

Abstract
A field test was conducted on the accuracy of an eight-microphone acoustic location system designed to triangulate the position of duetting rufous-and-white wrens (Thryothorus rufalbus) in Costa Rica’s humid evergreen forest. Eight microphones were set up in the breeding territories of twenty pairs of wrens, with an average inter-microphone distance of 75.2±2.6 m. The array of microphones was used to record antiphonal duets broadcast through stereo loudspeakers. The positions of the loudspeakers were then estimated by evaluating the delay with which the eight microphones recorded the broadcast sounds. Position estimates were compared to coordinates surveyed with a global-positioning system (GPS). The acoustic location system estimated the position of loudspeakers with an error of 2.82±0.26 m and calculated the distance between the “male” and “female” loudspeakers with an error of 2.12±0.42 m. Given the large range of distances between duetting birds, this relatively low level of error demonstrates that the acoustic location system is a useful tool for studying avian duets. Location error was influenced partly by the difficulties inherent in collecting high accuracy GPS coordinates of microphone positions underneath a lush tropical canopy, and partly by the complicating influence of irregular topography and thick vegetation on sound transmission.
I. INTRODUCTION
An acoustic location system consists of an array of microphones that facilitates spatial monitoring of animals based on the sounds they produce. Acoustic location systems offer special research advantages because animals’ natural movement patterns are not inhibited by the passive monitoring process and animals can be located even in environments characterized by visual obstructions (McGregor et al. 1997). Furthermore, the signals exchanged between animals are recorded as part of the acoustic location process, facilitating detailed studies of communication processes between multiple individuals simultaneously (McGregor et al. 1997, Burt & Vehrencamp 2005). Acoustic location systems have been used extensively to monitor the behaviour of many marine organisms, with particular focus on large mammals (e.g. Stafford et al. 1998, Mellinger & Clark 2003, Clark & Clapham 2004). In contrast, research employing terrestrial acoustic location systems is uncommon (but see Spiesberger 1999, Hayes et al. 2000, Mohl et al. 2001, Wahlberg et al. 2003, Bower & Clark 2005, Burt & Vehrencamp 2005). Here we test the accuracy of a passive acoustic location system for monitoring the position of duetting songbirds in neotropical forest.
Duets occur when two animals, usually breeding partners, coordinate their songs in overlapping or exactly-alternating bouts of vocalizations. Duets are produced in many taxa, including insects (Bailey 2003), frogs (Emerson & Boyd 1999), mammals (e.g. Geissmann 2002), and birds (Hall 2004). Duetting occurs in more than 220 species of birds and ranks among the most highly coordinated behaviours known in the animal kingdom (Thorpe 1963, Hall 2004). However, much controversy and little consensus surrounds the ecological and evolutionary significance of duetting, as well as the mechanics of vocal coordination by duetting partners. This is, in part, because many duetting birds live in densely vegetated tropical forest where traditional tools for following and observing animals, such as visual tracking or radiotelemetry, are difficult. Nevertheless, spatial analyses of duetting may allow for an advanced understanding of the significance of this intriguing behaviour. We were interested in studying duetting behaviour in neotropical rufous-and-white wrens (Thryothorus rufalbus) from a spatial perspective. To that end, we developed an eight-microphone acoustic location system for spatial monitoring of the duetting behaviour of birds in the humid forests of northwest Costa Rica. In this study we present a test of this system by quantifying the accuracy with which microphone array recordings allowed us to triangulate the position of loudspeakers broadcasting simulated rufous-and-white wren duets.
II. METHODS
A. Study species and study site
Rufous-and-white wrens live in forested and semi-forested habitat throughout much of Central America and northwestern South America. Males and females coordinate their songs to produce both antiphonal and polyphonal duets. The voices of the two sexes are similar, although male songs have significantly lower frequency characteristics and are significantly longer than female songs (Mennill & Vehrencamp 2005). Rufous-and-white wrens are largely ground-dwelling songbirds (Ahumada 2001) and typically remain in very low vegetation, occasionally ascending to higher perches to sing.
At Santa Rosa National Park in northwest Costa Rica (10°40′N, 85°30′W), our study population of rufous-and-white wrens inhabits the fragments of evergreen moist forest that remain within the mosaic of regenerating forests that constitute the Guanacaste Conservation Area. The study site is characterized by thick vegetation, including a rich vertical structure and a lush canopy dominated by guapinol (Hymenea courbaril), milk (Brosiumu utile), and chicle (Manilkara chicle) trees. The study site also has diverse topography – the wrens’ territories follow the sloped stream-sides and small escarpments that are associated with the park’s evergreen moist forest. These biotic and abiotic features of rufous-and-white wren territories provide a challenging model system in which to test an acoustic location system; rich understory vegetation complicates sound transmission, thick forest canopy makes high-accuracy survey coordinates difficult to obtain, and topographic diversity complicates both sound transmission and satellite reception. Therefore, if an acoustic location system can be used to accurately monitor the position of duetting birds in this habitat, it will be useful for monitoring animals in many other habitats as well.
B. Field methods
Our acoustic location system consisted of an array of eight omni-directional microphones which were connected by 1200 m of microphone cable to a centrally-located computer. Microphone input was digitized using a multi-channel data acquisition card (model: National Instruments DAQ-6260). The microphones were housed in rain guards made of PVC tubing and speaker cloth, mounted on top of 3 m wooden poles via 30 cm shelf brackets. Stakes were elevated off the ground and attached to trees with bungee cords (Fig. 1). We attempted to space microphones evenly throughout each rufous-and-white wren territory, mounting microphones opportunistically where vegetation allowed. Within each array, the average between-microphone distance was 75.2 ± 2.6 m (calculated pairwise for all microphones in each array), the maximum distance was 137.6 ± 5.9 m and the minimum distance was 31.5 ± 1.4 m (mean ± SE; n=20 array configurations).
Figure 1.

Photograph of one of the eight microphones used to record playback of rufous-and-white wren duets. Microphones were placed on 3.0 m wooden poles and attached to trees within wren territories. The average between-microphone distance within each of the 20 array configurations was 75.2 ± 5.7 m.
Between April 18 and July 8, 2003 and 2004 we used the eight-channel microphone array to record duets played through stereo loudspeakers within the breeding territories of 20 different pairs of wrens. Recording sessions were conducted between 0630 and 0845 h. The left and right loudspeakers (model: Sony SRS-A37) were separated by a distance of 10 to 20 m (average distance between loudspeakers: 16.3 ± 0.5 m), a distance within the observed range of distances between duetting rufous-and-white wrens. The left loudspeaker broadcast one bird’s duet contribution and the right loudspeaker broadcast the partner’s duet contribution. The loudspeakers were oriented upwards and mounted on poles at a height of 1.0 m. Stimuli were broadcast from a portable CD player (model: Sony D-SJ301). The volume of broadcast songs was held constant across all trials, at a natural sound pressure level of 80 dB at 1 m (measured at a horizontal distance from the upwards-oriented loudspeaker using a Realistic 33-2050 sound level meter on slow setting).
The playback stimuli were duets recorded from the 20 resident pairs. We recorded stimuli as monaural WAV files (16 bit sampling, 22050 Hz sample rate) with a directional microphone (model: Sennheiser MKH-70) and a solid state digital recorder (model: Marantz PMD-670) during spontaneous bouts of early-morning singing. The male and female duet components were isolated from each another using the “frequency cursor filter” function of Syrinx-PC (J. Burt, Seattle, WA). The amplitudes of both the male and female songs were then subtly adjusted to the same maximum amplitude using the “amplitude normalize” feature of CoolEdit 2000 (Syntrillium, Phoenix, AZ). Male and female songs were then pasted into the left and right channels of 16-bit stereo WAV files (Fig. 2). We alternated whether the male or female song was broadcast through the left or right channel from one playback session to the next. Rufous-and-white wren duets occur in several different varieties: the male may begin singing and the female may join in part way through the male song or immediately after it; the sex roles may be reversed; or both birds may begin singing at the same time. Male-initiated duets are the most common variety (Mennill & Vehrencamp 2005) and for this reason we used male-initiated duets in this test of location accuracy. Within each array, we recorded the broadcast of six duets, as well as six male solos and six female solos for comparison. The solo stimuli were identical to the male and female contributions to the duet.
Figure 2.

Sound spectrograms of rufous-and-white wren duet stimuli broadcast within the acoustic location system. Spectrograms depict stereo sound files with the left channel on top and the right channel below. Examples of three different pairs’ duets are shown.
Traditional survey methods are difficult given the thick vegetation at our study site. We surveyed microphone and loudspeaker positions using an Ashtech ProMark II Global Positioning System (GPS; Thales Navigation, Santa Clara, CA). This system consists of twin receiver units with external antennae which collect data in tandem at two positions, allowing for differential correction based on the data collected at the two sites simultaneously. We mounted the GPS antennae on tripods at a height of 1.70 m. In an attempt to overcome the error introduced by GPS sampling underneath a thick forest canopy, we sampled points extensively and repeatedly, sampling each site two to six times between 2003 and 2005. Combined over three years of GPS sampling, we sampled the position of each microphone for 85.9 ± 1.3 minutes (n=160 microphone positions) and the position of each loudspeaker for 74.7 ± 1.6 minutes (n=40 loudspeaker positions). We used Ashtech Solutions Software (Thales Navigation, Santa Clara, CA) to calculate an average UTM coordinate from each loudspeaker and microphone position. We report error around these coordinates as the standard error given as output by the Ashtech Solutions Software following post-processing adjustment.
C. Array analysis
In the laboratory, we used Syrinx-PC to browse eight-channel spectrograms of field recordings. We used the time and frequency cursors of Syrinx-PC to draw a selection box around non-overlapping sections of male and female duet contributions. As is typical of rufous-and-white wren duets (Mennill & Vehrencamp 2005), our broadcast stimuli involved duets where the terminal section of the male song was overlapped by the beginning section of the female song. Therefore, we selected only the introductory portion of male duet contributions and the terminal portion of the female duet contributions. For our analyses of the solo songs, in contrast, we were able to select the entire male or female song.
We used software written in MatLab (Mathworks Inc, Natick, MA) to estimate the location of the loudspeakers that broadcast male and female songs. The localization process consists of two components: computing cross-correlation functions and searching for the best location estimate. The correlation component begins by copying the sound data from the selected channel and applying an FIR filter to the data to restrict the signal energy to the frequency limits of the selection box. The digital samples that fall outside of the temporal bounds of the selected region are replaced with zeroes. This edited signal is cross-correlated against the remaining seven channels. These correlations are resistant to the presence of other sounds within the data segment because the extent of the selection is used to mask out all the other signals in the selected channel. Twenty one additional correlation functions are computed between all pairs of microphones that do not include the selected channel. For these correlations, it is advantageous to utilize the channels with the strongest signals to provide the masked template that is correlated against the remaining channels. The amplitudes of peaks in the initial seven correlations are used to order the seven channels in terms of estimated signal strength. The remaining correlations are computed as follows: the channel with the highest correlation peak is correlated with the remaining six channels, the channel with the second highest correlation peak is correlated with the remaining five channels, etc. The locations of the correlation peaks are used to apply masking, to help reduce the impact of other signals that occur in the data segment. The software can deal with multiple correlation peaks that complicate the masking process, but this complication was not an issue in this study.
Once the correlation functions are computed, the estimated location of the selected sound is obtained by a stochastic search which involves the GPS coordinates of the 8 microphones. For each candidate source location, the predicted time delays are computed using the Euclidean distances between the source location and the microphone locations, combined with the speed of sound. The values of the correlation functions associated with the predicted delays are extracted and these 28 values are summed. The location with the maximal sum of these correlation values provides the estimated location. This optimization criterion is algebraically identical to finding the point in space that produces the highest energy output from a delay-and-sum beamforming system (Johnson and Dudgeon 1993). Our array analysis process assumes that the speakers lie in a plane (two dimensional localization) and deviations from this assumption contribute to location errors. For all analyses we fixed the speed of sound at 348 m/sec (corresponding to air temperature of 25 °C and 90 % relative humidity, as is typical at our study site at the time of recording; see below). We evaluate how variation in air temperature and humidity – and therefore speed of sound – influenced position estimates by plotting temperature and humidity against location accuracy.
D. Location accuracy
To test the accuracy of the acoustic location system, we compared the loudspeakers’ GPS coordinates to the positions estimated by the array analysis software. We report the difference as “location accuracy”; the Euclidean distance between the GPS coordinates of the loudspeaker and the array-estimated coordinates of the sound source (in meters).
To evaluate environmental variables that may have influenced the accuracy of our array-based location estimates, we compared location accuracy measurements across the 20 arrays to four features of the acoustic environment: (1) air temperature at the time of the recording, (2) relative humidity of the air at the time of the recording, (3) recording date (a rough measure of leaf phenology), and (4) variance in the height of the eight microphones within each array (a rough measure of topographic variation within each recording area). We measured temperature and humidity with a data logger (model: Hobo Pro, Onset Technology, Pocasset, MA), which was hung at a height of 1.0 m near the center of each array and sampled the air condition every five minutes. We also compare our error measurements to two features of the microphone arrays themselves: (5) the density of microphones (measured as the average distance between all pairs of microphones in each array), and (6) the distance between the loudspeaker and the nearest microphone (a feature which has been shown to influence recording accuracy in a study of temperate microphone arrays; McGregor et al. 1997).
E. Statistical methods
All analyses were conducted in JMP 5.0 (SAS Institute, Cary, NC). We compare the location error of female versus male songs broadcast as solos versus duets with two-factor ANOVA. We test for associations between accuracy and environmental variables with Pearson correlations. All tests are two-tailed. All values are reported as mean ± SE.
III. RESULTS
A. GPS accuracy
Using a global position system (GPS) to survey the positions of eight microphones within each of the 20 microphone arrays, we achieved a latitudinal accuracy of 1.26 ± 0.05 m and a longitudinal accuracy of 1.36 ± 0.05 m, for an average accuracy of 1.31 ± 0.05 m (n=160 microphone positions). For loudspeaker positions, we achieved a latitudinal accuracy of 2.49 ± 0.09 m and a longitudinal accuracy of 2.68 ± 0.09 m, for an average accuracy of 2.59 ± 0.09 (n=40 microphone positions; lower accuracy in loudspeaker coordinates arose from shorter sampling periods). The accuracy of all GPS coordinates was constrained by the thick canopy, as has been found in other studies involving GPS-based research in tropical forest environments (e.g. Phillips et al. 1998, Dominy & Duncan 2001).
B. Location accuracy
Using the eight-channel acoustic location system, we triangulated the position of loudspeakers broadcasting rufous-and-white wren song with an average location error of 2.82 ± 0.26 m (n=40 loudspeakers, average location error calculated across all stimuli broadcast from each loudspeaker; Fig. 3). It is noteworthy that this value is fractionally larger than the estimated GPS location error for the speaker locations, suggesting that the combined effects of all errors in the acoustic location system are smaller than the GPS survey error for the speakers. The location error was similar for locations based on the broadcast of male solos, male duet contributions, female solos, and female duet contributions (Fig. 4; two-factor ANOVA: whole model F3,76=0.4, p=0.73) despite the significantly lower frequency characteristics of male songs (see Mennill & Vehrencamp 2005). The six songs broadcast from each loudspeaker generated tightly clustered position estimates, demonstrating that our position estimates were repeatable.
Figure 3.
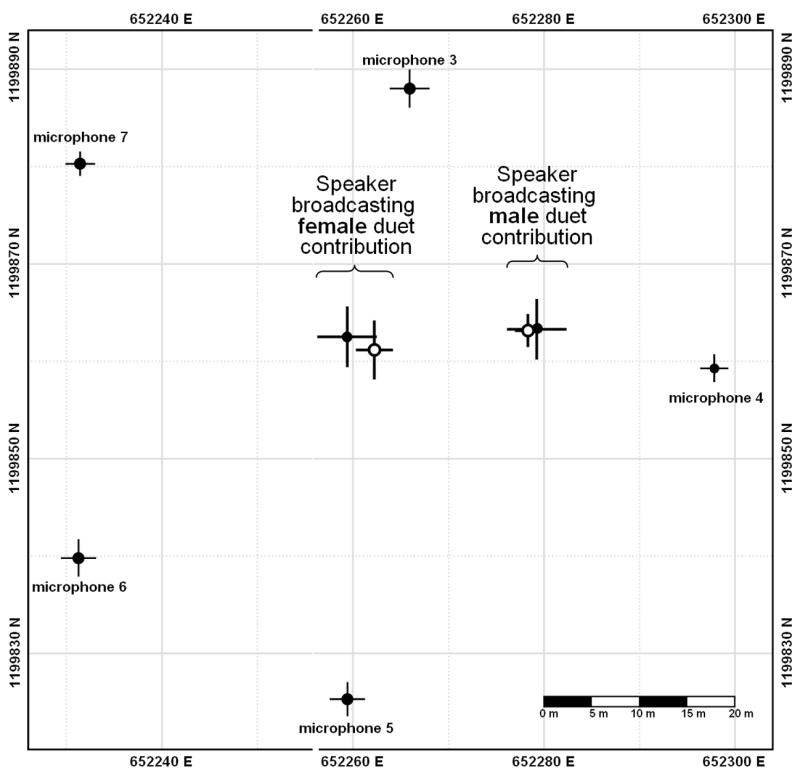
Map of a portion of a microphone array, showing five microphones and two loudspeaker positions. Stereo loudspeakers were set up near the center of the area enclosed by the microphones and stereo duet stimuli were broadcast through the left and right channels. The solid circles show positions of loudspeakers and microphones measured by GPS. The open circles show the position of the loudspeakers estimated using the acoustic location system. Horizontal and vertical lines show standard error.
Figure 4.
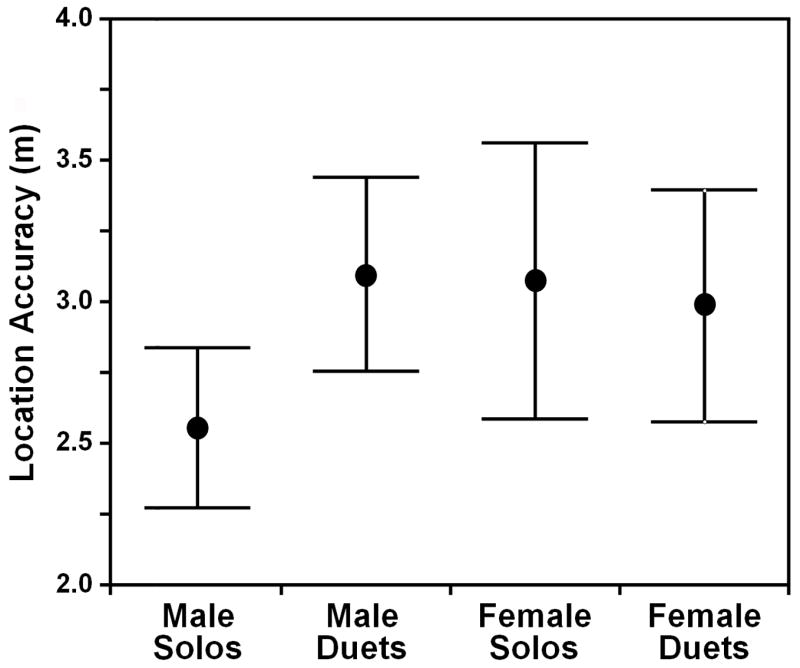
Location accuracy for n=40 loudspeakers which broadcast rufous-and-white wren vocalizations and were recorded within 20 microphone arrays. Location accuracy was similar when loudspeakers broadcast male and female vocalizations, whether as solos or duets. Points show means and vertical bars show standard error.
Array recordings accurately estimated the distance between a pair of loudspeakers simulating the male and female duet contributions of a pair of rufous-and-white wrens. The discrepancy in the estimated distance between the male and female loudspeakers, as measured by GPS versus the acoustic location system, was 2.12 ± 0.42 m (n=20).
C. Factors influencing location accuracy
None of the variables we measured showed a relationship with location accuracy (Fig. 5). Location accuracy did not vary significantly across a temperature range of 22.1 to 25.2 degrees Celsius (r=0.07, p=0.67, n=40), a range typical of the early to mid morning at our study site. Location accuracy did not vary significantly across a humidity range of 58.3 to 99.2 % (r=0.12, p=0.45, n=40). The acoustic location system was remarkably robust and functioned effectively even at the high humidity levels within this tropical forest. Location accuracy did not change with the time of year (r=0.02, p=0.92, n=40), indicating that leaf phenology had little influence on the accuracy of the acoustic location system. This is not surprising, as the leaf structure of the study site remains similar throughout most of the year, varying primarily with bursts of leaf production in the canopy, as is the case for many tropical forests (van Schaik et al. 1993). Despite the variation in topography at the study site, location accuracy did not show a significant relationship with topographic variation, measured as the variance in the heights of the microphones (r=0.18, p=0.26, n=40). Similarly, location accuracy did not vary significantly with microphone density (r=0.04, p=0.81, n=40) or the distance between the loudspeaker and the center of the array (r=0.09, p=0.59, n=40).
Figure 5.
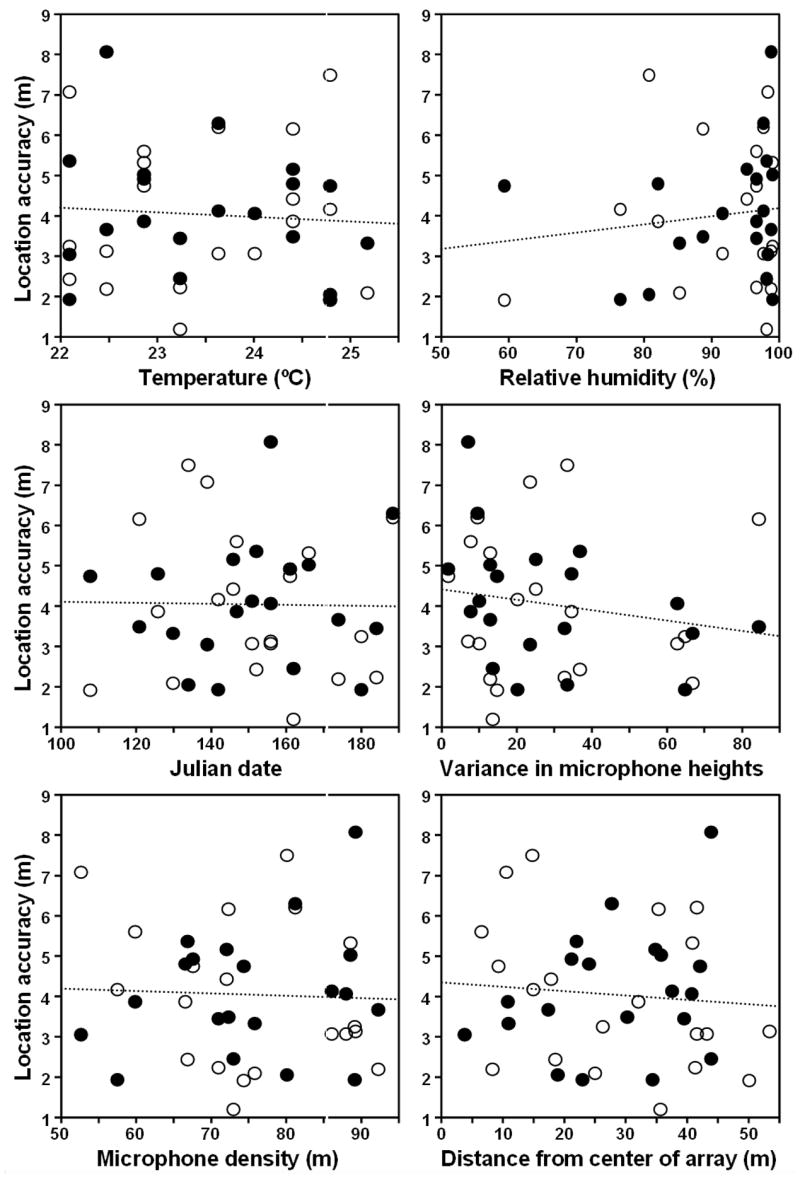
Location accuracy did not show a strong relationship with four features of the recording environment or two features of array configuration: air temperature, the relative humidity of the air, Julian date (a rough measure of leaf phenology), variance in microphone height (a rough measure of topographic variation), the density of microphones within the array (measured as the average distance between microphones), or the distance between the loudspeaker and the center of the array. Open circles show n=20 loudspeakers which broadcast female duet contributions and closed circles show n=20 loudspeakers which broadcast male duet contributions.
IV. DISCUSSION
The acoustic location system provided accurate estimates of the position of loudspeakers broadcasting rufous-and-white wren duets in neotropical humid forest. Triangulating the position of loudspeakers based on the delay in time with which eight microphones recorded playback stimuli, we achieved an average location error of 2.82 meters. We estimated the distance between the loudspeakers broadcasting male and female duet components to within 2.12 meters. Therefore, our test of acoustic location system accuracy establishes microphone array technology as a useful tool for spatial analysis of the vocal behaviour of animals in tropical forests, even within areas characterized by dense vegetation, thick canopy cover, and irregular topography.
When participating in vocal duets, the distance between male and female rufous-and-white wrens varies tremendously (D. J. Mennill unpublished data). Because the error of our acoustic location system is small, the accuracy of this acoustic location system will be revealing for monitoring duetting behaviour in these Neotropical wrens. Recording natural duets with the acoustic location system will allow us to test hypotheses for the function of avian duets by evaluating how close together birds are when they duet, whether birds approach one another during a bout of duetting, and where birds sing duets relative to territory boundaries and other habitat features. Whereas single channel recordings are predicated on the idea of monitoring one individual at a time, a multi-microphone system may be used to monitor multiple individuals simultaneously, and therefore to understand behaviours such as duetting. Moreover, because multi-microphone recordings permit spatial analysis of animal vocal behaviour, acoustic location systems can be used to monitor the movement and spacing of animals in habitats where traditional techniques are impractical.
Error in our ability to triangulate exactly the position of the loudspeakers arose from at least two sources. First, the accuracy of our acoustic location system is dependent on the accuracy with which we surveyed microphone positions. Our GPS coordinates of microphone positions were accurate to only 1.31 meters despite extensive surveying across multiple years. Tropical forest ranks among the most challenging environments for collecting high-accuracy GPS coordinates because of the presence of dense canopy vegetation (Blake et al. 2001, Phillips et al. 1998, Dominy & Duncan 2001). When surveying areas near our study site that lacked a dense canopy cover, we found that our GPS was capable of achieving sub-meter accuracy, supporting the idea that it is the canopy layer within the study site that made high-accuracy GPS coordinates difficult to obtain, not satellite geometry or any other feature of the tropics per se. When future research questions require more precise GPS coordinates for points within canopied tropical forest, raising GPS antennae high into the canopy on telescoping poles may facilitate sub-meter level accuracy.
The second source of error in our location accuracy probably stemmed from the influence of dense vegetation and diverse topography on sound transmission within the study site. Our acoustic location system triangulates the position of animals in two dimensions, effectively treating the recorded area as a flat plane. In reality, the study site has many hills and valleys which are likely to influence patterns of sound reverberation for songs transmitted in this environment. We found no relationship between the variance in the height of the eight microphones in each array (a rough measure of the topographic variation within each array) and location accuracy. However, we do not interpret this as evidence that topography had no influence on the acoustic location system, but rather that topography had equivalent effects across all twenty array configurations. There are no analytical software tools yet developed to account for both three-dimensional transmission of sound together with a variable habitat surface. This is an important area for future enhancement in acoustic location system accuracy.
Both McGregor et al. (1997) and Bower and Clark (2005) evaluated the accuracy of a four-microphone array for triangulating the position of songbirds. By recording a variety of sounds in an open meadow and an open woodland in Europe, McGregor et al. (1997) evaluated the accuracy of an acoustic location system in two temperate environments. They report an average location error of 3.68 ± 0.29 m in open meadow and 6.76 ± 0.56 in open woodland, although accuracy improved substantially when the broadcast sound was closer to the center of their four-element arrays (McGregor et al. 1997). Bower and Clark (2005) recorded songbirds in a re-grown farm field in North America with a similar array of four microphones. They report an average location error of 0.82 ± 0.29 m for birds recorded near the center of their four-element array and 2.13 ± 1.30 m for birds recorded just outside of their array. Both McGregor et al. (1997) and Bower and Clark (2005) report a relationship between location error and the distance between the loudspeaker and the center of the array. We did not find this effect in our analyses, however all of our stimuli were recorded from positions near the center of the 20 arrays, whereas these other analyses included sounds recorded outside the area bounded by their four microphones. We expect that location accuracy would decline if duets were recorded from outside of the area encompassed by the array of microphones. McGregor et al. (1997) and Bower and Clark (2005) recorded only on windless days in environments which induced little reverberation, whereas we conducted recordings through a variety of inclement weather, including substantial winds and persistent dripping water, and in an environment which was likely to induce much reverberation. Our analyses provide a realistic test of the abilities of an acoustic location system, and demonstrate that an acoustic location system can be used in suboptimal recording conditions.
Our understanding of the behaviour and ecology of tropical animals remains poorly developed, despite the tremendous diversity of animal life in the tropics and the urgency for tropical research in light of the conservation threats to tropical forests (Kroodsma et al. 1996). Acoustic location technology is a highly valuable tool for enhancing our understanding of the ecology and evolution of tropical animals. Microphone array recordings may be useful for studying extremely secretive animals or animals that are difficult to observe because of dense vegetation, large home ranges, or nocturnal activity. Microphone arrays also offer a valuable tool for studying network-level communication, and have been used to study neighbourhoods of song sparrows (Bower 2000) and banded wrens (Burt & Vehrencamp 2005). We believe that acoustic location technology may be used to study many other systems and provide ecologists and ethologists with a valuable and versatile tool for tropical research.
Acknowledgments
We thank a dedicated field team who assisted with the difficult task of operating microphone arrays and collecting GPS data: C. Calaustro, V. Connolly, S. Doucet, A. Lindo, D. Moseley, R. Richter, and S. Topp. We thank R. Blanco and the staff of the Guanacaste Conservation Area for logistical support in the field. We thank everyone at Cornell University’s Bioacoustics Research Program for logistical support during the design and analysis stages, including K. Dunsmore, A. Gururajan, and K. Cortopassi for implementing and testing the localization algorithm. We thank J. Cuthbert, S. Doucet, L. Reed, K. Swiston, S. Topp, and an anonymous reviewer for comments on the manuscript. Financial support for this project was provided by grants from the National Geographic Society, the Natural Science and Engineering Research Council (NSERC) of Canada, the American Museum of Natural History’s Frank M. Chapman Memorial fund, the Birmingham Audubon Society’s Walter F. Coxe Research Fund, and the Explorers Club’s Exploration Fund to D.J.M., and by a grant from the National Institutes of Health (R01-MH60461) to S.L.V.
Contributor Information
Daniel J. Mennill, Department of Biological Sciences, University of Windsor, 401 Sunset Ave, Windsor, Ontario, Canada N9B3P4 (dmennill@uwindsor.ca)
John M. Burt, Psychology Department, University of Washington, Seattle, Washington, 98195 (quill@u.washington.edu)
Kurt M. Fristrup, Natural Sounds Program Center, National Park Service, 1201 Oakridge Drive, Suite 100, Fort Collins, Colorado, 80525 (kurt_fristrup@nps.gov)
Sandra L. Vehrencamp, Lab of Ornithology, Cornell University, 159 Sapsucker Woods Road, Ithaca, New York, 14850 (slv8@cornell.edu)
References
- Ahumada JA. Comparison of the reproductive biology of two neotropical wrens in an unpredictable environemnt in northeastern Colombia. Auk. 2001;118:191–210. [Google Scholar]
- Bailey WJ. Insect duets: underlying mechanisms and their evolution. Physiol Entomol. 2003;28:157–174. [Google Scholar]
- Blake S, Douglas-Hamilton I, Karesh WB. GPS telemetry for forest elephants in Central Africa: results of a preliminary study. Afr J Ecol. 2001;39:178–186. [Google Scholar]
- Bower JL. PhD thesis. Cornell Unviersity; New York: 2000. Acoustic interaction sduring naturally occurring territorial conflict in a song sparrow neighbourhood. [Google Scholar]
- Bower JL, Clark CW. A field test of the accuracy of a passive acoustic location system. Bioacoustics. 2005;15:1–14. [Google Scholar]
- Burt JM, Vehrencamp SL. Dawn chorus as an interactive communication network. In: McGregor PK, editor. Animal Communication Networks. Cambridge University Press; 2005. pp. 321–343. [Google Scholar]
- Clark CW, Clapham PJ. Acoustic monitoring on a humpback whale (Megaptera novaeangliae) feeding ground shows continual singing into late Spring. Proc Roy Soc Lond B. 2004;271:1051–1057. doi: 10.1098/rspb.2004.2699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dominy NJ, Duncan B. GPS and GIS methods in an African rain forest: Applications to tropical ecology and conservation. Conserv Ecol. 2001;5:6. [Google Scholar]
- Emerson SB, Boyd SK. Mating vocalizations of female frogs: Control and evolutionary mechanisms. Brain Behav Evol. 1999;53:187–197. doi: 10.1159/000006594. [DOI] [PubMed] [Google Scholar]
- Geissmann T. Duet-splitting and the evolution of gibbon songs. Biol Rev. 2002;77:57–76. doi: 10.1017/s1464793101005826. [DOI] [PubMed] [Google Scholar]
- Hall ML. A review of hypotheses for the functions of avian duetting. Behav Ecol Sociobiol. 2004;55:415–430. [Google Scholar]
- Hayes SA, Mellinger DK, Croll DA, Costa DP, Borsani JF. An inexpensive passive acoustic system for recording and localizing wild animal sounds. J Acoust Soc Am. 2000;107:3552–3555. doi: 10.1121/1.429424. [DOI] [PubMed] [Google Scholar]
- Johnson DJ, Dudgeon DE. Array Signal Processing: Concepts and Techniques. Prentice-Hall; Englewood Cliffs, NJ: 1993. [Google Scholar]
- Kroodsma DE, Vielliard JM, Stiles FG. Study of bird sounds in the neotropics: Urgency and opportunity. In: Kroodsma DE, Miller EH, editors. Ecology and Evolution of Acoustic Communication in Birds. Cornell University Press; Ithaca: 1996. pp. 269–281. [Google Scholar]
- McGregor PK, Dabelsteen T, Clark CW, Bower JL, Tavares JP, Holland J. Accuracy of a passive acoustic location system: Empirical studies in terrestrial habitats. Ethol Ecol Evol. 1997;9:269–286. [Google Scholar]
- Mellinger DK, Clark CW. Blue whale (Balaenoptera musculus) sounds from the North Atlantic. J Acoust Soc Am. 2003;114:1108–1119. doi: 10.1121/1.1593066. [DOI] [PubMed] [Google Scholar]
- Mennill DJ, Vehrencamp SL. Sex differences in the singing and duetting behavior of neotropical Rufous-and-white Wrens (Thryothorus rufalbus) Auk. 2005;122:175–186. [Google Scholar]
- Mohl B, Wahlberg M, Heerfordt A. A large-aperture array of non-linked receivers for acoustic positioning of biological sound sources. J Acoust Soc Am. 2001;109:434–437. doi: 10.1121/1.1323462. [DOI] [PubMed] [Google Scholar]
- Phillips KA, Elvey CR, Abercrombie CL. Applying GPS to the study of primate ecology: a useful tool? Am J Primatol. 1998;46:167–172. doi: 10.1002/(SICI)1098-2345(1998)46:2<167::AID-AJP6>3.0.CO;2-U. [DOI] [PubMed] [Google Scholar]
- Spiesberger JL. Locating animals from their sounds and tomography of the atmosphere: Experimental demonstration. J Acoust Soc Am. 1999;106:837–846. doi: 10.1121/1.427100. [DOI] [PubMed] [Google Scholar]
- Stafford KM, Fox CG, Clark DS. Long-range acoustic detection and localization of blue whale calls in the northeastern Pacific Ocean. J Acoust Soc Am. 1998;104:3616–3625. doi: 10.1121/1.423944. [DOI] [PubMed] [Google Scholar]
- Thorpe WH. Antiphonal singing in birds as evidence for avian auditory reaction time. Nature. 1963;197:774–776. [Google Scholar]
- van Schaik CP, Terborgh JW, Wright JS. The Phenology of tropical forests: Adaptive significance and consequences for primary consumers. Ann Rev Ecol Sys. 1993;24:353–377. [Google Scholar]
- Wahlberg M, Mohl B, Madsen PT. Estimating source position accuracy of a large aperture hydrophone array for bioacoustics. J Acoust Soc Am. 2001;109:397–406. [Google Scholar]
- Wahlberg M, Tougaard J, Mohl B. Localising Bitterns Botaurus stellaris with an array of non-linked microphones. Bioacoustics. 2003;13:233–246. [Google Scholar]