

Abstract
The renin-angiotensin system (RAS) participates in the compensation of major blood pressure disturbances such as haemorrhage and is involved in the tonic long-term (> 1 day) maintenance of mean arterial blood pressure (MABP). Since its contribution to the short-term (< 1 h) buffering of normal blood pressure variability is not known, this was investigated in resting conscious dogs.
The regulatory efficiency and the response time of the RAS were studied by an acute step reduction of renal artery pressure to 70 mmHg for 1 h using a suprarenal aortic cuff. After a delay of at least 100 s, MABP rose exponentially by 22 ± 5 mmHg in normal dogs (n = 4), by 6 ± 3 mmHg after angiotensin converting enzyme (ACE) inhibition (n = 4), and by 25 ± 5 mmHg after ganglionic blockade (n = 4). MABP returned to control after release of the cuff with similar time courses. The time constants of the MABP responses were in the range of 20 min. Thus, possible feedback oscillations of the RAS would be expected around 0.0025 Hz (1/(4 × 100 s)); a buffering effect would be possible below this frequency.
Blood pressure variability was investigated by spectral analysis of MABP from 3.75 h recordings in the frequency ranges of 0.002–0.003 Hz (feedback oscillations) and below 0.002 Hz (buffering effect).
ACE inhibition (n = 7) decreased MABP by 11 ± 2 mmHg (P < 0.05), but in both frequency ranges integrated spectral density was not affected. ACE inhibition also failed to significantly change spectral density in either of the two frequency ranges under the following conditions: (1) during ganglionic blockade (n = 7), (2) during a low-sodium diet (except for a very slight elevation below 0.002 Hz) (n = 7), and (3) when the fall of MABP induced by ACE inhibition was compensated by an angiotensin II infusion (n = 7).
It is concluded that in spite of its high regulatory efficiency with an adequate response time the RAS does not directly contribute to the short-term buffering of blood pressure variability, nor does it give rise to feedback oscillations under normal resting conditions. Even if the RAS is stimulated by sodium restriction its contribution to short-term blood pressure buffering is only marginal.
The contribution of the renin-angiotensin system (RAS) to the maintenance of mean arterial blood pressure (MABP) becomes apparent when it is strongly stimulated by sodium deprivation (Johnson & Davis, 1973; Fray et al. 1977; Kopelman et al. 1983) or during pathophysiological conditions which provoke major circulatory disturbances such as haemorrhage (Brough et al. 1975; Kopelman et al. 1983). More recently, a continuous tonic influence, as evidenced by a fall in MABP after angiotensin converting enzyme (ACE) inhibition (MacGregor et al. 1981; Kopelman et al. 1983; Ehmke et al. 1987; Hasser & Bishop, 1988; Persson et al. 1993), has also been observed under physiological conditions. This tonic effect of the RAS on the level of MABP appears to be controlled, at least in part, by the pressure-dependent mechanism of renin release, for the following reasons. (1) The levels of long-term MABP and threshold pressure for pressure-dependent renin release are closely related in dogs (Finke et al. 1983; Farhi et al. 1983; Ehmke et al. 1987) and in humans (Fiorentini et al. 1981; Hohenbleicher et al. 1997). (2) In conscious dogs, the level of arterial plasma renin activity (PRA) was found to be strongly dependent on the actual difference between MABP and threshold pressure, with higher PRA values being present when MABP was closer to the threshold pressure (Ehmke et al. 1987). (3) When MABP was artificially reduced to below the threshold pressure by inducing a constant pressure reduction between the suprarenal aortic and the renal arterial blood pressure (RABP) in conscious dogs, both aortic and renal arterial pressure rose within 2 h until RABP was almost completely restored (Persson et al. 1993). Therefore, the pressure-dependent mechanism of renin release might very well participate in the buffering of the spontaneous blood pressure variability.
Studies in humans (Dutrey-Dupagne et al. 1991), dogs (Akselrod et al. 1985; Brown et al. 1991) and rats (Cerutti et al. 1991; Grichois et al. 1992; Ponchon & Elghozi, 1996) have investigated the influence of the RAS on blood pressure variability in various frequency ranges extending down to 0.008 Hz (2 min cycle length) (Brown et al. 1991). However, previous studies on dogs suggest a response time of the RAS in the range of 5 min for the first effects (Gutmann et al. 1973) and up to 60 min to approach the maximum (Cowley et al. 1971; Gutmann et al. 1973; Liard et al. 1974; Fray et al. 1977). Therefore, a direct regulatory impact of the RAS would at most be expected for fluctuations occurring with a repetition time slower than this, i.e. in a frequency range of below 0.003 Hz (1 per 5 min). Thus, the report that activation of the RAS by sodium restriction failed to alter blood pressure variability down to 0.00003 Hz (8.5 h) in dogs (Shimada & Marsh, 1979) seems to be the only information about possible buffering effects of the RAS available to date. In addition, due to its response characteristics comprising a delay and a dynamic response, the RAS may also give rise to feedback oscillations which would increase blood pressure variability (Madwed et al. 1989; Wagner et al. 1997). Such an effect may arise when the cycle length of the fluctuation is in the range 4 times longer than the delay (Madwed et al. 1989; Wagner et al. 1997). In a previous study on dogs, we had observed the appearance of slow oscillations of MABP at 0.01 Hz, when the buffering effect of the baroreceptor reflex had been suppressed by ganglionic blockade (Wagner et al. 1997). These oscillations may have been derived from feedback oscillations of the RAS.
The aim of the present study therefore was 2-fold. First, an attempt was made to identify the frequency ranges in which a direct regulatory influence or feedback oscillations of the RAS are to be expected. Second, the role of the RAS in the exaggeration or the buffering of MABP variability was investigated in these ranges. In order to determine the frequency ranges, the time course of the pressure response during a step reduction of RABP was studied. Blood pressure variability was then measured within the identified frequency ranges using spectral analysis both before and after blockade of the RAS. To exclude a possible interaction with the buffering action of the baroreceptor reflex, the experiments were repeated after ganglionic blockade. Furthermore, since the regulatory capacity of pressure-dependent renin release is known to be enhanced after sodium restriction (Fray et al. 1977; Farhi et al. 1983), experiments were also done in dogs on a low-sodium diet. Finally, the possibility that an increase in blood pressure variability might be blunted by a vasodilatation (Jacob et al. 1991; Julien et al. 1993) induced by ACE inhibition was investigated by experiments in which the decrease in MABP was compensated by a concurrent infusion of angiotensin II (Ang II).
METHODS
All data were derived from sixty-eight experiments on twenty conscious, chronically instrumented foxhounds (24–43 kg body weight, 15–24 months of age, either sex). The dogs were kept under an artificial light-dark cycle (06.00–18.00 h light) and were held on a standard dog diet (SSNIFF Spezialdiäten GmbH, Soest, Germany). All experiments and procedures were done in accordance with the national law for the care and use of research animals (State license No. 37–9185.81/121/93, Regierungspräsidium Karlsruhe, Germany).
Surgical procedures
The dogs were surgically prepared under sterile conditions. After premedication with atropine (0.5 mg s.c.; Braun, Melsungen, Germany) and propionylpromazine (0.64 mg kg−1s.c.; Combelen, Bayer, Leverkusen, Germany) anaesthesia was induced by pentobarbitone sodium (20 mg kg−1i.v.; Nembutal, Sanofi, Libourne Cedex, France) and maintained by halothane (0.8–1.0 %; Fluothane, ZENECA, Planckstadt, Germany) and N2O (0.5 l min−1). Depth of anaesthesia was checked by repeated testing of the eyelid reflex. At the induction, masseter relaxation was also assessed.
Through a left flank incision the left renal artery and the abdominal aorta were exposed retroperitoneally. A polyurethane catheter was implanted into the abdominal aorta below the renal arteries. Most of the dogs also received a silicone catheter implanted into the left renal vein to facilitate later drug infusions. Four dogs further received an inflatable cuff, placed around the aorta above the origins of the renal arteries as well as a second catheter which was implanted proximal to the cuff. The catheter(s) were subcutaneously led to the animal's neck, where they were exteriorized. At least 10 days were allowed before the first experiments were done. During the first 9 days, the dogs received a combination of benzylpenicillin and sulfatolamide (3 ml s.c. every third day; Tardomycel, Bayer). The catheter(s) were flushed every second or third day and filled with a solution of heparin (1700 i.u. ml−1) and cephtazidim (16 mg ml−1; Fortum, Glaxo, Bad Oldesloh, Germany) in 0.9 % saline.
Measurements
All experiments were done between 07.30 and 14.00 h, while the dogs rested on their right side, as they had been trained to do beforehand. MABP was measured by a pressure transducer (Statham P23Db or P23XL, Gould, Valley View, OH, USA) with a calibrated amplifier (Gould Pressure Processor) connected to the catheter. All data were continuously recorded on a computer (80286, or 80386 plus Labtech Note Book v 7.11 (Spectra GmbH, Germany)) at a sampling rate of 20 Hz, after A/D conversion (DAS-16, Keithley-Metrabyte, Taunton, MA, USA). After all experiments on a respective dog had been done the animal was killed by an overdose of pentobarbitone.
Experimental protocols
Regulatory capacity of the RAS
In order to define the temporal characteristics of the regulatory effect of the RAS, the blood pressure response to a sudden reduction of the RABP was investigated. In the four dogs with the implanted aortic cuff arterial blood pressures proximal (MABP) and distal (RABP) to the cuff were measured continuously for a duration of 240 min. After a control period of 60 min, RABP was reduced to 70 mmHg by a controlled inflation of the cuff using an electropneumatic control device. Sixty minutes later, the cuff was released. Blood samples were drawn from the proximal aortic catheter at 57, 117, 177 and 237 min after the start of the experiment. PRA (expressed as nanograms angiotensin I (Ang I) per millilitre per hour) was determined by radioimmunoassay.
In each of the four dogs, this protocol was followed under three different conditions on separate days in random order. At least one additional day of recovery was allowed after ACE inhibition or ganglionic blockade. The dogs were on a normal-sodium diet.
Control
The experiments were done under control conditions.
ACE inhibition
In order to test whether the pressure response was effectively blocked by ACE inhibition, the experiment was done after ramipril (Hoechst, Frankfurt a.M., Germany) had been given (2 mg kg−1 per os 60 min and 1 mg kg−1 10 min before the experiment was started).
Ganglionic blockade
In order to eliminate the influence of the baroreceptor reflex, the ganglionic blocking agent hexamethonium bromide (Sigma-Aldrich, Deisenhofen, Germany) was given as a bolus of 5 mg kg−1i.v., immediately followed by a continuous infusion of 5 mg kg−1 h−1i.v. The control period was begun 20 min after the start of the infusion.
Blood pressure variability
Arterial blood pressure was recorded continuously for 3.75 h (13 500 s) under four different conditions (see below) in paired experiments with and without ACE inhibition. Experiments on the same dog were done on separate days, the order of the experimental conditions being randomly assigned. In those dogs in which experiments for more than one of the experimental groups, including the low-sodium group, were done, the order of the experiments for the different groups was also randomly assigned. At least 1 day was allowed for recovery after ACE inhibition or ganglionic blockade.
Control
(n = 7). Experiments were done in dogs on a normal-sodium diet.
Ganglionic blockade
(n = 7). The experiments were done during infusion of hexamethonium bromide (5 mg kg−1 bolus i.v. followed by infusion of 5 mg kg−1 h−1i.v.). The experimental recording was begun 20 min after the start of the hexamethonium infusion.
Low-sodium diet
(n = 7). The normal food was replaced by a diet preparation with a restricted-sodium content (Limited Ingredients Diets, canine; IVD Natures Recipe Pet Foods, Corona, CA, USA; 850 g day−1). This was started at least 9 days prior to the experiments. To accelerate sodium depletion bolus injections of 40 mg furosemide (frusemide) i.v. (LasixR, Hoechst AG, Frankfurt, Germany) were given on the first 2 days of the diet. Sodium intake was less than 30 mequiv day−1 under this diet, while it was about 100 mequiv day−1 with the normal food. In six of the dogs, blood samples were taken during the normal- and low-sodium diet periods for determination of PRA. At least 10 days of normal-sodium intake were allowed for recovery from the low-sodium diet.
Ang II infusion
(n = 7). In this group the administration of ramipril (see above) was combined with an exogenous substitution of Ang II at a constant rate (5 ng kg−1 min−1i.v.; Olsen et al. 1985). This infusion was begun 30 min after the first dose of ramipril (i.e. 20 min before the second dose, 30 min before starting the recording). The experiments without Ang II were done under the same conditions as those in the control group.
Data handling
Regulatory capacity of the RAS
The 20 Hz data were converted into consecutive block-averages over 100 s (Fig. 1) or over 10 s (Fig. 2). Heart rate (HR) was determined off-line from the pulse intervals (PIs) detected in the 20 Hz blood pressure signal by a computer program (written by A. Just). The increase in systemic MABP during the RABP reduction was calculated from the difference between MABP during the first 200 s and the last 200 s of the 60 min period of reduced RABP for each dog. The compensatory gain (open-loop gain) of the RAS was estimated for each dog from this pressure response divided by the difference between RABP from the 60 min period prior to RABP reduction and RABP from the 60 min period during RABP reduction. The pressure decrease after release of the cuff was calculated from the difference between MABP during the 200 s immediately after release and during the last 200 s 120 min after release.
Figure 1. Pressure responses to a reduction of renal artery pressure.

Mean arterial blood pressure (MABP) and mean renal artery blood pressure (MRABP) before, during and after a servo-controlled reduction of RABP to 70 mmHg for 60 min. Values are means ± s.e.m. from experiments in 4 conscious dogs during control conditions (A), after ACE inhibition (B) and after ganglionic blockade (C). Large filled circles, MABP; small filled circles, MRABP.
Figure 2. Pressure response (from Fig. 1) after ganglionic blockade at higher temporal resolution.
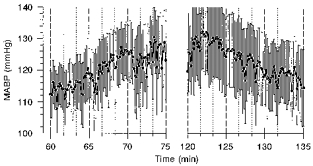
The first 15 min of the rise of MABP during reduction of RABP (left) and the first 15 min of the fall of MABP after release of the cuff (right) are shown in 4 dogs during ganglionic blockade. •, mean values averaged over 10 s (± s.e.m.). Vertical lines denote periods of 100 s.
To assess the time constants of the pressure responses, both the increase and the decrease in MABP were normalized for each dog to the respective change in MABP. The time constants were estimated from the time at which 63 % (1 - e−1) of the respective change had been completed. No time constants were determined for the responses during ACE inhibition.
Blood pressure variability
Spectral analysis
The 20 Hz data were digitally low-pass filtered (cut-off (frequency) = 1.6 Hz; recurrent finite-impulse-response type, order 100) and then resampled at 2.5 Hz. Although this would produce aliasing above 0.9 Hz, it allowed us to use a small ratio of cut-off to sampling frequency and kept the inaccuracies in the vicinity of the cut-off frequency far outside of the frequency range of interest. Power density spectra were calculated from these 2.5 Hz data by the Blackman-Tukey algorithm (Bendat & Piersol, 1986). In order to expand the spectral resolution as far as possible into the low frequency range, a single spectrum was calculated from all 215 values of these data, corresponding to 13 107 s, yielding a frequency resolution of 0.000076 Hz. The lowest two frequencies were discarded due to their limited accuracy, so that the smallest frequency investigated was 0.000229 Hz (cycle length, 73 min). Because MABP substantially increased over time during the Ang II infusion, in this group the spectral density was calculated after the trend (linear regression) had been subtracted. In order to estimate the total bood pressure variability in the frequency range deduced from the experiments with the RABP reduction, in all groups spectral density was integrated in two frequency ranges by the trapezoidal rule: at 0.002–0.003 Hz in order to investigate possible feedback oscillations of the RAS, as well as at 0.0002–0.002 Hz in order to test its buffering effect. To quantify the effect of the ACE inhibition on the blood pressure variability in these frequency ranges, the ratio of the respective integrated spectral density values of the experiment with ramipril divided by those without ramipril was calculated for each dog, in decibels (10 log(ratio)).
Time domain
Since the non-stationarities of the blood pressure fluctuations may limit the validity of the spectral analysis in the low frequency range, the total blood pressure variability below 0.002 Hz was also estimated in the time domain. For this purpose the 2.5 Hz data were further low-pass filtered (cut-off 0.25 Hz; recurrent finite-impulse-response type, order 100) and then resampled at 0.5 Hz (6514 values). Further smoothing was done by a sliding average over 250 of these values, i.e. corresponding to a period of 500 s or 0.002 Hz, and moving by 1 value each, thus keeping the sampling rate of 0.5 Hz. From the resulting 6265 values, the standard deviation was calculated for each recording. As described for the spectral densities above, for the group with Ang II infusion the standard deviation was calculated after trend correction. The change in variability was expressed in decibels as the ratio of the standard deviation with ACE inhibition divided by that without it (20 log(ratio)). The factor of 20 takes into account that the standard deviation directly reflects the amplitude, while the spectral density (factor 10 used, see above) corresponds to the squared amplitude of the fluctuations.
Test of ganglionic blockade
In order to test the effectiveness of the ganglionic blockade (see above), the responses of MABP and PI to bolus injections of nitrotriglycerate (0.5 or 1.0 μg i.v.; Nitrolingual, Pohl-Boskamp, Hohenlockstedt, Germany) and of methoxamine (24 μg kg−1i.v.; Sigma-Aldrich) were investigated in three dogs. The MABP response was determined from the average value of MABP during the first 3 min after each bolus compared with the MABP during the 60 s immediately before each bolus. This was done to obtain an estimate of the regulatory efficiency of the baroreceptor reflex. The period of 3 min was chosen because, in contrast to the baroreceptor reflex (Just et al. 1995), the RAS is not able to contribute to the control of MABP within this time span (see Results of the present study). Thus it was possible to estimate the regulation independent of the RAS without the need to block the RAS. The PI response was determined from the maximum change in PI during the first 60 s after the bolus compared with the mean PI during the 60 s before the bolus. This was done to estimate the maximum regulatory effort of the baroreceptor reflex. Arterial blood pressure was recorded continuously at 20 Hz for 3.75 h. PI was determined off-line from the pressure data. The nitrotriglycerate bolus was given at 30 min and again at 3.5 h after the start of the experiment. In both cases methoxamine was given 5–7 min after the nitrotriglycerate.
Under control conditions, MABP fell by 4 ± 2 mmHg and PI decreased by 573 ± 77 ms after the nitrotriglycerate. MABP rose by 4 ± 1 mmHg and PI increased by 1184 ± 444 ms in response to methoxamine. The MABP responses were enhanced to −25 ± 5 mmHg (nitrotriglycerate) and +36 ± 6 mmHg (methoxamine) after 20 min of infusion of hexamethonium. After 3.5 h infusion of hexamethonium the MABP responses were −31 ± 4 mmHg (nitrotriglycerate) and +39 ± 4 mmHg (methoxamine). The PI responses were greatly attenuated to −66 ± 21 and +27 ± 8 ms at the beginning, and to −52 ± 1 and +14 ± 2 ms at the end of the infusion of hexamethonium. Thus, the dose of hexamethonium was sufficient to substantially attenuate the baroreceptor reflex-mediated regulation of MABP.
Statistical analysis
Differences between paired experiments in the same dogs were tested by Student's t test for paired samples. For the analysis of the temporal changes of HR and PRA during RABP reduction, one-way ANOVA was used in conjunction with Student-Newman- Keuls test. For the comparison of resting PRA during normal- and low-sodium diet the Mann-Whitney U test was used. In all cases, a P value of 0.05 was considered significant. All values are expressed as means ±s.e.m.
RESULTS
Regulatory capacity of the RAS
Under control conditions, MABP was 98 ± 4 mmHg during the 60 min control period and increased significantly within the following 60 min, when RABP was reduced to 70 mmHg (Fig. 1A). In all dogs the increase started after a latency of at least 200 s. The increase in MABP from the first 200 s to the last 200 s of the period of reduced RABP was 22 ± 5 mmHg (P < 0.05), corresponding to an open-loop gain of −0.90 ± 0.21. Sixty-three per cent of this response was completed after about 32 min. After release of the cuff, MABP decreased in an approximately exponential manner by 18 ± 1 mmHg (P < 0.05), after a delay of about 200 s. The time constant for the falling phase of MABP was about 18 min. PRA averaged 1.5 ng Ang I ml−1 h−1 during the control period, increased to a 4-fold higher level during the pressure reduction, and was restored 1 h after release of the cuff (Table 1). HR did not change over time and was on average 66 ± 4 beats min−1.
Table 1.
Arterial plasma renin activity (PRA) before, during and after the reduction of renal artery pressure
| PRA (ng Ang I ml−1 h−1) | ||||
|---|---|---|---|---|
| Group | 1 h | 2 h | 3 h | 4 h |
| Control (n = 4) | 1.5 ± 0.3 | 6.0 ± 1.8* | 1.8 ± 0.5 | 1.5 ± 0.4 |
| ACE inhibition (n = 4) | 5.3 ± 2.6 | 19.9 ± 3.9* | 3.9 ± 0.8 | 5.2 ± 1.7 |
| Ganglionic blockade (n = 4) | 1.0 ± 0.2 | 7.9 ± 3.3 | 1.4 ± 0.1 | 1.1 ± 0.2 |
PRA as measured at the end of the first (1 h), second (2 h), third (3 h) and fourth (4 h) hour of the experiment. Data aremeans ± s.e.m.;n, number of dogs.
P < 0.05, significantly different from 1 h.
After ACE inhibition, the increase in MABP during RABP reduction was abolished (+6 ± 3 mmHg; Fig. 1B). PRA increased 3.8-fold during the reduction of RABP, and was restored within 1 h after release of the cuff (Table 1). HR remained unchanged at 80 ± 8 beats min−1.
After ganglionic blockade, MABP was 103 ± 7 mmHg during the control period. After inflation of the aortic cuff, it increased immediately by 8 ± 1 mmHg (P < 0.05). MABP then rose in an exponential manner by an additional 25 ± 5 mmHg (P < 0.05; Fig. 1C), corresponding to an open-loop gain of −0.76 ± 0.12. As can be seen from Fig. 2, which shows the first 15 min of this response at a 10 times higher temporal resolution, this increase did not start before a delay of at least 100 s. The time constant for the rise was approximately 21 min. After release of the cuff, MABP fell immediately by 11 ± 2 mmHg (P < 0.05), remained there for about 200 s (Figs 1C and 2), and then fell exponentially by an additional 18 ± 4 mmHg (P < 0.05; Fig. 1C). The time constant for the falling phase of pressure was about 22 min. PRA rose almost 8-fold during reduced RABP (although this was not statistically significant due to the great interindividual variation), and was restored within 1 h after release of the cuff (Table 1). HR remained unchanged at 127 ± 7 beats min−1.
From these temporal characteristics, the frequency ranges in which feedback oscillations and in which a direct regulatory influence of the RAS are to be expected in the conscious dog were deduced. Feedback oscillations may arise for fluctuations with a cycle length about 4 times longer than the delay (Madwed et al. 1989; Wagner et al. 1997), i.e. around 0.0025 Hz, and was thus assessed between 0.002 and 0.003 Hz. A regulatory effect can only be expected below this frequency and was thus investigated between 0.0002 and 0.002 Hz.
Blood pressure variability
The physiological blood pressure spectrum under control conditions in the control group (Fig. 3A, open circles) shows a typical 1/f distribution with the prominent representation of lower frequencies (Fig. 3, note logarithmic scales). ACE inhibition lowered MABP by 11 ± 2 mmHg (P < 0.05) in the control group, while HR remained unchanged (Table 2). The mean spectrum of blood pressure variability below 0.04 Hz was not overtly altered by ACE inhibition (Fig. 3A). Therefore, as a more sensitive marker, the relative change of spectral density was calculated (in decibels) for each individual dog (Fig. 4A). Again, no increase in blood pressure variability was detected. Integrated spectral density in the range 0.002–0.003 Hz (feedback oscillations) was not significantly altered by ACE inhibition (Table 3). In the frequency range below 0.002 Hz (buffering) a slight, non-significant reduction of variability was found after ACE inhibition in the integrated spectral density as well as in the standard deviation of MABP (Fig. 5).
Figure 3. Effect of ACE inhibition on mean spectra of blood pressure variability.
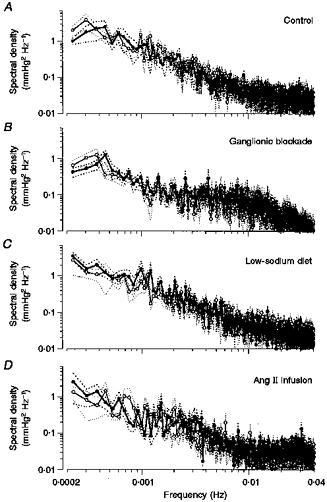
The effect of ACE inhibition on mean spectral density of blood pressure between 0.0002 and 0.04 Hz is shown from paired experiments under control conditions (n = 7; A), ganglionic blockade (n = 7; B), low-sodium diet (n = 7; C) and Ang II infusion (n = 7; D). ○, control; •, ACE inhibition; dotted lines, s.e.m.
Table 2.
Haemodynamic data during the 3.75 h recordings
| MABP (mmHg) | HR (beats min−1) | |||
|---|---|---|---|---|
| Group | Control | ACE inhibition | Control | ACE inhibition |
| Control (n = 7) | 103 ± 4 | 92 ± 4* | 77 ± 3 | 83 ± 8 |
| Ganglionic blockade (n = 7) | 100 ± 3 | 93 ± 3* | 126 ± 3 | 122 ± 2 |
| Low-sodium diet (n = 7) | 97 ± 8 | 86 ± 4* | 68 ± 5 | 73 ± 5* |
| Ang II infusion (n = 7) | 97 ± 3 | 114 ± 3* | 69 ± 3 | 68 ± 3 |
Mean arterial blood pressure (MABP) and heart rate (HR) before and after ACE inhibition for the experiments under normal conditions (Control), and after ganglionic blockade, low-sodium diet and Ang II infusion. Data are means ±s.e.m.;n, number of dogs.
P < 0.05, significantly different from control.
Figure 4. Changes of spectral density induced by ACE inhibition.
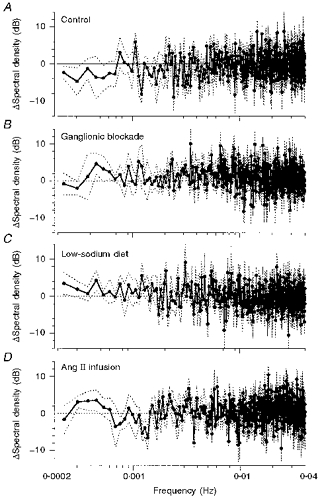
Relative changes (in dB) of spectral density induced by ACE inhibition under control conditions (n = 7; A), ganglionic blockade (n = 7; B) low-sodium diet (n = 7; C) and Ang II infusion (n = 7; D). •, mean change; dotted lines, s.e.m.
Table 3.
Changes of integrated spectral density of blood pressure between 0.002 and 0.003 Hz induced by ACE inhibition
| Group | ΔSpectral density (dB) |
|---|---|
| Control (n = 7) | −0.11 ± 1.47 |
| Ganglionic blockade (n = 7) | +1.60 ± 0.96 |
| Low-sodium diet (n = 7) | +0.29 ± 1.20 |
| Ang II infusion (n = 7) | +0.03 ± 0.77 |
Relative changes of spectral density induced by ACE inhibition. Means ±s.e.m. for experiments under normal conditions (Control), and after ganglionic blockade, low-sodium diet and Ang II infusion. n, number of dogs. None of the changes was significantly different from 0 dB.
Figure 5. Changes of integrated spectral density and standard deviation of blood pressure below 0.002 Hz induced by ACE inhibition.
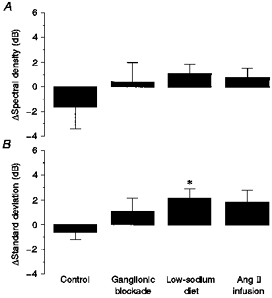
The columns depict average changes induced by ACE inhibition under control conditions (n = 7) ganglionic blockade (n = 7), low-sodium diet (n = 7) and Ang II infusion (n = 7). A, changes in spectral density of blood pressure integrated between 0.0002 and 0.002 Hz. B, changes in the standard deviation of blood pressure calculated from consecutive 8.3 min mean values of MABP. Data are presented as means ± s.e.m. *P < 0.05.
Ganglionic blockade increased HR, but did not change MABP (Table 2). Baseline variability of MABP in the total range below 0.04 Hz was slightly, but not significantly, smaller (1.70 ± 0.42 mmHg2) compared with the fourteen pooled control experiments of the control and Ang II infusion groups (2.61 ± 0.45 mmHg2). ACE inhibition after ganglionic blockade induced a fall in MABP by 7 ± 2 mmHg (P < 0.05) without changing HR (Table 2). The mean spectrum of arterial blood pressure was not altered by ACE inhibition (Fig. 3B). The paired comparison of the single spectra (Fig. 4B) also failed to reveal any change in blood pressure variability. Integrated spectral densities also neither decreased at 0.002–0.003 Hz (Table 3), nor increased at 0.0002–0.002 Hz after ACE inhibition (Fig. 5A). The same was true for the standard deviation (Fig. 5B).
In accordance with a previous observation (Wagner et al. 1997), distinct oscillations of MABP appeared sporadically with a frequency around 0.01 Hz after hexamethonium infusion. Spectral density at 0.005–0.015 Hz in relation to the total power below 0.04 Hz was significantly higher (0.222 ± 0.033) than in the pooled controls (0.132 ± 0.010). However, the spectral density in this frequency range was not significantly affected by the additional ACE inhibition (+1.29 ± 0.58 dB; not significant).
In sodium-restricted dogs, MABP and HR were not significantly different from the values in normal dogs (Table 2). PRA was elevated from 1.6 ± 0.1 to 4.5 ± 1.3 ng Ang I ml−1 h−1 (P < 0.05) by the low-sodium diet. ACE inhibition lowered MABP by 11 ± 2 mmHg (P < 0.05) and slightly elevated HR (Table 2). In the paired comparison of the spectra (Fig. 4C), a slight but consistent increase in spectral density was detected below about 0.002 Hz. The integrated spectral density (P = 0.21) and the standard deviation (P < 0.05) in this frequency range were slightly elevated (Fig. 5). There was no significant change of spectral density at 0.002–0.003 Hz (Table 3).
Infusion of Ang II during ACE inhibition immediately increased MABP to 104 ± 5 mmHg (mean over the first 20 min of infusion, data not shown; P < 0.05vs. mean level of control experiments). During the following 2 h MABP gradually further rose and then remained at a steady-state level of 116 ± 3 mmHg. Over the entire recording period MABP was 17 ± 1 mmHg higher than in the control experiments (Table 2). HR did not change (Table 2). The combination of ACE inhibition with Ang II infusion did not cause any obvious change in the mean spectrum of arterial blood pressure (Fig. 3D) or in the paired comparison of the spectra (Fig. 4D). The slight elevation in the integrated spectral density below 0.002 Hz and in the standard deviation did not reach statistical significance (Fig. 5). There was also no significant change of integrated spectral density at 0.002–0.003 Hz (Table 3).
DISCUSSION
Regulatory capacity of the RAS
The RABP reduction induced a substantial increase in systemic MABP of more than 20 mmHg, which demonstrates that the RAS is capable of effectively compensating for such a disturbance. The fact that this response was abolished after ACE inhibition in spite of even higher levels of PRA indicates that the observed rise in MABP was indeed induced by an activation of the RAS. Therefore, the observed time courses are representative of the regulatory activity of the RAS.
The time course of both the rising phase of MABP during RABP reduction (on-response), as well as its falling phase after release of the cuff (off-response of the RAS) followed an approximately exponential pattern both under control conditions and after ganglionic blockade. The on-response appeared somewhat slower under control conditions than without a functional baroreceptor reflex. This corresponds to other reports (Liard et al. 1974) and can most easily be explained by a compensatory influence which is brought about by other control systems, but which decreases in strength over time. The most probable cause for such an effect may derive from a pressure-dependent or Ang II-induced resetting of the baroreceptor reflex (Liard et al. 1974). Thus, the observed time course of the on-response in baroreflex intact dogs most probably reflects the dynamics of the gradually fading regulatory influence of the baroreceptor reflex due to the process of resetting, rather than the time course of the activation of the RAS itself. Accordingly, the temporal characteristics of the RAS are probably more reliably reflected by the response observed under ganglionic blockade. Under these conditions, both the on- and the off-response of the RAS followed an exponential pattern with an initial delay of at least 100 s and time constants in the range of 20 min. Within the 60 min period of stimulation the response appeared to be completed. Similar time courses in response to a RABP reduction without the interference of the baroreceptor reflex have also been found by others (Cowley et al. 1971; Liard et al. 1974), with indeed no further changes in MABP for up to 6 h (Liard et al. 1974). Since both the onset of renin release (Gutmann et al. 1973; Nafz et al. 1997) as well as the conversion of angiotensin I (Ng & Vane, 1968) are known to be completed within 2–4 min, the rate limiting step is most probably the accumulation of renin in the plasma. Although the delay cannot be determined exactly from the present data, it seems safe to conclude that 100 s is the minimum time which passes before the RAS can exert any substantial impact on MABP, after its activation or deactivation.
From these temporal characteristics the following conclusions can be drawn concerning the frequency range in which the RAS would be able to exert an impact on blood pressure variability. (1) The estimated delay of at least 100 s excludes any impact on fluctuations occurring with a repetition time faster than this, and hence confines this frequency range below a maximum upper limit of 0.01 Hz. (2) The strong regulatory effect and the symmetric exponential response with a delay indicate that the RAS might give rise to feedback oscillations. From the delay of at least 100 s, such an effect would be expected around 0.0025 Hz (1/(4 × 100 s)). (3) A buffering effect on MABP fluctuations can only become effective at lower frequencies (< 0.001 Hz).
Blood pressure variability
Feedback oscillations
During ACE inhibition, no decrease in the blood pressure variability was observed in the frequency range 0.002–0.003 Hz. This was also true when a possible influence of the baroreceptor reflex had been suppressed by ganglionic blockade, or when the sensitivity of the RAS had been increased by sodium restriction. This indicates that the RAS does not contribute to the blood pressure variability in this frequency range.
In a previous study (Wagner et al. 1997), we had observed the appearance of distinct oscillations of MABP in dogs after ganglionic blockade at a frequency of about 0.01 Hz. The same observation was made in the present investigation. Although the almost sinusoidal rhythmicity of these oscillations suggests some kind of feedback oscillation, the temporal characteristics of the RAS appear to be too slow to serve as the underlying mechanism. Indeed the oscillations were not affected by ACE inhibition. Thus, the nature of these MABP oscillations still remains obscure.
Blood pressure buffering effect
In the mean spectra of arterial blood pressure, no differences in blood pressure variability were observed with and without ACE inhibition in any of the experimental groups. However, due to the large interindividual variation a possible change induced by blockade of the RAS might have been blurred in the mean spectra. Therefore, the relative changes of spectral density were also calculated for each individual dog (Fig. 4). Furthermore, due to the non-stationarities of the pressure fluctuations (Marsh et al. 1990), the validity of the spectral analysis at these low frequencies may be limited. For this reason, the variability in the range below 0.002 Hz was also estimated in the time domain from the standard deviation of MABP. However, in the control group, neither of these measures revealed any elevation of blood pressure variability in the frequency range below 0.002 Hz after blockade of the RAS.
With regard to the strong regulatory capability of the RAS in response to a constant reduction of the RABP (Cowley et al. 1971; Gutmann et al. 1973) and in view of the well-documented regulatory function under closed-loop conditions (Persson et al. 1993), this lack of a regulatory influence under physiological conditions seems surprising. It is even more remarkable, since MABP was consistently lowered by about 10 mmHg after ACE inhibition, indicating that the RAS contributed significantly to basal vascular tone in all groups.
Since a potential increase in blood pressure variability after blockade of the RAS might have been absorbed by the buffering action of the baroreceptor reflex, the experiments were repeated after ganglionic blockade. Because some evidence in the literature suggests that pressure-dependent activation of the RAS may critically depend on resting sympathetic tone (Hopf et al. 1994), it was important to test that the ganglionic blockade did not at the same time blunt the function of the RAS. The strong increase in MABP in response to the reduction in RABP (Fig. 1C) demonstrates that in the present study the response of the RAS was still functional after ganglionic blockade. Nevertheless, under these conditions also, ACE inhibition did not induce any increase of blood pressure variability.
After chronic sodium deprivation the contribution of the RAS to the tonic maintenance of MABP is known to be enhanced (Kopelman et al. 1983), the pressure-dependent activation of renin release is stronger (Farhi et al. 1983), and the regulatory function in response to RABP reductions is exaggerated (Fray et al. 1977). Accordingly, the contribution of the RAS to blood pressure buffering might be more important and may only become detectable when the RAS is activated. Indeed, even though the integrated spectral density did not show any significant increase of variability with ACE inhibition, in the time domain a significant elevation of blood pressure variability was detected in the sodium-restricted dogs (Fig. 5). This indicates that during low-sodium intake the RAS may have a small regulatory role in blood pressure buffering. However, this effect is only marginal compared with the large influence of the baroreceptor reflex. The change of standard deviation by 2.2 dB indicates that blood pressure variability was at most 30 % higher after blockade of the RAS in the sodium-restricted dogs. In contrast, in a previous study on dogs of the same strain (Just et al. 1995) the spectral density of MABP between 0.0002 and 0.1 Hz had been observed to be elevated more than 6-fold and the standard deviation of MAPB almost 3-fold after denervation of the baroreceptors, indicating that in this case the variability increased by at least 150 %.
Studies in rats have shown that the amount of blood pressure variability is promoted by a higher vascular tone (Jacob et al. 1991; Julien et al. 1993). Although in dogs this does not seem to be the case after ganglionic blockade (Just et al. 1995), such an effect may still play a role after ACE inhibition. It is conceivable that a potential increase in blood pressure fluctuations after blockade of the RAS had been ablated by a loss of variability due to the concomitant vasodilatation. However, when Ang II was infused at a constant rate after ACE inhibition, the blood pressure variability remained virtually unchanged, even though the MABP was even higher than under control conditions. This was the case whether the variability was calculated in the frequency or the time domain.
Conclusions
The present study has confirmed the previous finding (Cowley et al. 1971; Gutmann et al. 1973; Persson et al. 1993) that the RAS provides a strong regulatory capacity in response to a reduction of the RABP, which is rapid enough to exert most of this effect within 1 h. Furthermore, the present study shows that the response follows a fairly symmetric exponential pattern with a delay of at least 100 s and a time constant in the range of 20 min.
Although these characteristics provide the potential for feedback oscillations around 0.0025 Hz, the RAS does not give rise to such an exaggeration of blood pressure variability under normal resting conditions. The RAS is also not involved in the MABP oscillations around 0.01 Hz which can occur after ganglionic blockade.
Furthermore, the temporal characteristics delineate the frequency range in which a direct regulatory influence can be achieved by the RAS, to the range below about 0.002 Hz. However, in spite of the substantial regulatory capability and irrespective of the obvious tonic influence which it exerts on the mean pressure, the RAS does not directly contribute to the short-term buffering of the normal blood pressure variability occurring in this frequency range in the conscious resting dog. Even after chronic sodium restriction the direct regulatory influence of the RAS in this frequency range is only marginal.
Although the present study was confined to resting conditions, the conclusions drawn here seem to apply also to those situations including orthostatic challenges, since at least under normal-sodium conditions the pressure response to upright posture is not significantly altered by blockade of the RAS (Sancho et al. 1976; Reinhart & Lohmeier, 1996). By contrast, the observation that the day-to-day variation of MABP was closely related to the same changes of the threshold pressure for renin release (Ehmke et al. 1987), suggests that the RAS may rather promote active fluctuations of arterial blood pressure. It is conceivable for example, that the transmission of fluctuations, which are transferred to the circulation by the autonomic nervous system (Just et al. 1995), is supported by the RAS via a sympathetically mediated shift of the threshold pressure (Ehmke et al. 1998).
Acknowledgments
We gratefully acknowledge the expert technical assistance of I. Keller, L. Mahl, E. Röbel and A. Klein. We thank Professor U. Hilgenfeld, Pharmakologisches Institut, University of Heidelberg for the determination of plasma renin activity. We thank Hoechst Marion Roussel Inc., Frankfurt a.M., for donating ramipril. Programs for digital filtering and Fourier analysis were written by Diplom-Informatiker der Medizin U. Wittmann. This study was supported by the German Research Foundation (SFB 320, A/01).
References
- Akselrod S, Gordon D, Madwed JB, Snidman NC, Shannon DC, Cohen RJ. Hemodynamic regulation: investigation by spectral analysis. American Journal of Physiology. 1985;249:H867–875. doi: 10.1152/ajpheart.1985.249.4.H867. [DOI] [PubMed] [Google Scholar]
- Bendat JS, Piersol AG. Random Data Analysis and Measurement Procedures. New York: Wiley; 1986. [Google Scholar]
- Brough RB, Cowley AWJ, Guyton AC. Quantitative analysis of the acute response to hemorrhage of the renin-angiotensin vasoconstrictor feedback loop in areflexic dogs. Cardiovascular Research. 1975;9:722–733. doi: 10.1093/cvr/9.6.722. [DOI] [PubMed] [Google Scholar]
- Brown DR, Yingling JD, Randall DC, Aral HM, Evans JM, Charles JB, Knapp CF, Raisch R, Ott CE. Angiotensin II does not contribute to rapid reflex control of arterial pressure. American Journal of Physiology. 1991;261:R473–477. doi: 10.1152/ajpregu.1991.261.2.R473. [DOI] [PubMed] [Google Scholar]
- Cerutti C, Gustin MP, Paultre CZ, Lo M, Julien C, Vincent M, Sassard J. Autonomic nervous system and cardiovascular variability in rats: a spectral analysis approach. American Journal of Physiology. 1991;261:H1292–1299. doi: 10.1152/ajpheart.1991.261.4.H1292. [DOI] [PubMed] [Google Scholar]
- Cowley AW, Jr, Miller JP, Guyton AC. Open loop analysis of the renin-angiotensin system in the dog. Circulation Research. 1971;28:568–581. [Google Scholar]
- Dutrey-Dupagne C, Girard A, Ulmann A, Elghozi JL. Effects of the converting enzyme inhibitor trandolapril on short-term variability of blood pressure in essential hypertension. Clinical Autonomic Research. 1991;1:303–307. doi: 10.1007/BF01819836. [DOI] [PubMed] [Google Scholar]
- Ehmke H, Gerke B, Just A, Kirchheim HR. Identification of a neuro-renal positive feedback in medium-term blood pressure control. FASEB Journal. 1998;12:A364 (abstract). [Google Scholar]
- Ehmke H, Persson P, Kirchheim H. A physiological role for pressure-dependent renin release in long-term blood pressure control. Pflügers Archiv. 1987;410:450–456. doi: 10.1007/BF00586524. [DOI] [PubMed] [Google Scholar]
- Farhi ER, Cant JR, Barger AC. Alteration of renal baroreceptor by salt intake in control of plasma renin activity in conscious dogs. American Journal of Physiology. 1983;245:F119–122. doi: 10.1152/ajprenal.1983.245.1.F119. [DOI] [PubMed] [Google Scholar]
- Finke R, Gross R, Hackenthal E, Huber J, Kirchheim HR. Threshold pressure for the pressure-dependent renin release in the autoregulating kidney of conscious dogs. Pflügers Archiv. 1983;399:102–110. doi: 10.1007/BF00663904. [DOI] [PubMed] [Google Scholar]
- Fiorentini C, Guazzi MD, Olivari MT, Bartotelli A, Necchi G, Magrini F. Selective reduction of renal perfusion pressure and blood flow in man: humoral and hemodynamic effects. Circulation. 1981;63:973–978. doi: 10.1161/01.cir.63.5.973. [DOI] [PubMed] [Google Scholar]
- Fray JCS, Johnson MD, Barger AC. Renin release and pressor response to renal arterial hypotension: effect of dietary sodium. American Journal of Physiology. 1977;233:H191–195. doi: 10.1152/ajpheart.1977.233.2.H191. [DOI] [PubMed] [Google Scholar]
- Grichois ML, Blanc J, Deckert V, Elghozi JL. Differential effects of enalapril and hydralazine on short-term variability of blood pressure and heart rate in rats. Journal of Cardiovascular Pharmacology. 1992;19:863–869. doi: 10.1097/00005344-199206000-00004. [DOI] [PubMed] [Google Scholar]
- Gutmann FD, Tagawa H, Haber E, Barger AC. Renal arterial pressure, renin secretion, and blood pressure control in trained dogs. American Journal of Physiology. 1973;224:66–72. doi: 10.1152/ajplegacy.1973.224.1.66. [DOI] [PubMed] [Google Scholar]
- Hasser EM, Bishop VS. Neurogenic and humoral factors maintaining arterial pressure in conscious dogs. American Journal of Physiology. 1988;255:R693–698. doi: 10.1152/ajpregu.1988.255.5.R693. [DOI] [PubMed] [Google Scholar]
- Hohenbleicher H, Klostermann F, Schorr U, Seyfert S, Persson PB, Sharma AM. Identification of a renin threshold and its relationship to salt intake in a patient with pure autonomic failure. Hypertension. 1997;30:1068–1071. doi: 10.1161/01.hyp.30.5.1068. [DOI] [PubMed] [Google Scholar]
- Hopf HB, Schlaghecke R, Peters J. Sympathetic neural blockade by thoracic epidural anesthesia suppresses renin release in response to arterial hypotension. Anesthesiology. 1994;80:992–999. doi: 10.1097/00000542-199405000-00007. [DOI] [PubMed] [Google Scholar]
- Jacob HJ, Alper RH, Grosskreutz CL, Lewis SJ, Brody MJ. Vascular tone influences arterial pressure lability after sinoaortic deafferentiation. American Journal of Physiology. 1991;260:R359–367. doi: 10.1152/ajpregu.1991.260.2.R359. [DOI] [PubMed] [Google Scholar]
- Johnson JA, Davis JO. Angiotensin II: important role in the maintenance of arterial blood pressure. Science. 1973;179:906–907. doi: 10.1126/science.179.4076.906. [DOI] [PubMed] [Google Scholar]
- Julien C, Zhang ZQ, Barres C. Role of vasoconstrictor tone in arterial pressure lability after chronic sympathectomy and sinoaortic denervation in rats. Journal of the Autonomic Nervous System. 1993;42:1–10. doi: 10.1016/0165-1838(93)90336-s. [DOI] [PubMed] [Google Scholar]
- Just A, Wagner CD, Ehmke H, Kirchheim HR, Persson PB. On the origin of low-frequency blood pressure variability in the conscious dog. The Journal of Physiology. 1995;489:215–223. doi: 10.1113/jphysiol.1995.sp021043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kopelman RI, Dzau VJ, Shimabukuro S, Barger AC. Compensatory response to hemorrhage in conscious dogs on normal and low salt intake. American Journal of Physiology. 1983;244:H351–356. doi: 10.1152/ajpheart.1983.244.3.H351. [DOI] [PubMed] [Google Scholar]
- Liard JF, Cowley AWJ, McCaa RE, McCaa CS, Guyton AC. Renin, aldosterone, body fluid volumes, and the baroreceptor reflex in the development and reversal of Goldblatt hypertension in conscious dogs. Circulation Research. 1974;34:549–560. doi: 10.1161/01.res.34.4.549. [DOI] [PubMed] [Google Scholar]
- MacGregor GA, Markandu ND, Roulston JE, Jones JC, Morton JJ. Maintenance of blood pressure by the renin-angiotensin-system in normal man. Nature. 1981;291:329–331. doi: 10.1038/291329a0. [DOI] [PubMed] [Google Scholar]
- Madwed JB, Albrecht P, Mark RG, Cohen RJ. Low-frequency oscillations in arterial pressure and heart rate: a simple computer model. American Journal of Physiology. 1989;256:H1573–1579. doi: 10.1152/ajpheart.1989.256.6.H1573. [DOI] [PubMed] [Google Scholar]
- Marsh DJ, Osborn JL, Cowley AWJr. 1/f fluctuations in arterial pressure and regulation of renal blood flow in dogs. American Journal of Physiology. 1990;258:F1394–1400. doi: 10.1152/ajprenal.1990.258.5.F1394. [DOI] [PubMed] [Google Scholar]
- Nafz B, Berthold H, Ehmke H, Hackenthal E, Kirchheim HR, Persson PB. Flow versus pressure in the control of renin release in conscious dogs. American Journal of Physiology. 1997;273:F200–205. doi: 10.1152/ajprenal.1997.273.2.F200. [DOI] [PubMed] [Google Scholar]
- Ng KKF, Vane JR. Fate of angiotensin I in the circulation. Nature. 1968;218:144–150. doi: 10.1038/218144a0. [DOI] [PubMed] [Google Scholar]
- Olsen ME, Hall JE, Montani JP, Guyton AC, Langford HG, Cornell JE. Mechanisms of angiotensin II natriuresis and antinatriuresis. American Journal of Physiology. 1985;249:F299–307. doi: 10.1152/ajprenal.1985.249.2.F299. [DOI] [PubMed] [Google Scholar]
- Persson PB, Ehmke H, Kirchheim HR, Lempinen M, Nafz B. The role of the kidney in canine blood pressure control: direct assessment of the closed-loop gain. The Journal of Physiology. 1993;464:121–130. doi: 10.1113/jphysiol.1993.sp019626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ponchon P, Elghozi JL. Contribution of the renin-angiotensin and kallikrein-kinin systems to short-term variability of blood pressure in two-kidney, one-clip hypertensive rats. European Journal of Pharmacology. 1996;297:61–70. doi: 10.1016/0014-2999(95)00721-0. [DOI] [PubMed] [Google Scholar]
- Reinhart GA, Lohmeier TE. Role of the renin-angiotensin system in mediating the effects of posture on renal function. American Journal of Physiology. 1996;271:R282–288. doi: 10.1152/ajpregu.1996.271.1.R282. [DOI] [PubMed] [Google Scholar]
- Sancho J, Re R, Burton J, Barger AC, Haber E. The role of the renin-angiotensin-aldosterone system in cardiovascular homeostasis in normal human subjects. Circulation. 1976;53:400–405. doi: 10.1161/01.cir.53.3.400. [DOI] [PubMed] [Google Scholar]
- Shimada S, Marsh DJ. Oscillations in mean arterial pressure in conscious dogs. Circulation Research. 1979;44:692–700. doi: 10.1161/01.res.44.5.692. [DOI] [PubMed] [Google Scholar]
- Wagner CD, Just A, Nafz B, Persson PB. Very low frequency oscillations in arterial blood pressure after autonomic blockade in conscious dogs. American Journal of Physiology. 1997;272:R2034–2039. doi: 10.1152/ajpregu.1997.272.6.R2034. [DOI] [PubMed] [Google Scholar]