

Abstract
The experimental question is whether or not sugar can be a substance of abuse and lead to a natural form of addiction. “Food addiction” seems plausible because brain pathways that evolved to respond to natural rewards are also activated by addictive drugs. Sugar is noteworthy as a substance that releases opioids and dopamine and thus might be expected to have addictive potential. This review summarizes evidence of sugar dependence in an animal model. Four components of addiction are analyzed. “Bingeing”, “withdrawal”, “craving” and cross-sensitization are each given operational definitions and demonstrated behaviorally with sugar bingeing as the reinforcer. These behaviors are then related to neurochemical changes in the brain that also occur with addictive drugs. Neural adaptations include changes in dopamine and opioid receptor binding, enkephalin mRNA expression and dopamine and acetylcholine release in the nucleus accumbens. The evidence supports the hypothesis that under certain circumstances rats can become sugar dependent. This may translate to some human conditions as suggested by the literature on eating disorders and obesity.
Keywords: binge eating, dopamine, acetylcholine, opioid, nucleus accumbens, withdrawal, craving, behavioral sensitization, rat
1. OVERVIEW
Neural systems that evolved to motivate and reinforce foraging and food intake also underlie drug-seeking and self-administration. The fact that some of these drugs can cause addiction raises the logical possibility that some foods might also cause addcition. Many people claim that they feel compelled to eat sweet foods, similar in some ways to how an alcoholic might feel compelled to drink. Therefore, we developed an animal model to investigate why some people have difficulty moderating their intake of palatable foods, such as sweet beverages.
In this animal model, rats are food deprived daily for 12 h, then after a delay of 4 h into their normal circadian-driven active period, they are given 12-h access to a sugar solution and chow. As a result, they learn to drink the sugar solution copiously, especially when it first becomes available each day.
After a month on this intermittent-feeding schedule, the animals show a series of behaviors similar to the effects of drugs of abuse. These are categorized as “bingeing”, meaning unusually large bouts of intake, opiate-like “withdrawal” indicated by signs of anxiety and behavioral depression (Colantuoni et al., 2001, 2002), and “craving” measured during sugar abstinence as enhanced responding for sugar (Avena et al., 2005). There are also signs of both locomotor and consummatory “cross-sensitization” from sugar to drugs of abuse (Avena et al., 2004, Avena and Hoebel, 2003b). Having found these behaviors that are common to drug dependency with supporting evidence from other laboratories (Gosnell, 2005, Grimm et al., 2005, Wideman et al., 2005), the next question is why this happens.
A well-known characteristic of addictive drugs is their ability to cause repeated, intermittent increases in extracellular dopamine (DA) in the nucleus accumbens (NAc) (Di Chiara and Imperato, 1988, Hernandez and Hoebel, 1988, Wise et al., 1995). We find that rats with intermittent access to sugar will drink in a binge-like manner that releases DA in the NAc each time, like the classic effect of most substances of abuse (Avena et al., 2006, Rada et al., 2005b). This consequently leads to changes in the expression or availability of DA receptors (Colantuoni et al., 2001, Spangler et al., 2004).
Intermittent sugar access also acts by way of opioids in the brain. There are changes in opioid systems such as decreased enkephalin mRNA expression in the accumbens (Spangler et al., 2004). Signs of withdrawal seem to be largely due to the opioid modifications since withdrawal can be obtained with the opioid antagonist naloxone. Food deprivation is also sufficient to precipitate opiate-like withdrawal signs (Avena, Bocarsly, Rada, Kim and Hoebel, unpublished, Colantuoni et al., 2002). This withdrawal state involves at least two neurochemical manifestations. First is a decrease in extracellular DA in the accumbens, and second is the release of acetylcholine (ACh) from accumbens interneurons. These neurochemical adaptations in response to intermittent sugar intake mimic the effects of opiates.
The theory is formulated that intermittent, excessive intake of sugar can have dopaminergic, cholinergic and opioid effects that are similar to psychostimulants and opiates, albeit smaller in magnitude. The overall effect of these neurochemical adaptations is mild, but well-defined, dependency (Hoebel et al., 1999, Leibowitz and Hoebel, 2004, Rada et al., 2005a). This review compiles studies from our laboratory and integrates related results obtained by others using animal models, clinical accounts and brain imaging to answer the question: can sugar, in some conditions, be “addictive”?
2. DEFINING ADDICTION
Throughout this review we use several terms with definitions for which there is not universal agreement. Addiction research traditionally focuses on drugs of abuse, such as morphine, cocaine, nicotine and alcohol. However, recently a variety of “addictions” to non-drug entities, including gambling, sex, and in this review, food, have been investigated (Bancroft and Vukadinovic, 2004, Comings et al., 2001, Petry, 2006). The term “addiction” implies psychological dependence and thus is a mental or cognitive problem, not just a physical ailment. “Addiction” is often used synonymously with the term “dependence” (Nelson et al., 1982) as defined by DSM-IV-TR (American Psychiatric Association, 2000). We will use the term dependence in its all-encompassing meaning to describe the results of a battery of animal studies that model human drug addiction in each of its major phases (Koob and Le Moal, 2005).
Drug dependence is characterized by compulsive, sometimes uncontrollable, behaviors that occur at the expense of other activities and intensify with repeated access. Dependence is difficult to demonstrate convincingly in laboratory animals, but criteria have been suggested using animal models. We have used models that were developed with rats for studying drug dependence and adapted them to test for signs of sugar dependence.
Bingeing
The diagnostic criteria for addiction can be grouped into three stages (American Psychiatric Association, 2000, Koob and Le Moal, 1997). The first, bingeing, is defined as the escalation of intake with a high proportion of intake at one time, usually after a period of voluntary abstinence or forced deprivation. Enhanced intake in the form of binges may result from both sensitization and tolerance to the sensory properties of a substance of abuse that occurs with its repeated delivery. Sensitization, which is described in greater detail below, is an increase in responsiveness to a repeatedly presented stimulus. Tolerance is a gradual decrease in responsiveness, such that more of the substance is needed to produce the same effect (McSweeney et al., 2005). Both are thought to influence the powerful, acute reinforcing effects of drugs of abuse and are important at the beginning of the addiction cycle since both can increase responding and intake (Koob and Le Moal, 2005).
Withdrawal
Signs of withdrawal become apparent when the abused substance is no longer available or chemically blocked. We will discuss withdrawal in terms of opiate withdrawal, which has a clearly defined set of symptoms (Martin et al., 1963, Way et al., 1969). Anxiety can be operationally defined and measured in animals using the elevated plus-maze, in which anxious animals will avoid spending time on the open arms of the maze (File et al., 2004). This test has been extensively validated for both general anxiety (Pellow et al., 1985) and anxiety induced by drug withdrawal (File and Andrews, 1991). Behavioral depression in animals can also be inferred, without reference to emotion, using the forced-swim test, which measures swimming escape efforts vs. passive floating (Porsolt et al., 1978). When signs of opiate withdrawal are precipitated with naloxone, it suggests that inactivation of opioid receptors is the cause. When the same signs are produced spontaneously during abstinence, one can surmise that it is due to lack of stimulation of some opioid system.
Craving
The third stage of addiction, craving, occurs when motivation is enhanced, usually after an abstinence period (Vanderschuren and Everitt, 2005, Weiss, 2005). “Craving” remains a poorly defined term that is often used to describe the intense desire to self-administer drugs in humans (Wise, 1988). For lack of a better word, we will use the term “craving” as defined by increased efforts to obtain a substance of abuse or its associated cues as a result of addiction and abstinence. “Craving” often has reference to extreme motivation, which can be measured using operant conditioning. If abstinence makes the animal significantly increase its lever pressing, one can take this as a sign of enhanced motivation.
Sensitization
In addition to the above diagnostic criteria, behavioral sensitization is thought to underlie some aspects of drug dependence (Vanderschuren and Kalivas, 2000). Behavioral sensitization is typically measured as increased locomotion in response to repeated administrations of a drug. For example, after repeated doses of amphetamine followed by abstinence, a challenge dose, which has little or no effect in naïve animals, causes marked hyperactivity (Antelman and Caggiula, 1996, Glick et al., 1986). Animals sensitized to one substance often show cross-sensitization, which is defined as an increased locomotor response to a different drug or substance. Cross-sensitization can also be manifest in consummatory behavior (Piazza et al., 1989). Animals sensitized to one drug may show increased intake of a different drug. In other words, one drug acts as a “gateway” to another. For example, animals sensitized to amphetamine show accelerated escalation of cocaine intake (Ferrario and Robinson, 2007), and animals sensitized to nicotine consume more alcohol compared with non-sensitized animals (Blomqvist et al., 1996). This behavior is thought to occur when different drugs activate the same neural circuitry, and it is the reason why many clinicians require complete drug abstention as a condition of treatment for addicts (Wise, 1988).
The first question addressed by this review is whether any of these operationally defined behavioral characteristics of substance dependence can be found with intermittent sugar access. The second question explores neural systems to discover how sugar could have effects like a drug of abuse.
3. DRUGS OF ABUSE AND PALATABLE FOOD ACTIVATE A COMMON SUBSET OF NEURAL SYSTEMS
Overlaps in the brain circuitry activated by food and drug intake suggests that different types of reinforcers (natural and artificial) stimulate some of the same neural systems (Hoebel, 1985, Hernandez and Hoebel, 1988, Kelley et al., 2002, Le Magnen, 1990, Volkow and Wise, 2005, Wise, 1988, 1989). There are several regions in the brain involved in the reinforcement of both feeding and drug intake (Hernandez and Hoebel, 1988, Kalivas and Volkow, 2005, Kelley et al., 2005, Koob and Le Moal, 2005, Mogenson and Yang, 1991, Wise, 1997, Yeomans, 1995), and many neurotransmitters, as well as hormones, have been studied in these and related brain regions (Harris et al., 2005, Kalivas, 2004, Leibowitz and Hoebel, 2004, Schoffelmeer et al., 2001, Stein and Belluzzi, 1979). This review will focus on DA, the opioids, and ACh in the NAc shell, which so far, are the neurotransmitters that we have found to be involved with the reinforcing effects of intermittent sugar intake.
3.A. Dopamine
It is well established that addictive drugs activate DA-containing neurons in areas of the brain that process behavior reinforcement. This was shown for drugs delivered systemically (Di Chiara and Imperato, 1988, Radhakishun et al., 1983), and for drugs micro-injected or infused locally (Hernandez and Hoebel, 1988, Mifsud et al., 1989). The mesolimbic DA projection from the ventral tegmental area (VTA) to the NAc is frequently implicated in reinforcement functions (Wise and Bozarth, 1984). The NAc is important for several components of “reward” including food seeking and reinforcement of learning, incentive motivation, stimulus salience and signaling a stimulus change (Bassareo and Di Chiara, 1999, Berridge and Robinson, 1998, Salamone, 1992, Schultz et al., 1997, Wise, 1988). Any neurotransmitter that directly or indirectly stimulates DA cell bodies in the VTA reinforces local self-administration, including opioids such as enkephalin (Glimcher et al., 1984), non-opioid peptides such as neurotensin (Glimcher et al., 1987) and many drugs of abuse (Bozarth and Wise, 1981, Gessa et al., 1985, McBride et al., 1999). Some addictive drugs also act at DA terminals (Cheer et al., 2004, Mifsud et al., 1989, Nisell et al., 1994, Westerink et al., 1987, Yoshimoto et al., 1992). Thus, any substance that repeatedly causes the release of DA or reduces DA reuptake at terminals via these circuits may be a candidate for abuse.
A variety of foods can release DA in the NAc, including lab chow, sugar, saccharin, and corn oil (Bassareo and Di Chiara, 1997, Hajnal et al., 2004, Liang et al., 2006, Mark et al., 1991, Rada et al., 2005b). The rise in extracellular DA can outlast the meal in food-deprived rats (Hernandez and Hoebel, 1988). However, in satiated animals, this DA release appears to be contingent on novelty since it wanes with repeated access, even when the food is palatable (Bassareo and Di Chiara, 1997, Rada et al., 2005b). An exception, which is described below (Section 5.C.), is when animals are food deprived and fed sugar intermittently.
Extracellular DA decreases in reaction to drug withdrawal (Acquas et al., 1991, Acquas and Di Chiara, 1992, Rada et al., 2004, Rossetti et al., 1992). The symptoms of withdrawal from dopaminergic drugs are less well-defined than those observed during withdrawal from opiates. Therefore, it may be easier to discern the signs of withdrawal when using foods that release both DA and opioids. Sugar is one such food.
3.B. Opioids
Opioid peptides are heavily expressed throughout the limbic system and linked to DA systems in many parts of the forebrain (Haber and Lu, 1995, Levine and Billington, 2004, Miller and Pickel, 1980). The endogenous opioid systems exert some of their effects on reinforcement processing by interacting with DA systems (Bozarth and Wise, 1986, Di Chiara and Imperato, 1986, Leibowitz and Hoebel, 2004). The opioid peptide enkephalin in the NAc has been related to reward (Bals-Kubik et al., 1989, Bozarth and Wise, 1981, Olds, 1982, Spanagel et al., 1990) and can activate both mu and delta receptors to increase the release of DA (Spanagel et al., 1990). Morphine alters gene expression of endogenous opioid peptides while increasing opioid peptide production in the NAc (Przewlocka et al., 1996, Spangler et al., 2003,Turchan et al., 1997). Opioids are also important components of this system as cotransmitters with GABA in some accumbens and dorsal striatal outputs (Kelley et al., 2005).
Repeated use of opiates, or even some non-opiate drugs, can result in mu-opioid receptor sensitization in several regions, including the NAc (Koob et al., 1992, Unterwald, 2001). A mu-receptor antagonist injected into the NAc will attenuate the rewarding effects of heroin (Vaccarino et al., 1985), and systemically such drugs have been used as a treatment for alcoholism and heroin dependence (Deas et al., 2005, Foster et al., 2003, Martin, 1975, O’Brien, 2005, Volpicelli et al., 1992).
Ingestion of palatable foods has effects via endogenous opioids in a variety of sites (Dum et al., 1983, Mercer and Holder, 1997, Tanda and Di Chiara, 1998), and the injection of mu-opioid agonists in the NAc increases intake of palatable foods rich in fat or sugar (Zhang et al., 1998, Zhang and Kelley, 2002). Opioid antagonists, on the other hand, decrease ingestion of sweet food and shorten meals of palatable, preferred foods, even at doses that have no effect on standard chow intake (Glass et al., 1999). This opioid-palatability link is further characterized by theories in which the reinforcing effect is dissociated into a dopaminergic system for incentive motivation and an opioid “liking” or “pleasure” system for hedonic responses (Berridge, 1996, Robinson and Berridge, 1993, Stein, 1978). Evidence that opioids in the NAc influence hedonic reactions comes from data showing that morphine enhances rats’ positive facial taste reactivity for a sweet solution in the mouth (Pecina and Berridge, 1995). The dissociation between the “wanting” and “liking” systems is also suggested by studies in humans (Finlayson et al., 2007).
3.C. Acetylcholine
Several cholinergic systems in the brain have been implicated in both food and drug intake, and related to DA and the opioids (Kelley et al., 2005, Rada et al., 2000, Yeomans, 1995). Focusing on ACh interneurons in the NAc, systemic administration of morphine decreases ACh turnover (Smith et al., 1984), a finding that was confirmed by in vivo microdialysis in freely-behaving rats (Fiserova et al., 1999, Rada et al., 1991a, 1996). Cholinergic interneurons in the NAc may selectively modulate enkephalin gene expression and peptide release (Kelley et al., 2005). During morphine withdrawal, extracellular ACh increases in the NAc while DA is low, suggesting that this neurochemical state could be involved in the aversive aspects of withdrawal (Pothos et al., 1991, Rada et al., 1991b, 1996). Likewise, both nicotine and alcohol withdrawal increase extracellular ACh, while decreasing DA in the NAc (De Witte et al., 2003, Rada et al., 2001, 2004). This withdrawal state may involve behavioral depression, because M1-receptor agonists injected in the NAc can cause depression in the forced-swim test (Chau et al., 1999). The role of ACh in drug withdrawal has been further demonstrated with systemically administered acetylcholinesterase inhibitors, which can precipitate withdrawal signs in non-dependent animals (Katz and Valentino, 1984, Turski et al., 1984).
ACh in the NAc has also been implicated in food intake. We theorize that its overall muscarinic effect is to inhibit feeding at M1 receptors since local injection of the mixed muscarinic agonist arecholine will inhibit feeding, and this effect can be blocked by the relatively specific M1 antagonist pirenzapine (Rada and Hoebel, unpublished). Feeding to satiety increases extracellular ACh in the NAc (Avena et al., 2006, Mark et al., 1992). A conditioned taste aversion also increases ACh in the NAc and simultaneously lowers DA (Mark et al., 1991, 1995). D-fenfluramine combined with phentermine (Fen-Phen) increases extracellular ACh in the NAc at a dose that inhibits both eating and cocaine self-administration (Glowa et al., 1997, Rada and Hoebel, 2000). Rats with accumbal ACh toxin-induced lesions are hyperphagic relative to non-lesioned rats (Hajnal et al., 2000).
DA/ACh balance is controlled in part by hypothalamic systems for feeding and satiety. Norepinephrine and galanin, which induce eating when injected in the paraventricular nucleus (PVN), lower accumbens ACh (Hajnal et al., 1997, Rada et al., 1998). An exception is neuropeptide-Y, which fosters eating when injected into the PVN, but does not increase DA release nor lower ACh (Rada et al., 1998). In accord with the theory, the satiety-producing combination of serotonin plus CCK injection into the PVN increases accumbens ACh (Helm et al., 2003).
It is very interesting that when DA is low and extracellular ACh is high, this apparently creates not satiety, but instead an aversive state (Hoebel et al., 1999), as during behavioral depression (Zangen et al., 2001, Rada et al., 2006), drug withdrawal (Rada et al., 1991b, 1996, 2001, 2004) and conditioned taste aversion (Mark et al., 1995). We conclude that when ACh acts as a post-synaptic M1 agonist it has effects opposite to DA, and thus may act as a “brake” on dopaminergic functions (Hoebel et al., 1999, Rada et al., 2007) causing satiety when DA is high and behavioral depression when DA is relatively low.
4. BEHAVIORAL SIMILARITIES BETWEEN DRUG SELF-ADMINISTRATION AND INTERMITTENT, EXCESSIVE SUGAR INTAKE
The concept of “sugar addiction” has been bandied about for many years. Clinical accounts of “sugar addiction” have been the topic of many best-selling books and the focus for popular diet programs (Appleton, 1996, DesMaisons, 2001, Katherine, 1996, Rufus, 2004). In these accounts, people describe symptoms of withdrawal when they deprive themselves of sugar-rich foods. They also describe food craving, particularly for carbohydrates, chocolate, and sugar, which can trigger relapse and impulsive eating. This leads to a vicious cycle of self-medication with sweet foods that may result in obesity or an eating disorder.
Although food addiction has been popular in the media and proposed to be based on brain neurochemistry (Hoebel et al., 1989, Le Magnen, 1990), this phenomenon has only recently been systematically studied in the laboratory.
As outlined in the overview in Section 1, we use a feeding schedule that induces rats to binge on a sugar solution, then apply the criteria for drug dependence that are presented in Section 2 and test for the behavioral and neurochemical commonalties given in Section 3. Rats are given 12-h daily access to an aqueous 10% sucrose solution (25% glucose in some experiments) and lab chow, followed by 12 h of deprivation for three or more weeks (i.e., Daily Intermittent Sugar and Chow). These rats are compared with control groups such as Ad libitum Sugar and Chow, Ad libitum Chow, or Daily Intermittent Chow (12-h deprivation followed by 12-h access to lab chow). For the intermittent access groups, availability is delayed 4 h into the animal’s active period in order to stimulate feeding, which normally ensues at the onset of the dark cycle. Rats maintained on the Daily Intermittent Sugar and Chow regimen enter a state that resembles drug dependence on several dimensions. These are divided into behavioral (Section 4) and neurochemical (Section 5) similarities to drug dependence.
4.A. “Bingeing”: Escalation of daily sugar intake and large meals
Escalation of intake is a characteristic of drugs of abuse. This may be a combination of tolerance, in which more of an abused substance is needed to produce the same euphoric effects (Koob and Le Moal, 2005), and sensitization, such as locomotor sensitization, in which the substance produces enhanced behavioral activation (Vezina et al., 1989). Studies using drug self-administration usually limit access to a few hours per day, during which animals will self-administer in regular intervals that vary as a function of the dose received (Gerber and Wise, 1989) and in a manner that keeps extracellular DA elevated above a baseline, or “trigger point” in the NAc (Ranaldi et al., 1999, Wise et al., 1995). The length of daily access has been shown to critically affect subsequent self-administration behavior. For example, the most cocaine is self-administered during the first 10 min of a session when access is at least 6 h per day (Ahmed and Koob, 1998). Limited periods of access, to create “binges”, have been useful, because the pattern of self-administration behavior that emerges is similar to that of a “compulsive” drug user (Markou et al., 1993, Mutschler and Miczek, 1998, O’Brien et al., 1998). Even when drugs, such as cocaine, are given with unlimited access, humans or laboratory animals will self-administer them in repetitive episodes or “binges” (Bozarth and Wise, 1985, Deneau et al., 1969). However, experimenter-imposed intermittent access is better than ad libitum access for experimental purposes, since it becomes very likely that the animal will take at least one large binge at the onset of the drug-availability period. Moreover, a period of food restriction can enhance drug intake (Carr, 2006, Carroll, 1985) and has been shown to produce compensatory neruoadaptations in the mesoaccumbens DA system (Pan et al., 2006).
The behavioral findings with sugar are similar to those observed with drugs of abuse. Rats fed daily intermittent sugar and chow escalate their sugar intake and increase their intake during the first hour of daily access, which we define as a “binge” (Colantuoni et al., 2001). The animals with ad libitum access to a sugar solution tend to drink it throughout the day, including their inactive period. Both groups increase their overall intake, but the limited-access animals consume as much sugar in 12 h as ad libitum-fed animals do in 24 h. Detailed meal pattern analysis using operant conditioning (fixed-ratio 1) reveals that the limited animals consume a large meal of sugar at the onset of access, and larger, fewer meals of sugar throughout the access period, compared with animals drinking sugar ad libitum (Fig. 1; Avena and Hoebel, unpublished). Rats fed Daily Intermittent Sugar and Chow regulate their caloric intake by decreasing their chow intake to compensate for the extra calories obtained from sugar, which results in a normal body weight (Avena, Bocarsly, Rada, Kim and Hoebel, unpublished, Avena et al., 2003b, Colantuoni et al., 2002).
Figure 1.
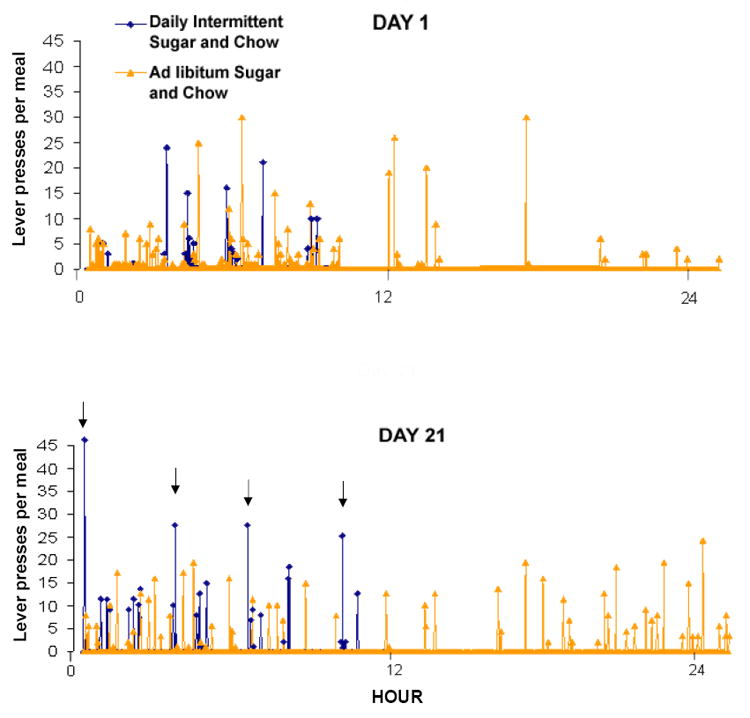
Meal analysis of two representative rats living in operant chambers. The one maintained on Daily Intermittent Sucrose and Chow (black lines) had an increased intake of sugar compared with one given Ad libitum Sucrose and Chow (grey lines). Hour 0 is 4 h into the dark phase. Each lever press delivers 0.1 mL of 10% sucrose. A sugar meal is defined as ending when the rat does not press for 2 min. Both rats consume several meals of about equal size on day 1 (top panel). Note that the rat with sugar available 24 h also drinks during the inactive (light) phase. By day 21 (bottom panel), the rat with sucrose and chow available for only 12 h consumes an initial “binge” of sucrose (indicated by the first arrow), followed by fewer, but larger meals, than the rat with sucrose and chow ad libitum. Sugar-bingeing rats are the ones that show signs of dependency in a battery of tests.
4.B. “Withdrawal”: Anxiety and behavioral depression induced by an opioid-antagonist or food deprivation
As described in Section 2, animals can show signs of opiate withdrawal after repeated exposure when the substance of abuse is removed, or the appropriate synaptic receptor is blocked. For example, an opioid antagonist can be used to precipitate withdrawal in the case of opiate dependency (Espejo et al., 1994, Koob et al., 1992). In rats, opiate withdrawal causes severe somatic signs (Martin et al., 1963, Way et al., 1969), decreases in body temperature (Ary et al., 1976), aggression (Kantak and Miczek, 1986), and anxiety (Schulteis et al., 1998), as well as a motivational syndrome characterized by dysphoria and depression (De Vries and Shippenberg, 2002, Koob and Le Moal, 1997).
These signs of opioid withdrawal have been noted after intermittent access to sugar when withdrawal is precipitated with an opioid antagonist, or when food and sugar are removed. When administered a relatively high-dose of the opioid antagonist naloxone (3 mg/kg, s.c.), somatic signs of withdrawal, such as teeth chattering, forepaw tremor, and head shakes are observed (Colantuoni et al., 2002). These animals are also anxious, as measured by reduced time spent on the exposed arm of an elevated plus-maze (Colantuoni et al., 2002) (Fig. 2).
Figure 2.
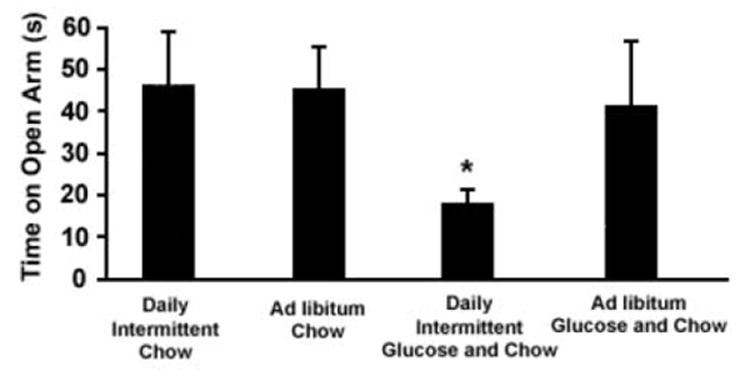
Time spent on the open arms of an elevated plus-maze. Four groups of rats were maintained on their respective diets for one month and then received naloxone (3 mg/kg, s.c.). The Daily Intermittent Glucose and Chow group spent less time on the open arms of the maze. *p<0.05 compared with the Ad libitum Chow group. From Colantuoni et al., 2002.
Behavioral depression has also been found during naloxone-precipitated withdrawal in intermittent sugar-fed rats. In this experiment, rats were given an initial 5-min forced-swim test in which escape (swimming and climbing) and passive (floating) behaviors were measured. Then the rats were divided into four groups that were fed Daily Intermittent Sucrose and Chow, Daily Intermittent Chow, Ad libitum Sucrose and Chow, or Ad libitum Chow for 21 days. On day 22, at the time that the intermittent-fed rats would normally receive their sugar and/or chow, all rats were instead injected with naloxone (3 mg/kg, s.c.) to precipitate withdrawal and were then placed in the water again for another test. In the group that had been fed Daily Intermittent Sucrose and Chow, escape behaviors were significantly suppressed compared with Ad libitum Sucrose and Chow and Ad libitum Chow controls (Fig. 3; Kim, Avena and Hoebel, unpublished). This decrease in escape efforts that were replaced by passive floating suggests the rats were experiencing behavioral depression during withdrawal.
Figure 3.
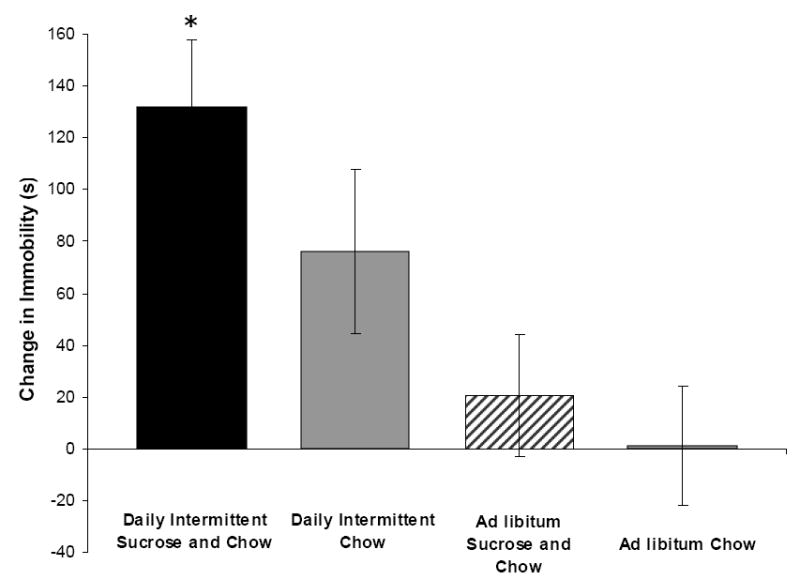
Rats that have been maintained on Daily Intermittent Sucrose and Chow are more immobile than control groups in a forced-swim test during naloxone-precipitated withdrawal. *p<0.05 compared with Ad libitum Sugar and Chow and Ad libitum Chow groups.
Signs of opiate-withdrawal also emerge when all food is removed for 24 h. Again this includes somatic signs such as teeth chattering, forepaw tremor and head shaking (Colantuoni et al., 2002) and anxiety as measured with an elevated plus-maze (Avena, Bocarsly, Rada, Kim and Hoebel, unpublished). Spontaneous withdrawal from the mere remove of sugar has been reported using decreased body temperature as the criterion (Wideman et al., 2005). Also, signs of aggressive behavior have been found during withdrawal of a diet that involves intermittent sugar access (Galic and Persinger, 2002).
4.C. “Craving”: Enhanced responding for sugar following abstinence
As described in Section 2, “craving” in laboratory animals can be defined as enhanced motivation to procure an abused substance (Koob and Le Moal, 2005). After self-administering drugs of abuse and then being forced to abstain, animals often persist in unrewarded operant responding (i.e., resistance to response extinction), and increase their responding for cues previously associated with the drug that grows with time (i.e., incubation) (Bienkowski et al., 2004, Grimm et al., 2001, Lu et al., 2004). Additionally, if the drug becomes available again, animals will take more than they did prior to abstinence (i.e., the “deprivation effect”) (Sinclair and Senter, 1968). This increase in motivation to procure a substance of abuse may contribute to relapse. The power of “craving” is evidenced by results showing that animals will sometimes face adverse consequences to obtain a substance of abuse such as cocaine or alcohol (Deroche-Gamonet et al., 2004, Dickinson et al., 2002, Vanderschuren and Everitt, 2004). These signs in laboratory animals mimic those observed with humans in which the presentation of stimuli previously associated with a drug of abuse increases self-reports of craving and the likelihood of relapse (O’Brien et al., 1977, 1998).
We used the “deprivation effect” paradigm to investigate consumption of sugar after abstinence in rats that had been bingeing on sugar. Following 12-h daily access to sugar, rats lever press for 23% more sugar in a test after 2 wks of abstinence than they ever did before (Fig. 4; Avena et al., 2005). A group with 0.5-h daily access to sucrose did not show the effect. This provides a cogent control group in which rats are familiar with the taste of sucrose, but have not consumed it in a manner that leads to a deprivation effect. The results suggest a change in the motivational impact of sugar that persists throughout two weeks of abstinence, leading to enhanced intake.
Figure 4.
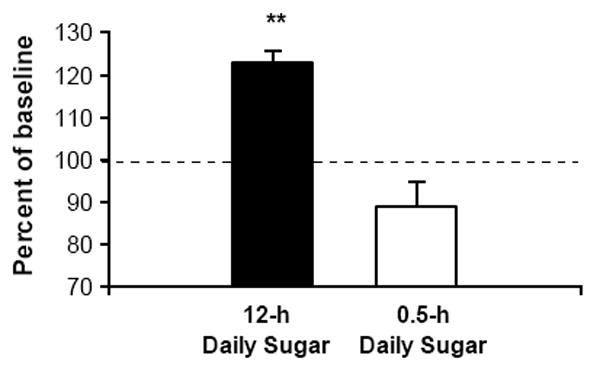
After 14 days of abstinence from sugar, rats that previously had 12-h daily access significantly increased lever pressing for glucose to 123% of pre-abstinence responding, indicating increased motivation for sugar. The group with 0.5-h daily access did not show increased responding after abstinence. **p<0.01. From Avena et al., 2004.
Additionally, like the drugs described above, the motivation to obtain sugar appears to “incubate”, or grow, with the length of abstinence (Shalev et al., 2001). Using operant conditioning, Grimm and colleagues (2005) find that sucrose seeking (lever pressing in extinction and then for a sucrose-paired cue) increases during abstinence in rats after intermittent sugar access for 10 days. Remarkably, responding for the cue was greater after 30 days of sugar abstinence compared with 1 week or 1 day. These results suggest the gradual emergence of long-term changes in the neural circuitry underlying motivation as a result of sugar self-administration and abstinence.
4.D. “Cross-sensitization”: Increased locomotor response to psychostimulants during sugar abstinence
Drug-induced sensitization may play a role in the enhancement of drug self-administration and is implicated as a factor contributing to drug addiction (Robinson and Berridge, 1993). In a typical sensitization experiment, the animal receives a drug daily for about a week, then the procedure stops. However, in the brain there are lasting, even growing, changes apparent a week or more later when a low, challenge dose of the drug results in hyperlocomotion (Kalivas et al., 1992). Additionally, cross-sensitization from one drug to another has been demonstrated with several drugs of abuse, including amphetamine sensitizing rats to cocaine or phencyclidine (Greenberg and Segal, 1985, Kalivas and Weber, 1988, Pierce and Kalivas, 1995, Schenk et al., 1991), cocaine cross-sensitizing with alcohol (Itzhak and Martin, 1999), and heroin with cannabis (Pontieri et al., 2001). Other studies have found this effect with non-drug substances. Behavioral cross-sensitization between cocaine and stress has been demonstrated (Antelman and Caggiula, 1977, Covington and Miczek, 2001, Prasad et al., 1998). Also, increases in food intake (Bakshi and Kelley, 1994) or sexual behaviors (Fiorino and Phillips, 1999, Nocjar and Panksepp, 2002) have been observed in animals with a history of drug sensitization.
We and others have found that Intermittent sugar intake cross-sensitizes with drugs of abuse. Rats sensitized with daily amphetamine injections (3 mg/kg, i.p.) are hyperactive one week later in response to tasting 10% sucrose (Avena and Hoebel, 2003a). Conversely, rats fed Daily Intermittent Sugar and Chow show locomotor cross-sensitization to amphetamine. Specifically, such animals are hyperactive in response to a low, challenge dose of amphetamine (0.5 mg/kg, i.p.) that has no effect on naïve animals, even after 8 days of abstinence from sugar (Fig. 5; Avena and Hoebel, 2003b). Rats maintained on this feeding schedule but administered saline were not hyperactive, nor were rats in control groups (Daily Intermittent Chow, Ad libitum Sugar and Chow, Ad libitum Chow) given the challenge dose of amphetamine. Intermittent sucrose access also cross-sensitizes with cocaine (Gosnell, 2005) and facilitates the development of sensitization to the DA agonist quinpirole (Foley et al., 2006). Thus, results with three different DA agonists from three different laboratories support the theory that the DA system is sensitized by intermittent sugar access, as evidenced by cross-sensitization. This is important since enhanced mesolimbic dopaminergic neurotransmission plays a major role in the behavioral effects of sensitization as well as cross-sensitization (Robinson and Berridge, 1993), and may contribute to addiction and comorbidity with poly-substance abuse.
Figure 5.
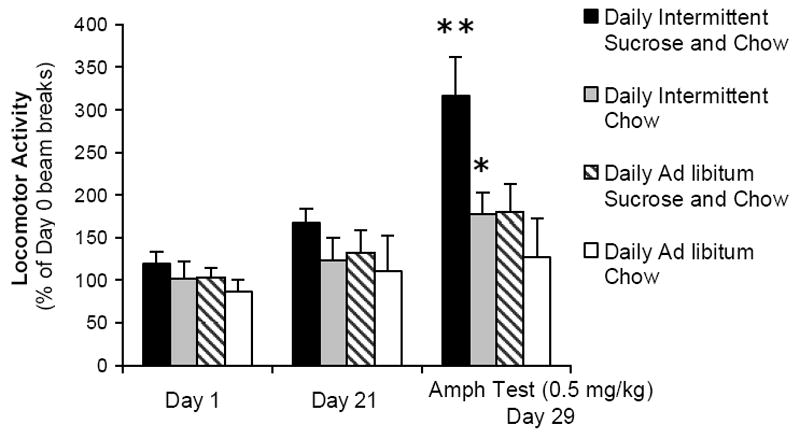
Locomotor activity in a photocell cage plotted as percent of baseline beam breaks on day 0. Rats were maintained for 21 days on the specified diets regimens. Rats maintained on Daily Intermittent Sucrose and Chow were hyperactive nine days later in response to a low dose of amphetamine, compared with control diet groups. **p<0.01. From Avena and Hoebel, 2003.
4.E. “Gateway effect”: Increased alcohol intake during sugar abstinence
Numerous studies have found that sensitization to one drug can lead not only to hyperactivity, but also to subsequent increased intake of another drug or substance (Ellgren et al., 2006, Henningfield et al., 1990, Hubbell et al., 1993, Liguori et al., 1997, Nichols et al., 1991, Piazza et al., 1989, Vezina, 2004, Vezina et al., 2002, Volpicelli et al., 1991). We refer to this phenomenon as “consummatory cross-sensitization”. In the clinical literature, when one drug leads to taking another, this is known as a “gateway effect”. It is particularly noteworthy when a legal drug (e.g. nicotine) acts as a gateway to an illegal drug (e.g. cocaine) (Lai et al., 2000).
Rats maintained on intermittent sugar access and then forced to abstain, subsequently show enhanced intake of 9% alcohol (Avena et al., 2004). This suggests that intermittent access to sugar can be a gateway to alcohol use. Others have shown that animals that prefer sweet-taste will self-administer cocaine at a higher rate (Carroll et al., 2006). As with the locomotor cross-sensitization described above, underlying this behavior are presumably neurochemical alterations in the brain, such as adaptations in DA and perhaps opioid functions.
5. NEUROCHEMICAL SIMILARITIES BETWEEN DRUG SELF-ADMINISTRATION AND INTERMITTENT SUGAR INTAKE
The studies described above suggest that intermittent sugar access can produce numerous behaviors that are similar to those observed in drug-dependent rats. In this section, we describe neurochemical findings that may underlie sugar dependency. To the extent that these brain alterations match the effects of drugs of abuse, it strengthens the case that sugar can resemble a substance of abuse.
5.A. Intermittent sugar intake alters D1, D2 and mu-opioid receptor binding and mRNA expression
Drugs of abuse can alter DA and opioid receptors in the mesolimbic regions of the brain. Pharmacological studies with selective D1, D2 and D3 receptor antagonists and gene knockout studies have revealed that all three receptor subtypes mediate the reinforcing effects drugs of abuse. There is an up-regulation of D1 receptors (Unterwald et al., 1994) and increase in D1 receptor binding (Alburges et al., 1993, Unterwald et al., 2001) in response to cocaine. Conversely, D2 receptor density is lower in NAc of monkeys that have a history of cocaine use (Moore et al., 1998). Drugs of abuse can also produce changes in gene expression of DA receptors. Morphine and cocaine have been shown to decrease accumbens D2 receptor mRNA (Georges et al., 1999, Turchan et al., 1997), and an increase in D3 receptor mRNA (Spangler et al., 2003). These finding with laboratory animals support clinical studies, which have revealed that D2 receptors are down-regulated in cocaine addicts (Volkow et al., 1996a, 1996b, 2006).
Similar changes have been reported with intermittent access to sugar. Autoradiography reveals increased D1 in the NAc and decreased D2 receptor binding in the striatum (Fig. 6; Colantuoni et al., 2001). This was relative to chow-fed rats, so it is not known whether ad libitum sugar would also show this effect. Others have reported a decrease in D2 receptor binding in the NAc of rats with restricted access to sucrose and chow compared with rats fed restriced chow only (Bello et al., 2002). Rats with intermittent sugar and chow access also have decreases in D2 receptor mRNA in the NAc compared with ad libitum chow controls (Spangler et al., 2004). mRNA levels of D3 receptor mRNA in the NAc are increased in the NAc and caudate-putamen.
Figure 6.
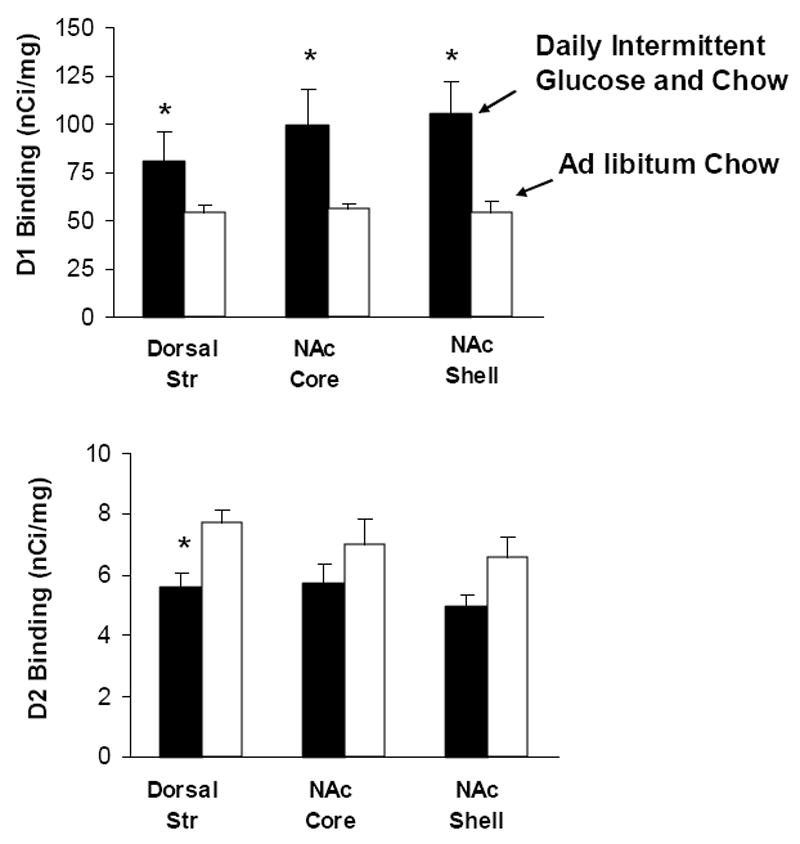
Intermittent sugar access alters DA receptor binding at the level of the striatum. D1 receptor binding (top panel) increases in the NAc core and shell of animals exposed to Daily Intermittent Glucose and Chow (black bars) for 30 days compared with control animals fed chow ad libitum (white bars). D2 receptor binding (bottom panel) decreases in the dorsal striatum in sections from taken the same animals. *p<0.05. From Colantuoni et al., 2001.
Regarding the opioid receptors, mu-receptor binding is increased in response to cocaine and morphine (Bailey et al., 2005, Unterwald et al., 2001, Vigano et al., 2003). Mu-opioid receptor binding is also significantly enhanced after three weeks on the intermittent sugar diet, compared with ad libitum chow. This effect was observed in the accumbens shell, cingulate, hippocampus and locus coeruleus (Colantuoni et al., 2001).
5.B. Intermittent sugar intake alters enkephalin mRNA expression
Enkephalin mRNA in the striatum and the NAc is decreased in response to repeated injections of morphine (Georges et al., 1999, Turchan et al., 1997, Uhl et al., 1988). These changes within opioid systems are similar to those observed in cocaine-dependent human subjects (Zubieta et al., 1996).
Rats with intermittent sugar access also display a significant decrease in enkephalin mRNA, although it is difficult to judge its functional significance (Spangler et al., 2004). This decrease in enkephalin mRNA is consistent with findings observed in rats with limited daily access to a sweet-fat, liquid diet (Kelley et al., 2003). Assuming this decrease in mRNA results in less enkephalin peptide being synthesized and released, it could account for a compensatory increase in mu-opioid receptors, as cited above.
5.C. Daily intermittent sugar intake repeatedly releases dopamine in the accumbens
One of the strongest neurochemical commonalities between intermittent sugar access and drugs of abuse has been found using in vivo microdialysis to measure extracellular DA. The repeated increase in extracellular DA is a hallmark of drugs that are abused. Extracellular DA increases in the NAc in response to both addictive drugs (De Vries and Shippenberg, 2002, Di Chiara and Imperato, 1988, Everitt and Wolf, 2002, Hernandez and Hoebel, 1988, Hurd et al., 1988, Picciotto and Corrigall, 2002, Pothos et al., 1991, Rada et al., 1991a) and drug-associated stimuli (Ito et al., 2000). Unlike drugs of abuse, which exert their effects on DA release each time they are administered (Pothos et al., 1991, Wise et al., 1995), the effect of eating palatable food on DA release wanes with repeated access when the food is no longer novel, unless the animal is food deprived (Bassareo and Di Chiara, 1999, Di Chiara and Tanda, 1997, Rada et al., 2005b). Thus normally feeding is very different than taking drugs because the DA response during feeding is phased out.
However, and this is very important, rats fed daily intermittent sugar and chow apparently release DA every day as measured on days 1, 2 and 21 of access (Fig. 7; Rada et al., 2005b). As controls, rats fed sugar or chow ad libitum, rats with intermittent access to just chow, or rats that taste sugar only two times, develop a blunted DA response as is typical of a food that looses it novelty. These results are supported by findings of alterations in accumbens DA turnover and DA transporter in rats maintained on an intermittent sugar-feeding schedule (Bello et al., 2003, Hajnal and Norgren, 2002). Together, these results suggest that intermittent access to sugar and chow causes recurrent increases in extracellular DA in a manner that is more like a drug of abuse than a food.
Figure 7.
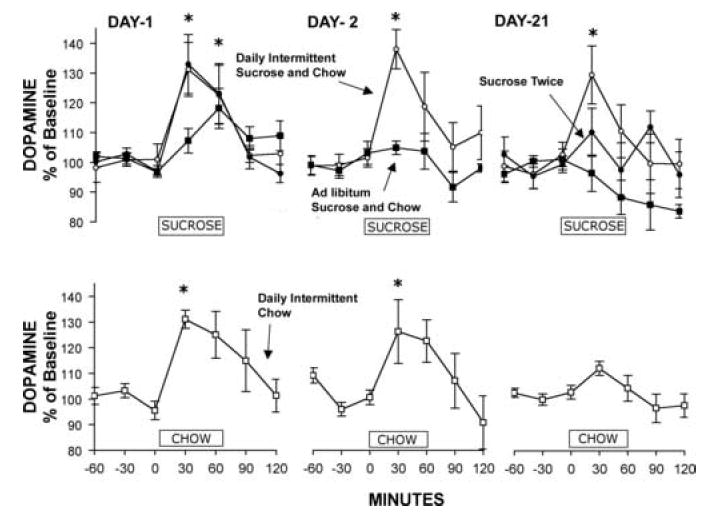
Rats with intermittent access to sugar release DA in response to drinking sucrose for 60 min on day 21. Dopamine, as measured by in vivo microdialysis, increases for the Daily Intermittent Sucrose and Chow rats (open circles) on days 1, 2 and 21; in contrast, DA release was attenuated on day 21 in four control groups as follows: a group that only had 1-h access to sucrose on day 1 and 21 with ad libitum chow in the interim (Sucrose Twice), Ad libitum Sucrose and Chow group, and Daily Intermittent Chow group (bottom panel). The bar on the ordinate indicates the hour (0-60 min) that sucrose or chow was available for the tests. *p<0.05. From Rada et al., 2005.
An interesting question is whether the neurochemical effects observed with intermittent sugar access are due to its postingestive properties or whether the taste of sugar can be sufficient. To investigate orosensory effects of sugar, we used the sham feeding preparation. Rats that are sham feeding with an open gastric fistula can ingest foods but not fully digest them (Smith, 1998). Sham feeding does not completely eliminate post-ingestive effects (Berthoud and Jeanrenaud, 1982, Sclafani and Nissenbaum, 1985), however it does allow the animals to taste the sugar while retaining almost no calories.
The results of sham feeding sugar for the first hour of access each day show that DA is released in the NAc, even after three weeks of daily bingeing, simply due to the taste of sucrose (Avena et al., 2006). Sham feeding does not further enhance the typical sugar-induced DA release. This supports other work showing that the amount of DA release in the NAc is proportional to the sucrose concentration, not the volume consumed (Hajnal et al., 2004).
5.D. Accumbens acetylcholine release is delayed during sugar binges and eliminated during sham feeding
Sham-feeding revealed interesting results with ACh. As described in Section 3.C., accumbens ACh increases in the midst of a meal when feeding slows down and then stops (Mark et al., 1992). One could predict that when an animal takes a very large meal, as with the first meal of a sugar solution and chow, the release of ACh should be delayed until the satiation process begins as reflected in gradual termination of the meal. This is what was observed; ACh release occurred when this initial “binge” meal was drawing to a close (Rada et al., 2005b).
Next we measured ACh release when the animal could take a large meal of sugar while sham feeding. Purging the stomach contents drastically reduced the release of ACh (Avena et al., 2006). This is predictable based on the theory that ACh is normally important for the satiation process (Hoebel et al., 1999, Mark et al., 1992). It also suggests that by purging, one eliminates the ACh response that opposes DA. Thus when “bingeing” on sugar is accompanied by purging, the behavior is reinforced by DA without ACh, which is more like taking a drug and less like normal eating.
5.E. Sugar withdrawal upsets dopamine/acetylcholine balance in the accumbens
Behavioral signs of drug withdrawal are usually accompanied by alterations in DA/ACh balance in the NAc. During withdrawal, DA decreases while ACh is increased. This imbalance has been shown during chemically-induced withdrawal with several drugs of abuse, including morphine, nicotine and alcohol (Rada et al., 1996, 2001, 2004). Mere abstinence from an abused substance is also sufficient to elicit neurochemical signs of withdrawal. For example, rats that are forced to abstain from morphine or alcohol have decreased extracellular DA in the NAc (Acquas and Di Chiara, 1992, Rossetti et al., 1992) and ACh increases during spontaneous morphine withdrawal (Fiserova et al., 1999). While withdrawal from an anxyolitic drug (diazepam) precipitated by a bendodiazepine-receptor antagonist does not lower extracellular DA, it does release accumbens ACh, which may contribute to benzodiazepine dependency (Rada and Hoebel, 2005)
Rats that have intermittent access to sugar and chow show the morphine-like neurochemical imbalance in DA/ACh during withdrawal. This was produced two ways. As shown in Fig. 8, when they are given naloxone to precipitate opioid withdrawal, there is a decrease in accumbens DA release coupled with an increase in ACh release (Colantuoni et al., 2002). The same thing occurs after 36 h of food deprivation (Avena, Bocarsly, Rada, Kim, Hoebel, unpublished). One way to interpret deprivation-induced withdrawal is to suggest that without food to release opioids, the animal suffers the same type of withdrawal seen when the up-regulated mu-opioid receptors are blocked with naloxone.
Figure 8.
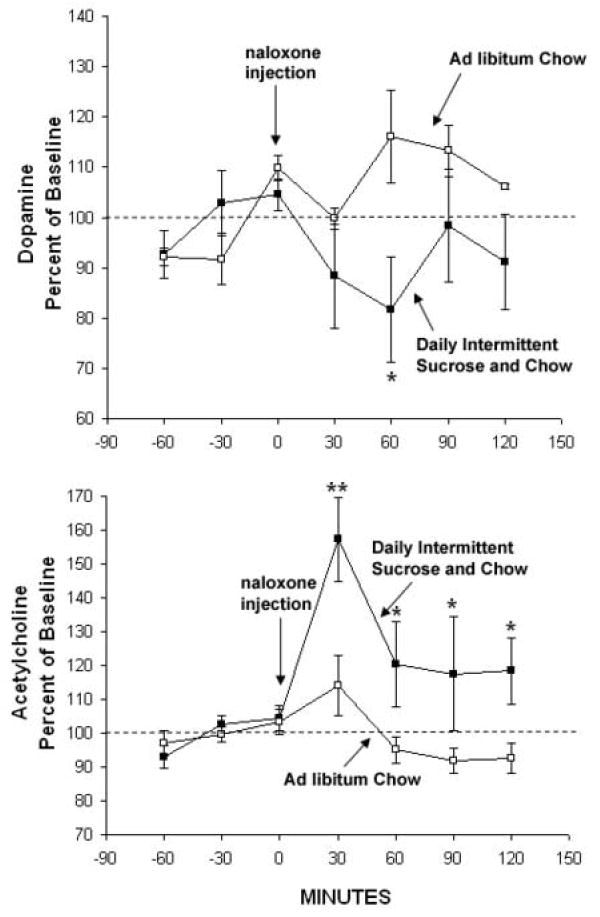
Extracellular DA (upper graph) decreased to 81% of baseline after naloxone injection (3 mg/kg, s.c.) in rats with a history of Daily Intermittent Sucrose and Chow. Acetylcholine (lower graph) increased to 157% in the same intermittent sugar-access rats. No effects were seen in a control group with Ad libitum Chow followed by a naloxone injection. *p<0.05, **p<0.01. From Colantuoni et al., 2002.
6. DISCUSSION AND CLINICAL IMPLICATIONS
Food is not ordinarily like a substance of abuse, but intermittent bingeing and deprivation changes that. Based on the observed behavioral and neurochemical similarities between the effects of intermittent sugar access and drugs of abuse, we suggest that sugar, as common as it is, nonetheless meets the criteria for a substance of abuse and may be “addictive” for some individuals when consumed in a “binge-like” manner. This conclusion is reinforced by the changes in limbic system neurochemistry that are similar for the drugs and for sugar. The effects we observe are smaller in magnitude than those produced by drug of abuse such as cocaine or morphine; however, the fact that these behaviors and neurochemical changes can be elicited with a natural reinforcer is interesting. It is not clear from this animal model if intermittent sugar access can result in neglect of social activities as required by the definition of dependency in the DSM-IV-TR (American Psychiatric Association, 2000). Nor is it known whether rats will continue to self-administer sugar despite physical obstacles, such as enduring pain to obtain sugar, as some rats do for cocaine (Deroche-Gamonet et al., 2004). Nonetheless, the extensive series of experiments revealing similarities between sugar-induced and drug-induced behavior and neurochemistry, as chronicled in Sections 4 and 5, lends credence to the concept of “sugar addiction”, gives precision to its definition, and provides a testable model.
6.A. Bulimia nervosa
The feeding regimen of Daily Intermittent Sugar and Chow shares some aspects of the behavioral pattern of people diagnosed with binge-eating disorder or bulimia. Bulimics often restrict intake early in the day and then binge later in the evening, usually on palatable foods (Drewnowski et al., 1992, Gendall et al., 1997). These patients later purge the food, either by vomiting or laxative use, or in some cases by strenuous exercise (American Psychiatric Association, 2000). Bulimic patients have low β-endorphin levels (Brewerton et al., 1992, Waller et al., 1986), which might foster eating with a preference or craving for sweets. They also have decreased mu-opioid receptor binding in the insula compared with controls, which correlates with recent fasting behavior (Bencherif et al., 2005). This contrasts with the increase observed in rats following a binge. Cyclic bingeing and food deprivation may produce alterations in mu-opioid receptors, which help perpetuate bingeing behavior.
We used the sham feeding preparation to mimic the purging associated with bulimia. The finding described in Section 5.C., that intermittent sugar access repeatedly releases DA in response to the taste of sugar, may be important for understanding the bingeing behaviors associated with bulimia. DA has been implicated in bulimia by comparing it to hypothalamic self-stimulation, which also releases DA without calories (Hoebel et al., 1992). Bulimic patients have low central DA activity as reflected in analysis of DA metabolites in the spinal fluid, which also indicates a role for DA in their abnormal responses to food (Jimerson et al., 1992).
The overall similarlites in behavior and brain adaptations with sugar bingeing and drug intake described above support the theory that obesity and eating disorders, such as bulimia and anorexia, may have properties of an “addiction” in some individuals (Davis and Claridge, 1998, Gillman and Lichtigfeld, 1986, Marrazzi and Luby, 1986, Mercer and Holder, 1997, Riva et al., 2006). The auto-addiction theory proposed that some eating disorders can be an addiction to endogenous opioids (Heubner, 1993, Marrazzi and Luby, 1986, 1990). In support, appetite dysfunctions in the form of binge eating and self-starvation can stimulate endogenous opioid activity (Aravich et al., 1993).
Bulimic patients will binge on excessive amounts of non-caloric sweeteners (Klein et al., 2006), suggesting that they derive benefits from sweet orosensory stimulation. We have shown that purging leaves DA unopposed by satiety-associated ACh in the accumbens (Section 5.D.). This neurochemical state may be conducive to exaggerated binge eating. Moreover, the findings that intermittent sugar intake cross-sensitizes with amphetamine and fosters alcohol intake (Sections 4.D. and 4.E.) may be related to the comorbidity between bulimia and substance abuse (Holderness et al., 1994).
6.B. Obesity
Sugar and obesity
Obesity is one of the leading preventable causes of death in the US (Mokdad et al., 2004). Several studies have correlated the rise in the incidence of obesity with an increase in sugar consumption (Bray et al., 1992, Elliott et al., 2002, Howard and Wylie-Rosett, 2002, Ludwig et al., 2001). The US Department of Agriculture has reported that per capita soft-drink consumption has increased by almost 500% in the past 50 years (Putnam and Allhouse, 1999). Sugar intake may lead to an increased number of and/or affinity for opioid receptors, which in turn leads to further ingestion of sugar and may contribute to obesity (Fullerton et al., 1985). Indeed, rats maintained on the diet of intermittent sugar access show opioid receptor changes (Section 5.A.); however, after one month on the diet using 10% sucrose or 25% glucose, these animals do not become overweight (Colantuoni et al., 2001, Avena and Hoebel, 2003b), although others have reported a metabolic syndrome (Toida et al., 1996), a loss of fuel efficiency (Levine et al., 2003) and an increase in body weight in rats fed sucrose (Bock et al., 1995, Kawasaki et al., 2005) and glucose (Wideman et al., 2005). Most studies of sugar intake and body weight do not use a binge-inducing diet, and the translation to human obesity is complex (Levine et al., 2003). As described in Section 4.A., it appears that rats in our model compensate for sucrose or glucose calories by decreasing chow intake (Avena, Bocarsly, Rada, Kim and Hoebel, unpublished). They gain weight at a normal rate (Colantuoni et al., 2002). This may not be true of all sugars.
Fructose is a unique sweetener that has different metabolic effects on the body than glucose or sucrose. Fructose is absorbed further down the intestine, and whereas circulating glucose releases insulin from the pancreas (Sato et al., 1996, Vilsboll et al., 2003), fructose stimulates insulin synthesis but does not release it (Curry, 1989, Le and Tappy, 2006, Sato et al., 1996). Insulin modifies food intake by inhibiting eating (Schwartz et al., 2000) and by increasing leptin release (Saad et al., 1998), which also can inhibit food intake. Meals of high-fructose corn syrup can reduce circulating insulin and leptin levels (Teff et al., 2004), contributing to increased body weight. Thus, fructose intake might not result in the degree of satiety that would normally ensue with an equally caloric meal of glucose or sucrose. Since high-fructose corn syrup has become a major constituent in the American diet (Bray et al., 2004) and lacks some effects on insulin and leptin, it may be a potential agent for producing obesity when given intermittently to rats. Whether or not signs of dependency on fructose are apparent when it is offered intermittently has yet to be determined. However, based on our results showing that sweet taste is sufficient to elicit the repeated release of DA in the NAc (see Section 5.C.), we hypothesize that any sweet taste consumed in a binge-like manner is a candidate for producing signs of dependence.
Fat and obesity
While we have chosen to focus on sugar, the question arises as to whether non-sweet, palatable foods could produce signs or dependence. The evidence is mixed. It appears that some signs of dependence are apparent with fat, while others have not been shown. Fat bingeing in rats occurs with intermittent access to pure fat (vegetable shortening), sweet-fat cookies (Boggiano et al., 2005, Corwin, 2006), or sweet-fat chow (Berner, Avena and Hoebel, unpublished). Repeated, intermittent access to oil releases DA in the NAc (Liang et al., 2006). Like sugar, bingeing on a fat-rich diet is known to affect the opioid system in the accumbens by decreasing enkephalin mRNA, an effect that is not observed with acute access (Kelley et al., 2003). Also, treatment with baclofen (GABA-B agonist), which reduces drug intake, also reduces binge eating of fat (Buda-Levin et al., 2005).
This all implies that fat dependency is a real possibility, but withdrawal from fat-bingeing is not as apparent as it is with sugar. Le Magnen (1990) noted naloxone could precipitate withdrawal in rats on a cafeteria-style diet, which contains a variety of fat- and sugar-rich foods (e.g., cheese, cookies, chocolate chips). However, we have not observed signs of naloxone-precipitated or spontaneous withdrawal in rats fed pure fat (vegetable shortening) or a sugar-fat combination, nor has such a result been published by others. Further studies are needed to fully understand the differences between sugar and fat bingeing and their subsequent effects on behavior. Just as different classes of drugs (e.g., dopamine agonists vs. opiates) have specific behavioral and physiological withdrawal signs, it may be that different macronutrients may also produce specific withdrawal signs. Since craving of fat or cross-sensitization between fat intake and drugs of abuse has yet to be documented in animals, sugar is currently the only palatable substance for which bingeing, withdrawal, abstinence-induced enhanced motivation and cross-sensitization have all been demonstrated (Sections 4 and 5).
Brain imaging
Recent findings using positron emission tomography (PET) and functional magnetic resonance imaging (fMRI) in humans have supported the idea that aberrant eating behaviors, including those observed in obesity, may have similarities to drug dependence. Craving-related changes in fMRI signal have been identified in response to palatable foods, similar to drug craving. This overlap occurred in the hippocampus, insula, and caudate (Pelchat et al., 2004). Similarly, PET scans reveal that obese subjects show a reduction in striatal D2 receptor availability that is associated with the body weight of the subject (Wang et al., 2004b). This decrease in D2 receptors in obese subjects is similar in magnitude to the reductions reported in drug-addicted subjects (Wang et al., 2001). The involvement of the DA system in reward and reinforcement has led to the hypothesis that alterations in DA activity in obese subjects dispose them to excessive use of food. Exposure to especially palatable foods, such as cake and ice cream, activates the several brain regions including the anterior insula and right orbitofrontal cortex (Wang et al., 2004a), which may underlie the motivation to procure food (Rolls, 2006).
7. CONCLUSION
From an evolutionary perspective, it is in the best interest of humans to have an inherent desire for food for survival. However, this desire may go awry, and certain people, including some obese and bulimic patients in particular, may develop an unhealthy dependence on palatable food that interferes with well-being. The concept of “food addiction” materialized in the diet industry on the basis of subjective reports, clinical accounts and case studies described in self-help books. The rise in obesity, coupled with the emergence of scientific findings of parallels between drugs of abuse and palatable foods has given credibility to this idea. The reviewed evidence supports the theory that, in some circumstances, intermittent access to sugar can lead to behavior and neurochemical changes that resemble the effects of a substance of abuse. According to the evidence in rats, intermittent access to sugar and chow is capable of producing a “dependency”. This was operationally defined by tests for bingeing, withdrawal, craving and cross-sensitization to amphetamine and alcohol. The correspondence to some people with binge eating disorder or bulimia is striking, but whether or not it is a good idea to call this a “food addiction” in people is both a scientific and societal question that has yet to be answered. What this review demonstrates is that rats with intermittent access to food and a sugar solution can show both a constellation of behaviors and parallel brain changes that are characteristic of rats that voluntarily self-administer addictive drugs. In the aggregrate, this is evidence that sugar can be addictive.
Acknowledgments
This reseach was supported by USPHS grant MH-65024 (B.G.H.), DA-10608 (B.G.H.), DA-16458 (fellowship to N.M.A) and the Lane Foundation.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Acquas E, Carboni E, Di Chiara G. Profound depression of mesolimbic dopamine release after morphine withdrawal in dependent rats. Eur J Pharmacol. 1991;193:133–134. doi: 10.1016/0014-2999(91)90214-b. [DOI] [PubMed] [Google Scholar]
- Acquas E, Di Chiara G. Depression of mesolimbic dopamine transmission and sensitization to morphine during opiate abstinence. J Neurochem. 1992;58:1620–1625. doi: 10.1111/j.1471-4159.1992.tb10033.x. [DOI] [PubMed] [Google Scholar]
- Ahmed SH, Koob GF. Transition from moderate to excessive drug intake: change in hedonic set point. Science. 1998;282:298–300. doi: 10.1126/science.282.5387.298. [DOI] [PubMed] [Google Scholar]
- Alburges ME, Narang N, Wamsley JK. Alterations in the dopaminergic receptor system after chronic administration of cocaine. Synapse. 1993;14:314–323. doi: 10.1002/syn.890140409. [DOI] [PubMed] [Google Scholar]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders Fouth Edition Text Revision (DSM-IV-TR) American Psychiatric Association; Washington, DC: 2000. [Google Scholar]
- Antelman SM, Caggiula AR. Norepinephrine-dopamine interactions and behavior. Science. 1977;195:646–653. doi: 10.1126/science.841304. [DOI] [PubMed] [Google Scholar]
- Antelman SM, Caggiula AR. Oscillation follows drug sensitization: implications. Crit Rev Neurobiol. 1996;10:101–117. doi: 10.1615/critrevneurobiol.v10.i1.50. [DOI] [PubMed] [Google Scholar]
- Appleton N. Lick the sugar habit. Nancy Appleton; Santa Monica: 1996. [Google Scholar]
- Aravich PF, Rieg TS, Lauterio TJ, Doerries LE. Beta-endorphin and dynorphin abnormalities in rats subjected to exercise and restricted feeding: relationship to anorexia nervosa? Brain Res. 1993;622:1–8. doi: 10.1016/0006-8993(93)90794-n. [DOI] [PubMed] [Google Scholar]
- Ary M, Chesarek W, Sorensen SM, Lomax P. Naltrexone-induced hypothermia in the rat. Eur J Pharmacol. 1976;39:215–220. doi: 10.1016/0014-2999(76)90129-1. [DOI] [PubMed] [Google Scholar]
- Avena NM, Carrillo CA, Needham L, Leibowitz SF, Hoebel BG. Sugar-dependent rats show enhanced intake of unsweetened ethanol. Alcohol. 2004;34:203–209. doi: 10.1016/j.alcohol.2004.09.006. [DOI] [PubMed] [Google Scholar]
- Avena NM, Hoebel BG. Amphetamine-sensitized rats show sugar-induced hyperactivity (cross-sensitization) and sugar hyperphagia. Pharmacol Biochem Behav. 2003a;74:635–639. doi: 10.1016/s0091-3057(02)01050-x. [DOI] [PubMed] [Google Scholar]
- Avena NM, Hoebel BG. A diet promoting sugar dependency causes behavioral cross-sensitization to a low dose of amphetamine. Neuroscience. 2003b;122:17–20. doi: 10.1016/s0306-4522(03)00502-5. [DOI] [PubMed] [Google Scholar]
- Avena NM, Long KA, Hoebel BG. Sugar-dependent rats show enhanced responding for sugar after abstinence: evidence of a sugar deprivation effect. Physiol Behav. 2005;84:359–362. doi: 10.1016/j.physbeh.2004.12.016. [DOI] [PubMed] [Google Scholar]
- Avena NM, Rada P, Moise N, Hoebel BG. Sucrose sham feeding on a binge schedule releases accumbens dopamine repeatedly and eliminates the acetylcholine satiety response. Neuroscience. 2006;139:813–820. doi: 10.1016/j.neuroscience.2005.12.037. [DOI] [PubMed] [Google Scholar]
- Bailey A, Gianotti R, Ho A, Kreek MJ. Persistent upregulation of mu-opioid, but not adenosine, receptors in brains of long-term withdrawn escalating dose “binge” cocaine-treated rats. Synapse. 2005;57:160–166. doi: 10.1002/syn.20168. [DOI] [PubMed] [Google Scholar]
- Bakshi VP, Kelley AE. Sensitization and conditioning of feeding following multiple morphine microinjections into the nucleus accumbens. Brain Res. 1994;648:342–346. doi: 10.1016/0006-8993(94)91139-8. [DOI] [PubMed] [Google Scholar]
- Bals-Kubik R, Herz A, Shippenberg TS. Evidence that the aversive effects of opioid antagonists and kappa-agonists are centrally mediated. Psychopharmacology (Berl) 1989;98:203–206. doi: 10.1007/BF00444692. [DOI] [PubMed] [Google Scholar]
- Bancroft J, Vukadinovic Z. Sexual addiction, sexual compulsivity, sexual impulsivity, or what? Toward a theoretical model. J Sex Res. 2004;41:225–234. doi: 10.1080/00224490409552230. [DOI] [PubMed] [Google Scholar]
- Bassareo V, Di Chiara G. Differential influence of associative and nonassociative learning mechanisms on the responsiveness of prefrontal and accumbal dopamine transmission to food stimuli in rats fed ad libitum. J Neurosci. 1997;17:851–861. doi: 10.1523/JNEUROSCI.17-02-00851.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bassareo V, Di Chiara G. Modulation of feeding-induced activation of mesolimbic dopamine transmission by appetitive stimuli and its relation to motivational state. Eur J Neurosci. 1999;11:4389–4397. doi: 10.1046/j.1460-9568.1999.00843.x. [DOI] [PubMed] [Google Scholar]
- Bello NT, Lucas LR, Hajnal A. Repeated sucrose access influences dopamine D2 receptor density in the striatum. Neuroreport. 2002;13:1575–1578. doi: 10.1097/00001756-200208270-00017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bello NT, Sweigart KL, Lakoski JM, Norgren R, Hajnal A. Restricted feeding with scheduled sucrose access results in an upregulation of the rat dopamine transporter. Am J Physiol Regul Integr Comp Physiol. 2003;284:R1260–1268. doi: 10.1152/ajpregu.00716.2002. [DOI] [PubMed] [Google Scholar]
- Bencherif B, Guarda AS, Colantuoni C, Ravert HT, Dannals RF, Frost JJ. Regional mu-opioid receptor binding in insular cortex is decreased in bulimia nervosa and correlates inversely with fasting behavior. J Nucl Med. 2005;46:1349–1351. [PubMed] [Google Scholar]
- Berridge KC. Food reward: brain substrates of wanting and liking. Neurosci Biobehav Rev. 1996;20:1–25. doi: 10.1016/0149-7634(95)00033-b. [DOI] [PubMed] [Google Scholar]
- Berridge KC, Robinson TE. What is the role of dopamine in reward: hedonic impact, reward learning, or incentive salience? Brain Res Brain Res Rev. 1998;28:309–369. doi: 10.1016/s0165-0173(98)00019-8. [DOI] [PubMed] [Google Scholar]
- Berthoud HR, Jeanrenaud B. Sham feeding-induced cephalic phase insulin release in the rat. Am J Physiol. 1982;242:E280–285. doi: 10.1152/ajpendo.1982.242.4.E280. [DOI] [PubMed] [Google Scholar]
- Bienkowski P, Rogowski A, Korkosz A, Mierzejewski P, Radwanska K, Kaczmarek L, Bogucka-Bonikowska A, Kostowski W. Time-dependent changes in alcohol-seeking behaviour during abstinence. Eur Neuropsychopharmacol. 2004;14:355–360. doi: 10.1016/j.euroneuro.2003.10.005. [DOI] [PubMed] [Google Scholar]
- Blomqvist O, Ericson M, Johnson DH, Engel JA, Soderpalm B. Voluntary ethanol intake in the rat: effects of nicotinic acetylcholine receptor blockade or subchronic nicotine treatment. Eur J Pharmacol. 1996;314:257–267. doi: 10.1016/s0014-2999(96)00583-3. [DOI] [PubMed] [Google Scholar]
- Bock BC, Kanarek RB, Aprille JR. Mineral content of the diet alters sucrose-induced obesity in rats. Physiol Behav. 1995;57:659–668. doi: 10.1016/0031-9384(94)00312-2. [DOI] [PubMed] [Google Scholar]
- Boggiano MM, Chandler PC, Viana JB, Oswald KD, Maldonado CR, Wauford PK. Combined dieting and stress evoke exaggerated responses to opioids in binge-eating rats. Behav Neurosci. 2005;119:1207–1214. doi: 10.1037/0735-7044.119.5.1207. [DOI] [PubMed] [Google Scholar]
- Bozarth MA, Wise RA. Intracranial self-administration of morphine into the ventral tegmental area in rats. Life Sci. 1981;28:551–555. doi: 10.1016/0024-3205(81)90148-x. [DOI] [PubMed] [Google Scholar]
- Bozarth MA, Wise RA. Toxicity associated with long-term intravenous heroin and cocaine self-administration in the rat. JAMA. 1985;254:81–83. [PubMed] [Google Scholar]
- Bozarth MA, Wise RA. Involvement of the ventral tegmental dopamine system in opioid and psychomotor stimulant reinforcement. NIDA Res Monogr. 1986;67:190–196. [PubMed] [Google Scholar]
- Bray GA, Nielsen SJ, Popkin BM. Consumption of high-fructose corn syrup in beverages may play a role in the epidemic of obesity. Am J Clin Nutr. 2004;79:537–543. doi: 10.1093/ajcn/79.4.537. [DOI] [PubMed] [Google Scholar]
- Bray GA, York B, DeLany J. A survey of the opinions of obesity experts on the causes and treatment of obesity. Am J Clin Nutr. 1992;55:151S–154S. doi: 10.1093/ajcn/55.1.151s. [DOI] [PubMed] [Google Scholar]
- Brewerton TD, Lydiard RB, Laraia MT, Shook JE, Ballenger JC. CSF beta-endorphin and dynorphin in bulimia nervosa. Am J Psychiatry. 1992;149:1086–1090. doi: 10.1176/ajp.149.8.1086. [DOI] [PubMed] [Google Scholar]
- Buda-Levin A, Wojnicki FH, Corwin RL. Baclofen reduces fat intake under binge-type conditions. Physiol Behav. 2005;86:176–184. doi: 10.1016/j.physbeh.2005.07.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carr KD. Chronic food restriction: Enhancing effects on drug reward and striatal cell signaling. Physiol Behav. 2006 doi: 10.1016/j.physbeh.2006.09.021. [DOI] [PubMed] [Google Scholar]
- Carroll ME. The role of food deprivation in the maintenance and reinstatement of cocaine-seeking behavior in rats. Drug Alcohol Depend. 1985;16:95–109. doi: 10.1016/0376-8716(85)90109-7. [DOI] [PubMed] [Google Scholar]
- Carroll ME, Anderson MM, Morgan AD. Regulation of intravenous cocaine self-administration in rats selectively bred for high (HiS) and low (LoS) saccharin intake. Psychopharmacology (Berl) 2006 doi: 10.1007/s00213-006-0600-3. [DOI] [PubMed] [Google Scholar]
- Chau D, Rada PV, Kosloff RA, Hoebel BG. Cholinergic, M1 receptors in the nucleus accumbens mediate behavioral depression. A possible downstream target for fluoxetine. Ann N Y Acad Sci. 1999;877:769–774. doi: 10.1111/j.1749-6632.1999.tb09320.x. [DOI] [PubMed] [Google Scholar]
- Cheer JF, Wassum KM, Heien ML, Phillips PE, Wightman RM. Cannabinoids enhance subsecond dopamine release in the nucleus accumbens of awake rats. J Neurosci. 2004;24:4393–4400. doi: 10.1523/JNEUROSCI.0529-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colantuoni C, Rada P, McCarthy J, Patten C, Avena NM, Chadeayne A, Hoebel BG. Evidence that intermittent, excessive sugar intake causes endogenous opioid dependence. Obes Res. 2002;10:478–488. doi: 10.1038/oby.2002.66. [DOI] [PubMed] [Google Scholar]
- Colantuoni C, Schwenker J, McCarthy J, Rada P, Ladenheim B, Cadet JL, Schwartz GJ, Moran TH, Hoebel BG. Excessive sugar intake alters binding to dopamine and mu-opioid receptors in the brain. Neuroreport. 2001;12:3549–3552. doi: 10.1097/00001756-200111160-00035. [DOI] [PubMed] [Google Scholar]
- Comings DE, Gade-Andavolu R, Gonzalez N, Wu S, Muhleman D, Chen C, Koh P, Farwell K, Blake H, Dietz G, MacMurray JP, Lesieur HR, Rugle LJ, Rosenthal RJ. The additive effect of neurotransmitter genes in pathological gambling. Clin Genet. 2001;60:107–116. doi: 10.1034/j.1399-0004.2001.600204.x. [DOI] [PubMed] [Google Scholar]
- Corwin RL. Bingeing rats: a model of intermittent excessive behavior? Appetite. 2006;46:11–15. doi: 10.1016/j.appet.2004.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Covington HE, Miczek KA. Repeated social-defeat stress, cocaine or morphine. Effects on behavioral sensitization and intravenous cocaine self-administration “binges”. Psychopharmacology (Berl) 2001;158:388–398. doi: 10.1007/s002130100858. [DOI] [PubMed] [Google Scholar]
- Curry DL. Effects of mannose and fructose on the synthesis and secretion of insulin. Pancreas. 1989;4:2–9. doi: 10.1097/00006676-198902000-00002. [DOI] [PubMed] [Google Scholar]
- Davis C, Claridge G. The eating disorders as addiction: a psychobiological perspective. Addict Behav. 1998;23:463–475. doi: 10.1016/s0306-4603(98)00009-4. [DOI] [PubMed] [Google Scholar]
- De Vries TJ, Shippenberg TS. Neural systems underlying opiate addiction. J Neurosci. 2002;22:3321–3325. doi: 10.1523/JNEUROSCI.22-09-03321.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Witte P, Pinto E, Ansseau M, Verbanck P. Alcohol and withdrawal: from animal research to clinical issues. Neurosci Biobehav Rev. 2003;27:189–197. doi: 10.1016/s0149-7634(03)00030-7. [DOI] [PubMed] [Google Scholar]
- Deas D, May MP, Randall C, Johnson N, Anton R. Naltrexone treatment of adolescent alcoholics: an open-label pilot study. J Child Adolesc Psychopharmacol. 2005;15:723–728. doi: 10.1089/cap.2005.15.723. [DOI] [PubMed] [Google Scholar]
- Deneau G, Yanagita T, Seevers MH. Self-administration of psychoactive substances by the monkey. Psychopharmacologia. 1969;16:30–48. doi: 10.1007/BF00405254. [DOI] [PubMed] [Google Scholar]
- Deroche-Gamonet V, Belin D, Piazza PV. Evidence for addiction-like behavior in the rat. Science. 2004;305:1014–1017. doi: 10.1126/science.1099020. [DOI] [PubMed] [Google Scholar]
- DesMaisons K. Your last diet!: The sugar addict’s weight-loss plan. Random House; Toronto: 2001. [Google Scholar]
- Di Chiara G, Imperato A. Preferential stimulation of dopamine release in the nucleus accumbens by opiates, alcohol, and barbiturates: studies with transcerebral dialysis in freely moving rats. Ann N Y Acad Sci. 1986;473:367–381. doi: 10.1111/j.1749-6632.1986.tb23629.x. [DOI] [PubMed] [Google Scholar]
- Di Chiara G, Imperato A. Drugs abused by humans preferentially increase synaptic dopamine concentrations in the mesolimbic system of freely moving rats. Proc Natl Acad Sci U S A. 1988;85:5274–5278. doi: 10.1073/pnas.85.14.5274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di Chiara G, Tanda G. Blunting of reactivity of dopamine transmission to palatable food: a biochemical marker of anhedonia in the CMS model? Psychopharmacology (Berl) 1997;134:351–353. doi: 10.1007/s002130050465. [DOI] [PubMed] [Google Scholar]
- Dickinson A, Wood N, Smith JW. Alcohol seeking by rats: action or habit? Q J Exp Psychol B. 2002;55:331–348. doi: 10.1080/0272499024400016. [DOI] [PubMed] [Google Scholar]
- Drewnowski A, Krahn DD, Demitrack MA, Nairn K, Gosnell BA. Taste responses and preferences for sweet high-fat foods: evidence for opioid involvement. Physiol Behav. 1992;51:371–379. doi: 10.1016/0031-9384(92)90155-u. [DOI] [PubMed] [Google Scholar]
- Dum J, Gramsch C, Herz A. Activation of hypothalamic beta-endorphin pools by reward induced by highly palatable food. Pharmacol Biochem Behav. 1983;18:443–447. doi: 10.1016/0091-3057(83)90467-7. [DOI] [PubMed] [Google Scholar]
- Ellgren M, Spano SM, Hurd YL. Adolescent cannabis exposure alters opiate intake and opioid limbic neuronal populations in adult rats. Neuropsychopharmacology. 2006 doi: 10.1038/sj.npp.1301127. Epub ahead of print. [DOI] [PubMed] [Google Scholar]
- Elliott SS, Keim NL, Stern JS, Teff K, Havel PJ. Fructose, weight gain, and the insulin resistance syndrome. Am J Clin Nutr. 2002;76:911–922. doi: 10.1093/ajcn/76.5.911. [DOI] [PubMed] [Google Scholar]
- Espejo EF, Stinus L, Cador M, Mir D. Effects of morphine and naloxone on behaviour in the hot plate test: an ethopharmacological study in the rat. Psychopharmacology (Berl) 1994;113:500–510. doi: 10.1007/BF02245230. [DOI] [PubMed] [Google Scholar]
- Everitt BJ, Wolf ME. Psychomotor stimulant addiction: a neural systems perspective. J Neurosci. 2002;22:3312–3320. doi: 10.1523/JNEUROSCI.22-09-03312.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrario CR, Robinson TE. Amphetamine pretreatment accelerates the subsequent escalation of cocaine self-administration behavior. Eur Neuropsychopharmacol. 2007;17:352–357. doi: 10.1016/j.euroneuro.2006.08.005. [DOI] [PubMed] [Google Scholar]
- File SE, Andrews N. Low but not high doses of buspirone reduce the anxiogenic effects of diazepam withdrawal. Psychopharmacology (Berl) 1991;105:578–582. doi: 10.1007/BF02244384. [DOI] [PubMed] [Google Scholar]
- File SE, Lippa AS, Beer B, Lippa MT. Unit 8.4 Animal tests of anxiety. In: Crawley JN, et al., editors. Current Protocols in Neuroscience. John Wiley & Sons, Inc.; Indianapolis: 2004. [DOI] [PubMed] [Google Scholar]
- Finlayson G, King N, Blundell JE. Is it possible to dissociate ‘liking’ and ‘wanting’ for foods in humans? A novel experimental procedure. Physiol Behav. 2007;90:36–42. doi: 10.1016/j.physbeh.2006.08.020. [DOI] [PubMed] [Google Scholar]
- Fiorino DF, Phillips AG. Facilitation of sexual behavior and enhanced dopamine efflux in the nucleus accumbens of male rats after D-amphetamine-induced behavioral sensitization. J Neurosci. 1999;19:456–463. doi: 10.1523/JNEUROSCI.19-01-00456.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fiserova M, Consolo S, Krsiak M. Chronic morphine induces long-lasting changes in acetylcholine release in rat nucleus accumbens core and shell: an in vivo microdialysis study. Psychopharmacology (Berl) 1999;142:85–94. doi: 10.1007/s002130050866. [DOI] [PubMed] [Google Scholar]
- Foley KA, Fudge MA, Kavaliers M, Ossenkopp KP. Quinpirole-induced behavioral sensitization is enhanced by prior scheduled exposure to sucrose: A multivariable examination of locomotor activity. Behav Brain Res. 2006;167:49–56. doi: 10.1016/j.bbr.2005.08.015. [DOI] [PubMed] [Google Scholar]
- Foster J, Brewer C, Steele T. Naltrexone implants can completely prevent early (1-month) relapse after opiate detoxification: a pilot study of two cohorts totalling 101 patients with a note on naltrexone blood levels. Addict Biol. 2003;8:211–217. doi: 10.1080/1355621031000117446. [DOI] [PubMed] [Google Scholar]
- Fullerton DT, Getto CJ, Swift WJ, Carlson IH. Sugar, opioids and binge eating. Brain Res Bull. 1985;14:673–680. doi: 10.1016/0361-9230(85)90117-0. [DOI] [PubMed] [Google Scholar]
- Galic MA, Persinger MA. Voluminous sucrose consumption in female rats: increased “nippiness” during periods of sucrose removal and possible oestrus periodicity. Psychol Rep. 2002;90:58–60. doi: 10.2466/pr0.2002.90.1.58. [DOI] [PubMed] [Google Scholar]
- Gendall KA, Sullivan PE, Joyce PR, Carter FA, Bulik CM. The nutrient intake of women with bulimia nervosa. Int J Eat Disord. 1997;21:115–127. doi: 10.1002/(sici)1098-108x(199703)21:2<115::aid-eat2>3.0.co;2-o. [DOI] [PubMed] [Google Scholar]
- Georges F, Stinus L, Bloch B, Le Moine C. Chronic morphine exposure and spontaneous withdrawal are associated with modifications of dopamine receptor and neuropeptide gene expression in the rat striatum. Eur J Neurosci. 1999;11:481–490. doi: 10.1046/j.1460-9568.1999.00462.x. [DOI] [PubMed] [Google Scholar]
- Gerber GJ, Wise RA. Pharmacological regulation of intravenous cocaine and heroin self-administration in rats: a variable dose paradigm. Pharmacol Biochem Behav. 1989;32:527–531. doi: 10.1016/0091-3057(89)90192-5. [DOI] [PubMed] [Google Scholar]
- Gessa GL, Muntoni F, Collu M, Vargiu L, Mereu G. Low doses of ethanol activate dopaminergic neurons in the ventral tegmental area. Brain Res. 1985;348:201–203. doi: 10.1016/0006-8993(85)90381-6. [DOI] [PubMed] [Google Scholar]
- Gillman MA, Lichtigfeld FJ. The opioids, dopamine, cholecystokinin, and eating disorders. Clin Neuropharmacol. 1986;9:91–97. doi: 10.1097/00002826-198602000-00011. [DOI] [PubMed] [Google Scholar]
- Glass MJ, Billington CJ, Levine AS. Opioids and food intake: distributed functional neural pathways? Neuropeptides. 1999;33:360–368. doi: 10.1054/npep.1999.0050. [DOI] [PubMed] [Google Scholar]
- Glick SD, Shapiro RM, Drew KL, Hinds PA, Carlson JN. Differences in spontaneous and amphetamine-induced rotational behavior, and in sensitization to amphetamine, among Sprague-Dawley derived rats from different sources. Physiol Behav. 1986;38:67–70. doi: 10.1016/0031-9384(86)90133-2. [DOI] [PubMed] [Google Scholar]
- Glimcher PW, Giovino AA, Hoebel BG. Neurotensin self-injection in the ventral tegmental area. Brain Res. 1987;403:147–150. doi: 10.1016/0006-8993(87)90134-x. [DOI] [PubMed] [Google Scholar]
- Glimcher PW, Giovino AA, Margolin DH, Hoebel BG. Endogenous opiate reward induced by an enkephalinase inhibitor, thiorphan, injected into the ventral midbrain. Behav Neurosci. 1984;98:262–268. doi: 10.1037//0735-7044.98.2.262. [DOI] [PubMed] [Google Scholar]
- Glowa JR, Rice KC, Matecka D, Rothman RB. Phentermine/fenfluramine decreases cocaine self-administration in rhesus monkeys. Neuroreport. 1997;8:1347–1351. doi: 10.1097/00001756-199704140-00006. [DOI] [PubMed] [Google Scholar]
- Gosnell BA. Sucrose intake enhances behavioral sensitization produced by cocaine. Brain Res. 2005;1031:194–201. doi: 10.1016/j.brainres.2004.10.037. [DOI] [PubMed] [Google Scholar]
- Greenberg BD, Segal DS. Acute and chronic behavioral interactions between phencyclidine (PCP) and amphetamine: evidence for a dopaminergic role in some PCP-induced behaviors. Pharmacol Biochem Behav. 1985;23:99–105. doi: 10.1016/0091-3057(85)90137-6. [DOI] [PubMed] [Google Scholar]
- Grimm JW, Fyall AM, Osincup DP. Incubation of sucrose craving: effects of reduced training and sucrose pre-loading. Physiol Behav. 2005;84:73–79. doi: 10.1016/j.physbeh.2004.10.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grimm JW, Hope BT, Wise RA, Shaham Y. Neuroadaptation. Incubation of cocaine craving after withdrawal. Nature. 2001;412:141–142. doi: 10.1038/35084134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haber SN, Lu W. Distribution of preproenkephalin messenger RNA in the basal ganglia and limbic-associated regions of the monkey telencephalon. Neuroscience. 1995;65:417–429. doi: 10.1016/0306-4522(94)00490-v. [DOI] [PubMed] [Google Scholar]
- Hajnal A, Mark GP, Rada PV, Lenard L, Hoebel BG. Norepinephrine microinjections in the hypothalamic paraventricular nucleus increase extracellular dopamine and decrease acetylcholine in the nucleus accumbens: relevance to feeding reinforcement. J Neurochem. 1997;68:667–674. doi: 10.1046/j.1471-4159.1997.68020667.x. [DOI] [PubMed] [Google Scholar]
- Hajnal A, Norgren R. Repeated access to sucrose augments dopamine turnover in the nucleus accumbens. Neuroreport. 2002;13:2213–2216. doi: 10.1097/00001756-200212030-00010. [DOI] [PubMed] [Google Scholar]
- Hajnal A, Smith GP, Norgren R. Oral sucrose stimulation increases accumbens dopamine in the rat. Am J Physiol Regul Integr Comp Physiol. 2004;286:R31–R37. doi: 10.1152/ajpregu.00282.2003. [DOI] [PubMed] [Google Scholar]
- Hajnal A, Szekely M, Galosi R, Lenard L. Accumbens cholinergic interneurons play a role in the regulation of body weight and metabolism. Physiol Behav. 2000;70:95–103. doi: 10.1016/s0031-9384(00)00236-5. [DOI] [PubMed] [Google Scholar]
- Harris GC, Wimmer M, Aston-Jones G. A role for lateral hypothalamic orexin neurons in reward seeking. Nature. 2005;437:556–559. doi: 10.1038/nature04071. [DOI] [PubMed] [Google Scholar]
- Helm KA, Rada P, Hoebel BG. Cholecystokinin combined with serotonin in the hypothalamus limits accumbens dopamine release while increasing acetylcholine: a possible satiation mechanism. Brain Res. 2003;963:290–297. doi: 10.1016/s0006-8993(02)04051-9. [DOI] [PubMed] [Google Scholar]
- Henningfield JE, Clayton R, Pollin W. Involvement of tobacco in alcoholism and illicit drug use. Br J Addict. 1990;85:279–291. doi: 10.1111/j.1360-0443.1990.tb03084.x. [DOI] [PubMed] [Google Scholar]
- Hernandez L, Hoebel BG. Food reward and cocaine increase extracellular dopamine in the nucleus accumbens as measured by microdialysis. Life Sci. 1988;42:1705–1712. doi: 10.1016/0024-3205(88)90036-7. [DOI] [PubMed] [Google Scholar]
- Heubner H. Endorphins, eating disorders and other addictive behaviors. W. W. Norton; New York: 1993. [Google Scholar]
- Hoebel BG. Brain neurotransmitters in food and drug reward. Am J Clin Nutr. 1985;42:1133–1150. doi: 10.1093/ajcn/42.5.1133. [DOI] [PubMed] [Google Scholar]
- Hoebel BG, Hernandez L, Schwartz DH, Mark GP, Hunter GA. Microdialysis studies of brain norepinephrine, serotonin, and dopamine release during ingestive behavior: theoretical and clinical implications. In: Schneider LH, et al., editors. The Psychobiology of Human Eating Disorders: Preclinical and Clinical Perspectives. Vol. 575. Annals of the New York Academy of Sciences; New York: 1989. pp. 171–193. [DOI] [PubMed] [Google Scholar]
- Hoebel BG, Leibowitz SF, Hernandez L. Neurochemistry of anorexia and bulimia. In: Anderson H, editor. The biology of feast and famine: relevance to eating disorders. Academic Press; New York: 1992. pp. 21–45. [Google Scholar]
- Hoebel BG, Rada P, Mark GP, Pothos E. Neural systems for reinforcement and inhibition of behavior: Relevance to eating, addiction, and depression. In: Kahneman D, et al., editors. Well-being: the Foundations of Hedonic Psychology. Russell Sage Foundation; New York: 1999. pp. 558–572. [Google Scholar]
- Holderness CC, Brooks-Gunn J, Warren MP. Co-morbidity of eating disorders and substance abuse review of the literature. Int J Eat Disord. 1994;16:1–34. doi: 10.1002/1098-108x(199407)16:1<1::aid-eat2260160102>3.0.co;2-t. [DOI] [PubMed] [Google Scholar]
- Howard BV, Wylie-Rosett J. Sugar and cardiovascular disease: A statement for healthcare professionals from the Committee on Nutrition of the Council on Nutrition, Physical Activity, and Metabolism of the American Heart Association. Circulation. 2002;106:523–527. doi: 10.1161/01.cir.0000019552.77778.04. [DOI] [PubMed] [Google Scholar]
- Hubbell CL, Mankes RF, Reid LD. A small dose of morphine leads rats to drink more alcohol and achieve higher blood alcohol concentrations. Alcohol Clin Exp Res. 1993;17:1040–1043. doi: 10.1111/j.1530-0277.1993.tb05661.x. [DOI] [PubMed] [Google Scholar]
- Hurd YL, Kehr J, Ungerstedt U. In vivo microdialysis as a technique to monitor drug transport: correlation of extracellular cocaine levels and dopamine overflow in the rat brain. J Neurochem. 1988;51:1314–1316. doi: 10.1111/j.1471-4159.1988.tb03103.x. [DOI] [PubMed] [Google Scholar]
- Ito R, Dalley JW, Howes SR, Robbins TW, Everitt BJ. Dissociation in conditioned dopamine release in the nucleus accumbens core and shell in response to cocaine cues and during cocaine-seeking behavior in rats. J Neurosci. 2000;20:7489–7495. doi: 10.1523/JNEUROSCI.20-19-07489.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Itzhak Y, Martin JL. Effects of cocaine, nicotine, dizocipline and alcohol on mice locomotor activity: cocaine-alcohol cross-sensitization involves upregulation of striatal dopamine transporter binding sites. Brain Res. 1999;818:204–211. doi: 10.1016/s0006-8993(98)01260-8. [DOI] [PubMed] [Google Scholar]
- Jimerson DC, Lesem MD, Kaye WH, Brewerton TD. Low serotonin and dopamine metabolite concentrations in cerebrospinal fluid from bulimic patients with frequent binge episodes. Arch Gen Psychiatry. 1992;49:132–138. doi: 10.1001/archpsyc.1992.01820020052007. [DOI] [PubMed] [Google Scholar]
- Kalivas PW. Glutamate systems in cocaine addiction. Curr Opin Pharmacol. 2004;4:23–29. doi: 10.1016/j.coph.2003.11.002. [DOI] [PubMed] [Google Scholar]
- Kalivas PW, Striplin CD, Steketee JD, Klitenick MA, Duffy P. Cellular mechanisms of behavioral sensitization to drugs of abuse. Ann N Y Acad Sci. 1992;654:128–135. doi: 10.1111/j.1749-6632.1992.tb25961.x. [DOI] [PubMed] [Google Scholar]
- Kalivas PW, Volkow ND. The neural basis of addiction: a pathology of motivation and choice. Am J Psychiatry. 2005;162:1403–1413. doi: 10.1176/appi.ajp.162.8.1403. [DOI] [PubMed] [Google Scholar]
- Kalivas PW, Weber B. Amphetamine injection into the ventral mesencephalon sensitizes rats to peripheral amphetamine and cocaine. J Pharmacol Exp Ther. 1988;245:1095–1102. [PubMed] [Google Scholar]
- Kantak KM, Miczek KA. Aggression during morphine withdrawal: effects of method of withdrawal, fighting experience, and social role. Psychopharmacology (Berl) 1986;90:451–456. doi: 10.1007/BF00174059. [DOI] [PubMed] [Google Scholar]
- Katherine A. Anatomy of a food addiction: an effective program to overcome compulsive eating. Gurze Books; Carlsbad: 1996. [Google Scholar]
- Katz JL, Valentino RJ. The opiate quasiwithdrawal syndrome in rhesus monkeys: comparison of naloxone-precipitated withdrawal to effects of cholinergic agents. Psychopharmacology (Berl) 1984;84:12–15. doi: 10.1007/BF00432015. [DOI] [PubMed] [Google Scholar]
- Kawasaki T, Kashiwabara A, Sakai T, Igarashi K, Ogata N, Watanabe H, Ichiyanagi K, Yamanouchi T. Long-term sucrose-drinking causes increased body weight and glucose intolerance in normal male rats. Br J Nutr. 2005;93:613–618. doi: 10.1079/bjn20051407. [DOI] [PubMed] [Google Scholar]
- Kelley AE, Bakshi VP, Haber SN, Steininger TL, Will MJ, Zhang M. Opioid modulation of taste hedonics within the ventral striatum. Physiol Behav. 2002;76:365–377. doi: 10.1016/s0031-9384(02)00751-5. [DOI] [PubMed] [Google Scholar]
- Kelley AE, Baldo BA, Pratt WE. A proposed hypothalamic-thalamic-striatal axis for the integration of energy balance, arousal, and food reward. J Comp Neurol. 2005;493:72–85. doi: 10.1002/cne.20769. [DOI] [PubMed] [Google Scholar]
- Kelley AE, Will MJ, Steininger TL, Zhang M, Haber SN. Restricted daily consumption of a highly palatable food (chocolate Ensure(R)) alters striatal enkephalin gene expression. Eur J Neurosci. 2003;18:2592–2598. doi: 10.1046/j.1460-9568.2003.02991.x. [DOI] [PubMed] [Google Scholar]
- Klein DA, Boudreau GS, Devlin MJ, Walsh BT. Artificial sweetener use among individuals with eating disorders. Int J Eat Disord. 2006;39:341–345. doi: 10.1002/eat.20260. [DOI] [PubMed] [Google Scholar]
- Koob GF, Le Moal M. Drug abuse: hedonic homeostatic dysregulation. Science. 1997;278:52–58. doi: 10.1126/science.278.5335.52. [DOI] [PubMed] [Google Scholar]
- Koob GF, Le Moal M. Neurobiology of Addiction. Academic Press; San Diego: 2005. [Google Scholar]
- Koob GF, Maldonado R, Stinus L. Neural substrates of opiate withdrawal. Trends Neurosci. 1992;15:186–191. doi: 10.1016/0166-2236(92)90171-4. [DOI] [PubMed] [Google Scholar]
- Lai S, Lai H, Page JB, McCoy CB. The association between cigarette smoking and drug abuse in the United States. J Addict Dis. 2000;19:11–24. doi: 10.1300/J069v19n04_02. [DOI] [PubMed] [Google Scholar]
- Le KA, Tappy L. Metabolic effects of fructose. Curr Opin Clin Nutr Metab Care. 2006;9:469–475. doi: 10.1097/01.mco.0000232910.61612.4d. [DOI] [PubMed] [Google Scholar]
- Le Magnen J. A role for opiates in food reward and food addiction. In: Capaldi PT, editor. Taste, Experience, and Feeding. American Psychological Association; Washington, D. C.: 1990. pp. 241–252. [Google Scholar]
- Leibowitz SF, Hoebel BG. Behavioral neuroscience and obesity. In: Bray G, et al., editors. The Handbook of Obesity. Marcel Dekker; New York: 2004. pp. 301–371. [Google Scholar]
- Levine AS, Billington CJ. Opioids as agents of reward-related feeding: a consideration of the evidence. Physiol Behav. 2004;82:57–61. doi: 10.1016/j.physbeh.2004.04.032. [DOI] [PubMed] [Google Scholar]
- Levine AS, Kotz CM, Gosnell BA. Sugars: hedonic aspects, neuroregulation, and energy balance. Am J Clin Nutr. 2003;78:834S–842S. doi: 10.1093/ajcn/78.4.834S. [DOI] [PubMed] [Google Scholar]
- Liang NC, Hajnal A, Norgren R. Sham feeding corn oil increases accumbens dopamine in the rat. Am J Physiol Regul Integr Comp Physiol. 2006;291:R1236–R1239. doi: 10.1152/ajpregu.00226.2006. [DOI] [PubMed] [Google Scholar]
- Liguori A, Hughes JR, Goldberg K, Callas P. Subjective effects of oral caffeine in formerly cocaine-dependent humans. Drug Alcohol Depend. 1997;49:17–24. doi: 10.1016/s0376-8716(97)00133-6. [DOI] [PubMed] [Google Scholar]
- Lu L, Grimm JW, Hope BT, Shaham Y. Incubation of cocaine craving after withdrawal: a review of preclinical data. Neuropharmacology. 2004;47(Suppl 1):214–226. doi: 10.1016/j.neuropharm.2004.06.027. [DOI] [PubMed] [Google Scholar]
- Ludwig DS, Peterson KE, Gortmaker SL. Relation between consumption of sugar-sweetened drinks and childhood obesity: a prospective, observational analysis. Lancet. 2001;357:505–508. doi: 10.1016/S0140-6736(00)04041-1. [DOI] [PubMed] [Google Scholar]
- Mark GP, Blander DS, Hoebel BG. A conditioned stimulus decreases extracellular dopamine in the nucleus accumbens after the development of a learned taste aversion. Brain Res. 1991;551:308–310. doi: 10.1016/0006-8993(91)90946-s. [DOI] [PubMed] [Google Scholar]
- Mark GP, Rada P, Pothos E, Hoebel BG. Effects of feeding and drinking on acetylcholine release in the nucleus accumbens, striatum, and hippocampus of freely behaving rats. Journal of Neurochemistry. 1992;58:2269–2274. doi: 10.1111/j.1471-4159.1992.tb10973.x. [DOI] [PubMed] [Google Scholar]
- Mark GP, Weinberg JB, Rada PV, Hoebel BG. Extracellular acetylcholine is increased in the nucleus accumbens following the presentation of an aversively conditioned taste stimulus. Brain Res. 1995;688:184–188. doi: 10.1016/0006-8993(95)00401-b. [DOI] [PubMed] [Google Scholar]
- Markou A, Weiss F, Gold LH, Caine SB, Schulteis G, Koob GF. Animal models of drug craving. Psychopharmacology (Berl) 1993;112:163–182. doi: 10.1007/BF02244907. [DOI] [PubMed] [Google Scholar]
- Marrazzi MA, Luby ED. An auto-addiction opioid model of chronic anorexia nervosa. Int J Eat Disord. 1986;5:191–208. [Google Scholar]
- Marrazzi MA, Luby ED. The neurobiology of anorexia nervosa: an auto-addiction? In: Cohen M, Foa P, editors. The Brain as an Endocrine Organ. Springer-Verlag; New York: 1990. pp. 46–95. [Google Scholar]
- Martin WR. Treatment of heroin dependence with naltrexone. Curr Psychiatr Ther. 1975;15:157–161. [PubMed] [Google Scholar]
- Martin WR, Wikler A, Eades CG, Pescor FT. Tolerance to and Physical Dependence on Morphine in Rats. Psychopharmacologia. 1963;4:247–260. doi: 10.1007/BF00408180. [DOI] [PubMed] [Google Scholar]
- McBride WJ, Murphy JM, Ikemoto S. Localization of brain reinforcement mechanisms: intracranial self-administration and intracranial place-conditioning studies. Behav Brain Res. 1999;101:129–152. doi: 10.1016/s0166-4328(99)00022-4. [DOI] [PubMed] [Google Scholar]
- McSweeney FK, Murphy ES, Kowal BP. Regulation of drug taking by sensitization and habituation. Exp Clin Psychopharmacol. 2005;13:163–184. doi: 10.1037/1064-1297.13.3.163. [DOI] [PubMed] [Google Scholar]
- Mercer ME, Holder MD. Food cravings, endogenous opioid peptides, and food intake: a review. Appetite. 1997;29:325–352. doi: 10.1006/appe.1997.0100. [DOI] [PubMed] [Google Scholar]
- Mifsud JC, Hernandez L, Hoebel BG. Nicotine infused into the nucleus accumbens increases synaptic dopamine as measured by in vivo microdialysis. Brain Res. 1989;478:365–367. doi: 10.1016/0006-8993(89)91518-7. [DOI] [PubMed] [Google Scholar]
- Miller RJ, Pickel VM. Immunohistochemical distribution of enkephalins: interactions with catecholamine-containing systems. Adv Biochem Psychopharmacol. 1980;25:349–359. [PubMed] [Google Scholar]
- Mogenson GJ, Yang CR. The contribution of basal forebrain to limbic-motor integration and the mediation of motivation to action. Adv Exp Med Biol. 1991;295:267–290. doi: 10.1007/978-1-4757-0145-6_14. [DOI] [PubMed] [Google Scholar]
- Mokdad AH, Marks JS, Stroup DF, Gerberding JL. Actual causes of death in the United States, 2000. Jama. 2004;291:1238–1245. doi: 10.1001/jama.291.10.1238. [DOI] [PubMed] [Google Scholar]
- Moore RJ, Vinsant SL, Nadar MA, Poorino LJ, Friedman DP. Effect of cocaine self-administration on dopamine D2 receptors in rhesus monkeys. Synapse. 1998;30:88–96. doi: 10.1002/(SICI)1098-2396(199809)30:1<88::AID-SYN11>3.0.CO;2-L. [DOI] [PubMed] [Google Scholar]
- Mutschler NH, Miczek KA. Withdrawal from a self-administered or non-contingent cocaine binge: differences in ultrasonic distress vocalizations in rats. Psychopharmacology (Berl) 1998;136:402–408. doi: 10.1007/s002130050584. [DOI] [PubMed] [Google Scholar]
- Nelson JE, Pearson HW, Sayers M, Glynn TJ, editors. Guide to Drug Abuse Research Terminology. National Institute on Drug Abuse; Rockville: 1982. [Google Scholar]
- Nichols ML, Hubbell CL, Kalsher MJ, Reid LD. Morphine increases intake of beer among rats. Alcohol. 1991;8:237–240. doi: 10.1016/0741-8329(91)90273-y. [DOI] [PubMed] [Google Scholar]
- Nisell M, Nomikos GG, Svensson TH. Systemic nicotine-induced dopamine release in the rat nucleus accumbens is regulated by nicotinic receptors in the ventral tegmental area. Synapse. 1994;16:36–44. doi: 10.1002/syn.890160105. [DOI] [PubMed] [Google Scholar]
- Nocjar C, Panksepp J. Chronic intermittent amphetamine pretreatment enhances future appetitive behavior for drug- and natural-reward: interaction with environmental variables. Behav Brain Res. 2002;128:189–203. doi: 10.1016/s0166-4328(01)00321-7. [DOI] [PubMed] [Google Scholar]
- O’Brien CP. Anticraving medications for relapse prevention: a possible new class of psychoactive medications. Am J Psychiatry. 2005;162:1423–1431. doi: 10.1176/appi.ajp.162.8.1423. [DOI] [PubMed] [Google Scholar]
- O’Brien CP, Childress AR, Ehrman R, Robbins SJ. Conditioning factors in drug abuse: can they explain compulsion? J Psychopharmacol. 1998;12:15–22. doi: 10.1177/026988119801200103. [DOI] [PubMed] [Google Scholar]
- O’Brien CP, Testa T, O’Brien TJ, Brady JP, Wells B. Conditioned narcotic withdrawal in humans. Science. 1977;195:1000–1002. doi: 10.1126/science.841320. [DOI] [PubMed] [Google Scholar]
- Olds ME. Reinforcing effects of morphine in the nucleus accumbens. Brain Res. 1982;237:429–440. doi: 10.1016/0006-8993(82)90454-1. [DOI] [PubMed] [Google Scholar]
- Pan Y, Berman Y, Haberny S, Meller E, Carr KD. Synthesis, protein levels, activity, and phosphorylation state of tyrosine hydroxylase in mesoaccumbens and nigrostriatal dopamine pathways of chronically food-restricted rats. Brain Res. 2006;1122:135–142. doi: 10.1016/j.brainres.2006.09.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pecina S, Berridge KC. Central enhancement of taste pleasure by intraventricular morphine. Neurobiology (Bp) 1995;3:269–280. [PubMed] [Google Scholar]
- Pelchat ML, Johnson A, Chan R, Valdez J, Ragland JD. Images of desire: food-craving activation during fMRI. Neuroimage. 2004;23:1486–1493. doi: 10.1016/j.neuroimage.2004.08.023. [DOI] [PubMed] [Google Scholar]
- Pellow S, Chopin P, File SE, Briley M. Validation of open:closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. J Neurosci Methods. 1985;14:149–167. doi: 10.1016/0165-0270(85)90031-7. [DOI] [PubMed] [Google Scholar]
- Petry NM. Should the scope of addictive behaviors be broadened to include pathological gambling? Addiction. 2006;101(Suppl 1):152–160. doi: 10.1111/j.1360-0443.2006.01593.x. [DOI] [PubMed] [Google Scholar]
- Piazza PV, Deminiere JM, Le Moal M, Simon H. Factors that predict individual vulnerability to amphetamine self-administration. Science. 1989;245:1511–1513. doi: 10.1126/science.2781295. [DOI] [PubMed] [Google Scholar]
- Picciotto MR, Corrigall WA. Neuronal systems underlying behaviors related to nicotine addiction: neural circuits and molecular genetics. J Neurosci. 2002;22:3338–3341. doi: 10.1523/JNEUROSCI.22-09-03338.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pierce RC, Kalivas PW. Amphetamine produces sensitized increases in locomotion and extracellular dopamine preferentially in the nucleus accumbens shell of rats administered repeated cocaine. J Pharmacol Exp Ther. 1995;275:1019–1029. [PubMed] [Google Scholar]
- Pontieri FE, Monnazzi P, Scontrini A, Buttarelli FR, Patacchioli FR. Behavioral sensitization to heroin by cannabinoid pretreatment in the rat. Eur J Pharmacol. 2001;421:R1–R3. doi: 10.1016/s0014-2999(01)01056-1. [DOI] [PubMed] [Google Scholar]
- Porsolt RD, Anton G, Blavet N, Jalfre M. Behavioural despair in rats: a new model sensitive to antidepressant treatments. Eur J Pharmacol. 1978;47:379–391. doi: 10.1016/0014-2999(78)90118-8. [DOI] [PubMed] [Google Scholar]
- Pothos E, Rada P, Mark GP, Hoebel BG. Dopamine microdialysis in the nucleus accumbens during acute and chronic morphine, naloxone-precipitated withdrawal and clonidine treatment. Brain Res. 1991;566:348–350. doi: 10.1016/0006-8993(91)91724-f. [DOI] [PubMed] [Google Scholar]
- Prasad BM, Ulibarri C, Sorg BA. Stress-induced cross-sensitization to cocaine: effect of adrenalectomy and corticosterone after short- and long-term withdrawal. Psychopharmacology (Berl) 1998;136:24–33. doi: 10.1007/s002130050535. [DOI] [PubMed] [Google Scholar]
- Przewlocka B, Turchan J, Lason W, Przewlocki R. The effect of single and repeated morphine administration on the prodynorphin system activity in the nucleus accumbens and striatum of the rat. Neuroscience. 1996;70:749–754. doi: 10.1016/s0306-4522(96)83012-0. [DOI] [PubMed] [Google Scholar]
- Putnam J, Allhouse JE. Food consumption, prices, and expenditures, 1970-1997. Food and Consumers Economics Division, Economics Research Service, US Department of Agriculture; Washington, D. C.: 1999. [Google Scholar]
- Rada P, Avena NM, Hoebel BG. Adicción al azúcar: ¿Mito ó realidad? Revision. Rev Venez Endocrinol Metab. 2005a;3:2–12. [Google Scholar]
- Rada P, Avena NM, Hoebel BG. Daily bingeing on sugar repeatedly releases dopamine in the accumbens shell. Neuroscience. 2005b;134:737–744. doi: 10.1016/j.neuroscience.2005.04.043. [DOI] [PubMed] [Google Scholar]
- Rada P, Colasante C, Skirzewski M, Hernandez L, Hoebel B. Behavioral depression in the swim test causes a biphasic, long-lasting change in accumbens acetylcholine release, with partial compensation by acetylcholinesterase and muscarinic-1 receptors. Neuroscience. 2006;141:67–76. doi: 10.1016/j.neuroscience.2006.03.043. [DOI] [PubMed] [Google Scholar]
- Rada P, Hoebel BG. Acetylcholine in the accumbens is decreased by diazepam and increased by benzodiazepine withdrawal: a possible mechanism for dependency. Eur J Pharmacol. 2005;508:131–138. doi: 10.1016/j.ejphar.2004.12.016. [DOI] [PubMed] [Google Scholar]
- Rada P, Jensen K, Hoebel BG. Effects of nicotine and mecamylamine-induced withdrawal on extracellular dopamine and acetylcholine in the rat nucleus accumbens. Psychopharmacology (Berl) 2001;157:105–110. doi: 10.1007/s002130100781. [DOI] [PubMed] [Google Scholar]
- Rada P, Johnson DF, Lewis MJ, Hoebel BG. In alcohol-treated rats, naloxone decreases extracellular dopamine and increases acetylcholine in the nucleus accumbens: evidence of opioid withdrawal. Pharmacol Biochem Behav. 2004;79:599–605. doi: 10.1016/j.pbb.2004.09.011. [DOI] [PubMed] [Google Scholar]
- Rada P, Mark GP, Hoebel BG. Galanin in the hypothalamus raises dopamine and lowers acetylcholine release in the nucleus accumbens: a possible mechanism for hypothalamic initiation of feeding behavior. Brain Res. 1998;798:1–6. doi: 10.1016/s0006-8993(98)00315-1. [DOI] [PubMed] [Google Scholar]
- Rada P, Mark GP, Pothos E, Hoebel BG. Systemic morphine simultaneously decreases extracellular acetylcholine and increases dopamine in the nucleus accumbens of freely moving rats. Neuropharmacology. 1991a;30:1133–1136. doi: 10.1016/0028-3908(91)90145-2. [DOI] [PubMed] [Google Scholar]
- Rada P, Paez X, Hernandez L, Avena NM, Hoebel BG. Microdialysis in the study of behavior reinforcement and inhibition. In: Westerink BH, Creamers T, editors. Handbook of Microdialysis: Methods, Application and Perspectives. Academic Press; New York: 2007. pp. 351–375. [Google Scholar]
- Rada P, Pothos E, Mark GP, Hoebel BG. Microdialysis evidence that acetylcholine in the nucleus accumbens is involved in morphine withdrawal and its treatment with clonidine. Brain Res. 1991b;561:354–356. doi: 10.1016/0006-8993(91)91616-9. [DOI] [PubMed] [Google Scholar]
- Rada PV, Hoebel BG. Supraadditive effect of d-fenfluramine plus phentermine on extracellular acetylcholine in the nucleus accumbens: possible mechanism for inhibition of excessive feeding and drug abuse. Pharmacol Biochem Behav. 2000;65:369–373. doi: 10.1016/s0091-3057(99)00219-1. [DOI] [PubMed] [Google Scholar]
- Rada PV, Mark GP, Taylor KM, Hoebel BG. Morphine and naloxone, i.p. or locally, affect extracellular acetylcholine in the accumbens and prefrontal cortex. Pharmacol Biochem Behav. 1996;53:809–816. doi: 10.1016/0091-3057(95)02078-0. [DOI] [PubMed] [Google Scholar]
- Rada PV, Mark GP, Yeomans JJ, Hoebel BG. Acetylcholine release in ventral tegmental area by hypothalamic self-stimulation, eating, and drinking. Pharmacol Biochem Behav. 2000;65:375–379. doi: 10.1016/s0091-3057(99)00218-x. [DOI] [PubMed] [Google Scholar]
- Radhakishun FS, Korf J, Venema K, Westerink BH. The release of endogenous dopamine and its metabolites from rat striatum as detected in push-pull perfusates: effects of systematically administered drugs. Pharm Weekbl Sci. 1983;5:153–158. doi: 10.1007/BF01961473. [DOI] [PubMed] [Google Scholar]
- Ranaldi R, Pocock D, Zereik R, Wise RA. Dopamine fluctuations in the nucleus accumbens during maintenance, extinction, and reinstatement of intravenous D-amphetamine self-administration. J Neurosci. 1999;19:4102–4109. doi: 10.1523/JNEUROSCI.19-10-04102.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riva G, Bacchetta M, Cesa G, Conti S, Castelnuovo G, Mantovani F, Molinari E. Is severe obesity a form of addiction? Rationale, clinical approach, and controlled clinical trial. Cyberpsychol Behav. 2006;9:457–479. doi: 10.1089/cpb.2006.9.457. [DOI] [PubMed] [Google Scholar]
- Robinson TE, Berridge KC. The neural basis of drug craving: an incentive-sensitization theory of addiction. Brain Res Brain Res Rev. 1993;18:247–291. doi: 10.1016/0165-0173(93)90013-p. [DOI] [PubMed] [Google Scholar]
- Rolls ET. Brain mechanisms underlying flavour and appetite. Philos Trans R Soc Lond B Biol Sci. 2006;361:1123–1136. doi: 10.1098/rstb.2006.1852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossetti ZL, Hmaidan Y, Gessa GL. Marked inhibition of mesolimbic dopamine release: a common feature of ethanol, morphine, cocaine and amphetamine abstinence in rats. Eur J Pharmacol. 1992;221:227–234. doi: 10.1016/0014-2999(92)90706-a. [DOI] [PubMed] [Google Scholar]
- Rufus E. Sugar addction: a step-by-step guide to overcoming sugar addiction. Elizabeth Brown Rufus; Bloomington, IN: 2004. [Google Scholar]
- Saad MF, Khan A, Sharma A, Michael R, Riad-Gabriel MG, Boyadjian R, Jinagouda SD, Steil GM, Kamdar V. Physiological insulinemia acutely modulates plasma leptin. Diabetes. 1998;47:544–549. doi: 10.2337/diabetes.47.4.544. [DOI] [PubMed] [Google Scholar]
- Salamone JD. Complex motor and sensorimotor functions of striatal and accumbens dopamine: involvement in instrumental behavior processes. Psychopharmacology (Berl) 1992;107:160–174. doi: 10.1007/BF02245133. [DOI] [PubMed] [Google Scholar]
- Sato Y, Ito T, Udaka N, Kanisawa M, Noguchi Y, Cushman SW, Satoh S. Immunohistochemical localization of facilitated-diffusion glucose transporters in rat pancreatic islets. Tissue Cell. 1996;28:637–643. doi: 10.1016/s0040-8166(96)80067-x. [DOI] [PubMed] [Google Scholar]
- Schenk S, Snow S, Horger BA. Pre-exposure to amphetamine but not nicotine sensitizes rats to the motor activating effect of cocaine. Psychopharmacology (Berl) 1991;103:62–66. doi: 10.1007/BF02244075. [DOI] [PubMed] [Google Scholar]
- Schoffelmeer AN, Wardeh G, Vanderschuren LJ. Morphine acutely and persistently attenuates nonvesicular GABA release in rat nucleus accumbens. Synapse. 2001;42:87–94. doi: 10.1002/syn.1104. [DOI] [PubMed] [Google Scholar]
- Schulteis G, Yackey M, Risbrough V, Koob GF. Anxiogenic-like effects of spontaneous and naloxone-precipitated opiate withdrawal in the elevated plus-maze. Pharmacol Biochem Behav. 1998;60:727–731. doi: 10.1016/s0091-3057(98)00034-3. [DOI] [PubMed] [Google Scholar]
- Schultz W, Dayan P, Montague PR. A neural substrate of prediction and reward. Science. 1997;275:1593–1599. doi: 10.1126/science.275.5306.1593. [DOI] [PubMed] [Google Scholar]
- Schwartz MW, Woods SC, Porte D, Jr, Seeley RJ, Baskin DG. Central nervous system control of food intake. Nature. 2000;404:661–671. doi: 10.1038/35007534. [DOI] [PubMed] [Google Scholar]
- Sclafani A, Nissenbaum JW. Is gastric sham feeding really sham feeding? Am J Physiol. 1985;248:R387–390. doi: 10.1152/ajpregu.1985.248.3.R387. [DOI] [PubMed] [Google Scholar]
- Shalev U, Morales M, Hope B, Yap J, Shaham Y. Time-dependent changes in extinction behavior and stress-induced reinstatement of drug seeking following withdrawal from heroin in rats. Psychopharmacology (Berl) 2001;156:98–107. doi: 10.1007/s002130100748. [DOI] [PubMed] [Google Scholar]
- Sinclair JD, Senter RJ. Development of an alcohol-deprivation effect in rats. Q J Stud Alcohol. 1968;29:863–867. [PubMed] [Google Scholar]
- Smith GP. Sham feeding in rats with chronic, reversible gastric fistulas. In: Crawley JN, et al., editors. Current Protocols in Neruoscience. Vol. 8.6. John Wiley and Sons, Inc.; New York: 1998. pp. D.1–D.6. [DOI] [PubMed] [Google Scholar]
- Smith JE, Co C, Lane JD. Limbic acetylcholine turnover rates correlated with rat morphine-seeking behaviors. Pharmacol Biochem Behav. 1984;20:429–442. doi: 10.1016/0091-3057(84)90282-x. [DOI] [PubMed] [Google Scholar]
- Spanagel R, Herz A, Shippenberg TS. The effects of opioid peptides on dopamine release in the nucleus accumbens: an in vivo microdialysis study. J Neurochem. 1990;55:1734–1740. doi: 10.1111/j.1471-4159.1990.tb04963.x. [DOI] [PubMed] [Google Scholar]
- Spangler R, Goddard NL, Avena NM, Hoebel BG, Leibowitz SF. Elevated D3 dopamine receptor mRNA in dopaminergic and dopaminoceptive regions of the rat brain in response to morphine. Brain Res Mol Brain Res. 2003;111:74–83. doi: 10.1016/s0169-328x(02)00671-x. [DOI] [PubMed] [Google Scholar]
- Spangler R, Wittkowski KM, Goddard NL, Avena NM, Hoebel BG, Leibowitz SF. Opiate-like effects of sugar on gene expression in reward areas of the rat brain. Brain Res Mol Brain Res. 2004;124:134–142. doi: 10.1016/j.molbrainres.2004.02.013. [DOI] [PubMed] [Google Scholar]
- Stein L. Brain endorphins: possible mediators of pleasure and reward. Neurosci Res Program Bull. 1978;16:556–563. [PubMed] [Google Scholar]
- Stein L, Belluzzi JD. Brain endorphins: possible role in reward and memory formation. Fed Proc. 1979;38:2468–2472. [PubMed] [Google Scholar]
- Tanda G, Di Chiara G. A dopamine-mu1 opioid link in the rat ventral tegmentum shared by palatable food (Fonzies) and non-psychostimulant drugs of abuse. Eur J Neurosci. 1998;10:1179–1187. doi: 10.1046/j.1460-9568.1998.00135.x. [DOI] [PubMed] [Google Scholar]
- Teff KL, Elliott SS, Tschop M, Kieffer TJ, Rader D, Heiman M, Townsend RR, Keim NL, D’Alessio D, Havel PJ. Dietary fructose reduces circulating insulin and leptin, attenuates postprandial suppression of ghrelin, and increases triglycerides in women. J Clin Endocrinol Metab. 2004;89:2963–2972. doi: 10.1210/jc.2003-031855. [DOI] [PubMed] [Google Scholar]
- Toida S, Takahashi M, Shimizu H, Sato N, Shimomura Y, Kobayashi I. Effect of high sucrose feeding on fat accumulation in the male Wistar rat. Obes Res. 1996;4:561–568. doi: 10.1002/j.1550-8528.1996.tb00270.x. [DOI] [PubMed] [Google Scholar]
- Turchan J, Lason W, Budziszewska B, Przewlocka B. Effects of single and repeated morphine administration on the prodynorphin, proenkephalin and dopamine D2 receptor gene expression in the mouse brain. Neuropeptides. 1997;31:24–28. doi: 10.1016/s0143-4179(97)90015-9. [DOI] [PubMed] [Google Scholar]
- Turski WA, Czuczwar SJ, Turski L, Sieklucka-Dziuba M, Kleinrok Z. Studies on the mechanism of wet dog shakes produced by carbachol in rats. Pharmacology. 1984;28:112–120. doi: 10.1159/000137951. [DOI] [PubMed] [Google Scholar]
- Uhl GR, Ryan JP, Schwartz JP. Morphine alters preproenkephalin gene expression. Brain Res. 1988;459:391–397. doi: 10.1016/0006-8993(88)90658-0. [DOI] [PubMed] [Google Scholar]
- Unterwald EM. Regulation of opioid receptors by cocaine. Ann N Y Acad Sci. 2001;937:74–92. doi: 10.1111/j.1749-6632.2001.tb03559.x. [DOI] [PubMed] [Google Scholar]
- Unterwald EM, Ho A, Rubenfeld JM, Kreek MJ. Time course of the development of behavioral sensitization and dopamine receptor up-regulation during binge cocaine administration. J Pharmacol Exp Ther. 1994;270:1387–1396. [PubMed] [Google Scholar]
- Unterwald EM, Kreek MJ, Cuntapay M. The frequency of cocaine administration impacts cocaine-induced receptor alterations. Brain Res. 2001;900:103–109. doi: 10.1016/s0006-8993(01)02269-7. [DOI] [PubMed] [Google Scholar]
- Vaccarino FJ, Bloom FE, Koob GF. Blockade of nucleus accumbens opiate receptors attenuates intravenous heroin reward in the rat. Psychopharmacology (Berl) 1985;86:37–42. doi: 10.1007/BF00431681. [DOI] [PubMed] [Google Scholar]
- Vanderschuren LJ, Everitt BJ. Drug seeking becomes compulsive after prolonged cocaine self-administration. Science. 2004;305:1017–1019. doi: 10.1126/science.1098975. [DOI] [PubMed] [Google Scholar]
- Vanderschuren LJ, Everitt BJ. Behavioral and neural mechanisms of compulsive drug seeking. Eur J Pharmacol. 2005;526:77–88. doi: 10.1016/j.ejphar.2005.09.037. [DOI] [PubMed] [Google Scholar]
- Vanderschuren LJ, Kalivas PW. Alterations in dopaminergic and glutamatergic transmission in the induction and expression of behavioral sensitization: a critical review of preclinical studies. Psychopharmacology (Berl) 2000;151:99–120. doi: 10.1007/s002130000493. [DOI] [PubMed] [Google Scholar]
- Vezina P. Sensitization of midbrain dopamine neuron reactivity and the self-administration of psychomotor stimulant drugs. Neurosci Biobehav Rev. 2004;27(8):827–839. doi: 10.1016/j.neubiorev.2003.11.001. [DOI] [PubMed] [Google Scholar]
- Vezina P, Giovino AA, Wise RA, Stewart J. Environment-specific cross-sensitization between the locomotor activating effects of morphine and amphetamine. Pharmacol Biochem Behav. 1989;32:581–584. doi: 10.1016/0091-3057(89)90201-3. [DOI] [PubMed] [Google Scholar]
- Vezina P, Lorrain DS, Arnold GM, Austin JD, Suto N. Sensitization of midbrain dopamine neuron reactivity promotes the pursuit of amphetamine. J Neurosci. 2002;22:4654–4662. doi: 10.1523/JNEUROSCI.22-11-04654.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vigano D, Rubino T, Di Chiara G, Ascari I, Massi P, Parolaro D. Mu opioid receptor signaling in morphine sensitization. Neuroscience. 2003;117:921–929. doi: 10.1016/s0306-4522(02)00825-4. [DOI] [PubMed] [Google Scholar]
- Vilsboll T, Krarup T, Madsbad S, Holst JJ. Both GLP-1 and GIP are insulinotropic at basal and postprandial glucose levels and contribute nearly equally to the incretin effect of a meal in healthy subjects. Regul Pept. 2003;114:115–121. doi: 10.1016/s0167-0115(03)00111-3. [DOI] [PubMed] [Google Scholar]
- Volkow ND, Ding YS, Fowler JS, Wang GJ. Cocaine addiction: hypothesis derived from imaging studies with PET. J Addict Dis. 1996a;15:55–71. doi: 10.1300/J069v15n04_04. [DOI] [PubMed] [Google Scholar]
- Volkow ND, Wang GJ, Fowler JS, Logan J, Hitzemann R, Ding YS, Pappas N, Shea C, Piscani K. Decreases in dopamine receptors but not in dopamine transporters in alcoholics. Alcohol Clin Exp Res. 1996b;20:1594–1598. doi: 10.1111/j.1530-0277.1996.tb05936.x. [DOI] [PubMed] [Google Scholar]
- Volkow ND, Wang GJ, Telang F, Fowler JS, Logan J, Childress AR, Jayne M, Ma Y, Wong C. Cocaine cues and dopamine in dorsal striatum: mechanism of craving in cocaine addiction. J Neurosci. 2006;26:6583–6588. doi: 10.1523/JNEUROSCI.1544-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Volkow ND, Wise RA. How can drug addiction help us understand obesity? Nat Neurosci. 2005;8:555–560. doi: 10.1038/nn1452. [DOI] [PubMed] [Google Scholar]
- Volpicelli JR, Alterman AI, Hayashida M, O’Brien CP. Naltrexone in the treatment of alcohol dependence. Arch Gen Psychiatry. 1992;49:876–880. doi: 10.1001/archpsyc.1992.01820110040006. [DOI] [PubMed] [Google Scholar]
- Volpicelli JR, Ulm RR, Hopson N. Alcohol drinking in rats during and following morphine injections. Alcohol. 1991;8:289–292. doi: 10.1016/0741-8329(91)90401-h. [DOI] [PubMed] [Google Scholar]
- Waller DA, Kiser RS, Hardy BW, Fuchs I, Feigenbaum LP, Uauy R. Eating behavior and plasma beta-endorphin in bulimia. Am J Clin Nutr. 1986;44:20–23. doi: 10.1093/ajcn/44.1.20. [DOI] [PubMed] [Google Scholar]
- Wang GJ, Volkow ND, Logan J, Pappas NR, Wong CT, Zhu W, Netusil N, Fowler JS. Brain dopamine and obesity. Lancet. 2001;357:354–357. doi: 10.1016/s0140-6736(00)03643-6. [DOI] [PubMed] [Google Scholar]
- Wang GJ, Volkow ND, Telang F, Jayne M, Ma J, Rao M, Zhu W, Wong CT, Pappas NR, Geliebter A, Fowler JS. Exposure to appetitive food stimuli markedly activates the human brain. Neuroimage. 2004a;21:1790–1797. doi: 10.1016/j.neuroimage.2003.11.026. [DOI] [PubMed] [Google Scholar]
- Wang GJ, Volkow ND, Thanos PK, Fowler JS. Similarity between obesity and drug addiction as assessed by neurofunctional imaging: a concept review. J Addict Dis. 2004b;23:39–53. doi: 10.1300/J069v23n03_04. [DOI] [PubMed] [Google Scholar]
- Way EL, Loh HH, Shen FH. Simultaneous quantitative assessment of morphine tolerance and physical dependence. J Pharmacol Exp Ther. 1969;167:1–8. [PubMed] [Google Scholar]
- Weiss F. Neurobiology of craving, conditioned reward and relapse. Curr Opin Pharmacol. 2005;5:9–19. doi: 10.1016/j.coph.2004.11.001. [DOI] [PubMed] [Google Scholar]
- Westerink BH, Tuntler J, Damsma G, Rollema H, de Vries JB. The use of tetrodotoxin for the characterization of drug-enhanced dopamine release in conscious rats studied by brain dialysis. Naunyn Schmiedebergs Arch Pharmacol. 1987;336:502–507. doi: 10.1007/BF00169306. [DOI] [PubMed] [Google Scholar]
- Wideman CH, Nadzam GR, Murphy HM. Implications of an animal model of sugar addiction, withdrawal and relapse for human health. Nutr Neurosci. 2005;8:269–276. doi: 10.1080/10284150500485221. [DOI] [PubMed] [Google Scholar]
- Wise RA. The neurobiology of craving: implications for the understanding and treatment of addiction. J Abnorm Psychol. 1988;97:118–132. doi: 10.1037//0021-843x.97.2.118. [DOI] [PubMed] [Google Scholar]
- Wise RA. Opiate reward: sites and substrates. Neurosci Biobehav Rev. 1989;13:129–133. doi: 10.1016/s0149-7634(89)80021-1. [DOI] [PubMed] [Google Scholar]
- Wise RA. Drug self-administration viewed as ingestive behaviour. Appetite. 1997;28:1–5. doi: 10.1006/appe.1996.0059. [DOI] [PubMed] [Google Scholar]
- Wise RA, Bozarth MA. Brain reward circuitry: four circuit elements “wired” in apparent series. Brain Res Bull. 1984;12:203–208. doi: 10.1016/0361-9230(84)90190-4. [DOI] [PubMed] [Google Scholar]
- Wise RA, Newton P, Leeb K, Burnette B, Pocock D, Justice JB., Jr Fluctuations in nucleus accumbens dopamine concentration during intravenous cocaine self-administration in rats. Psychopharmacology (Berl) 1995;120:10–20. doi: 10.1007/BF02246140. [DOI] [PubMed] [Google Scholar]
- Yeomans JS. Role of tegmental cholinergic neurons in dopaminergic activation, antimuscarinic psychosis and schizophrenia. Neuropsychopharmacology. 1995;12:3–16. doi: 10.1038/sj.npp.1380235. [DOI] [PubMed] [Google Scholar]
- Yoshimoto K, McBride WJ, Lumeng L, Li TK. Alcohol stimulates the release of dopamine and serotonin in the nucleus accumbens. Alcohol. 1992;9:17–22. doi: 10.1016/0741-8329(92)90004-t. [DOI] [PubMed] [Google Scholar]
- Zangen A, Nakash R, Overstreet DH, Yadid G. Association between depressive behavior and absence of serotonin-dopamine interaction in the nucleus accumbens. Psychopharmacology (Berl) 2001;155:434–439. doi: 10.1007/s002130100746. [DOI] [PubMed] [Google Scholar]
- Zhang M, Gosnell BA, Kelley AE. Intake of high-fat food is selectively enhanced by mu opioid receptor stimulation within the nucleus accumbens. J Pharmacol Exp Ther. 1998;285:908–914. [PubMed] [Google Scholar]
- Zhang M, Kelley AE. Intake of saccharin, salt, and ethanol solutions is increased by infusion of a mu opioid agonist into the nucleus accumbens. Psychopharmacology (Berl) 2002;159:415–423. doi: 10.1007/s00213-001-0932-y. [DOI] [PubMed] [Google Scholar]
- Zubieta JK, Gorelick DA, Stauffer R, Ravert HT, Dannals RF, Frost JJ. Increased mu opioid receptor binding detected by PET in cocaine-dependent men is associated with cocaine craving. Nat Med. 1996;2:1225–1229. doi: 10.1038/nm1196-1225. [DOI] [PubMed] [Google Scholar]