

Abstract
Theodor Boveri’s major intellectual contribution was his focus on the causality of nuclear chromosomal determinants for embryological development. His initial experimental attempt to demonstrate that the character of the developing embryo is determined by nuclear rather than cytoplasmic factors was launched in 1889. The experimental design was to fertilize enucleate sea urchin eggs with sperm of another species that produces a distinguishably different embryonic morphology. Boveri’s “hybrid merogone” experiment provided what he initially thought was empirical evidence for the nuclear control of development. However, for subtle reasons, the data were not interpretable and the experiment was repeated and contested. At the end of his life, Boveri was finally able to explain the technical difficulties that had beset the original experiment. However, by 1902 Boveri had carried out his famous polyspermy experiments, which provided decisive evidence for the role of nuclear chromosomal determinants in embryogenesis. Here we present the history of the hybrid merogone experiment as an important case of conceptual reasoning paired with (often difficult) experimental approaches. We then trace the further history of the merogone and normal species hybrid approaches that this experiment had set in train, and review their results from the standpoint of current insights. The history of Boveri’s hybrid merogone experiment suggests important lessons about the interplay between what we call “models”, the specific intellectual statements we conceive about how biology works, and the sometimes difficult task of generating experimental proof for these concepts.
Keywords: Hybrid merogones, Species hybrids, Genomic control, Sea urchin embryos
Introduction
In the spring of 1889, during his second visit to the Marine Zoological Station in Naples, Theodor Boveri performed a set of experiments with sea urchin eggs designed to address what he perceived as the fundamental question of biology: is the character of the developing embryo determined by nuclear chromosomal factors or by cytoplasmic factors? For many, including August Weismann, Wilhelm Roux, Carl Nägeli, and Oscar Hertwig the advances in cytology during the 1870s and 1880s—the observations of fertilization, the fusion of pronuclei, and the processes of cell division—all seemed to point to the nucleus as the causal force in embryogenesis. But, as Boveri noted in the first report of his 1889 Naples experiments, this claim, although widely accepted at that time, was still missing experimental verification (Boveri, 1889). Boveri, who certainly shared the theoretical conviction that the nucleus is causally relevant for determining embryological and adult characters, set out to solve this problem experimentally.
“Simple reflection shows moreover that the determination whether or not this Theory of Inheritance (Vererbungs-Theorie) is true, can be settled in one way alone, viz., to take two different sorts of cells, utilizing the nucleus of one and the protoplasm of the other, to form a new cell. If the nucleus and protoplasm are so constituted that they can exist together, then will the properties arising from this cell, made artificially, answer our question” (Boveri, 1893 [1889], p. 223, English translation by T.H. Morgan).
Shortly before, August Rauber (1886) had attempted to transplant the nucleus of a fertilized toad egg into the enucleated egg of a frog, and vice versa, but obtained no viable results. Boveri settled on sea urchin eggs as the experimental system on which to try a similar hybrid merogone approach. Two years earlier Boveri’s mentor Richard Hertwig had reported the results of a series of experiments performed together with his older brother Oscar (Hertwig and Hertwig, 1887). They observed that when they shook sea urchin eggs these eggs sometimes produced fragments that apparently lack a nucleus. And they furthermore demonstrated that these fragments could be fertilized and subsequently showed signs of cleavage. The Hertwigs, however, did not follow the development of these enucleated fertilized fragments, or merogones, any further.
For Boveri the possibility of fertilizing enucleated egg fragments represented a ‘natural experiment (Boveri, 1907)’ that made it possible to address the problem of nuclear versus cytoplasmatic control of cellular and embryonic characters; the sperm provides a natural delivery system for the nucleus. The requirements for experimental analysis of the theory of nuclear determination, then, would be two species that first, form a viable hybrid merogone and second, have sufficiently different embryonic characteristics so that it would be possible to clearly distinguish between nuclear, or paternal, and cytoplasmic, or maternal, characters. He found such a system in crosses between two sea urchin species from the Gulf of Naples, Echinus (now Psammechinus) microtuberculatus and Sphaerechinus granularis. These two species have clearly distinguishable larval forms and skeletal elements; the only drawback, as Boveri noted, was that the rate at which hybrid merogones could be generated is rather low, at less than 1 out of 1000.
Boveri reported the following results from the 1889 hybrid merogone experiments:
Normal hybrids between Sphaerechinus and Echinus always yield larvae with intermediate forms and skeletal elements.
Mass Fertilization of shaken Sphaerechinus eggs with Echinus sperm yields a mixture of larvae that includes (a) normal hybrids, which are the product of unaffected eggs; (b) smaller larvae of hybrid character, which are the product of egg fragments that contain a nucleus; and (c) smaller larvae that clearly resemble those of Echinus thus representing the paternal type (see Fig. 1).
Fig. 1.
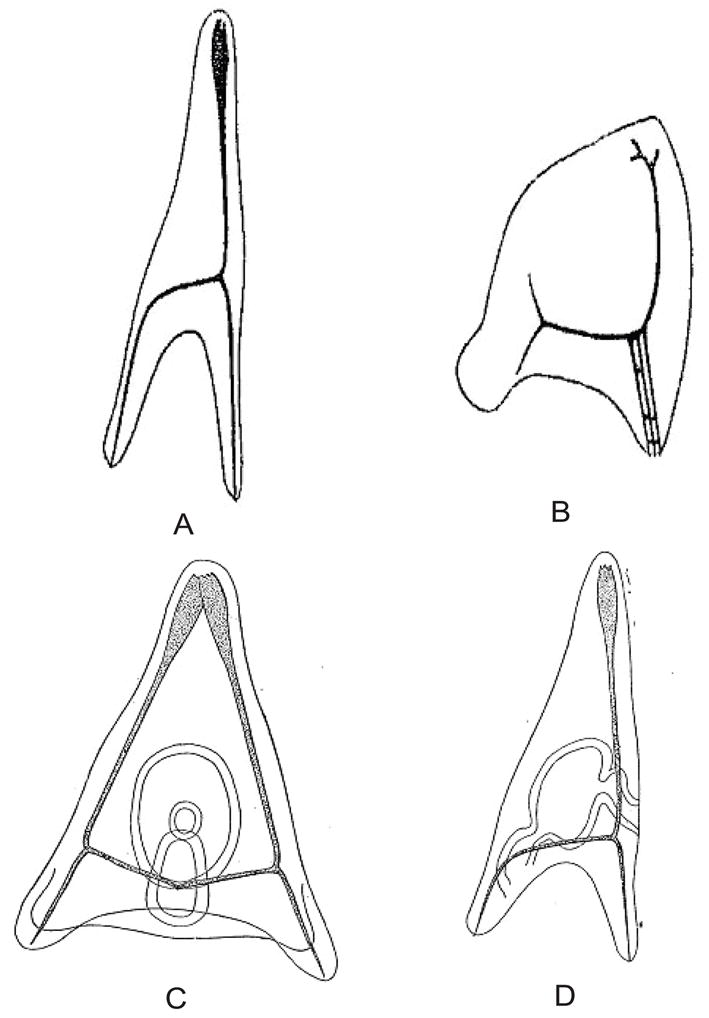
Drawings from the initial mergon experiment. (A) Normal Echninus pluteus. (B) Normal Sphearechinus pluteus. The plutei are shown upside down, as presented in the original paper; reproduced from Boveri, 1889. (C) Dwarf pluteus formed by insemination of a nucleated egg fragment Sphaerechinus with Echinus sperm. (D) Dwarf pluteus formed by insemination of an apparently enucleated Sphaerchinus egg fragment with Echinus sperm. Drawings from Boveri 1893 [1889], English translation of this paper by T.H. Morgan, which contains additional original drawings from Boveri.
The latter, Boveri deduced, must have been the product of fertilizing an enucleated Sphaerechinus fragment with Echinus sperm. As these larvae display only paternal characters the conclusion that the nucleus determines larval characters is thus supported. Boveri noted that a more direct experimental proof would involve rearing and fertilizing enucleated fragments in isolation. But due to the low fertilization rate of these hybrids he was unable to observe such an event during his 1889 stay at Naples. He did, however, offer an additional observation in support of this conclusion; the nucleus of those “bastard” larvae that clearly show the Echinus type was proportionally smaller than of those which showed the hybrid type. This observation is consistent with the assumption that only the sperm (Echinus) nucleus is present in those (presumably haploid) larvae.
Based on all these observation Boveri concluded in 1889 that the theory of nuclear determination is supported. His reasoning in the short report of his results represents a clear logical inference based on what he considered to be the right kind of observations and experiments needed to arrive at such a conclusion. Oscar and Richard Hertwig’s observation that enucleated egg fragments can be fertilized had suggested a way to deliver the nucleus into a cytoplasm; his own hybridization studies with the two sea urchin species (Sphaerechinus and Echinus) provided a baseline for the interpretations of the merogony results; and comparing the size of the nuclei in the different larvae that resulted from fertilizing shaken Sphaerechinus eggs established the predicted haploidy. But Boveri was well aware that he lacked direct proof, as he had not directly observed the fertilization of an enucleated egg, and he planned to return to Naples as soon as possible to repeat the experiments.
Not surprisingly, Boveri’s initial report of his results caused a sensation, as it seemed to show nothing less than that the nucleus is the determinant of cellular and embryological characteristics. But not everybody was convinced, and Boveri’s observations and interpretations were soon challenged. However, before discussing the further fate of Boveri’s 1889 experiment, it is useful to reconstruct briefly the conceptual and observational basis that had already led to the theory that nuclear determinants control embryonic development. This attractive theory, despite the absence of any truly pertinent direct evidence, produced the intellectual landscape on which the subsequent battles were fought.
After this background digression we return to the immediate critiques of Boveri’s 1889 experiment; and his decisive demonstration of nuclear chromosomal control more than a decade later by an entirely different protocol. But the difficulties with the hybrid merogone experiment continued to trouble Boveri, and we trace to the end of his life in 1916 his further considerations on this subject, and the explanation for the defects of the 1889 experiment. We then turn to the direct off-spring of Boveri’s hybrid merogone experiment, some of which extend all the way down to the “early modern” period of developmental biology in the 1950’s and 60’s. On the one hand, sea urchin merogones continued to be explored by better methods. On the other, a long and in many ways very fruitful exploration of species hybrid experiments ensued. It was learned what could be deduced from them, what could not, and what from a modern, mechanistic point of view actually underlay the observations that were reported. By the early modern period the species hybrid experiments had made two important contributions: they bolstered the argument for direct chromosomal control of embryonic developmental processes; and they pointed toward the discovery of maternal mRNA.
Earlier evidence: The prior embryological and cytological context of Boveri’s 1889 experiment
As Boveri mentioned in the 1889 paper, his actual experiment was motivated by a long series of earlier observations and theoretical speculations that had accumulated in cytology and embryology. Among those, several observations supported claims that the nucleus contains the hereditary determinants for development. One line of evidence came from the work of embryologists, such as Karl Ernst von Baer, Mauro Rusconi, Heinrich Rathke and Robert Remak, whose observations of the development of frog embryos clearly demonstrated the segmentation of the egg after fertilization. This observation was important, as it provided a crucial material continuity between development and heredity. The cells of the embryo descend directly from the fertilized egg, which, in turn, contains all the substances derived via the egg straight from the mother.
In this context it is important to remember that during the initial formulations of the cell theory in the late 1830s the processes of the origin and formation of new cells were not yet fully understood; indeed several different conceptions—from secretion and “crystallization” to fission were widely discussed (Harris, 1999). Today the origin of cells from other cells is associated with Rudolf Virchow’s dictum “omnis cellula e cellula (Virchow, 1858).” But it was actually Robert Remak who deserves most of the credit for discovering the details about the origin and formation of cells. In 19th century Germany Remak, an orthodox Jew of Polish descent who refused to be baptized, did not qualify for a distinguished professorship, despite the support he received from such dignitaries as Alexander von Humboldt. Remak and Virchow were both students of Johannes Müller in Berlin and had, initially at least, a collegial relationship with each other. By 1855 Remak had stated repeatedly his conviction that cells derive from other cells by binary fission and provided ample evidence to support his claim; however, it was Virchow in his Cellularpathologie (1858), who popularized this notion without giving Remak the credit he deserved. Both Remak and Virchow expanded their claims to pathology. In an earlier paper Remak concluded that “pathological tissues are not, any more than normal tissues, formed in an extracellular cytoblastem, but are the progeny or products of normal tissues in the organism.” This interpretation of pathological variation is also important for our concern here, as it implies that factors within the cells are responsible for differentiation and variation (including pathological variation.) The question, then, was which intracellular factors are involved in determining the specific features of the cells.
Once it was clear that cells originate through division from other cells the question of what happens to intracellular structures became the focus of attention. It was again Remak and others, such as Bagge, who observed in the 1840s and early 1850s that nuclear division precedes cell division. But there was less clarity about the actual mechanisms of nuclear division, which would remain an area of vibrant speculation during the following decades, as the details of both mitosis and meiosis were described in painstaking observations by Eduard Strasburger, Otto Bütschli and others (Harris, 1999). Of special interest were the mechanisms of longitudinal chromosomal division, that is, the replication of chromosomes at mitosis such that each element along the length of the chromosome is duplicated in its two daughters. However, as long as there was no direct conceptual link to patterns of heredity, the peculiar features of this process did not carry much significance. This changed after Hermann Fol observed in 1876 the fertilization of an egg by a single sperm (in both sea stars and sea urchins), something that Oscar Hertwig had already deduced from observations on eggs of the sea urchin Toxopneustes lividus a year earlier (Hertwig, 1875). In subsequent years there was much speculation about the fate of the two pronuclei and their role in the process of fertilization. Did they, for instance, fuse to form what Oscar Hertwig called a ‘Furchungskeim (cleavage nucleus)’? It was Edouard van Beneden who in 1883 brought these questions into focus. In a classic paper on Ascaris van Beneden concluded that fertilization is complete with the formation of two (one male one female) nuclear elements within the female egg and that there is no subsequent fusion between the male and the female chromatin (van Beneden, 1883). The latter conclusion was also expanded to suggest that chromosomal elements of male and female origin remain distinct in all subsequent nuclei derived from them. Boveri would spend a good part of his scientific career researching the implications of this suggestion. What is important for our question here is that the proposed individuality of male and female chromosomal elements suggested a correlation between these hereditary elements and the observed expression of paternal and maternal traits in the offspring.
Even though almost all of the early work in cytology and embryology up to the 1880’s was descriptive, these studies also introduced different organisms as experimental systems (Lederman and Burian, 1993; Rheinberger, 1997). Various species of nematodes (Ascaris) and sea urchins and their embryos proved to be essential for further progress in cytology and embryology. Note that by this time the scientists participating in this discourse realized that in processes which are this fundamental, what is true for any one species would be relevant to any other species. The new experimental orientation of biology, basing conclusions on observation, contrasted with the traditional fondness for grand speculation, as then exemplified by Ernst Haeckel. Nevertheless, it was Haeckel who first suggested in his ‘Generelle Morphologie’ (1866) that the nucleus contains the hereditary material, while the cytoplasm is responsible for accommodating the cell to its environment. Besides Haeckel’s speculations there was also a solid argument for the role of the nucleus in controlling development. This included Carl Nägeli’s conclusion, based on his own and Kölreuter’s observations of the equal contributions of the egg and the sperm to heredity, that the totality of the cytoplasm could not be involved in heredity. Nägeli introduced the term idioplasm for what he postulated to be the small part of the cellular material that is involved in heredity. A year earlier, Wilhelm Roux, the founder of Entwicklungsmechanik (developmental mechanics), had already claimed that the longitudinal division of the chromosomes not only assured the preservation of the quantity, but also the discrete quality of the nuclear material. It is important, he argued, for each daughter cell to receive the same hereditary factors. Roux, together with August Weismann, would later famously argue for qualitative differences in chromosomal division to account for embryological differentiation, but he never questioned that the hereditary substance lies inside the nucleus.
To sum up, by the time Boveri decided to address the problem of the nuclear control of development and heredity experimentally, a growing consensus had emerged that interpreted the known observations of cytology—the material continuity between the egg and the embryo, the cellular origins of pathologies, the integrity of maternal and paternal chromosomes, etc.—along these lines. But, by the late 1880s all these ideas were yet based on interpretation and speculation; there was no experimental proof, and alternative explanations that focused on the cytoplasm as the causal agent in development and heredity were still a logical possibility, especially as long as neo-Lamarckian explanations for the origin of phenotypic variation were still considered seriously.
The immediate reception and critique of Boveri’s experiment
Though Boveri’s 1889 paper promised to confirm experimentally what many of the leading researchers of the day already believed conceptually, namely that the nucleus controls development, these were only preliminary results. Soon several serious objections would be raised. Thomas Hunt Morgan set the tone in 1893 (see Gilbert, 1978; 1987, for discussions of Morgan’s response to Boveri). In the preface to his English translation of Boveri’s initial report Morgan acknowledged the importance of Boveri’s project and of his paper, “which will certainly become a classic”, but he also insisted “results of such importance must be verified over and over” (Boveri [Morgan] 1893, p. 222).
The first to raise specific objections was Oswald Seeliger (1894;1896) soon to be followed by Morgan (1895) and later also by Driesch (1903). In addition, Delage (1899) stated serious concerns about Boveri’s method for obtaining enucleated egg fragments. These criticisms, and there were others which offered similar objections, questioned several of the core premises of Boveri’s experiment. First, both Seeliger and Morgan claimed that they were unable to obtain similar results as Boveri. They also reported that they saw a much greater degree of variability in larval forms, in both pure and hybrid crosses. This, they suggested, seriously undermined Boveri’s experimental premises, as he claimed that he could clearly distinguish between hybrid larvae, dwarf hybrid larvae and dwarf paternal larvae. If this is in fact not possible, Boveri’s results must be considered inconclusive. Another issue concerned the size of the nuclei, a crucial element in Boveri’s arguments, as this allowed him to ascertain whether the dwarf paternal larvae were haploid and therefore only contained a paternal nucleus. Seeliger and Morgan claimed that the size of the nucleus also shows great variability and is therefore not a reliable criterion for haploidy. Finally, Delage’s observation that shaking of eggs can lead to the fragmentation of the nucleus and therefore to the presence of maternal nuclear material in supposedly enucleated egg fragments was probably the most serious objection, as there was no easy way to control for this effect nor to test for the presence of fragments of maternal nuclear material.
Boveri’s responses to his critics
Looking at Boveri’s substantial oeuvre it is difficult to imagine that his research was frequently interrupted by illness and finally cut short by an untimely death in 1916. These interruptions are nowhere more serious than in the controversy following the initial publication of his hybrid merogone experiment. The first paper was intended as a preliminary publication in a venue with limited circulation, as Morgan noted in the preface to his English translation. Returning from Naples, Boveri reported his result at a meeting of the Munich Society for Morphology and Physiology (Gesellschaft für Morphologie und Physiologie) on July 16, 1889 with 29 people in attendance. As was the custom of this society, the presentation was published in their proceedings (Boveri 1889). Boveri had every intention of returning to Naples the following spring to continue his experiments, hoping, for instance, to observe direct fertilization of an enucleated egg fragment. Illness and later his move to Würzburg interfered, so by the time the first criticisms were published in 1894/1895, Boveri had not been able to obtain any additional data. In addition, his plans to prepare a more substantial publication based on his first set of experimental results were severely complicated by that fact due to an acid contamination in the fixative he had used, the skeletal elements of the embryos on his slides had dissolved. He thus decided to wait until he could obtain new material, and was only pressed to publish a more substantial account (still without new material) by the barrage of criticism in 1895.
In this first response (Boveri, 1895) he mainly stood his ground, correcting what he perceived as misunderstandings of his claims and experimental procedures and giving a more detailed account of his data. He acknowledged that Morgan and Seeliger were unable to repeat his experiments and that they had seen a greater variation in larval forms, but he insisted that his logic as well as his experimental procedure were sound and that the observed results, especially the paternal-type dwarf larvae, were an indication of nuclear control of development. As Seeliger was working in Trieste rather than Naples, Boveri also suggested that variability between different populations might account for some of the discrepancies. He concluded that the basic problem, providing an experimental solution to the problem of nuclear control of development, can be addressed with this system, something he was planning to do as soon as he would be able to return to Naples. But he also acknowledged that his preliminary results were not as conclusive as he had originally hoped. Over the next two decades Boveri continued to defend his experiment. He was, for example, involved in a “priority dispute” with Delage, who coined the term “merogony” without specifically acknowledging the significance of Boveri’s earlier experiment as the first instance of this phenomenon. But he also began to realize the problems with his method, especially the fact that shaking might not lead to truly enucleated fragments.
But the hybrid merogone experiment was scarcely the only approach that Boveri took to address the problem of nuclear control. His whole scientific career can be seen as one long argument for the importance of the nucleus and the chromosomes in development and heredity (Baltzer, 1967; Moritz, 1993). In this context his sea urchin embryo polyspermy experiments, arguably, his most famous set of experiments, were decisive (Boveri, 1902, 1905, 1907). These experiments illustrate again Boveri’s unique combination of conceptual understanding, logical clarity and experimental skills. Boveri was convinced that a complete set of qualitatively distinct chromosomes is required for embryonic development; indeed, this is the proposition underlying the haploid development he thought he was seeing in the 1889 experiment. As shown clearly only much later by Horstadius, Harvey, and then Von Ubisch, haploid sea urchin embryos may develop (see below). The polyspermy experiment was designed specifically to test the idea that a complete set of chromosomes must be present in every cell of the embryo for normal development to occur. The premises were: first, that the chromosomes are qualitatively distinct and play distinct roles; second, that there are a sufficient number of these roles that any missing chromosomes in any of the cells of the embryo would cause developmental deficiencies on the part of those cells; third, that the developmental functions of every cell in the embryo depend on chromosomal determinants.
The sea urchin embryo offered an elegant experimental approach to this question. From his earlier studies on nuclear division, Boveri knew that each sperm nucleus forms its own aster and spindle apparatus. When two or three sperm enter a single egg di- and tri-polar mitoses form, and Boveri realized that the embryos which derive from these eggs are mosaics of blastomeres which contain different and usually incomplete sets of chromosomes. He carefully showed that different portions of such embryos fail to develop properly, and only the small fraction of embryos in which all blastomeres have inherited a complete chromosome set develop normally. He further showed that quantitatively, this fraction conforms to the predicted probabilities of normal, complete chromosome sets in dipolar and tri-polar eggs, respectively. An illustration from these experiments, and a recently made photomicrograph of one of Boveri’s original slides from this experiment can be seen in Fig., 2. Boveri drew the correct conclusion: in modern terms, different developmental defects occurred in embryonic clones that lacked different chromosomes; therefore a complete genome is required for normal development in every cell; therefore the genome provides inputs essential for development in every cell.
But despite this independent and widely acknowledged experimental verification of his theoretical convictions, Boveri continued to work on the hybrid merogone experiment. During visits to Naples in 1910/11 and 1914 Boveri, together with his wife, attempted to resolve the problems with the experiment. His final words on this matter were published posthumously in 1918. There he had to acknowledge that his initial conclusions could not be maintained (Boveri, 1918). Under his experimental conditions it is almost certain that nuclear material remains in the egg fragments; therefore the resulting larvae cannot be considered true merogones. With regard to the 1889 experiment, this was a devastating conclusion. However, the status of the underlying basic problem, the role of the nucleus during development and differentiation, was now entirely different, due to the accumulated evidence both from experimental embryology and genetics. Reviewing his own and others’ studies, Boveri concluded that there was decisive support for the theory that nuclear chromosomal determinants are required for differentiation and development. Boveri’s final sentence, before the manuscript abruptly ends, is rather prescient: “We have to conclude that inside the nucleus are combined different [genetic] parts each of which have different roles … (Boveri, 1918, p. 469).”
Some years after Boveri’s death his friend E.B. Wilson provided the then final summary of the details and implications of the merogony experiment in the third edition of The Cell in Development and Heredity (Wilson, 1925). As he had in every previous edition, Wilson emphasizes the significance and importance of Boveri’s results. As early as in the first edition he had noted that: “should they be positively confirmed, they would furnish practical demonstration of inheritance through the nucleus. (Wilson, 1896, p. 258).” And in the second edition, summarizing many of the problems raised by Seeliger, Morgan, and Driesch, he recognizes clearly the importance of an experimental approach to this important problem of development and inheritance, stating, that: “although his conclusions do not rest on absolutely certain ground, they at least open the way to a decisive test” and he concludes “it is to be hoped that this highly ingenious experiment may be repeated on other forms which may afford a decisive result (Wilson, 1900, p. 353).” In the third edition Wilson gives by far the most extensive treatment of the implications of the merogony experiment, discussing at length Boveri’s own conclusions in his final paper (Boveri, 1918). But Wilson then added a new section devoted specifically to the question of the determinant role of the chromosomes in development. Here in the context of his final discussion on “The mechanisms of development” Boveri’s now classic polyspermy experiment plays a central role.
Wilson’s analysis of the problem of nuclear vs. cytoplasmic determination displays a subtle shift in orientation that motivated much of the following work on hybrid merogones and on species hybrids. Many studies undertaken in the wake of Boveri’s initial experiment seemed to indicate the existence of a period early in embryogenesis controlled by maternal determinants, while paternal characteristics became visible only later in development. This was taken as evidence that there is a period of cytoplasmic control, followed by the onset of nuclear activity. This distinction was taken by some, such as Jacques Loeb and Edwin Grant Conklin, to imply that the cytoplasm is the carrier of species-specific hereditary factors, whereas the later activity of the nuclear factors of both parents accounts for the adult individual’s traits. But, as Wilson points out, Boveri realized that the cytoplasmic organization of the egg is actually “itself the product of an antecedent process of development” (i.e., oogenesis) “in the course of which, as we have every reason to believe, the chromosomes have played their part” (Wilson, 1925, p. 1108). For Boveri the chromosomes were the primary determinants of heredity and development throughout the whole life cycle.
The eventual solution to the technical problems of sea urchin hybrid merogones; how Boveri was both right and wrong
Hybrid merogones continued to be studied, but now the emphasis was on the extent of cytoplasmic inheritance in development, an argument that extended as well into the newly established field of genetics. In Germany, in particular, studies devoted to cytoplasmic inheritance attracted much attention (Harwood, 1994; Sapp, 1987). More often than not these studies were tied to theoretical debates about reductionism, mostly associated with the paradigm of chromosomal nuclear genes, vs. various forms of biological holism. None of these subsequent debates shared the conceptual clarity of Boveri’s experiments and interpretations, though they raised issues that kept the debate on nuclear vs cytoplasmic and organismal determinants of development alive. But by the late 1920s and early 1930s, the conclusions Boveri and Wilson had drawn seemed to have been buried and forgotten.
Study of sea urchin merogones advanced mainly through the introduction of new methods. To address the problem of generating truly enucleate eggs, Sven Hörstadius adopted microsurgery techniques, first developed by Hans Spemann, to remove the small portion of the egg that clearly contains the nucleus (Hörstadius, 1936). Initial attempts to produce true hybrid merogones with the same combination of species used by Boveri (Sphaerechinus and Echinus=Psammechinus) failed to produce larvae that developed to the point that distinct skeletal elements could be compared. But Hörstadius used less than 100 Sphaerechinus eggs in these experiments and his lack of success was well within the range of expectation reported by Boveri.
However, Hörstadius did succeed in creating hybrid merogones using a different species pair (Psammechinus microtuberculatus and Paracentrotus (originally called Strongylocentrotus) lividus). The problem with these species, as Boveri had already noted, is that their larval stages are very similar morphologically, and thus not suitable for the clear distinction of paternal characteristics. An additional complication in Hörstadius’ experiments was that different temperatures influenced the shape and other skeletal characteristics of the larvae, thus complicating interpretation of the results. Hörstadius thus was able to produce reliable merogones containing solely the paternal genome, but he failed to resolve the roles of nuclear vs. cytoplasmatic factors.
E. B Harvey (1933, 1940, 1951) introduced centrifugation as another technique for producing unequivocally enucleate egg halves, and then fertilized these with sperm of the same and other species. Her fertilized merogones underwent cleavage but thereafter developed poorly, neither intraspecific fertilized merogones nor interspecific hybrids generated the elegant skeletal structures figured in Boveri’s (1889) account, except rarely. In 1933 she was able to obtain plutei from enucleated egg fragments of Sphaerechinus fertilized with Paracentrotus sperm, but she did not describe their skeletal features in great detail. A year later, in 1934, with the help of Harvey, von Ubisch used this same technique to create hybrid merogones of Sphaerechinus and Echinus at the Naples Station. His results were again ambiguous, but most importantly he discovered that with centrifugation, as with shaking, the nucleus can break and therefore chromosomal elements could be present in the supposedly enucleated half of the egg. Thus by the mid 1930s the study of hybrid merogones between Sphaerechinus and Echinus was not significantly advanced beyond Boveri’s conclusions of 1918, though there had been some progress using other species.
But the story of Boveri’s experiment does not end here. After the interruptions caused by the political situation of the 1930s and WWII were finally past, von Ubisch returned to Naples in 1953, in order to once again attempt to produce hybrid merogones using Sphaerechinus and Echinus (Psammechinus) and Paracentrotus. This time he used his own technique for obtaining enucleated eggs, by microsurgically removing the nucleus (see von Ubisch, 1954 for detailed description). He was now able to increase the success rate of fertilization and subsequent development. From 2905 fertilized enucleated Sphaerechinus egg fragments (with either Echinus or Paracentrotus sperm) he was able to obtain 4 pre-pluteus and 16 pluteus larvae, some of which survived for 46 days. The results seemed to confirm some of Boveri’s initial observations, including a frequency of success of only 10−3, although it is of course, impossible to know what Boveri really saw. However, von Ubisch proved that he had generated true merogones, i.e., the larvae were haploid (although their nuclei were larger than expectation for haploids, which von Ubisch thought due to their interaction with Sphaerechinus cytoplasm). Most importantly von Ubisch was able to observe a haploid mitosis. The skeletons of the hybrid merogones were strictly paternal. Von Ubisch published his results in 1954, exactly 65 years after Boveri’s initial report. Ironically, he had proved Boveri’s 1899 theoretical expectation exactly correct, while at the same time his technological advances made it completely clear that Boveri could never have obtained that result in a convincing way by the experimental method he had used.
Boveri’s hybrid merogone experiment also led to a whole series of related experiments on amphibian eggs. These were more successful than the sea urchin merogony experiments, because the amphibian egg is more robust and allows for easier microsurgical removal of its nucleus or the insertion of a different nucleus. Indirectly, the fundamental logic of the merogony experiment contributed as well to the development of nuclear transfer (Briggs and King, 1952; 1957; Fishberg et al., 1958), and the subsequent advances in animal cloning. The historical link is evidenced by the suggestion of Michael Fishberg to his then graduate student John Gurdon that he should work on a merogony experiment for part of his thesis (John Gurdon, personal communication).
The echinoid species hybrid experiments
In the terms of long term scientific progress, another irony is that the control for Boveri’s 1889 experiment was of more importance than the experiment. For the control was a straight species hybrid experiment, which generated larvae of intermediate type. This by itself was excellent evidence for the role of chromosomal determinants since the sperm carries almost no other components in to the egg. In fact, lost in the debate over the veracity and reproducibility of Boveri’s hybrid merogone experiment was the point that even if he was in fact just making species hybrids rather than merogone hybrids, the intermediate and incompletely paternal forms he reported were also perfectly good evidence for chromosomal determinants of embryonic form. Soon after Boveri’s contentious hybrid merogone experiment, it occurred to a number of people in this field that there is an easier way to approach the same question: simply make hybrids using normal eggs of one species and sperm of another, the embryonic anatomy of which differed observably from that of the maternal species. For example Driesch (1898) crossed Psammechinus (then called Echinus) microtuberculatus and Spherechinus granularis reciprocally, and showed that the skeletal forms of the hybrid plutei were intermediate in morphology with respect to the parental forms. But on the other hand, he also noted that the number of skeletogenic micromeres is quantitatively exactly that of the maternal species in such crosses. These two conclusions nicely illustrate the dual significance of echinoderm species hybrid experiments in the classical period of what we would term today “genome oriented developmental biology.” The two objectives were of course intertwined: (1) to prove that chromosomal genetic determinants indeed control embryonic development; but (2) to determine when in development of the embryo genomic control takes over, since in all these experiments and many others done with other organisms only maternal species characters were observed at the earlier stages. Echinoderm species hybrid experiments continued to be done throughout this period and thereafter (see Harvey, 1956, for a comprehensive review).
In the first of these objectives, demonstration of the genomic control of development at least from gastrula on, the species hybrid experiments were decisively successful and their import incontrovertible. A major confusing factor that had confounded interpretation of some species hybrid results was resolved by Boveri’s student Baltzer (1910) (who was still studying echinoid species hybrids almost a half century later; e.g., Whiteley and Baltzer, 1958). Baltzer’s careful observations showed that in some species pairs the paternal set of chromosomes is thrown out of the mitotic apparatus during cleavage, while in others this does not happen. This had two consequences. First, it enabled consistent interpretations of the development of maternal vs. intermediate phenotypes; and second, it underlined the fundamental point that if you wanted to understand what is going on, you had better focus on the embryo chromosomes. In the 1950s and early 60s this main and inescapable conclusion was one of the foundation stones on which the modern science of genome oriented development was constructed. Boveri’s polyspermy experiment was another foundation stone
What we now would term Gain Of Function species hybrid demonstrations were the most convincing in this respect. Tennent (1914) produced a beautiful demonstration of this kind. He fertilized eggs of a cidaroid sea urchin with sperm of a typical thin-spined (euechinoid) species. The euechinoids diverged from the cidaroids (“pencil urchins”) after the great Permian Triassic extinction 250 million years ago, and they differ from the latter in many ways. Among these is that the skeletogenic cells of cidaroid sea urchin embryos arise together with other mesodermal cell types only at gastrulation, and produce skeleton only beginning late in postgastrular development (Tennent, 1914; Schroeder, 1981; Wray and McClay, 1988). Cidaroids lack a population of skeletogenic micromere descendants which ingress into the blastocoel prior to gastrulation, among the prominent features of embryogenesis in euechinoid sea urchins. In Tennent’s cideroid-euechinoid hybrids the precociously invaginating skeletogenic mesenchyme characteristic of the male species is generated on the euechinoid schedule, and in the euechinoid pattern, the causal output of the sperm genome. Thus the classical period ended with the basic, fundamental, and general truth established that genomes control embryonic development, and this conclusion provided the starting point for the current solution of the mechanism of development.
The classical idea that embryogenesis is initially controlled by maternal constituents inherited in the egg cytoplasm, another direct objective of the species hybrid experiments, also foreshadowed very important modern discoveries made only after 1965. Maternal mRNA was discovered and authenticated in sea urchin eggs in the 1960’s (reviewed by Davidson, 1968). Maternal transcripts are transcribed and stored in a stable form during oogenesis. It is general to all animal eggs as are a multitude of other RNAs and proteins, all the constituents of cells and their organelles. Furthermore, most animal eggs utilize some form of anisotropically localized, cytoplasmic gene regulatory factors of maternal origin, which directly or indirectly affect gene expression, to set up the initial functional asymmetries of the embryo (Davidson, 2006). Classical concepts of the dominance in early development of maternal, cytoplasmically borne mechanisms were qualitatively valuable, in that they immediately and directly influenced modern considerations of regulatory egg organization (see e.g., Davidson, 1986, Chapter 6; Davidson, 1990).
However, the more quantitative question that many of the species hybrid experiments were designed to address, the point in embryogenesis when genomic control becomes dominant, was never answered correctly by this means. There is a design issue which causes this to be an intrinsically slippery slope: as Davidson (1968) commented in reviewing these experiments, “Hybrids between closely related species which may be the most likely to succeed… are also the least likely to display early hybrid genome control over morphogenesis, since… the mechanics of early development will of course tend to be more similar the more closely related are the species.” But in 1968 we had no way of knowing how far off were the results of classical species hybrid experiments. They generally yielded the conclusion that maternal (i.e., cytoplasmic) dominance extends up to the late blastula period or the onset of gastrulation. But now we know: the slippery slope resulted in a wrong answer, in that genomic control of the events of early echinoid development begins right away, in earliest cleavage.
Contemporary retrospective
As a final point, it is informative to look back and reconsider the relation between the underlying mechanisms and the experimental observations made in the course of more than a half-century of echinoid species hybrid experiments, beginning with Boveri’s 1889 control. Short of molecular level explication of the developmental process in terms of the genomic regulatory program, embryological experiments are all phenomenological. But we can now know what was the real meaning of the phenomena our predecessors studied so intensively in the species hybrid experiments.
Three illustrative questions are (1) When do the embryo genomes actually assume control of development; (2) How is the formation of skeletogenic micromeres controlled; (3) What accounts for the form of the skeleton? These issues all figured largely in classical considerations and in the arguments that for many years swirled around the species hybrid experiments.
The greatest failure of the species hybrid experiments, as noted above, was in respect to the first of these matters, the timing of onset of genomic control. In sea urchins the embryo genomes open up transcriptionally even before fertilization and within a few cleavages the rate of RNA synthesis per nucleus is as high as it ever becomes (reviewed by Davidson, 1986). The gene regulatory network now available for this embryonic system (Davidson, 2006; for current version see http://sugp.caltech.edu/endomes/) provides dozens of examples of regulatory genes whose transcriptional expression during cleavage is essential for specific aspects of embryonic development, in the skeletogenic territory and in the endomesodermal territories from which the gut and mesodermal cell types much later develop. The regulatory state specification of these territories takes place while to an observer lacking molecular markers the embryo appears a largely featureless ball of cells, later on a hollow ball. Lack of tools for observation, and lack of suspicion that there might be more than meets the eye, compounded with the design flaw noted above, resulted in the wrong prediction that embryo genomes only begin to affect development about the time of gastrulation.
The most completely understood aspect of the endomesodermal gene regulatory network is that portion depicting the mechanism by which the skeletogenic micromeres are specified, and then differentiate by expression of gene batteries encoding biomineralization and other proteins (Oliveri and Davidson, 2004; http://sugp.caltech.edu/endomes/). This much is well understood. But with regard to species differences, there has been little modern exploration. No one yet knows why in some species there are three further divisions of the micromeres prior to ingression and in others four, the difference exploited by Driesch in his 1898 experiment. Nor have the considerable differences in skeletogenic process in cidaroid sea urchins with respect to the euechinoids that were exploited by Tennent (1914) been much analyzed, though it is clear that the basic aspects of skeletogenesis are similar (Wray and McClay, 1988).
Boveri, Driesch, Tennent, Baltzer and many other experimentalists relied on the species specific morphologies of the late embryo skeleton as the main readout in their echinoid species hybrid experiments. Herein lies a further tale. We now know that the micromere lineage determines the structure of the skeletal rods themselves, whether they are smooth or fenestrated, for example. But the shape of the skeleton, where its arms go and what the angles they form, is determined by interactions between the migratory skeletogenic mesenchyme cells and the ectodermal wall over which they crawl (Armstrong et al., 1993; Armstrong and McClay, 1994). The skeletogenic cells evidently read signals expressed by ectodermal cells at the correct locations, and thereby determine their disposition and thus the form of the rods they produce. Molecular analysis is closing in on the processes of ectoderm specification and the signaling to which the skeletogenic cells respond, and the details of this morphogenetic interaction will not remain unknown for long. It was the externally perceptible outcome of these processes that classical experimentalists took as their indicators of genomic control of development, and in this conclusion they were exactly right. We can see that there are many regulatory components to skeletal morphogenesis, and in the event it was an excellent metric.
Conclusion
The hybrid merogone experiment and the associated problem of nuclear control of development preoccupied Boveri throughout his entire career. While he was unable to provide the simple and elegant solution to the problem for which he designed his initial experiment, he nevertheless set in motion a research program that contributed decisively to both the experimental and conceptual clarification of the initial problem, and that ultimately provided crucial evidence for the role of chromosomes in development and heredity.
Boveri’s experiments demonstrate the fundamental importance of conceptualization in science. In approaching the problem in the data poor, relatively confused scientific environment of 1889, he clearly summarized the logically possible alternative explanations of development, and designed an experiment that would in theory have provided clear-cut experimental evidence for one or the other position. In this light he interpreted the first set of data as providing support for nuclear control. As it subsequently became clear that the complexities of the experimental system did not allow for an unambiguous interpretation of his data, Boveri continued to follow his “logic rules.” He designed additional experiments that provided further evidence for the role of the nucleus, and more specifically the chromosomes, in controlling differentiation. The famous polyspermy experiment proving unequivocally that a complete set of chromosomes is required for embryological development did just that. He also continued to work intermittently on his original experiment, searching for factual reasons why his initial elegant design did not yield clear-cut results.. As he noted in his last paper in 1918 the reasons are complications due to the complexities of the experimental system, and cannot affect the underlying logic supporting the conceptual arguments for nuclear control of development.
With the perspective of a century Boveri’s long experiment represents an exemplary case for theoretical and experimental biology. It began a conceptual pathway which led to the modern explanation of development in terms of genomic functions. By 1902–1904 the alternative approach it had forced on Boveri had provided experimental evidence that was incontrovertible and could not be ignored. In the 21st century biology of the field that Boveri initiated, genomic control of development, everything has changed except for the fundamental relationship between concept and experiment. What has changed most in recent times is the enormous, unforeseen enhancement in technological capabilities for large-scale measurement. But, as current literature amply demonstrates, masses of measurements that are not illuminated by concept fail utterly to generate novel insights into developmental processes. Effective use of modern experimental technologies requires conceptual guidance and logical construction of experimental perturbation protocols, just as Boveri used concept in the design of his polyspermy experiment. But in following a conceptual pathway to new experimental knowledge, the previous concepts are a living part of the process. Thus the confrontation of apparently contradictory facts and arguments frequently requires retreat to the last conceptual position than can be regarded as bed-rock. Conceptual advance, and conceptual history, can never truly be separated.
Supplementary Material
Fig. 2.
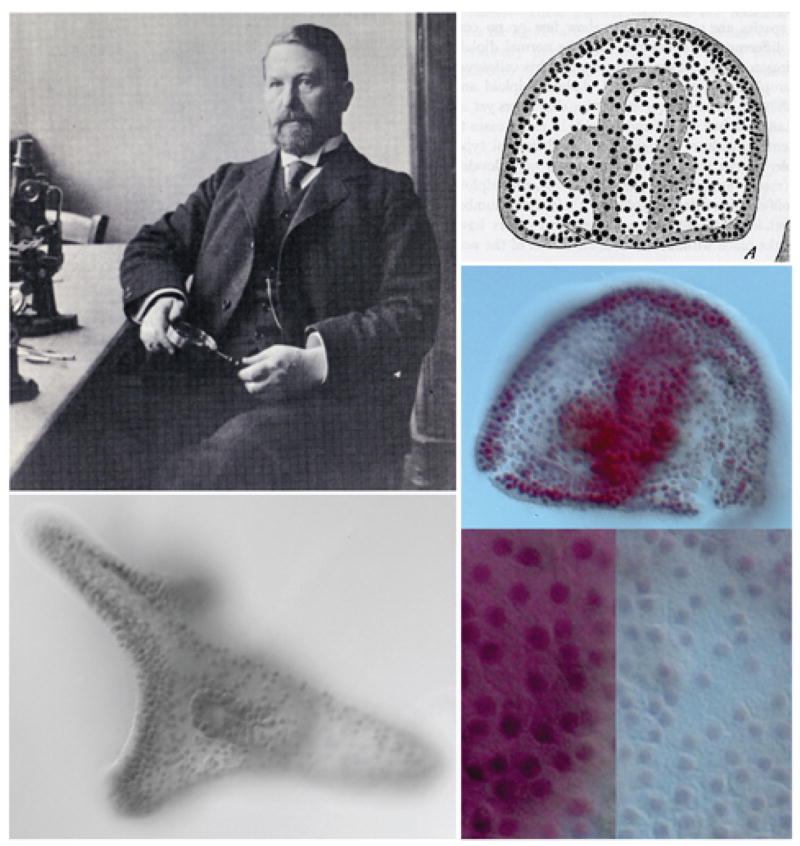
Provided by Dr. Andrew Ransick. Boveri at his laboratory at the Institute of Zoology in Würzburg in 1907, reproduced from Baltzer, 1967. Boveri’s drawing at upper right shows an abnormal aneuploid gastrula stage embryo containing nuclei of two different sizes. Below is the same embryo, photographed by Dr. Andrew Ransick from one of Boveri’s original 1904 slides, which had been kindly provided to the author by Professor Ulrich Scheer, University of Würzburg. In the magnification below the smaller nuclei in the blue region to the right can clearly be distinguished. Lower left shows an approximately normal pluteus larva from the same series of experiments.
Acknowledgments
We would like to thank Jane Maienschein for valuable feedback, and Jed Buchwald for suggesting our collaboration. This work has been supported by the following grants: NSF SES 0645729 and NSF SES 0623176 to MDL and NIH grant HD-37105 to EHD.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Armstrong N, McClay DR. Skeletal pattern is specified autonomously by the primary mesenchyme cells in sea urchin embryos. Dev Biol. 1994;162:329–338. doi: 10.1006/dbio.1994.1090. [DOI] [PubMed] [Google Scholar]
- Armstrong N, Hardin J, McClay DR. Cell-cell interactions regulate skeleton formation in the sea urchin embryo. Development. 1993;119:833–840. doi: 10.1242/dev.119.3.833. [DOI] [PubMed] [Google Scholar]
- Baltzer F. Über die Beziehung zwischen dem Chromatin und der Entwicklung und Vererbungsrichtung bei Echinodermenbastarden. Arch Zellforsch. 1910;5:497–621. [Google Scholar]
- Baltzer F. Theodor Boveri Life and Work of a Great Biologist. University of California Press; Berkeley and Los Angeles: 1967. [Google Scholar]
- Boveri T. Ein geschlechtlich erzeugter Organismus ohne mütterliche Eigenschaften. Sitz Gesel Morph u Physiol Müchen 5, 73–83 Trans by T H Morgan 1893, as “An organism produced sexually without characteristics of the mother”. Am Naturalist. 1889;27:222–232. [Google Scholar]
- Boveri T. Über die Befruchtungs- und Entwicklungsfähigkeit kernloser Seeigel -Eier und über die Möglichkeit ihrer Bastardirung. Archiv für Entwicklungsmechanik der Organismen. 1895;2:394–443. [Google Scholar]
- Boveri T. Merogoneoie (Y. Delange) und Ephebogenesis (B. Rawitz), neue Namen für eine alte Sache. Anatomischer Anzeiger. 1901;19:156–172. [Google Scholar]
- Boveri T. Über Mehrpolige Mitosen als Mittel zur Analyse des Zellkerns. Verhandlungen der Physikalische-medizinischen Gesellschaft zu Würzburg. 1902;35:67–90. [Google Scholar]
- Boveri T. Noch ein Wort über Seeigelbastarde. Archiv für Entwicklungsmechanik der Organismen. 1904;17:521–525. [Google Scholar]
- Boveri T. In: Die Entwicklung dispermer Seeigeleier Ein Beitrag zur Befruchtungslehre und zur Theorie des Kerns Gustav. Zellenstudien VI, editor. Fischer; Jena: 1907. [Google Scholar]
- Boveri T. Zwei Fehlerquellen bei Merogoneieversuchen und die Entwicklungsfähigheit merogoneischer und partiellmerogoneischer Seigelbastarde. Arch Entwicklungsmech Organ. 1918;44:417–471. [Google Scholar]
- Briggs R, King TJ. Transplantation of living nuclei from blastula cells into enucleated frogs eggs. Proc Natl Acad Sci USA. 1952;38:455–463. doi: 10.1073/pnas.38.5.455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Briggs R, King TJ. Changes in the nuclei of differentiating endoderm cells as revealed by nuclear transplantation. Journal of Morphology. 1957;100:269–311. [Google Scholar]
- Davidson EH. Gene Activity in Early Development. Academic Press; New York, NY: 1968. [Google Scholar]
- Davidson EH. Gene Activity in Early Development. 3. Academic Press; Orlando, Florida: 1986. [Google Scholar]
- Davidson EH. How embryos work: A comparative view of diverse modes of cell fate specification. Development. 1990;108:365–389. doi: 10.1242/dev.108.3.365. [DOI] [PubMed] [Google Scholar]
- Davidson EH. The Regulatory Genome Gene Regulatory Networks in Development and Evolution. Academic Press/Elsevier; San Diego, CA: 2006. [Google Scholar]
- Delage Y. Etudes sur la merogoneie. Arch zool exp et gen. 1899;7 [Google Scholar]
- Driesch H. Über rein-mütterliche Charaktere an Bastardlarven von Echiniden. Arch Entwicklungsmech Organ. 1898;7:65–102. [Google Scholar]
- Gilbert SF. The embryological origins of the gene theory. J Hist Biol. 1978;11:307–51. doi: 10.1007/BF00389303. [DOI] [PubMed] [Google Scholar]
- Gilbert SF. In friendly disagreement: Wilson, Morgan, and the embryological origins of the gene theory. Am Zool. 1987;27:787–806. [Google Scholar]
- Gurdon JB, Elsdale TR, Fischberg M. Sexually mature individuals of Xenopus laevis from the transplantation of single somatic nuclei. Nature. 1958;182:64–65. doi: 10.1038/182064a0. [DOI] [PubMed] [Google Scholar]
- Haeckel EHPA. Generelle Morphologie der Organismen: allgemeine Grundzüge der organischen Formen-Wissenschaft, mechanisch begründet durch die von Charles Darwin reformirte Descendenz-Theorie. G. Reimer; Berlin. 1866. [Google Scholar]
- Harris H. The Birth of the Cell. Yale University Press; New Haven: 1999. [Google Scholar]
- Harvey E. Development of half of the parts of sea urchin eggs separated by centrifugal force. Biol Bull. 1933;64:125–148. [Google Scholar]
- Harvey E. A comparison of the development of nucleate and non nucelated eggs of Arbacia punctulata. Biol Bull. 1940;79:166–187. [Google Scholar]
- Harvey E. Cleavage in centrifuged eggs and in partheongenetic merogones. Ann NY Acad Sci. 1951;51:1336–1348. doi: 10.1111/j.1749-6632.1951.tb30068.x. [DOI] [PubMed] [Google Scholar]
- Harvey EB. The American Arbacia and Other Sea Urchins. Princeton University Press; Princeton, NJ: 1956. [Google Scholar]
- Hertwig O. 1875 Beiträge zur Kenntniss der Bildung, Befruchtung und Theilung des thierischen Eies. W. Engelmann; Leipzig: [Google Scholar]
- Hertwig O, Hertwig R. 1887 Über den Befruchtungs - und Teilungsvorgang des tierischen Eies Gustav. Fischer; Jena: [Google Scholar]
- Hörstadius S. Studien über heterosperme Seeigelmerogonee nebst Bemerkungen über einige Keimblattchimären. Mem Muse Royal d’ nat de Belgique Ser 2 Fac. 1936;3:800–88. [Google Scholar]
- Lederman M, Burian R. The right organism for the job - introduction. Journal of the History of Biology. 1993;26:235–237. [Google Scholar]
- Morgan TH. Studies on the partial larvae of Sphaerechinus. Archiv für Entwicklungsmechanik der Organismen. 1895;2:81–125. [Google Scholar]
- Moritz KB. Theodor Boveri (1862–1915), Pionier der modernen Zell- und Entwicklungsbiologie. Gustav Fischer; Stuttgart: 1993. [Google Scholar]
- Oliveri P, Carrick DM, Davidson EH. A regulatory gene network that directs micromere specification in the sea urchin embryo. Dev Biol. 2002;246:209–228. doi: 10.1006/dbio.2002.0627. [DOI] [PubMed] [Google Scholar]
- Rauber A. Personaltheil und Germinaltheil des Individuum. Zoologischer Anzeiger. 1886;9:166–171. [Google Scholar]
- Revilla-i-Domingo R, Oliveri P, Davidson EH. A missing link in the sea urchin embryo gene regulatory network: hesC and the double-negative specification of micromeres. Proc Natl Acad Sci USA. 2007;104:12383–12388. doi: 10.1073/pnas.0705324104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rheinberger HJ. Toward a history of epistemic things: synthesizing proteins in the test tube. Stanford University Press; Stanford, Calif: 1997. [Google Scholar]
- Roux W. 1881 Der Kampf der Theile im Organismus: ein Beitrag zur Vervollständigung der mechanischen Zweckmässigkeitslehre Wilhelm. Engelmann; Leipzig: [Google Scholar]
- Seeliger O. Gibt es geschlechtlich erzeugte Organismen ohne mütterliche Eigenschaften? Archiv für Entwicklungsmechanik der Organismen. 1894;1:203–224. [Google Scholar]
- Seeliger O. Bemerkungen über Bastardlarven der Seeigel. Archiv für Entwicklungsmechanik der Organismen. 1896;3:478–526. [Google Scholar]
- Schroeder TE. Development of a “primitive” sea urchin (Eucidaris tribuloides): Irregularities in the hyaline layer, micromeres, and primary mesenchyme. Biol Bull. 1981;161:141–151. [Google Scholar]
- Tennent DH. The early influence of the spermatozoan upon the characters of echinoid larvae. Carnegie Inst Wash Publ. 1914;18:129–138. [Google Scholar]
- Virchow RLK. Die cellularpathologie in ihrer begründung auf physiologische und pathologische gewebelehre. A. Hirschwald; Berlin: 1858. [PubMed] [Google Scholar]
- von Ubisch L. Über Seeigelmerogonee. Pub del Staz Zool di Nap. 1954;25:247–345. [Google Scholar]
- Wilson EB. 1896 The Cell in Development and Inheritance. Macmillan; New York: [Google Scholar]
- Wilson EB. The Cell in Development and Inheritance. Macmillan; New York: 1900. [Google Scholar]
- Wilson EB. The Cell in Development and Heredity. Macmillan; New York: 1925. [Google Scholar]
- Whiteley AH, Baltzer F. Development, respiratory rate and content of desoxribonucleic acid in the hybrid Paracentrotus ♀ x Arbacia ♂. Publ Staz Zool Napoli. 1958;30:402–457. [Google Scholar]
- Wray GA, McClay DR. The origin of spicule-forming cells in a ‘primitive’ sea urchin (Eucidaris tribuloides) which appears to lack primary mesnchyme cells. Development. 1988;103:305–315. doi: 10.1242/dev.103.2.305. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.