.3.679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melikyan GB, Deriy BN, Ok DC, Cohen FS. Voltage-dependent translocation of R18 and DiI across lipid bilayers leads to fluorescence changes. Biophys J. 1996;71:2680–2691. doi: 10.1016/S0006-3495(96)79459-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melikyan GB, Brener SA, Ok DC, Cohen FS. Inner but not outer membrane leaflets control the transition from glycosylphosphatidylinositol-anchored influenza hemagglutinin-induced hemifusion to full fusion. J Cell Biol. 1997a;136:995–1005. doi: 10.1083/jcb.136.5.995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melikyan GB, Jin H, Lamb RA, Cohen FS. The role of the cytoplasmic tail region of influenza virus hemagglutinin in formation and growth of fusion pores. Virology. 1997b;235:118–128. doi: 10.1006/viro.1997.8686. [DOI] [PubMed] [Google Scholar]
- Mohandas N, Wyatt J, Mel SF, Rossi ME, Shohet SB. Lipid translocation across the human erythrocyte membrane. Regulatory factors. J Biol Chem. 1982;257:6537–6543. [PubMed] [Google Scholar]
- Nanavati C, Markin VS, Oberhauser AF, Fernandez JM. The exocytotic fusion pore modeled as a lipidic pore. Biophys J. 1992;63:1118–1132. doi: 10.1016/S0006-3495(92)81679-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nüssler F, Clague MJ, Herrmann A. Meta-stability of the hemifusion intermediate induced by glycosylphosphatidylinositol-anchored influenza hemagglutinin. Biophys J. 1997;73:2280–2291. doi: 10.1016/S0006-3495(97)78260-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ratinov V, Plonsky I, Zimmerberg J. Fusion pore conductance: experimental approaches and theoretical algorithms. Biophys J. 1998;74:2374–2387. doi: 10.1016/S0006-3495(98)77946-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sambrook J, Rodgers L, White JM, Gething M-J. Lines of BPV-transformed murine cells that constitutively express influenza virus hemagglutinin. EMBO (Eur Mol Biol Organ) J. 1985;4:91–103. doi: 10.1002/j.1460-2075.1985.tb02322.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sheetz MP, Singer SJ. Biological membranes as bilayer couples. A molecular mechanism of drug-erythrocyte interactions. Proc Natl Acad Sci USA. 1974;71:4457–4461. doi: 10.1073/pnas.71.11.4457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spruce AE, Iwata A, Almers W. The first milliseconds of the pore formed by a fusogenic viral envelope protein during membrane fusion. Proc Natl Acad Sci USA. 1991;88:3623–3627. doi: 10.1073/pnas.88.9.3623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steck, T.L. 1989. Red cell shape. In Cell Shape: Determinants, Regulation, and Regulatory Role. W.D. Stein, and F. Bronner, editors. Academic Press, Inc., San Diego, CA. 205–246.
- Struik, D.J. 1961. Lectures on Classical Differential Geometry. 2nd ed. Addison-Wesley Publishing. Co., Reading, MA. 60–61.
- Suzuki Y, Nagao Y, Kato H, Matsumoto M, Nerome K, Nakajima K, Nobusawa E. Human influenza A virus hemagglutinin distinguishes sialyloligosaccharides in membrane-associated gangliosides as its receptor which mediates the adsorption and fusion process of virus infection. J Biol Chem. 1986;261:17057–17061. [PubMed] [Google Scholar]
- Tilcock CPS, Cullis PR. Lipid polymorphism. Ann NY Acad Sci. 1987;492:88–102. doi: 10.1111/j.1749-6632.1987.tb48657.x. [DOI] [PubMed] [Google Scholar]
- White JM. Membrane fusion. Science. 1992;258:917–924. doi: 10.1126/science.1439803. [DOI] [PubMed] [Google Scholar]
- Zhou Z, Misler S, Chow RH. Rapid fluctuations in transmitter release from single vesicles in bovine adrenal chromaffin cells. Biophys J. 1996;70:1543–1552. doi: 10.1016/S0006-3495(96)79718-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zimmerberg J, Blumenthal R, Sarkar DP, Curran M, Morris SJ. Restricted movement of lipid and aqueous dyes through pores formed by influenza hemagglutinin during cell fusion. J Cell Biol. 1994;127:1885–1894. doi: 10.1083/jcb.127.6.1885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zimmerberg J, Curran M, Cohen FS, Brodwick M. Simultaneous electrical and optical measurements show that membrane fusion precedes secretory granule swelling during exocytosis of beige mouse mast cells. Proc Natl Acad Sci USA. 1987;84:1585–1589. doi: 10.1073/pnas.84.6.1585. [DOI] [PMC free article] [PubMed] [Google Scholar]