

Abstract
Higher plant reproduction is unique because two cells are fertilized in the haploid female gametophyte. Egg and sperm nuclei fuse to form the embryo. A second sperm nucleus fuses with the central cell nucleus that replicates to generate the endosperm, a tissue that supports embryo development. To understand mechanisms that initiate reproduction, we isolated a mutation in Arabidopsis, f644, that allows for replication of the central cell and subsequent endosperm development without fertilization. When mutant f644 egg and central cells are fertilized by wild-type sperm, embryo development is inhibited, and endosperm is overproduced. By using a map-based strategy, we cloned and sequenced the F644 gene and showed that it encodes a SET-domain polycomb protein. Subsequently, we found that F644 is identical to MEDEA (MEA), a gene whose maternal-derived allele is required for embryogenesis [Grossniklaus, U., Vielle-Calzada, J.-P., Hoeppner, M. A. & Gagliano, W. B. (1998) Science 280, 446–450]. Together, these results reveal functions for plant polycomb proteins in the suppression of central cell proliferation and endosperm development. We discuss models to explain how polycomb proteins function to suppress endosperm and promote embryo development.
Keywords: reproduction, embryo, seed, apomixis
How fertilization activates reproductive development is a fundamental problem in biology. In higher plants, the ovule produces a female gametophyte that is composed of egg, central, synergid, and antipodal cells (Fig. 1 A and C). All are haploid, except the central cell, which contains two daughter nuclei that fuse before fertilization to form a diploid central cell nucleus. Surrounding and protecting the female gametophyte are maternal cell layers, integuments (Fig. 1C). Egg and sperm fuse to generate the embryo (Fig. 1I). Fusion of the adjacent central cell with a second sperm cell generates the primary triploid endosperm nucleus that replicates to form the endosperm (Fig. 1I), a tissue that supports the development of the embryo (1). Fertilization also activates maternal tissue differentiation. The seed coat (Fig. 1I) is generated from the ovule integuments. To support the development of the rapidly growing seeds, the ovary differentiates and elongates to form the fruit, or silique, in Arabidopsis (Fig. 2A).
Figure 1.

Endosperm and seed coat development in mutant Arabidopsis plants when fertilization is prevented. (A) Wild-type silique. (B) f644 homozygous silique. (C) Wild-type ovule. (D–H) f644 homozygous seed-like structures. (I) Wild-type seed with globular embryo 2 days after self-pollination. Siliques, ovules and seed-like structures in A–H were harvested 6 days after removal of anthers. CCN, central cell nucleus; ECN, egg cell nucleus; EM, embryo, EN, endosperm; ENN, one of the endosperm nuclei in the endosperm; FG, female gametophyte; I, integument; OV, ovule; SC, seed coat; SLS, seed-like structure. Unlabeled arrows indicate nuclei derived from the unfertilized central cell. [Bar = 0.5 mm (A–B) and 0.1 mm (C–I).]
Figure 2.

Seed development in wild-type and mutant Arabidopsis plants. Siliques and seeds shown in panels A–G were harvested 9 days after self-pollination. Those in panels H–I were harvested 5 days after self-pollination. Seeds in F–I were cleared and photographed by using Nomarski optics as described in Methods. (A) Portion of a wild-type silique. (B) Portion of an f644 heterozygous silique. (C) Portion of an f644 homozygous silique. (D and F) Seeds with wild-type phenotype from f644 heterozygous silique in B. (E and G) Seeds with mutant phenotype from f644 heterozygous silique in B. Arrows in Inset point to nuclei. (H) Wild-type seed. (I) Homozygous f644 seed. AE, abnormal embryo; EM, embryo; EN, endosperm; m, seed with a mutant phenotype; wt, seed with a wild-type phenotype; SC, seed coat. [Bar = 0.5 mm (A–C), 0.1 mm (D–I), and 0.04 mm (Inset in G).]
Embryo and endosperm follow distinct patterns of development. In Arabidopsis, the embryo passes through a series of stages that have been defined morphologically as preglobular (Fig. 1I), globular, heart, cotyledon (Fig. 2H), and maturation (Fig. 2F). During these stages, an axis of polarity is fixed, shoot and root meristems are formed, and storage organs are generated. In contrast, the primary endosperm nucleus mitotically divides and produces a syncytium of nuclei (Fig. 1I). These nuclei migrate within an expanding central cell (2), after which cytokinesis occurs, and endosperm nuclei and cytoplasm are partitioned into cells. The endosperm (Fig. 2H) produces storage proteins, starch, and lipids, all of which are taken up by the developing embryo (1).
We and others isolated Arabidopsis mutants, fertilization-independent endosperm (fie) (3) and fertilization-independent seed (fis) (4), that act in the female gametophyte and allow for aspects of reproductive development to occur without fertilization. When fertilization is prevented by the removal of pollen-producing anthers, the mutant central cell replicates to form a diploid endosperm, and the maternal ovary and ovule integuments generate the fruit and seed coat, respectively (3, 4). Recently, the FIE gene has been shown to encode a polycomb protein with WD repeats (5). Therefore, this class of polycomb protein functions to suppress central-cell nuclear replication and endosperm development in the female gametophyte until fertilization occurs.
Inheritance of the mutant fie or fis allele by a female gametophyte is deleterious for embryo development even when the pollen bears the wild-type allele (3, 4). In Arabidopsis, two other mutations have been identified based on a similar maternal “parent-of-origin” effect on embryogenesis, embryo-defective173 (emb173) (6) and medea (mea) (7).
To understand how fertilization initiates reproductive development, we have isolated the mutation f644. Central-cell nuclei with the f644 mutant allele replicate and initiate endosperm development without fertilization. When the f644 mutant egg and central cells are fertilized with mutant or wild-type male gametes, the embryo aborts, and endosperm accumulates to a higher than normal level. By using a map-based strategy, we cloned the F644 gene and showed that it is related to the family of SET-domain polycomb proteins (8). Subsequently, we became aware that the F644 gene was identical to the MEA gene that had been reported to control embryonic cell proliferation during the middle and late stages of embryogenesis (7). The phenotypes associated with the f644 mutation revealed functions for the MEA/F644 gene in the suppression of central cell proliferation and endosperm development.
METHODS
Plant Material.
Siliques, seed-like structures, and seeds were prepared and photographed as described (3). Approximate seed volume was calculated by using the equation 4/3πa2b, where a is the minor axis and b is the major axis of the ellipsoid-shaped seed.
Mapping the f644 Mutation.
The F644 mutation was mapped to position 3.0 on chromosome 1 between restriction fragment length polymorphism (RFLP) markers nga59 and 0846A by using procedures described previously (3). For fine structure mapping, we selected plants bearing recombinant chromosomes with breakpoints between f644 and the downstream morphological marker, axr3 (9). Eighty-one F2 recombinant plants were identified and used to map the f644 mutation relative to closely linked downstream RFLP markers. We also selected plants bearing recombinant chromosomes with breakpoints between f644 and the upstream morphological marker, emb60 (10). Twenty-six recombinant F2 plants were identified and used to map the f644 mutation relative to closely linked upstream RFLP markers.
Complementation.
Agrobacterium-mediated transformation of f644 heterozygous plants was performed as described (11). For cosmid 6–22, 10 transgenic T1 lines were obtained that displayed 25% seed abortion (e.g., 81 normal:22 aborted, 3:1, χ2 = 0.02, P > 0.9) in self-pollinated siliques, suggesting that an unlinked wild-type F644 allele on cosmid 6-22 complemented the mutant f644 allele. Lines homozygous for both the mutant f644 allele and the transgenic wild-type F644 allele displayed no seed abortion in self-pollinated siliques. After fertilization was prevented by the removal of anthers, all ovules (n = 177) displayed a central cell with a single nucleus.
DNA Sequencing.
Cosmid 6-22 DNA sequence was determined by LARK Sequencing Technologies (Houston). To identify mutant alleles, homozygous mutant genomic regions were amplified by using PCR, subcloned, and sequenced. To eliminate PCR-induced sequence errors, a minimum of three independent PCR products were sequenced. The f644 mutant sequence was also confirmed by converting it into a codominant cleaved amplified polymorphic sequence (12). cDNA clones were obtained by using reverse transcription–PCR.
RNA Methods.
Flower bud tissue from stage 0 to 12 (13), open flowers (stage 13), young rosette leaves, stems, and roots were harvested, and total RNA was isolated by using Trizol reagent (Life Technologies, Grand Island, NY) and purified by using the RNeasy Mini Kit (Qiagen, Chatsworth, CA). All reverse transcription–PCR reactions were performed with 2 μg of RNA as starting material by using a RETROscript kit (Ambion, Austin, TX), blotted, and hybridized to a MEA cDNA probe using standard procedures. Primers used to amplify MEA RNA were CGAAGTGGATGTTTCGGAC and CGCAATATTTCTCGCAGCAAT. Primers used to amplify control GAPC (glyceraldehyde-3-phosphate dehydrogenase C) RNA (14) were CACTTGAAGGGTGGTGCCAAG and CCTGTTGTCGCCAACGAAGTC.
RESULTS
f644 Mutation Allows Endosperm Development Without Fertilization.
We previously described our procedure, based on silique elongation, for isolating mutant lines that display fertilization-independent development and our analysis of mutations in one gene, FIE, that mapped to chromosome 3 (3). Another mutation, f644, mapped to chromosome 1 and therefore represented a lesion in a distinct gene. Mature mutant f644 ovules (Fig. 1D) resembled wild-type ovules (Fig. 1C). The f644 female gametophyte contained the proper number of nuclei (data not shown), including a haploid egg and a diploid central cell nucleus that resulted from the fusion of two polar nuclei (Fig. 1C). When fertilization was prevented in heterozygous or homozygous f644 plants, siliques partially elongated and contained seed-like structures (Fig. 1B) that were not observed in wild-type controls (Fig. 1A). Within the seed-like structures, the large central cell replicated (Fig. 1 E and F) and multiple central cell nuclei were observed (Fig. 1G) within an expanded central cell surrounded by a seed coat (Fig. 1H). Taken together, these results show that the f644 mutation activated central-cell nuclear replication, as well as maternal sporophyte seed coat and silique elongation, without fertilization.
Fertilization-independent endosperm development in f644 mutant female gametophytes is a partially penetrant phenotype that is affected by genetic background. Approximately 18% of mutant female gametophytes displayed multinucleate central cells without fertilization in the Landsberg genetic background (Table 1). In control wild-type female gametophytes, regardless of the genetic background, no multinucleate central cells were observed (Fig. 1C and Table 1). The degree of penetrance was similar in heterozygous and homozygous f644 mutant plants (Table 1), suggesting that the genotype of the surrounding sporophyte (e.g., ovule integuments) had little influence on nuclear replication of the central cell in mutant gametophytes. These results suggest that in wild-type plants, the F644 gene acts, along with other factors, in the female gametophyte to suppress central-cell nuclear replication and endosperm formation before fertilization.
Table 1.
Penetrance of the fertilization-independent endosperm phenotype in the female gametophyte
| Ovule genotype (ecotype) | Multinucleate central cells, % | Gametophyte penetrance |
|---|---|---|
| Wild type | 0 | — |
| f644/+ (Ler) | 7.5 | 15.0 |
| f644/f644 (Ler) | 18.6 | 18.6 |
| f644/+ (Col) | 10.3 | 20.6 |
| f644/+ (WS) | 1.5 | 3.0 |
| emb173/+ (WS) | 1.8 | 3.6 |
Multinucleate central cells had 3 or more nuclei. Nuclei were counted 6 days after emasculation. Number of ovules checked were: Landsberg erecta (Ler), 515; Columbia glabrous1 (Col), 283; Wassilewskija (WS), 1739; f644/+ (Ler), 273; f644/f644 (Ler), 613; f644/+ (Col), 145; f644/+ (WS), 861; emb173/+ (WS), 954.
f644 Mutation Is Rarely Transmitted by the Female Gametophyte.
When heterozygous f644 females were crossed to wild-type males, siliques (Fig. 2B) were produced with equal numbers (81:87, 1:1, χ2 = 0.22, P > 0.95) of viable seed with normal green embryos (Fig. 2 D and F) and white or brown seeds with abnormal embryos (Fig. 2 E and G). Seeds from this cross were germinated, and of 81 progeny, only 3 were heterozygous for the f644 mutation. Thus, the f644 mutant allele was infrequently transmitted by the female gametophyte to the next generation. In contrast, when wild-type females were crossed to heterozygous f644 males, siliques had no aborted seed (data not shown), and a 1:1 segregation of wild-type and heterozygous progeny was observed (51:72, χ2 = 3.58, P > 0.06), indicating that the f644 mutation did not affect transmission by the male gametophyte. Rare transmission of the f644 mutant allele by the female gametophyte made it possible to obtain homozygous f644 plants that generated normal rosette leaves, an inflorescence, and produced siliques containing 100% white or brown defective seeds (Fig. 2C). Thus, the f644 mutation specifically affects maternal reproductive processes and is not an absolute requirement for vegetative growth and development.
f644 Mutation Suppresses Embryo Development.
Wild-type and mutant embryos developed in a similar fashion during the first 4 days after pollination (DAP). At 4 DAP, both wild-type and mutant embryos are heart-shaped and contain ≈500 cells (Fig. 3A). At 5 DAP, differences between wild-type and mutant embryo development could be discerned. For example, because of rapid cell proliferation in the cotyledons of wild-type embryos at 5 DAP (Fig. 2H), the embryonic cell number increased to ≈1,000 (Fig. 3A). In contrast, mutant embryos remained heart-shaped (Fig. 2I) with only ≈600 cells (Fig. 3A). At 9 DAP, the wild-type mature embryo (Fig. 2 D and F) filled the seed and contained >15,000 cells (Fig. 3A; ref. 15), whereas the mutant embryos were still heart-shaped (Fig. 2G) and comprised only ≈630 cells (Fig. 3A). Similar results were observed with homozygous mutant embryos and with heterozygous embryos that inherited a maternal-derived f644 mutant allele (data not shown). These results indicate that the maternal-derived f644 mutant allele, directly or indirectly, inhibits embryo cell proliferation and morphogenesis.
Figure 3.

Embryo and endosperm proliferation in wild-type and mutant Arabidopsis plants. Seeds were cleared and visualized with Nomarski optics, and the number of embryo cells (A) and endosperm nuclei (B) were counted. The average number of cells or nuclei (±SD, n = 10) is shown for each time point, except the number of cells in only a single wild-type embryo at 9 DAP was determined. Wild-type embryos were at the globular stage at 2 DAP, early heart stage at 4 DAP, early torpedo stage at 5 DAP, and mature stage at 9 DAP. ○, wild-type; ■, f644 mutant; ⋄, 7523 mutant. 7523 is a recessive zygotic embryo defective mutation in the Wassilewskja genetic background that arrests embryo development at the early heart stage.
One explanation for the defect in f644 seed development is that a mutant female gametophyte does not support double fertilization. To test this possibility, we pollinated mutant f644 pistils with pollen from plants homozygous for a seed-specific reporter gene, β-conglycinin promoter fused to β-glucuronidase-coding sequences (3). β-glucuronidase activity was detected in both the aborted embryo and endosperm (data not shown). These results indicate that both the mutant egg and central cell were fertilized.
f644 Mutation Promotes Endosperm Development.
Wild-type and mutant endosperm developed in a similar fashion during the first 4 DAP. At 4 DAP, wild-type and mutant seeds contained a syncytial endosperm composed of ≈350 nuclei (Fig. 3B). However, at 5 DAP, differences between wild-type and mutant endosperm development became apparent. In wild-type endosperm, replication ceased at 5 DAP (Fig. 3B) and cellularization began. Soon thereafter, the wild-type embryo absorbed the endosperm, and the number of nuclei decreased (data not shown). In contrast, in mutant endosperm, the period of nuclear replication was extended, resulting in endosperm with nearly 150% the number of nuclei (Fig. 3B). At 9 DAP, the volume of mutant seeds (Fig. 2E, 6.2 ± 1.3 mm3, n = 13), was almost 2-fold greater than a wild-type control mature seed (Fig. 2D, 3.4 ± 0.9 mm3, n = 13). Similar results were observed with homozygous mutant endosperm and with heterozygous endosperm that inherited a maternal-derived f644 mutant allele (data not shown). Ultimately, f644 mutant seed accumulated red and brown pigments, desiccated, and decreased in size (Fig. 2 B and C), similar to seeds with aborted embryos because of zygotic mutations (16).
One possibility to explain these results was that the increase in endosperm nuclei was due to the failure of the aborted embryo to absorb endosperm. However, the increase in the number of endosperm nuclei occurred between 4 and 5 days after pollination (Fig. 3B), before significant endosperm absorption takes place. Moreover, endosperm overproduction was not observed in seeds with aborted embryos caused by zygotic mutations (Fig. 3B; R.L.F., unpublished results). Taken together, these results suggest that mutations in the maternal-derived f644 allele have opposite effects on embryo and endosperm development, suppressing the former and promoting the latter.
emb173 Mutant Phenotype Is Similar to f644.
We examined the phenotype of another available female gametophyte mutation, emb173 (6). It had been shown that inheritance of the maternal-derived emb173 mutant allele resulted in the production of unusually large seeds with embryos arrested at the heart stage (6). We found that mutant emb173 seeds, like f644 mutant seeds, accumulated an unusually high level of endosperm (data not shown). Moreover, when fertilization was prevented in emb173 heterozygous or homozygous flowers, seed-like structures comprised of an endosperm surrounded by a seed coat were formed (data not shown). However, the frequency of emb173 seed-like structure formation was approximately one-fifth of that observed for f644 (Table 1). The reduced penetrance appears to be due to differences in the genetic backgrounds of f644 (Landsberg ecotype) and emb173 (Wassilewskija ecotype). When we introduced the f644 mutation into the Wassilewskija genetic background, there was a concomitant drop in penetrance to one-fifth the level observed in the Landsberg ecotype (Table 1). The decrease in penetrance appears to be specific to Wassilewskija, and introducing the f644 mutation into the Columbia genetic background did not decrease the penetrance (Table 1). Taken together, these results indicate that the f644 and emb173 mutations, both located at approximately position 3.0 on chromosome 1 (Methods and ref. 6), generated very similar phenotypes; promoting endosperm, and suppressing embryo, development.
Cloning the F644 Gene.
As shown in Fig. 4A, we used a map-based strategy to localize the F644 gene to a pair of overlapping bacterial artificial chromosome clones. Genetic complementation experiments indicated that the entire F644 gene resided on cosmid 6-22 (Fig. 4A). Because f644 is a gametophytic mutation, it was not possible to determine whether the mutant allele was dominant or recessive by using classical genetic procedures. However, these complementation experiments indicate that the mutant f644 allele is recessive to the wild-type F644 allele, suggesting that F644 suppresses endosperm development before fertilization.
Figure 4.
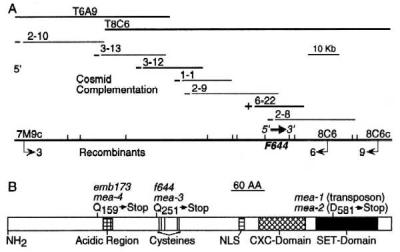
Genetic mapping and positional cloning and position of mutant mea alleles. (A) Position of the F644 gene relative to overlapping BAC clones. →, number of upstream recombinant chromosomes between indicated marker and f644; ←, number of downstream recombinant chromosomes between indicated marker and f644. −, no complementation; +, complementation. Tick marks represent BamHI restriction sites. Position and orientation of the F644 gene relative to cosmid clones is shown. (B) Position of mutant alleles relative to domains in the MEA protein. Position of the mea-1 and mea-2 mutations is from sequence data in Grossniklaus et al. (7). Position of conserved amino acids, acidic region, cysteines, CXC domain, SET domain, and the nuclear localization signal (NLS) are shown (21). NH2, amino end of protein.
The DNA sequence of the region spanned by the complementing cosmid 6-22 was determined, and computer analysis revealed an ORF. The DNA sequence of the ends of flanking noncomplementing cosmids 2-9 and 2-8 was determined and shown to terminate within the ORF. A cDNA clone corresponding to the ORF was isolated from floral bud RNA, sequenced, and shown to encode a 689aa polypeptide related to the family of SET-domain polycomb proteins (8). Comparison of the wild-type and f644 mutant sequence revealed a single base pair difference that generated a premature translation termination codon within the ORF, verifying that F644 is a polycomb gene (Fig. 4B). Moreover, comparison of the wild-type and emb173 mutant sequence revealed another single base pair difference that generated a premature translation termination codon (Fig. 4B). Thus, f644 and emb173 have similar phenotypes and are two new mutant alleles of the same polycomb gene. It is likely that the f644 and emb173 mutations represent loss-of-function alleles, consistent with their being recessive to wild-type, and further supporting the idea that the wild-type F644 allele prevents the central cell from initiating endosperm development. After completing the cloning and sequencing experiments, we became aware that F644 was identical to MEA, a gene reported to control embryonic cell proliferation during middle and late embryogenesis (7).
We compared the level of F644/MEA RNA accumulation in immature floral buds that contain developing female gametophytes, open flowers that are being fertilized, postfertilization young siliques that contain seeds with preglobular to heart stage embryos, and leaf, stem, and root from mature plants. RNA gel-blot experiments detected a low-abundance RNA of the expected size, 2.3 kb (data not shown). As shown in Fig. 5, a more sensitive procedure, amplifying F644/MEA RNA by reverse transcription–PCR followed by gel-blot hybridization to a labeled probe, showed that F644/MEA RNA is present in immature floral buds, open flowers, young siliques, stems, and roots. F644/MEA RNA also was detected in seedlings (data not shown). These results indicate that the F644/MEA gene is expressed in both reproductive and vegetative phases of the plant life cycle.
Figure 5.

Analysis of F644/MEA RNA accumulation. L, leaf; S, stem; R, root; IF, immature floral buds; OF, open flowers; YS, young siliques. As a control, glyceraldehyde-3-phosphate dehydrogenase C (GAPC) RNA was amplified.
DISCUSSION
We discovered a mutation, f644, that has multiple phenotypes: (i) fertilization-independent endosperm development and (ii) embryo abortion and endosperm overproduction after fertilization. We cloned the F644 gene and found that it encoded a SET-domain polycomb protein (8). Subsequently, we became aware that F644 was identical to the MEA polycomb protein (7). The identity between F644 and MEA was not predicted because, as discussed below, the f644 phenotype was distinct from mea. However, these results revealed functions for the MEA gene in the control of central cell proliferation and endosperm development. We now refer to f644 and emb173 as two mutant mea alleles, mea-3 and mea-4.
MEA Suppresses Replication of the Central Cell Nucleus in the Female Gametophyte.
The higher plant life cycle alternates between multicellular gametophyte and sporophyte generations. Morphologically, the gametophyte and sporophyte generations are quite distinct. This suggests the existence of regulatory proteins that function to ensure that the appropriate gametophyte-specific and sporophyte-specific developmental pathways are followed in each generation. Here we show that the mea-3 and mea-4 mutations allow for fertilization-independent replication of the central cell nucleus, the first step in endosperm development (Fig. 1). Thus, the MEA gene plays an important role in preventing endosperm formation, a sporophytic pattern of development, from occurring in the female gametophyte before fertilization occurs.
The MEA polypeptide is related to the family of SET-domain polycomb proteins (7). In mammals, insects, and fungi, these diverse polypeptides have been shown to participate in the formation of complexes at specific sites within the genome (17). By influencing the degree of localized chromatin condensation, polycomb proteins regulate gene transcription and cell proliferation (17). We have shown that endosperm development occurs in the female gametophyte as a result of loss-of-function mutations in the MEA gene. Hence, we propose that the MEA protein suppresses transcription of genes needed for replication of the central cell nucleus. Thus, loss-of-function mutations in the MEA gene may prevent, at least in part, the establishment or maintenance of silencing complexes in the female gametophyte, resulting in central-cell nuclear replication and the initiation of endosperm development without fertilization.
In support of the idea that complexes of polycomb proteins suppress endosperm development in the female gametophyte, it has recently been shown that the SET-domain polycomb-group proteins in insects and mammals physically interact with a different class of polycomb group protein that contains WD40 repeats (18–20). We have shown that mutations in the FIE gene allow for fertilization-independent endosperm development (3), and that the FIE gene encodes a WD40-motif polycomb protein (5). Thus, it is possible that the molecular partnership of SET-domain (i.e., MEA) and WD40-domain (i.e., FIE) polycomb proteins may also be conserved in plants, where these proteins function to suppress precocious endosperm development in the female gametophyte.
Genetic penetrance of the mea fertilization-independent endosperm phenotype is partial (Table 1). This suggests that other related SET-domain polycomb group proteins may have the capacity to substitute for the MEA protein, at least in part, in the female gametophyte. A search of the databases identified proteins, for example CURLY LEAF (21), with related SET- and CXC-domains (Fig. 4B), essential for polycomb protein activity. Thus, it is possible that the low penetrance is due to genetic redundancy.
MEA Suppresses Late Replication of Endosperm Nuclei.
We have found that mutant endosperm nuclear proliferation is prolonged in seeds that inherited a maternal-derived mea-3 (Fig. 3B) or mea-4 allele. Thus, we conclude that the function of wild-type MEA is to suppress late endosperm nuclear proliferation. It is not known whether MEA acts by the same mechanism to suppress nuclear replication of the central cell before fertilization and replication of endosperm nuclei after fertilization.
In contrast to what we observed in mea-3 and mea-4 seeds, endosperm nuclear divisions were reported to take place more slowly in mea-1 and mea-2 endosperm, with fewer nuclei generated (7). The mea-1 (i.e., transposon insertion) and mea-2 (i.e., premature translation termination codon) mutations are in the exon encoding the carboxyl-terminal SET domain (Fig. 4B). Thus, mea-1 and mea-2 mutant polypeptides, although probably lacking a functional SET domain, would be predicted to contain important upstream domains: conserved acidic and cysteine residues, a nuclear localization signal, and the CXC domain. In contrast, the mea-3 and mea-4 mutations generate translation stop codons in the amino-terminal portion of the MEA polypeptide (Fig. 4B). Thus, mea-3 and mea-4 mutant polypeptides would be predicted to lack most, or all, of the protein domains required for polycomb activity, respectively. Because mutant alleles were analyzed in the same genetic background (i.e., Landsberg for mea-1, mea-2, and mea-3), it is possible that the distinct endosperm phenotypes reflect differences in the mutant alleles.
MEA Is Required for Proper Embryonic Cell Proliferation.
In seeds that inherited a maternal-derived mea-1 or mea-2 allele, it was reported that the mutant embryo showed increased cell proliferation, and the authors concluded that the wild-type function of MEA is to restrict cell proliferation during embryogenesis (7). In contrast, when wild-type and mutant seeds of equal age were compared (Fig. 3B), we found that cell proliferation during the later stages of embryogenesis was severely reduced in seeds that inherited a maternal-derived mea-3 or mea-4 allele. Thus, the mea-3 and mea-4 mutant embryo phenotype, and the nature of the mea-3 and mea-4 lesions (Fig. 4B), indicates that wild-type MEA is required, either directly or indirectly, to attain the normal level of cell proliferation during embryogenesis.
Why Does the Mutant Maternal-Derived Allele Cause Embryo Abortion?
One explanation is that a single paternal-derived wild-type allele, in combination with two mutant maternal-derived alleles, is not sufficient for proper endosperm development, which in turn is required for embryo development. However, this model is not likely to be correct because increasing the number of paternal-derived wild-type alleles, either by the use of tetraploid lines (7) or transgenic lines (data not shown), did not rescue the aborted embryo. Alternatively, essential MEA gene expression for embryo development may occur before fertilization in the female gametophyte. Although this model cannot be ruled out, it is important to note that the MEA gene is expressed after fertilization in siliques and in vegetative tissues (Fig. 5). A third possibility is that the paternal-derived allele does not produce active MEA protein, perhaps because of transcriptional silencing (22).
The postfertilization phenotype of seeds that inherit a maternal-derived mutant allele, endosperm overproduction, and embryo abortion, supports the “silencing” model for parent-of-origin effects on seed development as described by Haig and Westoby (23). This theory accounts for similarities in genetic imprinting observed in mammals and flowering plants by proposing that the maternal and paternal genomes have conflicting interests in relation to the transfer of nutrients from the mother to the embryo via specialized acquisitive tissues (i.e., endosperm in plants). According to this theory, a gene that tends to suppress endosperm development, and therefore restricts nutrient flow to the embryo, would be preferentially expressed by the maternal allele. In contrast, a gene that tends to promote endosperm development, and therefore increases nutrient flow to the embryo, would be preferentially expressed by the paternal allele. In this way, the female genome saves resources for reproduction in the future, whereas the paternal genome promotes transmission of its genome. Analysis of the products of interploidy crosses in many plant species (24), including Arabidopsis (25), reveals that paternal genomic excess is associated with overproduction of endosperm, whereas maternal genomic excess is associated with the opposite. Ultimately, successful reproduction in many plant species (24), including Arabidopsis (25), requires the proper balance of maternal and paternal genomes within the endosperm, and sufficient maternal or paternal excess results in embryo abortion. Thus, it is possible that the wild-type maternal MEA allele supports the “interests” of the maternal genome by establishing complexes of polycomb proteins that tend to suppress endosperm development, a process needed for proper embryo development.
Apomixis and Polycomb Proteins.
Apomictic plant species asexually generate embryo, endosperm, maternal seed coat, and fruit (26). The fertilization-independent phenotype associated with mutations in genes such as MEA, FIE, and FIS reveals that Arabidopsis, a sexually reproducing plant, has the genetic potential for aspects of fertilization-independent reproductive development (3, 4). The fact that both FIE and MEA encode polycomb-group proteins suggests that alterations in the activity of polycomb proteins may be responsible, at least in part, for fertilization-independent reproductive development observed in naturally occurring apomictic plants.
Acknowledgments
We thank D. Meinke for emb60 and emb173 seed, E. Grill for an Arabidopsis cosmid library, and C. Shieh for help in gene cloning experiments. We gratefully acknowledge support from Ceres, Inc. (Malibu, CA) for DNA sequencing. This work was funded by a grant to R.L.F. and N.O. from the Binational Agricultural Research and Development Fund (97-34339-4954) and the U.S.-Israel Binational Science Fund (96-00242/1). T.K. was the recipient of a Japan Society for the Promotion of Science Postdoctoral Fellowship. R.Y. is a Department of Energy Biosciences Research Fellow of the Life Sciences Research Foundation.
ABBREVIATION
- DAP
days after pollination
Note Added in Proof
Luo et al. (27) show that the fertilization-independent seed1 (fis1) mutation is a mea allele.
References
- 1.Lopes M A, Larkins B A. Plant Cell. 1993;5:1383–1399. doi: 10.1105/tpc.5.10.1383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Mansfield S G. In: Arabidopsis: An Atlas of Morphology and Development. Bowman J, editor. New York: Springer; 1994. pp. 385–397. [Google Scholar]
- 3.Ohad N, Margossian L, Hsu Y-C, Williams C, Repetti P, Fischer R L. Proc Natl Acad Sci USA. 1996;93:5319–5324. doi: 10.1073/pnas.93.11.5319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Chaudhury A M, Ming L, Miller C, Craig S, Dennis E S, Peacock W J. Proc Natl Acad Sci USA. 1997;94:4223–4228. doi: 10.1073/pnas.94.8.4223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ohad, N., Yadegari, R., Kinoshita, T., Margossian, L., Hannon, M., Michaeli, D., Harada, J. J., Goldberg, R. B. & Fischer, R. L. (1999) Plant Cell11, in press. [DOI] [PMC free article] [PubMed]
- 6.Castle L A, Errampalli D, Atherton T L, Franzmann L H, Yoon E S, Meinke D W. Mol Gen Genet. 1993;241:504–514. doi: 10.1007/BF00279892. [DOI] [PubMed] [Google Scholar]
- 7.Grossniklaus U, Vielle-Calzada J-P, Hoeppner M A, Gagliano W B. Science. 1998;280:446–450. doi: 10.1126/science.280.5362.446. [DOI] [PubMed] [Google Scholar]
- 8.Jenuwein T, Laible G, Dorn R, Reuter G. Cell Mol Life Sci. 1998;54:80–93. doi: 10.1007/s000180050127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Leyser H M O, Pickett F B, Dharmasiri S, Estelle M. Plant J. 1996;10:403–413. doi: 10.1046/j.1365-313x.1996.10030403.x. [DOI] [PubMed] [Google Scholar]
- 10.Franzmann L H, Yoon E S, Meinke D W. Plant J. 1995;7:341–350. [Google Scholar]
- 11.Bent A F, Kunkel B N, Kahlbeck D, Brown K L, Schmidt R, Giraudat J, Leung J, Staskawicz B J. Science. 1994;265:1856–1860. doi: 10.1126/science.8091210. [DOI] [PubMed] [Google Scholar]
- 12.Konieczny A, Ausubel F M. Plant J. 1993;4:403–410. doi: 10.1046/j.1365-313x.1993.04020403.x. [DOI] [PubMed] [Google Scholar]
- 13.Smyth D R, Bowman J L, Meyerowitz E M. Plant Cell. 1990;2:755–767. doi: 10.1105/tpc.2.8.755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Shih M C, Heinrich P, Goodman H M. Gene. 1991;104:133–138. doi: 10.1016/0378-1119(91)90242-4. [DOI] [PubMed] [Google Scholar]
- 15.Lindsey K, Topping J F. J Exp Bot. 1993;44:359–374. [Google Scholar]
- 16.Meinke D W. Dev Genet. 1991;12:382–392. [Google Scholar]
- 17.Pirrotta V. Cell. 1998;93:333–336. doi: 10.1016/s0092-8674(00)81162-9. [DOI] [PubMed] [Google Scholar]
- 18.Jones C A, Ng J, Peterson A J, Morgan K, Simon J, Jones R S. Mol Cell Biol. 1998;18:2825–2834. doi: 10.1128/mcb.18.5.2825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.van Lohuizen M, Tijms M, Voncken J j, Schumacher A, Magnuson T, Wientjens E. Mol Cell Biol. 1998;18:3572–3579. doi: 10.1128/mcb.18.6.3572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Sewalt R G, van der Vlag J, Gunster M J, Hamer K M, den Blaauwen J L, Satijn D P, Hendrix T, van Driel R, Otte A P. Mol Cell Biol. 1998;18:3586–3595. doi: 10.1128/mcb.18.6.3586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Goodrich J, Puangsomlee P, Martin M, Long D, Meyerowitz E M, Coupland G. Nature (London) 1997;386:44–51. doi: 10.1038/386044a0. [DOI] [PubMed] [Google Scholar]
- 22.Martienssen R. Curr Opin Genet Dev. 1998;8:240–244. doi: 10.1016/s0959-437x(98)80147-0. [DOI] [PubMed] [Google Scholar]
- 23.Haig D, Westoby M. Am Nat. 1989;134:147–155. [Google Scholar]
- 24.Haig D, Westoby M. Phil Trans R Soc London B. 1991;333:1–14. [Google Scholar]
- 25.Scott R J, Spielman M, Bailey J, Dickinson H G. Development. 1998;125:3329–3341. doi: 10.1242/dev.125.17.3329. [DOI] [PubMed] [Google Scholar]
- 26.Koltunow A. Plant Cell. 1993;5:1425–1437. doi: 10.1105/tpc.5.10.1425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Luo M, Bilodeau P, Koltunow A, Dennis E S, Peacock W J, Chaudhury A M. Proc Natl Acad Sci USA. 1999;96:296–301. doi: 10.1073/pnas.96.1.296. [DOI] [PMC free article] [PubMed] [Google Scholar]