

Abstract
Tandem constructs are increasingly being used to restrict the composition of recombinant multimeric channels. It is therefore important to assess not only whether such approaches give functional channels, but also whether such channels completely incorporate the subunit tandems. We have addressed this question for neuronal nicotinic acetylcholine receptors, using a channel mutation as a reporter for subunit incorporation. We prepared tandem constructs of nicotinic receptors by linking α (α2–α4, α6) and β (β2, β4) subunits by a short linker of eight glutamine residues. Robust functional expression in oocytes was observed for several tandems (β4_α2, β4_α3, β4_α4, and β2_α4) when coexpressed with the corresponding β monomer subunit. All tandems expressed when injected alone, except for β4_α3, which produced functional channels only together with β4 monomer and was chosen for further characterization. These channels produced from β4_α3 tandem constructs plus β4 monomer were identical with receptors expressed from monomer α3 and β4 constructs in acetylcholine sensitivity and in the number of α and β subunits incorporated in the channel gate. However, separately mutating the β subunit in either the monomer or the tandem revealed that tandem-expressed channels are heterogeneous. Only a proportion of these channels contained as expected two copies of β subunits from the tandem and one from the β monomer construct, whereas the rest incorporated two or three β monomers. Such inaccuracies in concatameric receptor assembly would not have been apparent with a standard functional characterization of the receptor. Extensive validation is needed for tandem-expressed receptors in the nicotinic superfamily.
Keywords: oocytes, two-electrode voltage-clamp, reporter mutation approach, acetylcholine, ion channels
INTRODUCTION
Tandem constructs have been employed in the study of voltage-gated potassium channels (Isacoff et al., 1990), cyclic nucleotide–gated channels (Gordon and Zagotta, 1995; Zimmerman, 2002), P2X (Stoop et al., 1999), GABAA (Im et al., 1995), and NMDA receptors (Schorge and Colquhoun, 2003). To be a useful experimental preparation, “tandem” recombinant channels must express reproducibly and homogeneously, have a defined channel composition, but at the same time maintain normal function.
We set out to use tandem constructs to express neuronal nicotinic acetylcholine receptors (nAChRs), with a view ultimately to obtain efficient expression of complex heteromeric combinations and examined in detail if the assumptions of the technique were valid for the tandem constructs that gave the best expression.
Tandem constructs should be particularly useful for neuronal nAChR, which, in their native form, can contain as many as four different subunits within the same pentamer (Conroy and Berg, 1995; Forsayeth and Kobrin, 1997). Functional recombinant neuronal nAChRs can however be obtained from simpler combinations, homomeric (one α subunit) or heteromeric (combinations of α and β subunits; for review see Colquhoun et al., 2003). Heterologous expression should in principle give good results for homomers, for heteromeric “pair” receptors (formed by a single α-type together with a single β-type subunit), and for heteromeric “triplet” receptors provided the third subunit, α5 or β3, cannot form pair receptors (Ramirez-Latorre et al., 1996; Wang et al., 1996; Groot-Kormelink et al., 1998). However, it is difficult to see how the complex native neuronal nAChRs combinations can be reliably produced in vitro. Even for apparently simple “pair” combinations, such as α4β2, nAChR stoichiometry depends on the expression system (Nelson et al., 2003).
A technique for obtaining receptors with a defined composition would therefore be a major advance. In the simplest strategy used in the nicotinic superfamily, tandem constructs (i.e., two subunits connected by a linker) are expressed with the appropriate monomer to produce a “pure” receptor of known stoichiometry (Im et al., 1995; Baumann et al., 2001, 2002, 2003). It was recently shown (Zhou et al., 2003) that oocyte expression of linked α4 and β2 nicotinic subunits together with either an α or a β monomer construct does produce functional nAChRs.
We found that expressing α3β4 nAChRs from β4_α3 tandems together with β4 monomers produced a heterogeneous population of channels, because of incomplete incorporation of the β subunit from the tandem construct (βtandem). The cartoons in each panel of Fig. 1 show the three possible receptor assemblies, from full incorporation of all the βtandem subunits (left) to partial incorporation (middle) to no incorporation (right). In our experiments (Fig. 1, B–F), expression of a reporter mutation (star) in different subunits detected incomplete tandem incorporation, because of differences in the numbers of mutations in the channel gate (shown by the numbers under the cartoons). Only a proportion of linked subunit receptors fully incorporated the fusion protein (i.e., are assembled as shown in the cartoons on the left of the panels).
Figure 1.
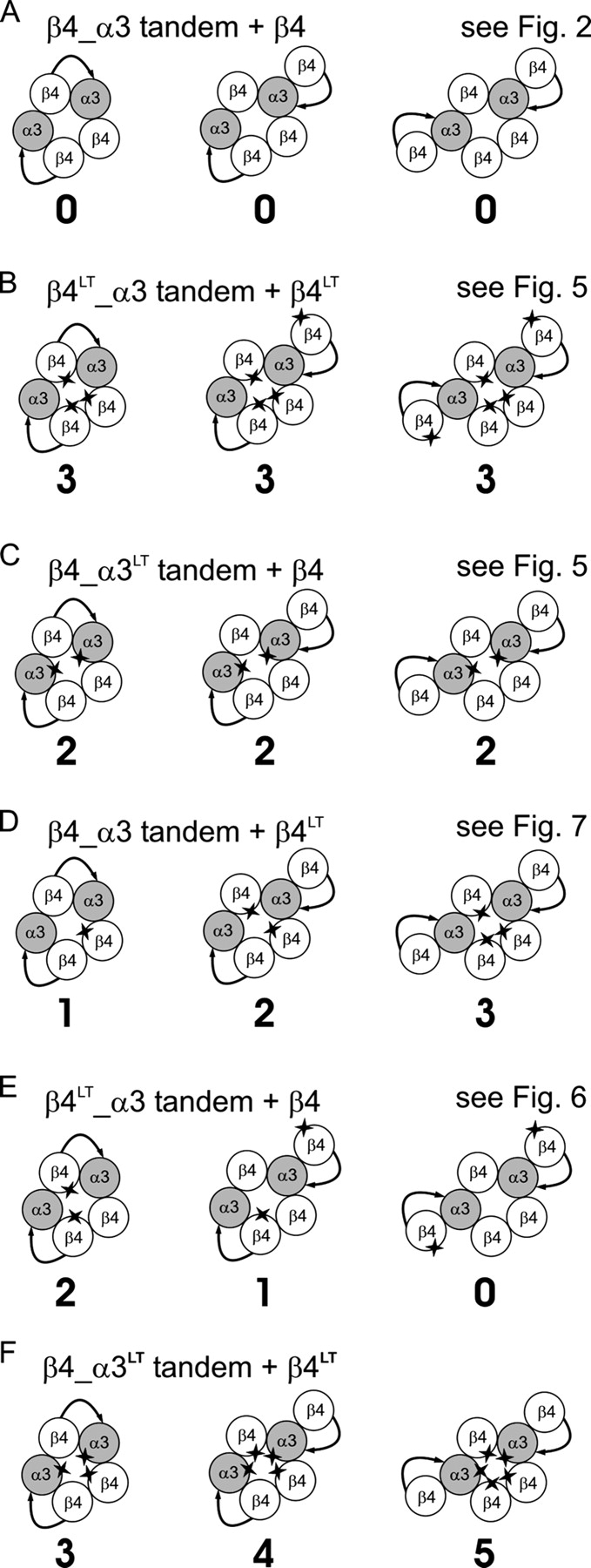
Diagrammatic representation of different nAChR assemblies which may be formed by complete (left) or partial (center and right) incorporation of linked constructs into the receptor. B–F show the effects of introducing a reporter mutation (a Thr in the 9′ position of the second transmembrane domain) in different subunits; references are given to the figures that show the results with each combination. The number under each cartoon shows the number of mutation copies expected to be in the gate for each receptor assembly. Note that no heterogeneity in the number of mutation copies is predicted if all α, all β, or no subunits are mutated (A–C) in accord with the experiments in Figs. 2 and 5. Mutating β only in the monomer construct or only in the tandem construct (D and E) can detect the different receptor forms (see the experiments in Figs. 7 and 6, respectively). If receptors are expressed in which both α and βmonomer (F) bear the mutation, some receptors will bear five copies of the mutation (see results).
MATERIALS AND METHODS
Cloning of Human Neuronal Nicotinic Subunit cDNAs
Previously we described the isolation and sequencing, using PCR-based techniques, of seven (α3, α4, α5, α7, β2, β3, and β4) human neuronal nAChR subunit cDNAs from the neuroblastoma cell lines SH-SY5Y and/or IMR-32 (Groot-Kormelink and Luyten, 1997). Using the same approach we have now also isolated full-length cDNAs for the α2 (from the IMR-32 cell line) and the α6 (from human prostate 5′STRETCH and human brain, thalamus 5′STRETCH PLUS cDNA, CLONTECH Laboratories, Inc.) subunits. Both the α2 and the α6 subunit cDNA coding sequences were identical to the corresponding subunit nucleotide sequences published by Elliott et al. (1996). Nucleotide sequences for all the nine neuronal nicotinic subunits used have been deposited in the Genbank database (EMBL/GenBank/DDBJ accession no. Y16281 for α2, Y08418 for α3, Y08421 for α4, Y08419 for α5, Y16282 for α6, Y08415 for β2, Y08417 for β3, and Y08416 for β4). All subunits, containing only coding sequences and an added Kozak consensus sequence (GCCACC) immediately upstream of the start codon (Groot-Kormelink and Luyten, 1997), were subcloned into the pcDNA3.1 vector (Invitrogen) and the pSP64GL vector. The pSP64GL vector contains 5′ and 3′ untranslated Xenopus β-globin regions (Akopian et al., 1996) to enhance protein expression in oocytes (Liman et al., 1992).
The mutants in 9′ (α3L279T and β4L272T, were L stands for leucine and T for threonine respectively) were created using the QuickChange™ site-directed mutagenesis kit (Stratagene) and their full-length sequence was verified.
Construction of Tandem Subunit cDNAs
All nine tandem constructs (α3_β4, β2_α2, β2_α3, β2_α4, β2_α6, β4_α2, β4_α3, β4_α4, and β4_α6) were made in an identical fashion. First, the coding region of each subunit (α2, α3, α4, α6, β2, β4) was amplified by PCR excluding the stop codon. The PCR primers directed to each corresponding start codon included the Kozak consensus sequence (GCCACC) and the EcoRI enzyme restriction site (5′-end PCR-fragment; CTGAATTC GCCACC ATG…). The primer directed to the coding sequence upstream of the stop codon included the NotI enzyme restriction site (basically the stop codon is replaced by the NotI restriction site). The resulting DNA fragments were purified and subcloned into the pcDNA3.1/Myc-His version C vector (Invitrogen), using the EcoRI and NotI restriction sites.
A linker DNA fragment was created (based on Im et al., 1995) by hybridization of two complementary oligonucleotides; 5′-GGCCGC TCAGCAACAGCAGCAACAGCAGCAAG-3′ and 5′-AATTCTTGCTGCTGTTGCTGCTGTTGCTGA GC-3′. The resulting double-strand DNA linker contains a 5′-end NotI restriction site overhang (underlined) and a 3′-end EcoRI restriction site overhang (underlined), separated by 25 nucleotides (the first nucleotide [bold] is inserted to bring the NotI site [8-cutter] back in the correct reading frame, whereas the next 24 nucleotides code for the eight glutamine amino acids).
The tandem constructs were created using three unique restriction sites; EcoRI (upstream of the start codon of all subunits and the 3′-end of the linker), NotI (downstream of the coding sequence of all subunits and the 5′-end of the linker), and AgeI (between the Myc and His epitope sequences in the pcDNA3.1/Myc-His version C vector). A three way ligation resulted in the following tandem circular plasmid: * [AgeI…His-epitope…stop codon…pcDNA3.1/Myc-His C vector…EcoRI − subunit A − NotI] * [NotI − linker − EcoRI] * [EcoRI − subunit B − NotI…Myc-epitope…AgeI] * (where * represents the ligation sites and [ ] represents purified DNA fragments digested with the restriction sites indicated in bold).
To remove the epitope tags (Myc- and His-) all tandems were subcloned in the corresponding pcDNA3.1 vector, using a unique restriction enzyme site in subunit B. For instance, cutting the tandem upstream of the start codon (of subunit A) and somewhere in subunit B and transferring this fragment in the same position of subunit B, previously cloned in the pcDNA3.1 vector. Finally, all the tandem constructs were also subcloned in the pSP64GL vector. The length of the extracellular linkage of the different tandem constructs differs depending on the extracellular region downstream of TM4 of the first subunit and the length of the signal peptide of the second subunit (Table I).
TABLE I.
Linker Region of Tandems
| Tandem | C′ domain | Linker | Signal peptide | Total |
|---|---|---|---|---|
| α3_β4 | 9 | 15 | 21 | 45 |
| β4_α2 | 19 | 15 | 29 | 63 |
| β4_α3 | 19 | 15 | 26 | 60 |
| β4_α4 | 19 | 15 | 28 | 62 |
| β4_α6 | 19 | 15 | 25 | 59 |
| β2_α2 | 23 | 15 | 25 | 63 |
| β2_α3 | 23 | 15 | 29 | 67 |
| β2_α4 | 23 | 15 | 28 | 66 |
| β2_α6 | 23 | 15 | 25 | 63 |
Linker region from presumed −NH2 end of TM4 region (extracellular region) of the first subunit up to start of the mature second subunit.
The 9′ mutant tandems were created by swapping the corresponding DNA fragments (using unique enzyme restriction sites upstream and downstream of the L9′T mutation) from α3L279T (for β4_α3L279T) and β4L272T (for β4L272T_α3).
The α3_β4, β4_α3, β4_α3L279T, and β4L272T_α3 tandem constructs cloned in the pSP64GL vector were sequenced fully to check for PCR and/or cloning artifacts. All other tandem constructs were sequenced only at the outer ends to check for cloning artifacts.
Mammalian Cell Culture and Transfection
HEK293 cells were obtained from the American Type Culture Collection (ATCC-CRL-1573). Cells were maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with sodium pyruvate (0.11 g/liter), 10% (vol/vol) heat-inactivated fetal bovine serum, 100 U/ml penicillin G, 100 μg/ml streptomycin sulfate, and 2 mM L-glutamine (all from GIBCO BRL) as described previously (Beato et al., 2002). Dishes (10-cm diameter) were plated 24 h before transfection in order to reach around 40% confluence (before transfection). Transfection with LipofectAMINE™ (GIBCO BRL) was performed according to the manufacturer's instructions. Briefly, for each dish 24 μl LipofectAMINE and 6 μg cDNA were added to two separate vials each containing serum-free medium (Optimem; GIBCO BRL) to a final volume of 600 μl. The contents of the LipofectAMINE and the cDNA vials were then mixed and incubated for 30 min at room temperature. Finally, 4.8 ml of Optimem medium was added to the mixture and the whole mixture was added dropwise to one dish of cells (washed with 6 ml Optimem and aspirated) and incubated for 40 h at 37°C and 5% CO2 (transfection mixture was replaced with fresh growth medium after 16 h, see above).
Preparation of In-Vitro Capped mRNA and RNA Gel-Electrophoresis
All cDNA/pSP64GL plasmids were linearized immediately downstream of the 3′ untranslated β-globin sequence, and capped cRNA was transcribed using the SP6 Mmessage Mmachine™ Kit (Ambion) according to the manufacturer's instructions. For RNA electrophoresis a 1.5% agarose gel was prepared using the 1× gel prep/running buffer (NorthernMax-Gly™ system; Ambion). RNA samples (including the 0.24–9.5 Kb RNA ladder; GIBCO BRL) were diluted 1:1 with Glyoxal sample loading dye (Ambion) and incubated at 50°C for 30 min before loading. Samples were separated at 5 V/cm for 3 h and RNAs were visualized by UV transillumination and a photograph taken.
Xenopus Oocyte Preparation
Female Xenopus laevis frogs were anesthetized by immersion in neutralized ethyl m-aminobenzoate solution (tricaine, methanesulphonate salt; 0.2% solution wt/vol; Sigma-Aldrich), and killed by decapitation and destruction of the brain and spinal cord (in accordance with Home Office guidelines) before removal of ovarian lobes. Clumps of stage V-VI oocytes were dissected in a sterile modified Barth's solution of composition (in mM): NaCl 88, KCl 1, MgCl2 0.82, CaCl2 0.77, NaHCO3 2.4, Tris-HCl 15, with 50 U/ml penicillin and 50 μg/ml streptomycin, pH 7.4 adjusted with NaOH. The dissected oocytes were treated with collagenase (type IA; Sigma-Aldrich; 65 min at 18°C, 245 collagen digestion U/ml in Barth's solution, 10–12 oocytes/ml), rinsed, stored at 4°C overnight, and manually defolliculated the following day before cRNA injection.
Xenopus Oocyte cRNA Injection
For Western blotting, oocytes (10 for each sample) were injected with 46 nl water or 500 ng cRNA (in 46 nl of RNase-free water per oocyte). For two-electrode voltage-clamp recording cRNA was injected at a ratio of 1:1 in order to express α3 + β4 pair receptors, and at a molar ratio of 2:1 (equals 4:1 cRNA mass) of tandem versus monomer, respectively, in order to express tandem-containing receptors (all in 46 nl of RNase-free water per oocyte). The total amount of cRNA to be injected for each combination was determined empirically with the aim of achieving a maximum ACh-evoked current of 1–2 μA and was 0.5–4 ng/oocyte, unless otherwise stated (see Fig. 3 and Table II). Oocytes were incubated for ∼60 h at 18°C in Barth's solution containing 5% heat-inactivated horse serum (GIBCO BRL; Quick and Lester, 1994) and then stored at 4°C. Two-electrode voltage-clamp experiments were performed at a room temperature of 18–20°C between 2.5 and 5 d from injection.
Figure 3.

Some tandem constructs form functional channels when expressed alone in Xenopus oocytes. Tandem constructs were expressed with or without monomer construct of the same β subunit they contain (β2 or β4; 2 ng/μl tandem to 0.5 ng/μl monomer; 46 nl injected/oocyte). The four tandem constructs shown here were the only ones to produce functional receptors when expressed with the corresponding β monomer (n = 5–10). Note that three out of these four tandem constructs produced inward currents to 1 mM ACh also when expressed alone (2 ng/μl injected, n = 4–6). Only the β4_α3 tandem construct failed to produce any inward currents (n = 6) when expressed alone.
TABLE II.
β4_α3 Tandem Expressed Together with Monomeric Subunits
| β4_α3 + | I max a | cRNA | n oocyte | n batch |
|---|---|---|---|---|
| nA | ng | |||
| — | 0 ± 0 | 2 | 6 | 2 |
| α2 | 13 ± 7 | 2:0.5 | 6 | 2 |
| α3 | 0 ± 0 | 2:0.5 | 6 | 2 |
| α4 | 0 ± 0 | 2:0.5 | 6 | 2 |
| α5 | 7 ± 4 | 2:0.5 | 5 | 2 |
| α6 | 0 ± 0 | 2:0.5 | 5 | 2 |
| β2 | 5 ± 5 | 2:0.5 | 6 | 2 |
| β3 | 0 ± 0 | 2:0.5 | 6 | 2 |
| β4 | 2,587 ± 702 | 2:0.5 | 10 | 3 |
The maximum current measured for the β4_α3 tandem constructs expressed in Xenopus oocytes alone (—) or in combination with any of the nicotinic monomers that can form heteromeric receptors.
Measured at 1 mM ACh, mean ± SEM.
Preparation of Oocytes and HEK293 Protein Lysates
Whole oocytes were homogenized (10 oocytes each sample) by vigorous pipetting in 500 μl oocyte lysis buffer (100 mM NaCl, 20 mM Tris-HCl, 2% Triton X-100, 1 tablet/5 ml of complete mini protease inhibitor cocktail tablets [Roche], pH 7.6) and rotating at 4°C for 1 to 2 h. To separate yolk and membrane fractions the samples were centrifuged at 20,000 g and 4°C for 30 min. A clear 200-μl supernatant sample was taken avoiding the lipid pellicle (by inserting pipette gently through the pellicle). Proteins were precipitated by adding 5 vol (1 ml) of acetone, incubating on ice for 10 min and centrifugation at 20,000 g for 5 min. Protein pellets were resuspended in 50 μl Laemmli sample buffer (Bio-Rad) containing 5% β-mercapto-ethanol (Bio-Rad Laboratories) and incubated for 1–2 h at RT.
Transfected HEK293 cells were (10-cm dish each sample) washed once with Hanks medium (GIBCO BRL) and dislodged by brief exposure to Trypsin (GIBCO BRL). After adding 1× PBS (GIBCO BRL), cell suspension was centrifuged (10 min, 1,000 rpm) and the pellet washed twice with 1 ml PBS. Pellets were homogenized by vigorous pipetting in 1 ml HEK293 lysis buffer (500 mM NaCl, 50 mM NaH2PO4, 1% Triton X-100, 1% protease inhibitor cocktail for mammalian tissues [Sigma-Aldrich], pH 8.0) and rotating at 4°C for 1–2 h. To pellet cellular debris and DNA the samples were centrifuged at 20,000 g and 4°C for 60 min. A clear 100-μl supernatant sample was taken and proteins precipitated by adding 400 μl methanol, 100 μl chloroform, and 300 μl water, respectively, and centrifugation at 20,000 g for 3 min at 4°C. Subsequently, most of the top layer was removed (protein is at the interface) and 100 μl methanol was added followed by centrifugation at 20,000 g for 5 min at 4°C. Finally, the protein pellets were resuspended in 50 μl Laemmli sample buffer (Bio-Rad Laboratories) containing 5% β-mercapto-ethanol (Bio-Rad Laboratories).
Western Blots
Both HEK293 and oocyte protein samples were incubated at 75°C for 8 min just before loading (20 μl each sample) on a 8% Tris-Glycine polyacrylamide gel containing 2% SDS together with the SeeBlue® prestained protein standard (Invitrogen). After PAGE-SDS the proteins were transferred to nitrocellulose membrane (0.2 μm, protran BA83; Schleicher-Schuell). The blots were probed with rabbit antiserum to α3 or β4 (diluted 1:200 from 5-ml stock solution; Research & Diagnostic antibodies, WR-5611 [α3] and WR-5656 [β4]) followed by HRP-labeled goat anti–rabbit IgG (diluted 1:10,000, 10 μg/ml stock solution from supersignal® west femto chemiluminescence substrate kit; Pierce Chemical Co.). After washing, blots were visualized using the supersignal® west femto chemiluminescence substrate kit (Pierce Chemical Co.) and exposure to biomax light films (Kodak).
Two-electrode Voltage-clamp Recording
Oocytes, held in a 0.2 ml bath, were perfused at 4.5 ml/min with modified Ringer solution (in mM): NaCl 150, KCl 2.8, HEPES 10, MgCl2 2, atropine sulfate 0.5 μM (Sigma-Aldrich), pH 7.2 adjusted with NaOH and voltage clamped at −70 mV, using the two-electrode clamp mode of an Axoclamp-2B amplifier (Axon Instruments, Inc.). Electrodes were pulled from Clark borosilicate glass GC150TF (Warner Instrument Corporation) and filled with 3 M KCl. The electrode resistance was 0.5–1 MΩ on the current-passing side. Experiments were terminated if the total holding current exceeded 2 μA, in order to reduce the effect of series resistance errors. We chose a nominally calcium-free solution in order to minimize the contribution of calcium-gated chloride conductance; this is endogenous to the Xenopus oocyte and may be activated by calcium entry through the neuronal nicotinic channels (Sands et al., 1993).
The agonist solution (acetylcholine chloride [Sigma-Aldrich], freshly prepared from frozen stock aliquots) was applied via the bath perfusion for a period sufficient to obtain a stable plateau response (at low concentrations) or the beginning of a sag after a peak (at the higher concentrations). The resulting inward current was recorded on a flat bed chart recorder (Kipp & Zonen) for later analysis. An interval of 5 min was allowed between ACh applications, as this was found to be sufficient to ensure reproducible responses. A descending dose protocol was used. To compensate for possible decreases in agonist sensitivity during the experiment, a standard concentration of ACh (approximately EC 20 for the particular combination used) was applied every third response for concentration-response curves. The experiment was started only after checking that this standard concentration gave reproducible responses. All the data shown in the study are compensated for the response rundown (Boorman et al., 2000). The average rundown in response to the standard concentrations for the different receptor constructs were: α3 + β4, 40 ± 3%; β4_α3 + β4, 39 ± 11%; β4_α3 + β4LT, 68 ± 5%; β4LT_α3 + β4, 45 ± 4%; β4_α3LT + β4, 57 ± 4% and β4LT_α3 + β4LT, 79 ± 3%.
To reassure ourselves that the lack of functional expression observed for some subunit combinations was true and not a false negative due to oocyte health or expression problems contingent to a given batch, oocyte data were obtained from a minimum of two separate oocyte batches for each combination. Each batch was from a different frog, processed on different experimental weeks: in every experimental batch at least one “control” highly expressing subunit combination was injected to check for expression efficiency.
Curve Fitting
Dose–response curves were fitted with the Hill equation:
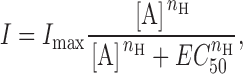 |
(1) |
where I is the response, measured at its peak, [A] is the agonist concentration, I max is the maximum response, EC 50 is the agonist concentration for 50% maximum response, and n H is the Hill coefficient. We used least squares fitting by the program CVFIT, courtesy of D. Colquhoun and I. Vais, available from http://www.ucl.ac.uk/Pharmacology/dc.html.
Fitting was done in stages, as follows. Each dose–response curve was fitted separately, individual responses being equally weighted, in order to obtain estimates for I max, EC 50, and n H (see results). The dose–response curves shown in Figs. 2 and 5–7 were obtained by normalizing each datapoint to the fitted maximum response in that oocyte before pooling and fitting the pooled data with the Hill equation (with weight given by the reciprocal of their variance). Parameter estimates were similar to those obtained by fitting each curve separately.
Figure 2.
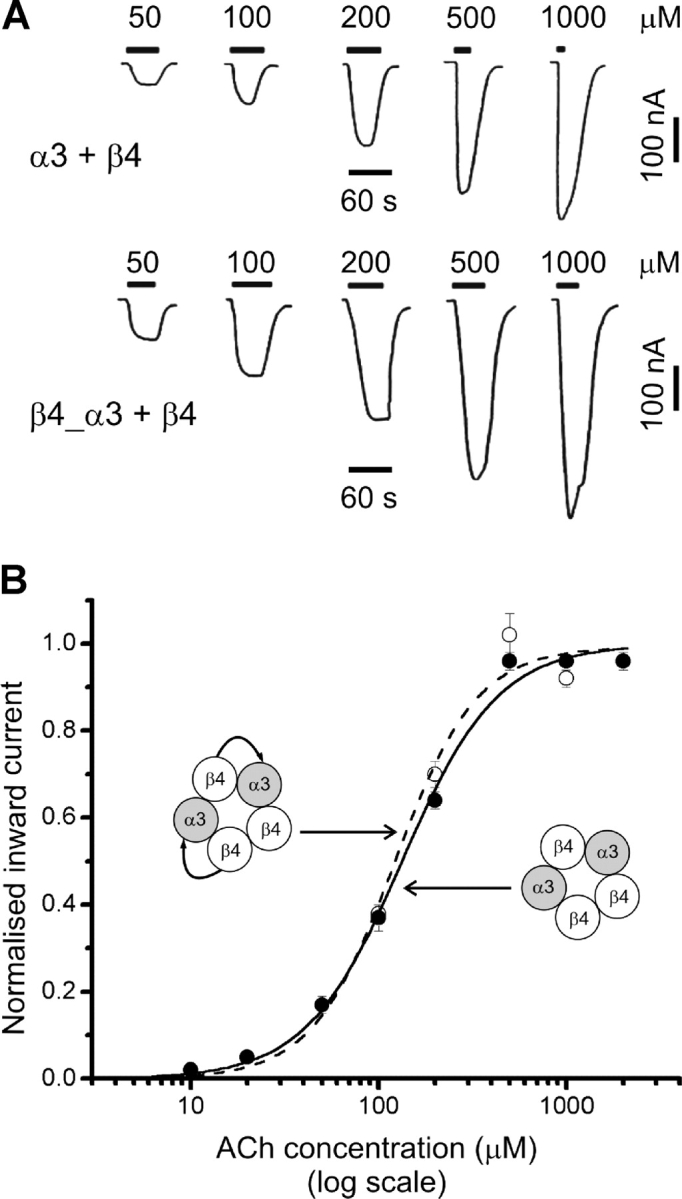
ACh concentration-response curves of α3β4 nAChR expressed in oocytes from monomer or tandem constructs are indistinguishable. (A) Traces are inward currents recorded at a holding potential of −70 mV in response to bath-applied ACh. (B) ACh concentration-response curves from experiments such as the ones shown in A, performed in oocytes injected with either α3 and β4 monomer cRNAs (filled circles, n = 9) or β4_α3 tandem together with β4 monomer cRNAs (open circles, n = 4). Full sets of peak responses to ACh from each oocyte were fitted with the Hill equation and normalised to the fitted maximum response before pooling (see materials and methods). The curves shown are the results of fitting the pooled data.
Figure 5.
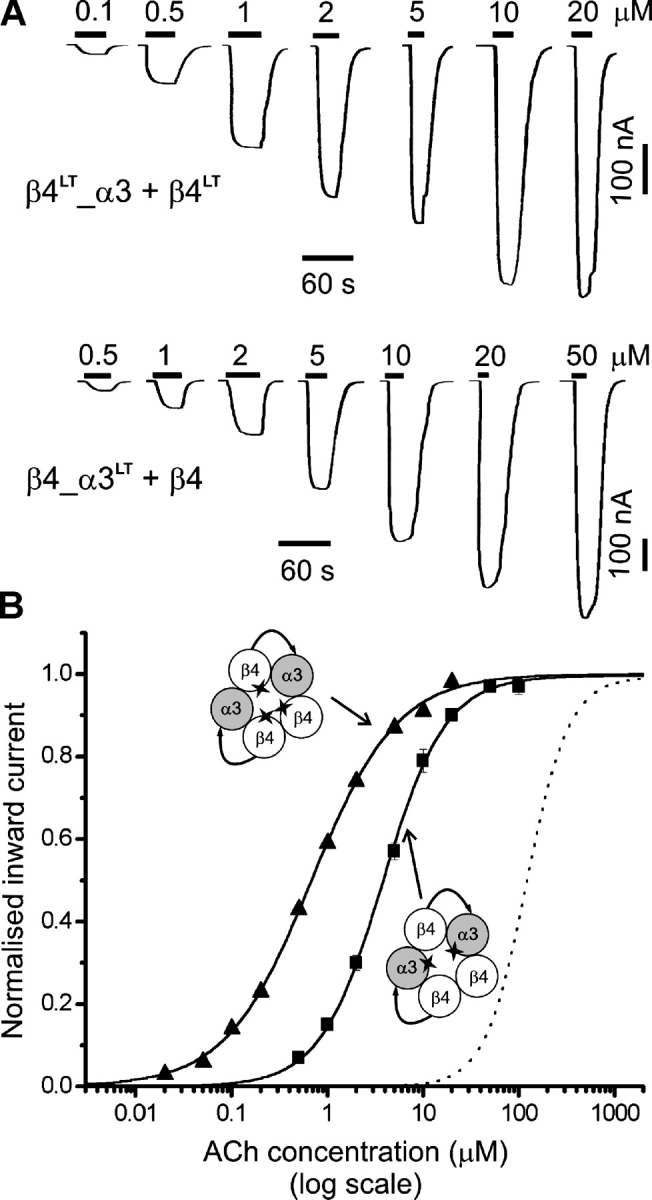
The effects of inserting a L9′T reporter mutation into all the α or all the β subunits in tandem construct receptors. (A) Examples of inward currents elicited by bath-applied ACh in oocytes expressing β4LT_α3 + β4LT (top) or β4_α3LT + β4 (bottom). (B) Concentration-response curves from oocytes injected with β4LT_α3 + β4LT cRNAs (filled triangles, n = 7) or β4_α3LT + β4 cRNAs (filled squares, n = 8). For details of the fitting see materials and methods and the legend to Fig. 2. The concentration-response curve for the β4_α3 + β4 wild-type nAChR (from Fig. 2 B) is shown for reference (dotted line). The EC 50 shifts produced by the mutation are similar to those observed in α3 + β4 receptors and suggest that the channel gate is made up of two α and three β subunits both in linked-subunit and in monomer construct receptors.
Figure 7.
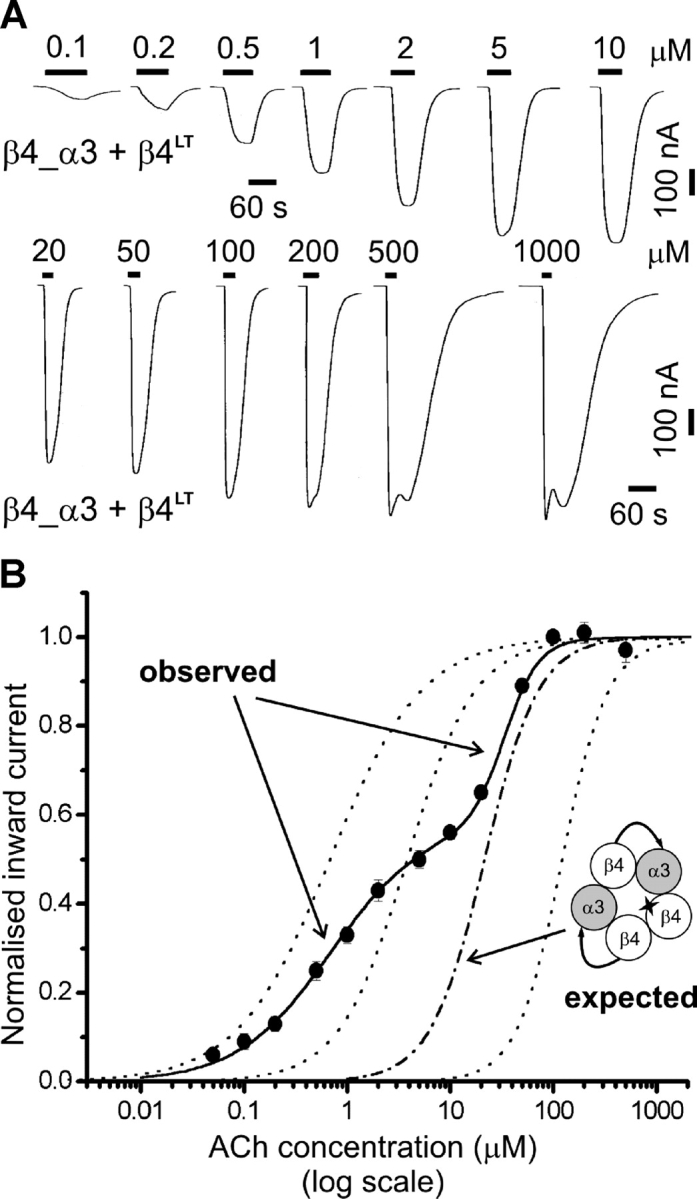
Inserting the L9′T reporter mutation in the β4monomer subunit of linked subunit nAChRs reveals multiple receptor populations. (A) Examples of inward currents elicited by bath-applied ACh in oocytes expressing β4_α3 + β4LT. (B) Concentration-response curve for oocytes injected with β4_α3 + β4LT cRNAs (filled circles, n = 4). Data were normalized and pooled as described in materials and methods and in the legend to Fig. 2. In this case the initial fit was a simultaneous fit of a two-component Hill equation to each dose–response curve, with the constraint of equal EC 50 and Hill slope across oocytes (the proportion of the first component was allowed to vary). The three dotted curves shown for reference are, respectively (right to left), the concentration-response curve for receptors with no mutations (β4_α3 + β4), two mutation copies (β4_α3LT + β4), or three mutation copies (β4LT_α3 + β4LT). The dashed curve shows the concentration-response curve expected for complete tandem incorporation as depicted in the cartoon (i.e., one mutation copy).
When two-components were detected in the concentration-response curve (see Fig. 7), free fits of the individual dose response curves were poorly defined because of the large number of parameters fitted. Good fits were obtained when all the concentration-response curves for this combination were fitted simultaneously with EC 50 and n H values for the two components constrained to be equal across oocytes, while the proportion of the current in the first component was allowed to vary from one oocyte to the other.
RESULTS
Receptors Formed from Linked α3β4 Subunits Reproduce the ACh Sensitivity of Channels Expressed from Unlinked α3 and β4
Because much of our recent work focused on the human α3β4 nicotinic receptor, we chose this pair combination for our first tests. In oocytes, recombinant α3β4 receptors have a stoichiometry of two α3 and three β4 subunits (Boorman et al., 2000), probably in an αβαββ arrangement (by analogy to the muscle nAChR). This means that, out of the four possible tandem constructs, namely α3_α3, β4_β4, α3_β4, and β4_α3, only α3_β4 and β4_α3 could form a functional receptor when expressed with a monomer subunit (β or α).
Therefore, we expressed either of the linked constructs α3_β4 or β4_α3 together with β4 (in a molar ratio of 2:1 tandem:monomer). After injection of the α3_β4 + β4 cRNA combination, there was no functional expression, i.e., no response to 1 mM ACh (n = 5). In contrast with that, the linked construct with the opposite orientation (β4_α3) expressed efficiently with β4, giving an average maximum inward current to ACh in oocytes of 2.6 μA (n = 10, Table II, Fig. 2, 2 ng β4_α3:0.5 ng β4 cRNA injected), whereas injection of β4_α3 tandem alone gave no expression (1 mM ACh, n = 6 oocytes from two different batches, 2 ng of cRNA, see Table II), suggesting proper incorporation of the tandem constructs.
Recombinant receptors with and without linked subunits were identical in their ACh sensitivity, as shown by the indistinguishable dose–response curves in Fig. 2. ACh EC 50 and n H were 122 ± 8 μM and 1.98 ± 0.21 for receptors containing linked subunits and 134 ± 5 μM and 1.70 ± 0.10 for α3 + β4 receptors (n = 4 and 9, respectively).
Linked subunits with opposite orientation differ in two points, namely in the position of the linker with respect to the agonist binding site domains (Zhou et al., 2003) and in the length of the effective linker (i.e., the introduced linker together with the COOH terminus extracellular domain of the leading subunit and the signal peptide of the second subunit), which is shorter in α3_β4 than in β4_α3 because of the differences in the COOH-terminal end/signal peptide of the two subunits (Table I). Either of these two factors may have prevented functional expression when the tandem orientation is α_β. Another possibility is that the α3_β4 orientation does not allow the assembly of the receptor subunits in the correct order (if assembly is similar to that of muscle nAChRs; see Green, 1999).
Expression of other Tandem Constructs Alone or Together with β2 or β4
We proceeded to test which other β_α + β combinations gave functional nAChR, bearing in mind that the subunits known to form “pair” nAChRs in oocytes are α2, α3, or α4 together with either β2 or β4. In all oocytes injected with linked constructs of the form β4_α + β4, ACh application gave rise to large inward currents, indicating efficient production of nAChRs (maximum currents in response to 1 mM ACh are shown in Fig. 3, gray bars). On the other hand, of all the β2-containing tandems, the only combination that produced significant currents was β2_α4 + β2 (Fig. 3). No functional response was observed when subunit dimers containing α6 (β2_α6 + β2 and β4_α6 + β4) were expressed (n = 6 and 5, respectively; not depicted).
When α4_β2 tandems are expressed without a monomer, they produce functional receptors that are “dimers of pentamers”, i.e., dipentamers made up of two nAChR with different stoichiometry (Zhou et al., 2003). We therefore checked also which of the tandem constructs gave functional expression when injected in oocytes alone (Fig. 3, black bars) and found that β4_α2, β4_α4, and β2_α4 all produced functional receptors, with maximum ACh responses between 5% and 45% of those observed when the same tandem was injected together with a monomer. The tandem construct β4_α3 was the exception, in that it gave good functional responses only if expressed together with the β4 monomer. This indicates that this particular construct does not produce dimers of pentamers when expressed alone and is therefore more likely to give receptors with homogeneous stoichiometry.
We next checked that the tandem cRNA used in the transfections was pure and had appropriate molecular weight. This is shown by the gel in Fig. 4 A, where cRNAs for the α3 or β4 monomers (lanes 1 and 2) and those for the α3_β4 and β4_α3 tandem constructs (lanes 3 and 4) migrate as single bands at the molecular weights expected (∼1.8 and 3.4 Kb for single subunits and tandems, respectively). We sought additional confirmation that the actual fusion protein formed by expression of the tandem construct β4_α3 does not undergo proteolysis by analyzing the protein formed by SDS-PAGE fractionation, Western blotting, and detection by antiserum raised against the β4 subunit. This antibody detected a single band at the appropriate molecular weight (∼56 kD) in proteins solubilized from oocytes or HEK-293 cells transfected with the β4 monomer subunit alone (Fig. 4, B and C). However, there was no signal in proteins from oocytes or cells transfected with the tandem construct β4_α3.
Figure 4.

cRNA gel-electrophoresis (A) and Western blots of expressed proteins in oocytes (B) and HEK293 cells (C). Approximately 1 μg of α3 (1), β4 (2), α3_β4 tandem (3), and β4_α3 tandem (4) besides the RNA ladder (M) were separated on a 1.5% agarose-gel (A). The Western blot in B was obtained from oocytes injected with water only (MQ), β4 only, or β4_α3 tandem (T) and the Western blot in C from HEK293 cells transfected with no DNA (ND), α3 only, β4 only, or β4_α3 tandem (T). Detection by β4 antibody and visualization by chemoluminescence. Bands for the β4 subunit were detected at the expected size of 56 kD for both blots after 30-s exposure. No breakdown products were observed for the tandem constructs in either blots, even for longer exposures up to 1 h. Note that the tandem fusion protein (predicted size of 115 kD) was not detected by the β4 antibody used (see text).
Similar results were obtained when we used antiserum raised against the α3 subunit (not depicted). The most likely explanation is that in both expression systems, the amount of antigens formed is too low for either of these antibodies to detect it above background. Similar observations have been reported for GABAA subunit dimers (Baumann et al., 2001). Clearly, our results cannot shed any light on whether there is proteolysis of the tandems into single subunits, followed by unconstrained assembly of these subunits into functional receptors. Nevertheless, this “breakdown” hypothesis is unlikely for several reasons. First, we did not obtain functional channels when we expressed either the α3_β4 or the β4_α3 tandem on its own, or the two tandem constructs together. Second, no functional channels were observed when we expressed the α3_β4 tandem with any of the other (α2 to α6, and β2 to β4) monomer subunits (not depicted). Finally, the β4_α3 tandem failed to produce functional channels when expressed with α2, α3, or α4 (a β4 “breakdown” subunit should easily have formed a pair receptor with either of these αs), or when expressed with β2 (an α3 “breakdown” product should easily have formed a pair α3β2 receptor). No functional responses were detected after expression of β4_α3 with any subunit other than β4 (see also Table II). This is somewhat surprising, given that nAChRs containing α3, β4, and either α5 or β3 in a 2:2:1 ratio are functional (Wang et al., 1996; Boorman et al., 2000; Groot-Kormelink et al., 2001).
In β4_α3 + β4 nAChR Two Copies of α3 and Three Copies of β4 Participate in Channel Gating
Another requisite for the validation of the tandem construct expression technique is that the receptor formed from linked subunits should have the same stoichiometry as receptors expressed from monomeric constructs. In the case of oocyte-expressed α3β4 nAChRs, this means two copies of α3 and three of β4 (Boorman et al., 2000). A simple way of checking stoichiometry is to apply to linked-subunit receptors the reporter mutation approach that we have previously exploited for the α3β4 and α3β4β3 combinations (Boorman et al., 2000). Mutating the 9′ hydrophobic amino acid (Leu or Val) in the pore-lining second transmembrane domain of a nicotinic type subunit to a hydrophilic residue (Thr or Ser) makes the resulting receptor more sensitive to agonists (Revah et al., 1991), probably by destabilizing the closed state. The extent to which the agonist EC 50 is reduced is approximately proportional to the number of mutant subunits in the pentamer (Labarca et al., 1995; Chang et al., 1996; Chang and Weiss, 1999).
Fig. 5 shows the effect of inserting a 9′ LT mutation in either all the β4 subunits (filled triangles, leftmost curve) or all the α3 subunits (filled squares, middle curve; the dashed curve shows for reference the position of the wild-type tandem receptor) expressed in the linked β4_α3 + β4 nAChR (see Fig. 2). The mutation produces a greater leftward shift in the ACh dose–response curve if it is expressed in the β4 subunit, reducing the wild-type EC 50 of 122 ± 8 μM to 0.68 ± 0.02 μM (β4LT_α3 + β4LT, n = 7). A smaller effect is seen when α3 subunits are mutated, with a decrease in the EC 50 to 3.81 ± 0.18 μM (β4_α3LT + β4, n = 8). This indicates clearly that more β than α subunits contribute to the channel gate in the linked subunit receptor. This pattern is identical to that seen in α3 + β4 nAChRs, where mutating β4 or α3 produced a 292-fold or a 37-fold reduction in EC 50, respectively (Boorman et al., 2000). It is therefore reasonable to conclude that linked-subunit nAChR contain three β4 and two α3 subunits in the channel gating domain.
Linked-subunit nAChRs Contain More Than One Copy of the Monomer Subunit
As shown in the cartoons in Fig. 1, B and C, mutating all the β or all the α subunits expressed will produce receptors with three or two copies of the mutation in the channel even if the efficiency of tandem incorporation is poor. Thus, we next tested whether all functional receptors contained two copies of the fusion protein and one of the monomer. For this to be true, of the three β4 subunits in the channel, two must come from the tandem construct (βtandem) and one from the monomer (βmonomer). Mutating only the β4tandem subunits should then produce a leftward shift in the EC 50 corresponding to two mutation copies and therefore similar in magnitude to the shift produced by mutating α3 (i.e., ∼30-fold; Fig. 1 E, left). This was not the case: the β4LT_α3 + β4 dose–response curve (Fig. 6, filled squares) is very close to that of wild-type linked subunit receptors (the dotted curve), but quite different from the curve expected for a receptor containing two copies of the mutation (dashed reference curve). Thus, mutating β4tandem has very little effect on the EC 50 (100 ± 4 μM; n = 11), suggesting that β4 subunits from the concatemer contribute little to the channel of linked subunit nAChRs.
Figure 6.

Low ACh sensitivity of linked subunit receptors carrying the L9′T reporter mutation in β4tandem only. (A) Examples of inward currents elicited by bath-applied ACh in oocytes expressing β4LT_α3 + β4. (B) Data from oocytes injected with β4LT_α3 + β4 cRNAs (filled squares, n = 7). Fits as in Fig. 2. The concentration-response curve for β4_α3 + β4 wild-type (see Fig. 2 B) is shown for reference (dotted line), together with the concentration-response curve for the β4_α3LT + β4 combination (from Fig. 4 B, dashed line): the latter shows the EC 50 shift expected with the incorporation of two L9′T mutations.
This explanation implies that the three β4 subunits in the nAChR channel come mostly or exclusively from the monomer construct, which must be present in more than one copy because of incomplete incorporation of the fusion protein. This hypothesis was confirmed by the results of expressing β4_α3 + β4LT (shown by the experiment in Fig. 7, filled circles). The two components of this curve suggest that functional receptors produced by this combination are a heterogeneous population. Fitting the curve with a two-component Hill equation gave EC 50 values of 0.67 ± 0.10 and 31.7 ± 1.9 μM (n = 8). The first EC 50 is virtually identical with that of receptors in which all β4 subunits carried the mutation (i.e., 0.68 μM for linked-subunit receptors, see above) and strongly suggests that in a significant portion of receptors all β subunits come from the monomer construct (Fig. 1 D, right). There was considerable variability between oocytes in the proportion of each component of the dose–response curve. On average, the component with higher ACh sensitivity carried 58.6 ± 7.3% of maximum current, with a range of 21–100% (n = 13). Note that, for display purposes, the data in Fig. 7 are from a subset of four oocytes that had roughly comparable proportions of high- and low-sensitivity current. As the relative maximum Popen of the different wild-type and mutated forms of the receptor is unknown, it is impossible to translate directly proportions of current into receptor numbers (even disregarding channel block and desensitization).
The EC 50 of the low-sensitivity component is close to that expected for a receptor containing one mutation copy (i.e., one β4 from the monomer construct; a situation similar to that seen for α3β4β3LT receptors; Boorman et al., 2000). In the ideal case of equivalent effects for mutations carried by different subunits, the EC 50 for a one mutant receptor would fall between the two mutant EC 50 and the wild-type EC 50 (at the geometric mean of the two values, 22 μM in this case, dashed reference curve in Fig. 7). Expression of β4_α3 + β4LT therefore shows that linked-subunit receptors are a mixed population, in which pentamers contain one, two, or three β4 subunits provided by the monomer construct.
This hypothesis was confirmed by further experiments in which the β4_α3LT tandem construct was expressed together with β4LT, a combination that should in principle give rise to a nAChR with three mutation copies (Fig. 1 F, left). However, poor survival was observed for oocytes injected with this combination over the 2 d of incubation; the few surviving oocytes (25%) had high holding current, which was reduced by 46 ± 6% (n = 3) by application of a high concentration of the nicotinic channel blocker trimetaphan (10 μM). Both the high holding current and the poor oocyte health are likely to be due to spontaneous opening of nAChRs and were similar to observation of receptors with 5 mutations (α3LT + β4LT; not depicted). Such phenomena were not detectable for nAChRs with three mutation copies or less (with or without linked subunits) and suggest again that the monomer construct contributes more than one copy of the β4 subunit to the linked subunit receptor complex (Fig. 1 F, right).
It is therefore clear that the concatemer technique fails the ultimate test for providing a method for producing nAChR of defined subunit composition, as β4_α3 + β4 receptors do not contain two copies of β4_α3 and one of β4. Simply checking the ACh sensitivity of the receptor was not sufficient to detect this receptor heterogeneity.
DISCUSSION
Tandem constructs have been used to restrict subunit composition of recombinant multimeric channels. In the present study we assessed the feasibility and accuracy of functional nAChRs expression from tandem constructs. This strategy is an initial step toward obtaining recombinant nAChRs of defined and uniform stoichiometry for more complex compositions (i.e., those that contain more than one type of β or α subunit) and for controlling the insertion of mutations in defined subunits.
Using a short linker, we obtained efficient functional expression for several pair-type combinations (α2β4, α3β4, α4β4, and α4β2). α3β4-type receptors were chosen for further characterization, as the tandem construct for this combination (β4_α3) gave no nicotinic currents if expressed alone, but showed robust expression if expressed together with β4, suggesting that the resulting receptor population may be constituted entirely by pentamers with two copies of α and three of β. These β4_α3 + β4 tandem nAChRs were very similar to their nontandem equivalent, α3 + β4 nAChRs, in that they had identical dose–response curves to ACh and responded in the same way to the introduction of a reporter mutation in the TM2 domain of either all the α or all the β subunits expressed. This indicated that the channel function was not grossly disrupted by the linker in either binding or gating and that the channel gate contained the same subunit types for both wild-type and linked subunit receptors, namely two α and three β subunits. Nevertheless, mutating only a subset of the β subunit constructs (i.e., only β4tandem or β4monomer) revealed heterogeneity in the ACh sensitivity of linked-subunit receptors. This was probably due to channels containing variable numbers of mutations (i.e., β subunits from monomers or tandems) and ultimately to incomplete incorporation of the β4tandem subunit. This means that a substantial fraction of the pentamers may incorporate only the COOH terminus α of the fusion protein into the channel.
This hypothesis is illustrated in Fig. 1, A–C, which shows that experiments in which all α or all β subunits are mutated do not give any information on the origin of these subunits. Fig. 1 D shows how a mix of the different types of pentamers (i.e., one, two, or three β subunits from monomer) explains well the presence and properties of more than one component in the ACh dose–response curve when β4monomer was mutated (Fig. 7). These components are likely to represent channels that contain either one or three mutation copies. Because of the limited sensitivity of fitting several components to a dose–response curve, we cannot exclude or quantify the presence of an intermediate fraction of receptors containing two copies of the βmonomer.
In the light of this hypothesis, it is hard to account fully for the results of the complementary experiment in which β4tandem was mutated. The ACh sensitivity of these receptors was only slightly greater than that of wild-type receptors, suggesting that most linked subunit receptors contained only β subunits from the monomer. This is in contrast with the estimate that the equivalent receptors when βmonomer carried the mutation (high-sensitivity component in Fig. 7) accounted for ∼60% of current. However, it is worth noting that in the latter experiment, there was great variability between oocytes in the proportion of high-sensitivity current and that in 3 out of 13 oocytes this was the only component detectable.
Can Alternate Stoichiometries Account for Our Results?
It was recently shown (Zhou et al., 2003) that tandem constructs of α4β2 subunits give functional expression in the absence of a monomer construct, a situation analogous to that described for the expression of P2X receptors (which are trimeric) from tandem constructs alone (Stoop et al., 1999; Nicke et al., 2003). Nicotinic α4β2 tandem constructs form dipentamers of receptors, half of which contain two α subunits and half three α (in accord with the possibility that this combination may exist in two different subunit stoichiometries; Nelson et al., 2003). While it was reported that these dipentamers disappeared when expressed with monomer constructs, we must consider the possibility that the receptors formed by our tandem constructs include a similar dipentamer alone, or together with a correctly assembled pentamer (i.e., one containing one β4monomer). It is unlikely that this possibility accounts for our findings for several reasons, first of all because the β4_α3 tandem alone does not produce functional receptors. If we assume that a dipentamer does nevertheless form, this would predict that receptors with all α or all β subunit mutated would contain two and a half mutation copies on average (as half would have two and half would have three). In turn this would suggest that, if mutations are equivalent, these different mutant combinations should have the same sensitivity to ACh (i.e., the two curves in Fig. 5 should coincide). Finally, receptors in the dipentamer are formed exclusively by tandem constructs. In the expression of receptors in which only the monomer is mutated this would predict a significant wild-type component (which we did not detect).
Similarly, it is unlikely that the results of mutating β4monomer can be explained by populations of receptors with different numbers of α subunits. If α3β4 nAChRs with three α subunits are present, they must represent a small proportion of the total, given the clear-cut results of mutating all α or all β subunits. Furthermore, a three–α subunit receptor could in principle form from the expression of tandem constructs of opposite orientation or of β4_α3 + α3, neither of which produced functional receptors.
Another possibility is that linked subunit receptors assemble correctly, but that the characteristics of the hydrophilic 9′ mutation in TM2 are altered in a way that curtails their usefulness in establishing subunit incorporation or stoichiometry. It could be that, even though the wild-type ACh sensitivity is unchanged, the conformational constraints introduced by linking the subunits distort the channel to such an extent that the magnitude of the effect of the reporter mutation becomes dependent on the subunit that carries it and on the number of mutations already in the pore, rendering the results uninterpretable. It is hard to see how that could explain the two components in the dose–response curve for the receptor from mutant β4monomer. At any rate, this hypothesis entails that the mutation has different properties in linked subunit receptors versus α3 + β4 receptors. That in itself would mean that the tandem technique has failed in reproducing this channel.
Expression of Concatenated Subunits in Other Channel Superfamilies
The strategy of linking subunits has been exploited in practically every channel family with favorable subunit topology (i.e., both NH2 and COOH termini on the same side of the membrane), yielding a variety of useful data (for review see Nicke et al., 2003). Inconsistencies have emerged, notably for K+ channels (Liman et al., 1992; McCormack et al., 1992; Hurst et al., 1995; Silverman et al., 1996) and for P2X receptors (Stoop et al., 1999; Nicke et al., 2003). Additionally, early results on the stoichiometry of cyclic nucleotide–gated channels obtained by linking subunits are contradicted by new studies (Zimmerman, 2002). Steric hindrance, leading to the incomplete assembly of concatenated subunits into a channel and/or to the formation of higher-order complexes of receptors was invoked to explain such results. Note that in the cartoon in Fig. 1, our working hypothesis, based on similar conjectures, represents unassembled β4tandem subunits as trailing outside the receptor for simplicity, although it cannot be excluded that they are cleaved before exit from the endoplasmic reticulum, as suggested for P2X1 receptors (Nicke et al., 2003). An interesting feature is that incomplete dimer incorporation in our case results in the COOH-terminal subunit of the dimer participating to the channel. This is in contrast with the common observation that the NH2-terminal subunit in a multimer incorporates most efficiently, and strengthens our conjecture that premature termination of translation or 3′ degradation of cRNA does not take place to produce truncated dimers.
In conclusion, our data show that the incomplete incorporation of the linked subunits and the consequent heterogeneity of the assembled receptor were not detected by a simple dose–response curve, but required extensive functional testing aided by reporter mutations. Clearly, the ultimate failure of our particular version of the linked subunit strategy does not exclude that it may be possible to engineer a different linker to produce homogeneous defined composition receptors in which the linked subunits incorporate efficiently. The difficulty of this task probably lies in balancing opposing demands on the linker, as a long flexible sequence will allow the formation of functional dimers of different pentamers, whereas a short linker would better constrain stoichiometry but may damage incorporation efficiency of the fusion protein. Nevertheless, our data clearly set out a minimum range of tests for a linked subunit receptor to be deemed to have successfully reproduced the characteristics of the unlinked receptor.
Acknowledgments
Our work was supported by the Wellcome Trust (Project Grant 064652 to Lucia G. Sivilotti) and the MRC (Ph.D. studentship to Steven D. Broadbent).
Angus C. Nairn served as editor.
James Boorman's present address is Department of Haematology, University of Cambridge, EABC Site, Long Road, Cambridge CB2 2PT, UK.
Abbreviation used in this paper: nAChR, nicotinic acetylcholine receptor.
References
- Akopian, A.N., L. Sivilotti, and J.N. Wood. 1996. A tetrodotoxin-resistant voltage-gated sodium-channel expressed by sensory neurons. Nature. 379:257–262. [DOI] [PubMed] [Google Scholar]
- Baumann, S.W., R. Baur, and E. Sigel. 2001. Subunit arrangement of γ-aminobutyric acid type A receptors. J. Biol. Chem. 276:36275–36280. [DOI] [PubMed] [Google Scholar]
- Baumann, S.W., R. Baur, and E. Sigel. 2002. Forced subunit assembly in α1β2γ2 GABAA receptors: insight into the absolute arrangement. J. Biol. Chem. 277:46020–46025. [DOI] [PubMed] [Google Scholar]
- Baumann, S.W., R. Baur, and E. Sigel. 2003. Individual properties of the two functional agonist sites in GABAA receptors. J. Neurosci. 23:11158–11166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beato, M., P.J. Groot-Kormelink, D. Colquhoun, and L.G. Sivilotti. 2002. Openings of the rat recombinant α1 homomeric glycine receptor as a function of the number of agonist molecules bound. J. Gen. Physiol. 119:443–466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boorman, J.P., P.J. Groot-Kormelink, and L.G. Sivilotti. 2000. Stoichiometry of human recombinant neuronal nicotinic receptors containing the β3 subunit expressed in Xenopus oocytes. J. Physiol. 529:565–577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang, Y., R. Wang, S. Barot, and D.S. Weiss. 1996. Stoichiometry of a recombinant GABAA receptor. J. Neurosci. 16:5415–5424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang, Y.C., and D.S. Weiss. 1999. Allosteric activation mechanism of the α1 β2 γ2 γ-aminobutyric acid type A receptor revealed by mutation of the conserved M2 leucine. Biophys. J. 77:2542–2551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colquhoun, D., N. Unwin, C. Shelley, C.J. Hatton, and L. Sivilotti. 2003. Nicotinic acetylcholine receptors. Burger's Medicinal Chemistry and Drug Discovery: Fundamentals of Medicinal Chemistry. D. Abraham, editor. John Wiley & Sons Inc., New York. 357–405.
- Conroy, W.G., and D.K. Berg. 1995. Neurons can maintain multiple classes of nicotinic receptors distinguished by different subunit compositions. J. Biol. Chem. 270:4424–4431. [DOI] [PubMed] [Google Scholar]
- Elliott, K.J., S.B. Ellis, K.J. Berckhan, A. Urrutia, L.E. Chavez-Noriega, E.C. Johnson, G. Velicelebi, and M.M. Harpold. 1996. Comparative structure of human neuronal α2-α7 and β2-β4 nicotinic acetylcholine receptor subunits and functional expression of the α2, α3, α4, α7, β2, and β4 subunits. J. Mol. Neurosci. 7:217–228. [DOI] [PubMed] [Google Scholar]
- Forsayeth, J.R., and E. Kobrin. 1997. Formation of oligomers containing the β3 and β4 subunits of the rat nicotinic receptor. J. Neurosci. 17:1531–1538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gordon, S.E., and W.N. Zagotta. 1995. Subunit interactions in coordination of Ni2+ in cyclic nucleotide-gated channels. Proc. Natl. Acad. Sci. USA. 92:10222–10226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Green, W.N. 1999. Ion channel assembly: creating structures that function. J. Gen. Physiol. 113:163–169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Groot-Kormelink, P.J., J.P. Boorman, and L.G. Sivilotti. 2001. Formation of functional α3β4α5 human neuronal nicotinic receptors in Xenopus oocytes: a reporter mutation approach. Br. J. Pharmacol. 134:789–796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Groot-Kormelink, P.J., and W.H.M.L. Luyten. 1997. Cloning and sequence of full-length cDNAs encoding the human neuronal nicotinic acetylcholine receptor (nAChR) subunits β3 and β4 and expression of seven nAChR subunits in the human neuroblastoma cell line SH-SY5Y and/or IMR-32. FEBS Lett. 400:309–314. [DOI] [PubMed] [Google Scholar]
- Groot-Kormelink, P.J., W.H.M.L. Luyten, D. Colquhoun, and L.G. Sivilotti. 1998. A reporter mutation approach shows incorporation of the “orphan” subunit β3 into a functional nicotinic receptor. J. Biol. Chem. 273:15317–15320. [DOI] [PubMed] [Google Scholar]
- Hurst, R.S., R.A. North, and J.P. Adelman. 1995. Potassium channel assembly from concatenated subunits: effects of proline substitutions in S4 segments. Receptors Channels. 3:263–272. [PubMed] [Google Scholar]
- Im, W.B., J.F. Pregenzer, J.A. Binder, G.H. Dillon, and G.L. Alberts. 1995. Chloride channel expression with the tandem construct of α6-β2 GABAA receptor subunit requires a monomeric subunit of α6 or γ2. J. Biol. Chem. 270:26063–26066. [DOI] [PubMed] [Google Scholar]
- Isacoff, E.Y., Y.N. Jan, and L.Y. Jan. 1990. Evidence for the formation of heteromultimeric potassium channels in Xenopus oocytes. Nature. 345:530–534. [DOI] [PubMed] [Google Scholar]
- Labarca, C., M.W. Nowak, H. Zhang, L. Tang, P. Deshpande, and H.A. Lester. 1995. Channel gating governed symmetrically by conserved leucine residues in the M2 domain of nicotinic receptors. Nature. 376:514–516. [DOI] [PubMed] [Google Scholar]
- Liman, E.R., J. Tytgat, and P. Hess. 1992. Subunit stoichiometry of a mammalian K+ channel determined by construction of multimeric cDNAs. Neuron. 9:861–871. [DOI] [PubMed] [Google Scholar]
- McCormack, K., L. Lin, L.E. Iverson, M.A. Tanouye, and F.J. Sigworth. 1992. Tandem linkage of Shaker K+ channel subunits does not ensure the stoichiometry of expressed channels. Biophys. J. 63:1406–1411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nelson, M.E., A. Kuryatov, C.H. Choi, Y. Zhou, and J. Lindstrom. 2003. Alternate stoichiometries of α4β2 nicotinic acetylcholine receptors. Mol. Pharmacol. 63:332–341. [DOI] [PubMed] [Google Scholar]
- Nicke, A., J. Rettinger, and G. Schmalzing. 2003. Monomeric and dimeric byproducts are the principal functional elements of higher order P2X1 concatamers. Mol. Pharmacol. 63:243–252. [DOI] [PubMed] [Google Scholar]
- Quick, M.W., and H.A. Lester. 1994. Methods for expression of excitability proteins in Xenopus oocytes. Ion Channels of Excitable Cells. T. Narahashi, editor. Academic Press, San Diego. 261–279.
- Ramirez-Latorre, J., C.R. Yu, X. Qu, F. Perin, A. Karlin, and L. Role. 1996. Functional contributions of α5 subunit to neuronal acetylcholine receptor channels. Nature. 380:347–351. [DOI] [PubMed] [Google Scholar]
- Revah, F., D. Bertrand, J.-L. Galzi, A. Devillers-Thiéry, C. Mulle, N. Hussy, S. Bertrand, M. Ballivet, and J.-P. Changeux. 1991. Mutations in the channel domain alter desensitization of a neuronal nicotinic receptor. Nature. 353:846–849. [DOI] [PubMed] [Google Scholar]
- Sands, S.B., A.C.S. Costa, and J.W. Patrick. 1993. Barium permeability of neuronal nicotinic receptor α7 expressed in Xenopus oocytes. Biophys. J. 65:2614–2621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schorge, S., and D. Colquhoun. 2003. Studies of NMDA receptor function and stoichiometry with truncated and tandem subunits. J. Neurosci. 23:1151–1158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silverman, S.K., H.A. Lester, and D.A. Dougherty. 1996. Subunit stoichiometry of a heteromultimeric G protein-coupled inward-rectifier K+ channel. J. Biol. Chem. 271:30524–30528. [DOI] [PubMed] [Google Scholar]
- Stoop, R., S. Thomas, F. Rassendren, E. Kawashima, G. Buell, A. Surprenant, and R.A. North. 1999. Contribution of individual subunits to the multimeric P2X2 receptor: estimates based on methanethiosulfonate block at T336C. Mol. Pharmacol. 56:973–981. [DOI] [PubMed] [Google Scholar]
- Wang, F., V. Gerzanich, G.B. Wells, R. Anand, X. Peng, K. Keyser, and J. Lindstrom. 1996. Assembly of human neuronal nicotinic receptor α5 subunits with α3, β2, and β4 subunits. J. Biol. Chem. 271:17656–17665. [DOI] [PubMed] [Google Scholar]
- Zhou, Y., M.E. Nelson, A. Kuryatov, C. Choi, J. Cooper, and J. Lindstrom. 2003. Human α4β2 acetylcholine receptors formed from linked subunits. J. Neurosci. 23:9004–9015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zimmerman, A.L. 2002. Two B or not two B? Questioning the rotational symmetry of tetrameric ion channels. Neuron. 36:997–999. [DOI] [PubMed] [Google Scholar]