

Abstract
We describe a theory that brain mechanisms underlying working memory for novel information include a buffer in parahippocampal cortices. Computational modelling indicates that mechanisms for maintaining novel information in working memory could differ from mechanisms for maintaining familiar information. Electrophysiological data suggest that the buffer for novel information depends on acetylcholine. Acetylcholine activates single-cell mechanisms that underlie persistent spiking of neurons in the absence of synaptic transmission, allowing maintenance of information without prior synaptic modification. fMRI studies and lesion studies suggest that parahippocampal regions mediate working memory for novel stimuli, and effects of cholinergic blockade impair this function. These intrinsic mechanisms in parahippocampal cortices provide an important alternative to theories of working memory based on recurrent synaptic excitation.
Keywords: acetylcholine, entorhinal cortex, sustained activity, fMRI, scopolamine
Research has demonstrated the existence of multiple memory systems, including working memory, which is defined as a limited capacity system for the temporary storage and manipulation of information for cognitive tasks [1,2]. Many initial fMRI studies of working memory in humans used highly familiar stimuli such as letters and words, and focused primarily on a system which includes prefrontal cortex (PFC) and parietal cortex [1-3]. However, electrophysiological and lesion studies in animals, and recent imaging studies suggest that temporal lobe structures play a critical role in working memory [4-6]. Here we present the theory that working memory for novel information differs from working memory for familiar information, proposing that novel stimuli require additional cellular mechanisms within the entorhinal and perirhinal cortex; whereas prefrontal and parietal systems are sufficient for maintaining familiar stimuli in working memory. It is critical to note that we are not proposing a double dissociation between the systems, but instead we suggest that the prefrontal-parietal system alone is insufficient for maintaining information that has no prior representation in the brain. We hypothesize that cholinergic activation of these single neuron mechanisms results in persistent spiking activity without excitatory synaptic transmission between neurons (see Figure 1 and Box). This theory provides an alternative to many physiological models of working memory, which use recurrent excitation between neurons to maintain persistent spiking [7]. Critically, models based on recurrent excitation can only maintain information consistent with previously formed representations (i.e. familiar information).
Figure 1.

Brain mechanisms underlying working memory (WM) for novel and familiar stimuli. Previous work suggests that working memory for familiar stimuli is mediated by excitatory recurrent connections between neurons in prefrontal cortex (PFC) and parietal cortex (PC), as indicated by the arrows connecting the circles that represent neurons. We propose that these recurrent connections are insufficient for working memory for novel stimuli. In contrast, we suggest that working memory for novel stimuli is mediated by neurons in parahippocampal cortices (ParaHC). These parahippocampal neurons have intrinsic mechanisms for persistent spiking activity, involving single cell regenerative mechanisms (represented by arrows within circles) which do not require prior synaptic modification.
In this article, we evaluate experimental data[6,8-10] and computational modelling[11-13] that support the proposal that working memory for novel stimuli requires additional cellular mechanisms activated by acetylcholine in the entorhinal cortex and other parahippocampal cortices that differ substantially from the mechanisms required for working memory for familiar stimuli. In each section we first evaluate evidence for the role of this cellular mechanism in the active maintenance of novel stimuli for working memory, and, if evidence is available, we consider a role for this mechanism in encoding of long-term memories.
Lesion studies
Working memory
Lesion studies suggest an important role for parahippocampal regions in working memory for novel stimuli. While cortical regions including prefrontal cortex and parietal cortex show activity during working memory for novel stimuli[14], these regions do not appear to be sufficient to perform working memory for novel stimuli at normal levels when parahippocampal regions are lesioned, though they are sufficient to maintain normal working memory for familiar stimuli. Many studies test working memory with delayed match to sample (DMS) or delayed non-match to sample (DNMS) tasks, in which a stimulus is presented as a sample, and after a delay period, the effective maintenance of the sample information is evaluated by presenting a test stimulus and requiring subjects to indicate whether the test matches the sample. Lesions of parahippocampal regions in monkeys do not cause impairments on DMS tasks with small numbers of highly familiar stimuli[15,16], and parahippocampal lesions in humans also do not cause impairments on tasks using highly familiar stimuli such as the digit span task[17]. In contrast, parahippocampal lesions in monkeys and humans do cause impairments when large sets of trial-unique (i.e. novel) stimuli are used in DMS[18] and delayed non-match to sample (DNMS)[19] tasks, even at delays as short as 8−10 seconds. This occurs despite the fact that matching tasks are easier (show lower error rates) with novel stimuli [5], because there is less interference from previous exposure to the stimuli, and because there is a stronger difference in item familiarity between the sample and the test stimuli. Importantly, parahippocampal lesions cause impairments in working memory for conjunctions and for complex non-verbalizable stimuli even at short retention intervals [20,21] and they disrupt working memory for novel configurations of stimuli in the Brown-Peterson task[17]. Parahippocampal lesions also disrupt memory for novel visual objects (see [22] for review). These data indicate that parahippocampal cortices are important for mediating working memory for novel stimuli, even over periods of 8−10 seconds, whereas areas outside of the parahippocampal region appear to be sufficient for normal delayed matching function with small numbers of highly familiar stimuli [15,16], but are not sufficient to support normal working memory for novel stimuli after parahippocampal lesions.
Encoding of long-term memory
The brain mechanisms necessary for working memory for novel stimuli may also be important for encoding new long-term memories. Parametric studies have shown that lesions to parahippocampal structures cause stronger impairments for matching at longer delays (e.g. 1 minute, 10 minutes) than at short delays (e.g. 0.5 seconds)[18,23]. These data indicate that parahippocampal structures are important for supporting long-lasting representations. Lesions of the hippocampus alone cause anterograde amnesia, as demonstrated by impairments of long-term memory, including free recall and paired associate memory[19], and these effects appear to be stronger in cases in which portions of the entorhinal and perirhinal cortex have also been affected[19]. These data suggest that the entorhinal and perirhinal cortices interact with the hippocampus in the formation of long-term memories, involving the strengthening of synapses to form a long-lasting representation[12,24,25].
In summary, behavioural data shows that parahippocampal lesions impair both working memory and encoding of novel stimuli, consistent with the data on fMRI activity in these regions that is reviewed in the next section.
Functional magnetic resonance imaging
Working memory
Functional neuroimaging studies support the idea that working memory for novel stimuli requires the additional recruitment of parahippocampal regions. Early functional neuroimaging studies of working memory emphasized the role of prefrontal and parietal cortices, and most studies were carried out using highly familiar stimuli, including letters, words, simple objects, and spatial locations[26]. Surprisingly, these early fMRI studies of working memory did not report activity within parahippocampal regions such as perirhinal or entorhinal cortex, although these areas had previously been shown to be necessary for DNMS and DMS tasks in monkeys[18,19,23]. The non-human primate lesion studies motivated an fMRI study by Stern et al. [5] that demonstrated differential activation for novel versus familiar stimuli during performance of a ‘two-back’ working memory task, in which subjects respond if a stimulus matches the stimulus seen two stimuli previously. This study showed that working memory for a highly familiar set of complex visual images primarily activated prefrontal and parietal cortices, whereas the same task using novel (trial-unique) visual images strongly activated parahippocampal structures in addition to prefrontal and parietal cortices[5]. Activation of parahippocampal structures associated with working memory for novel stimuli has also been shown in an event-related fMRI study using novel face stimuli[27,28].
Encoding of long-term memory
Increased parahippocampal activity for novel stimuli may also be important for encoding of long-term memories. A recent fMRI study by Schon et al. showed changes in blood flow in the entorhinal cortex and other parahippocampal cortices during the delay period in a DMS task with novel stimuli[6,29]. This delay period fMRI signal in parahippocampal cortices was correlated with subsequent memory function in a long-term recognition test at the end of the experiment [6] (Figure 2), and may be closely related to the persistent spiking activity described below in the section on Cellular Physiology[10,30]. Thus, persistent activity may facilitate the encoding of novel stimuli in humans. When subjects were injected with scopolamine, a muscarinic cholinergic antagonist, fMRI activation in parahippocampal regions which was correlated with subsequent memory was reduced[6], supporting the hypothesis that acetylcholine is necessary for the persistent activity that enhances encoding.
Figure 2.

Effect of scopolamine on delay period activity [6,29] in a delayed match to sample task, which includes Sample, Delay and Test periods. (a) Proposed pattern of delay period spiking activity which could underlie changes in blood flow detected by fMRI during performance of the task. (b1) Histogram bars show high levels of fMRI activity in parahippocampal regions during the sample, delay and test periods which is correlated with performance on the post-scan recognition test. (b2) Blockade of acetylcholine (ACh) receptors with scopolamine greatly reduces the activity in perirhinal/entorhinal regions correlated with performance on the post-scan recognition memory task. (b3) Location of significant activation in the perirhinal/entorhinal region. (c) Scanning occurred during performance of a delayed match to sample task, which includes Sample, Delay, and Test periods, with inter-trial intervals (ITI). After scanning, subjects were tested on a post-scan recognition memory task for the individual stimuli.
These data indicate that cholinergic modulation in parahippocampal cortices contributes to persistent activity for working memory and encoding of novel stimuli, consistent with results of other cholinergic manipulations reviewed in the next section.
Cholinergic manipulations
Working memory
Studies of the behavioural effects of drugs that block muscarinic acetylcholine receptors support a role for cholinergic modulation in the entorhinal cortex in the short-term retention of novel memories. Systemic injections of muscarinic acetylcholine blockers such as scopolamine impair DMS performance in monkeys at delays on the order of several seconds[31], and impair the arm choice behaviour of rats in an 8-arm radial maze when there is a delay between the individual choices[32]. In mice, selective knockouts of the m1 muscarinic receptor also cause deficits in DNMS behaviour[33].
Muscarinic blockade with drugs such as scopolamine in humans also causes deficits on DMS and N-back tasks [34-36] and on the Brown-Peterson task when letter trigrams that involve novel combinations of well-known elements are used [37]. However, scopolamine injections do not lead to impairments on simple measures of short-term memory such as digit span[38] and the recency component of a serial position curve[39], suggesting a lack of effect on working memory for highly familiar stimuli such as words or numbers. For these stimuli, previously modified synaptic connections might be sufficiently strong to maintain activity without cholinergic modulation.
An experiment by McGaughy et al. directly tested the hypothesis that cholinergic modulation regulates memory for novel but not familiar stimuli by analysing DMS performance in rats that were given injections of a neurotoxin into entorhinal cortex which selectively lesions the cholinergic innervation to this region [40]. These selective lesions did not impair DNMS function for familiar odours at delays of 15 minutes, but did cause a significant and selective deficit at 15 minutes for odours that were novel to the rats. This impairment could be due to loss of the cholinergic modulation of persistent spiking activity, coupled with a loss of cholinergic enhancement of synaptic modification [41]. In monkeys, selective lesions of the cholinergic innervation to the perirhinal cortex caused impairments in DNMS performance at a delay of 30 seconds for novel visual stimuli[42]. Parahippocampal neurons can maintain persistent spiking for periods longer than 15 minutes (Giocomo, Tahvildari and Hasselmo, unpublished data), but in the behavioural tasks with long delays it is likely that cholinergic regulation of persistent activity also enhances encoding of novel stimuli via long-term synaptic modification so that later matching can be performed even after persistent activity terminates.
Encoding into long-term memory
The cholinergic lesion studies extend earlier work in monkeys demonstrating that injections of scopolamine into the perirhinal cortex (which likely also affect the entorhinal cortex[43]) impair the encoding of new visual stimuli but not the recognition of stimuli learned before the scopolamine injection[43,44]. By contrast, infusion into the inferotemporal cortex or dentate gyrus did not cause impaired performance[43]. Crossed lesions of cholinergic innervation and cortical regions similarly impair learning of new scenes and objects [45]. Consistent with these data, systemic injections of scopolamine in humans strongly impair the encoding of new words into memory for subsequent free recall or paired associate memory, without affecting the recall of words learned before scopolamine injections[46]. In line with the effects of scopolamine, damage to the hippocampus does not impair digit span[17], but does impair performance on tasks requiring longer retention or retention of complex non-verbalizable stimuli [21]. The data summarized above indicate that scopolamine may be blocking a necessary mechanism for the formation of long-term memories involving novel stimuli in the hippocampus, possibly by inhibiting sustained spiking activity in the entorhinal cortex that provides input necessary to modify synapses in the hippocampus [25].
In summary, reductions of cholinergic modulation impair working memory for novel stimuli, possibly due to blockade of cellular effects reviewed in the next section.
Cellular physiology
The proposal of a working memory system for novel stimuli presented here was initially motivated by intracellular single-cell recordings of memory mechanisms[8,47]. Recordings in the laboratory of Angel Alonso demonstrated that single neurons in layer II of slice preparations of the entorhinal cortex, isolated from other neurons by pharmacological blockade of excitatory and inhibitory synaptic transmission, can maintain memory for prior input in the form of persistent spiking activity (Figure 3). This maintenance of persistent spiking depends on the application of acetylcholine or other drugs that activate muscarinic acetylcholine receptors[47] and is blocked by muscarinic antagonists such as atropine[8]. In the absence of acetylcholine, a depolarizing intracellular current injection causes the neuron to fire a train of spikes during the depolarization, but when the current injection is stopped, the membrane potential of the cell falls back to resting potential and the firing stops. However, in the presence of acetylcholine, a current injection of only a few hundred milliseconds in layer II causes the membrane potential to remain depolarized after the current injection is stopped, and the cell continues to generate spikes for an extended period of many seconds or even minutes without current injection or synaptic input.
Figure 3.
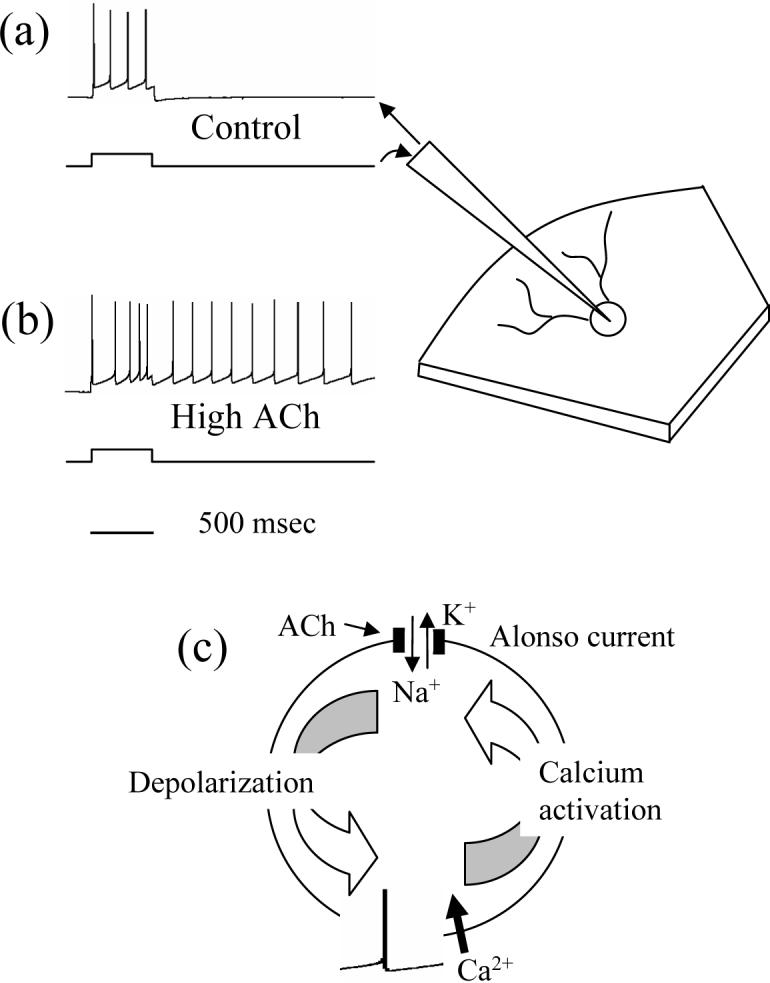
Schematic of persistent spiking observed with intracellular recording from neurons in layer II of entorhinal cortex slices. (a) In control conditions, an entorhinal cortical neuron responds to a brief current injection by spiking during the current injection, and falling silent after the current injection ends. (b) With activation of acetylcholine receptors, the same neuron responds to a current injection with spiking which persists for many seconds or minutes after the end of the current injection. (c) Intrinsic mechanisms of regenerative spiking. The Alonso current is activated by a combination of extracellular acetylcholine (ACh) and by spiking activity induced by current injection. Spiking activates voltage-sensitive calcium channels that increase intracellular calcium. Calcium activates the Alonso current which increases Na+ influx and causes more depolarization. This depolarization causes further spiking activity that causes calcium influx that further activates the Alonso current, completing a cycle which can underlie persistent spiking.
The phenomenon underlying self-sustained spiking is sometimes referred to as an afterdepolarization or plateau potential. This phenomenon continues even when both excitatory and inhibitory synaptic transmission are blocked, indicating its dependence on intrinsic mechanisms in a single neuron, rather than on excitatory recurrent connectivity between neurons [8]. The effect can return after brief periods of hyperpolarizing current injection, indicating resistance to transient distractors. A similar plateau potential phenomenon has also been described in layer V of prefrontal cortical slices[48]. However, the fact that the PFC is not sufficient to mediate working memory performance for novel stimuli after parahippocampal lesions[19,22] suggests that without prior familiarization, PFC neurons may not receive sufficient synaptic input to represent novel sensory stimuli, consistent with a weaker spiking response to novel stimuli compared to familiar stimuli in some experiments [49].
In the entorhinal cortex, plateau potentials appear to arise from a calcium-sensitive non-specific cation current[8,13] which strongly depolarizes neurons. Here we refer to this current in entorhinal cortex as the Alonso current, after Prof. Angel Alonso, who died in 2005. This mechanism for intrinsic working memory relies on the conjunction of acetylcholine and cellular spiking[8]. Current injection in the absence of acetylcholine does not initiate sustained spiking. But in the presence of acetylcholine, the generation of spiking by intracellular current injection or synaptic stimulation causes the neuron to enter an internal regenerative cycle of sustained spiking. Each new spike activates voltage-sensitive calcium channels, and the new influx of calcium activates the Alonso current[13]. This current causes additional depolarization, leading to another spike, which again activates voltage-sensitive calcium channels that further perpetuate the cycle (see Figure 3C). The Box summarizes computational modelling of these single cell mechanisms for working memory.
In summary, the cellular processes in parahippocampal cortices provide a mechanism of persistent spiking suitable for working memory for novel stimuli, which could also underlie some of the data from unit recording studies in parahippocampal structures reviewed in the next section.
Recording of spiking activity during behaviour
The theory that the Alonso current in the entorhinal cortex contributes to the maintenance of working memory is supported by recordings of the spiking activity of single neurons in animals performing delayed matching tasks. Sustained spiking activity during delay periods has been shown for neurons in parahippocampal cortices, including the entorhinal cortex, in rats performing a continuous DNMS task with odours[10], and in monkeys during performance of DMS tasks with complex visual stimuli[30]. Delay activity in the entorhinal cortex proves resistant to distractor stimuli presented between the sample and match stimulus[30]. This is consistent with data from entorhinal cortex slices showing that the spiking activity of single cells will restart after a brief hyperpolarizing current injection[8,13]. Interestingly, delay activity in the perirhinal cortex does not persist after intervening distractors[50]. No electrophysiological study has yet looked at effects of cholinergic blockade on persistent spiking for novel stimuli in the parahippocampal cortices.
In addition to sustained activity during delay periods, some entorhinal neurons show enhancement of spiking activity for a match stimulus relative to the response to the same stimulus presented as sample, whereas most neurons show suppression of activity in response to a match [10,30] and to repetitions of distractor stimulus [30,50,51]. Thus, there are multiple possible mechanisms for making a memory-guided decision in a matching task — including sustained delay activity, match enhancement and repetition suppression — which are not mutually exclusive. There is additional suppression as a novel stimulus becomes familiar [52], which may play a crucial role in selectively gating parahippocampal activity to only maintain novel stimuli [53,54].
In summary, unit recording data support the idea that sustained spiking in parahippocampal cortices could provide working memory due to cellular mechanisms of persistent spiking.
BOX: Computational modelling of single cell mechanisms for working memory
The hypothetical link between the cellular data and behavioural data presented here has been described and analyzed in detailed computational models. John Lisman initially proposed that intrinsic afterdepolarization such as that shown by Alonso[8] and others forms a cellular basis for working memory[11,55]. This section describes models showing how this cellular mechanism could underlie working memory[11-13,56] and encoding into episodic memory[24,25,55].
Working memory
Biophysical compartmental simulations of layer II non-stellate neurons[13] demonstrate how the Alonso current can underlie the sustained delay period spiking in the entorhinal cortex seen with extracellular unit recording in rats and monkeys during the performance of delayed matching tasks[10,30]. These detailed models extended earlier models showing that afterdepolarization could underlie working memory function[11,55]. Compartmental simulations of layer II neurons were combined in network simulations that included detailed models of inhibitory interneurons[13] to demonstrate the mechanisms for many additional response properties of single neurons recorded in the entorhinal cortex, including match and non-match enhancement and suppression[10,30].
Models also illustrate why intrinsic mechanisms for persistent spiking provide a mechanism for working memory for novel stimuli [40], in contrast to most models using excitatory recurrent connectivity which are primarily appropriate for familiar stimuli. Familiar stimuli can be retained in a circuit in which Hebbian synaptic modification of glutamatergic synapses has been previously induced, as shown in Figure 4A. Modified connections (arrows) allow reverberatory interactions between neurons in the pattern, causing persistent spiking (Figure 4A). In contrast, the neurons responding to novel stimuli do not have excitatory recurrent connections with other neurons coding the same pattern. However, as shown in Figure 4B, the internal regenerative process mediated by the Alonso current allows maintenance of spiking activity for novel stimuli in the absence of synaptic interactions. As shown in Figure 4C, blockade of cholinergic receptors will block the cellular mechanisms for persistent spiking, and thereby reduce the capacity to maintain working memory for novel stimuli, as suggested by impairments caused by scopolamine and cholinergic lesions[34,36,40,42].
Figure 4.

(a) Maintenance of familiar information does not require acetylcholine, because the familiar input matches the pattern of excitatory glutamatergic connectivity between neurons, allowing maintenance of persistent spiking. The arrows between circles represent excitatory recurrent synapses strengthened by prior learning, which spread activity back and forth between neurons representing the familiar pattern. (b) Maintenance of novel information is possible with cholinergic activation of intrinsic mechanisms for persistent spiking (arrows within circles), because individual neurons have internal regenerative mechanisms for spiking without excitatory recurrent connections (discussed in [40]). (c) Blockade of muscarinic acetylcholine receptors (for example by scopolamine) blocks the intrinsic mechanisms for persistent spiking, impairing working memory for novel stimuli.
Encoding of long-term memory
Modelling also demonstrates how these mechanisms could enhance encoding into long-term memory. In particular, the capacity of the entorhinal cortex for maintaining persistent spiking could allow spikes to fall within the narrow time window that is required for spike timing-dependent plasticity[12,24,25], thereby bridging behavioural input that is separated by long time intervals and enabling the formation of associations between sequential patterns. Models of these mechanisms utilize timing of intrinsic spiking relative to the phase of cortical theta rhythm oscillations to maintain information about the sequential order of an item [12,24,25], consistent with evidence for cholinergic modulation of cortical theta rhythm oscillations. Modeling also demonstrates potential mechanisms for the extended maintenance of graded levels of firing frequency in layer V of entorhinal cortex [57] which could provide a mechanism for maintaining temporal context for memory encoding [58]. End of Box.
Summary and outstanding questions
The experimental data and computational modelling reviewed here support the hypothesis that intrinsic single neuron mechanisms in parahippocampal cortices mediate working memory for novel stimuli. Studies using techniques at a number of different levels support this hypothesis. However, further work at the behavioural level is needed to test whether blockade of cholinergic receptors will selectively impair active maintenance of novel but not familiar stimuli in the same task in humans, possibly by using experimental designs that are similar to those used to show impairments of memory for novel visual objects in amnesic subjects[22].
The work presented here also generates specific predictions about the nature of unit activity in parahippocampal structures. In particular, during the delay period of a DMS task, unit activity should be more sensitive to muscarinic antagonists such as scopolamine for novel than for highly familiar stimuli. Studies have shown that scopolamine does not reduce repetition suppression of unit responses[59], but have not tested scopolamine effects on match enhancement or delay activity. Unfortunately, the ability to quantify firing rate responses to stimuli usually requires showing them repeatedly, thereby reducing their novelty[30], but the effect could be analysed with a large set of novel stimuli.
The cholinergic mechanisms in parahippocampal structures described here could act as a buffer for long-term encoding[1,12], which has the capacity to integrate and hold information from multiple modalities, to maintain information about the order of items in working memory, and to feed information into long-term episodic memory. The episodic buffer could be supported by a selected subset of neurons firing in response to the multi-modal convergent input to the entorhinal cortex. The Alonso current could allow entorhinal neurons coding multi-modal components of an episodic memory to persist in activity, representing the distinct relationship between events and items in a novel episode, and allowing encoding into episodic memory in the hippocampus.
The data described here provides a link between the behavioural function of working memory and possible mechanisms at the cellular and circuit level for holding a sustained representation of novel information across a temporal delay. Understanding these physiological mechanisms and relating them to behaviour is critical for our understanding of working memory.
Outstanding questions:
Do systemic injections of scopolamine cause stronger impairments of working memory for novel versus familiar stimuli in the same study in humans?
Does local infusion of scopolamine into entorhinal and perirhinal cortex selectively impair working memory for novel stimuli but not familiar stimuli in rats?
Does systemic or local infusion of scopolamine reduce single neuron spiking activity in parahippocampal cortices during the delay period of delayed match to sample tasks using novel stimuli, but not familiar stimuli?
Do mechanisms of persistent spiking shown with intracellular recording in vitro interact with in vivo theta rhythm oscillations in parahippocampal structures[12,25,60]?
Acknowledgments
Supported by NINDS 41636, NIMH 60013, NIMH 61492, NIMH 60450, NSF Science of Learning Center SBE0354378 and NIDA 16454 as part of the joint NSF/NIH program for Collaborative Research in Computational Neuroscience (CRCNS).
REFERENCES
- 1.Baddeley A. The episodic buffer: a new component of working memory? Trends Cogn Sci. 2000;4(11):417–423. doi: 10.1016/s1364-6613(00)01538-2. [DOI] [PubMed] [Google Scholar]
- 2.Eichenbaum H, Cohen NJ. From conditioning to conscious recollection: Memory systems of the brain. Oxford University Press; 2001. [Google Scholar]
- 3.Fuster JM. Memory networks in the prefrontal cortex. Prog Brain Res. 2000;122:309–316. doi: 10.1016/s0079-6123(08)62147-0. [DOI] [PubMed] [Google Scholar]
- 4.Ranganath C, D'Esposito M. Directing the mind's eye: prefrontal, inferior and medial temporal mechanisms for visual working memory. Curr Opin Neurobiol. 2005;15(2):175–182. doi: 10.1016/j.conb.2005.03.017. [DOI] [PubMed] [Google Scholar]
- 5.Stern CE, et al. Medial temporal and prefrontal contributions to working memory tasks with novel and familiar stimuli. Hippocampus. 2001;11(4):337–346. doi: 10.1002/hipo.1048. [DOI] [PubMed] [Google Scholar]
- 6.Schon K, et al. Scopolamine reduces persistent activity related to long-term encoding in the parahippocampal gyrus during delayed matching in humans. J. Neurosci. 2005;25(40):9112–9123. doi: 10.1523/JNEUROSCI.1982-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Durstewitz D, et al. Neurocomputational models of working memory. Nat Neurosci. 2000;3:1184–1191. doi: 10.1038/81460. [DOI] [PubMed] [Google Scholar]
- 8.Klink R, Alonso A. Muscarinic modulation of the oscillatory and repetitive firing properties of entorhinal cortex layer II neurons. J. Neurophysiol. 1997;77(4):1813–1828. doi: 10.1152/jn.1997.77.4.1813. [DOI] [PubMed] [Google Scholar]
- 9.Egorov AV, et al. Graded persistent activity in entorhinal cortex neurons. Nature. 2002;420(6912):173–178. doi: 10.1038/nature01171. [DOI] [PubMed] [Google Scholar]
- 10.Young BJ, et al. Memory representation within the parahippocampal region. J. Neurosci. 1997;17:5183–5195. doi: 10.1523/JNEUROSCI.17-13-05183.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lisman JE, Idiart MA. Storage of 7 +/− 2 short-term memories in oscillatory subcycles. Science. 1995;267(5203):1512–1515. doi: 10.1126/science.7878473. [DOI] [PubMed] [Google Scholar]
- 12.Jensen O, Lisman JE. Novel lists of 7+/−2 known items can be reliably stored in an oscillatory short-term memory network: interaction with long-term memory. Learning and Memory. 1996;3:257–263. doi: 10.1101/lm.3.2-3.257. [DOI] [PubMed] [Google Scholar]
- 13.Fransen E, et al. Simulations of the role of the muscarinic-activated calcium-sensitive nonspecific cation current INCM in entorhinal neuronal activity during delayed matching tasks. J Neurosci. 2002;22(3):1081–1097. doi: 10.1523/JNEUROSCI.22-03-01081.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Asaad WF, et al. Neural activity in the primate prefrontal cortex during associative learning. Neuron. 1998;21(6):1399–1407. doi: 10.1016/s0896-6273(00)80658-3. [DOI] [PubMed] [Google Scholar]
- 15.Eacott MJ, et al. Preserved recognition memory for small sets, and impaired stimulus identification for large sets, following rhinal cortex ablations in monkeys. Eur J Neurosci. 1994;6:1466–1478. doi: 10.1111/j.1460-9568.1994.tb01008.x. [DOI] [PubMed] [Google Scholar]
- 16.Otto T, Eichenbaum H. Complementary roles of the orbital prefrontal cortex and the perirhinal entorhinal cortices in an odor-guided delayed-nonmatching- to-sample task. Behavioral Neuroscience. 1992;106(5):762–775. doi: 10.1037//0735-7044.106.5.762. [DOI] [PubMed] [Google Scholar]
- 17.Corkin S. Lasting consequences of bilateral medial temporal lobectomy: Clinical course and experimental findings in H.M. Semin. Neurol. 1984;4:249–259. [Google Scholar]
- 18.Gaffan D, Murray EA. Monkeys (Macaca fascicularis) with rhinal cortex ablations succeed in object discrimination learning despite 24-hr intervals and fail at matching to sample despite double sample presentations. Behav Neurosci. 1992;106:30–38. doi: 10.1037//0735-7044.106.1.30. [DOI] [PubMed] [Google Scholar]
- 19.Squire LR, et al. The medial temporal lobe. Annu Rev Neurosci. 2004;27:279–306. doi: 10.1146/annurev.neuro.27.070203.144130. [DOI] [PubMed] [Google Scholar]
- 20.Olson IR, et al. Working memory for conjunctions relies on the medial temporal lobe. J Neurosci. 2006;26(17):4596–4601. doi: 10.1523/JNEUROSCI.1923-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Olson IR, et al. Visual working memory is impaired when the medial temporal lobe is damaged. J. Cogn. Neurosci. 2006;18:1087–1097. doi: 10.1162/jocn.2006.18.7.1087. [DOI] [PubMed] [Google Scholar]
- 22.Ranganath C, Blumenfeld RS. Doubts about double dissociations between short- and long-term memory. Trends Cogn Sci. 2005;9(8):374–380. doi: 10.1016/j.tics.2005.06.009. [DOI] [PubMed] [Google Scholar]
- 23.Alvarez P, et al. The animal model of human amnesia: long-term memory impaired and short-term memory intact. Proc. Natl. Acad. Sci. U S A. 1994;91(12):5637–5641. doi: 10.1073/pnas.91.12.5637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Koene RA, et al. Modeling goal-directed spatial navigation in the rat based on physiological data from the hippocampal formation. Neural Netw. 2003;16(5−6):577–584. doi: 10.1016/S0893-6080(03)00106-0. [DOI] [PubMed] [Google Scholar]
- 25.Jensen O, Lisman JE. Hippocampal sequence-encoding driven by a cortical multi-item working memory buffer. Trends Neurosci. 2005;28(2):67–72. doi: 10.1016/j.tins.2004.12.001. [DOI] [PubMed] [Google Scholar]
- 26.Braver TS, et al. A parametric study of prefrontal cortex involvement in human working memory. Neuroimage. 1997;5(1):49–62. doi: 10.1006/nimg.1996.0247. [DOI] [PubMed] [Google Scholar]
- 27.Ranganath C, D'Esposito M. Medial temporal lobe activity associated with active maintenance of novel information. Neuron. 2001;31(5):865–873. doi: 10.1016/s0896-6273(01)00411-1. [DOI] [PubMed] [Google Scholar]
- 28.Ranganath C, Rainer G. Neural mechanisms for detecting and remembering novel events. Nat Rev Neurosci. 2003;4(3):193–202. doi: 10.1038/nrn1052. [DOI] [PubMed] [Google Scholar]
- 29.Schon K, et al. Persistence of parahippocampal representation in the absence of stimulus input enhances long-term encoding: a functional magnetic resonance imaging study of subsequent memory after a delayed match-to-sample task. J Neurosci. 2004;24(49):11088–11097. doi: 10.1523/JNEUROSCI.3807-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Suzuki WA, et al. Object and place memory in the macaque entorhinal cortex. Neuroreport. 1997;7:2231–2235. doi: 10.1152/jn.1997.78.2.1062. [DOI] [PubMed] [Google Scholar]
- 31.Penetar DM, McDonough JH. Effects of cholinergic drugs on delayed match to sample performance of Rhesus monkeys. Pharmacol. Biochem. Behav. 1983;19:963–967. doi: 10.1016/0091-3057(83)90399-4. [DOI] [PubMed] [Google Scholar]
- 32.Bolhuis JJ, et al. Effects of scopolamine on performance of rats in a delayed-response radial maze task. Physiol Behav. 1988;43:403–409. doi: 10.1016/0031-9384(88)90111-4. [DOI] [PubMed] [Google Scholar]
- 33.Anagnostaras SG, et al. Selective cognitive dysfunction in acetylcholine M1 muscarinic receptor mutant mice. Nature Neuroscience. 2003;6:51–58. doi: 10.1038/nn992. [DOI] [PubMed] [Google Scholar]
- 34.Robbins TW, et al. Effects of scopolamine on delayed-matching-to-sample and paired associates tests of visual memory and learning in human subjects: comparison with diazepam and implications for dementia. Psychopharmacology (Berl) 1997;134(1):95–106. doi: 10.1007/s002130050430. [DOI] [PubMed] [Google Scholar]
- 35.Koller G, et al. Effects of scopolamine on matching to sample paradigm and related tests in human subjects. Neuropsychobiology. 2003;48(2):87–94. doi: 10.1159/000072883. [DOI] [PubMed] [Google Scholar]
- 36.Green A, et al. Muscarinic and nicotinic receptor modulation of object and spatial n-back working memory in humans. Pharmacol Biochem Behav. 2005;81(3):575–584. doi: 10.1016/j.pbb.2005.04.010. [DOI] [PubMed] [Google Scholar]
- 37.Beatty WW, et al. Patterns of memory failure after scopolamine treatment: Implications for cholinergic hypotheses of dementia. Behav. Neural Biol. 1986;45:196–211. doi: 10.1016/s0163-1047(86)90772-7. [DOI] [PubMed] [Google Scholar]
- 38.Broks P, et al. Modelling dementia: Effects of scopolamine on memory and attention. Neuropsychologia. 1988;26:685–700. doi: 10.1016/0028-3932(88)90004-8. [DOI] [PubMed] [Google Scholar]
- 39.Crow TJ, Grove-White IG. An analysis of the learning deficit following hyoscine administration to man. Br. J. Pharmacol. 1973;49:322–327. doi: 10.1111/j.1476-5381.1973.tb08379.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.McGaughy J, et al. Cholinergic deafferentation of the entorhinal cortex in rats impairs encoding of novel but not familiar stimuli in a delayed non-match to sample task (DNMS). J. Neurosci. 2005;25(44):10273–10281. doi: 10.1523/JNEUROSCI.2386-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Huerta PT, Lisman JE. Heightened synaptic plasticity of hippocampal CA1 neurons during a cholinergically induced rhythmic state. Nature. 1993;364:723–725. doi: 10.1038/364723a0. [DOI] [PubMed] [Google Scholar]
- 42.Turchi J, et al. Effects of cholinergic deafferentation of the rhinal cortex on visual recognition memory in monkeys. Proc Natl Acad Sci U S A. 2005;102(6):2158–2161. doi: 10.1073/pnas.0409708102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Tang Y, et al. Effects of muscarinic blockade in perirhinal cortex during visual recognition. Proc. Natl. Acad. Sci. USA. 1997;94(23):12667–12669. doi: 10.1073/pnas.94.23.12667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Aigner TG, et al. Comparison of the effects of scopolamine administered before and after acquisition in a test of visual recognition memory in monkeys. Behav. Neural Biol. 1991;55(1):61–67. doi: 10.1016/0163-1047(91)80127-z. [DOI] [PubMed] [Google Scholar]
- 45.Easton A, et al. Unilateral lesions of the cholinergic basal forebrain and fornix in one hemisphere and inferior temporal cortex in the opposite hemisphere produce severe learning impairments in rhesus monkeys. Cereb Cortex. 2002;12(7):729–736. doi: 10.1093/cercor/12.7.729. [DOI] [PubMed] [Google Scholar]
- 46.Atri A, et al. Blockade of central cholinergic receptors impairs new learning and increases proactive interference in a word paired-associate memory task. Behav Neurosci. 2004;118(1):223–236. doi: 10.1037/0735-7044.118.1.223. [DOI] [PubMed] [Google Scholar]
- 47.Shalinsky MH, et al. Muscarinic activation of a cation current and associated current noise in entorhinal-cortex layer-II neurons. J Neurophysiol. 2002;88(3):1197–1211. doi: 10.1152/jn.2002.88.3.1197. [DOI] [PubMed] [Google Scholar]
- 48.Haj-Dahmane S, Andrade R. Ionic mechanism of the slow afterdepolarization induced by muscarinic receptor activation in rat prefrontal cortex. J. Neurophysiol. 1998;80(3):1197–1210. doi: 10.1152/jn.1998.80.3.1197. [DOI] [PubMed] [Google Scholar]
- 49.Xiang JZ, Brown MW. Neuronal responses related to long-term recognition memory processes in prefrontal cortex. Neuron. 2004;42(5):817–829. doi: 10.1016/j.neuron.2004.05.013. [DOI] [PubMed] [Google Scholar]
- 50.Miller EK, et al. Activity of neurons in anterior inferior temporal cortex during a short-term memory task. J. Neurosci. 1993;13:1460–1478. doi: 10.1523/JNEUROSCI.13-04-01460.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Fahy FL, et al. Neuronal activity related to visual recognition memory: long term memory and the encoding of recency and familiarity information in the primate anterior and medial inferior temporal and rhinal cortex. Exp Brain Res. 1993;96:457–472. doi: 10.1007/BF00234113. [DOI] [PubMed] [Google Scholar]
- 52.Riches IP, et al. The effects of visual stimulation and memory on neurons of the hippocampal formation and the neighboring parahippocampal gyrus and inferior temporal cortex of the primate. J Neurosci. 1991;11(6):1763–1779. doi: 10.1523/JNEUROSCI.11-06-01763.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Warburton EC, et al. Cholinergic neurotransmission is essential for perirhinal cortical plasticity and recognition memory. Neuron. 2003;38(6):987–996. doi: 10.1016/s0896-6273(03)00358-1. [DOI] [PubMed] [Google Scholar]
- 54.Bogacz R, Brown MW. Comparison of computational models of familiarity discrimination in the perirhinal cortex. Hippocampus. 2003;13(4):494–524. doi: 10.1002/hipo.10093. [DOI] [PubMed] [Google Scholar]
- 55.Jensen O, Lisman JE. Theta/gamma networks with slow NMDA channels learn sequences and encode episodic memory: role of NMDA channels in recall. Learning and Memory. 1996;3:264–278. doi: 10.1101/lm.3.2-3.264. [DOI] [PubMed] [Google Scholar]
- 56.Lansner A, Fransén E. Modeling Hebbian cell assemblies comprised of cortical neurons. Network. 1992;3:105–119. [Google Scholar]
- 57.Fransen E, et al. Mechanism of graded persistent cellular activity of entorhinal cortex layer v neurons. Neuron. 2006;49(5):735–746. doi: 10.1016/j.neuron.2006.01.036. [DOI] [PubMed] [Google Scholar]
- 58.Howard MW, Kahana MJ. A distributed representation of temporal context. J. Math. Psychol. 2002;46(3):269–299. [Google Scholar]
- 59.Miller EK, Desimone R. Scopolamine affects short-term memory but not inferior temporal neurons. Neuroreport. 1993;4(1):81–84. doi: 10.1097/00001756-199301000-00021. [DOI] [PubMed] [Google Scholar]
- 60.Lee H, et al. Phase locking of single neuron activity to theta oscillations during working memory in monkey extrastriate visual cortex. Neuron. 2005;45:147–156. doi: 10.1016/j.neuron.2004.12.025. [DOI] [PubMed] [Google Scholar]