

Abstract
Purpose
To identify the underlying genetic defect in a three-generation family with five members affected with dominant bilateral congenital cataract and microcornea.
Methods
Detailed family history and clinical data were recorded. Mutation screening in the candidate genes, CRYAA, CRYBB1, MAF, GJA3, and GJA8, was performed by bidirectional sequencing of the amplified products.
Results
Affected individuals had a jellyfish-like cataract in association with microcornea. Sequencing of GJA8 (connexin 50) showed a novel, heterozygous c.134G→C change that resulted in the substitution of a highly conserved tryptophan by serine (p.W45S). This sequence change segregated completely with the disease phenotype and was not observed in 108 ethnically matched controls (216 chromosomes). However, an identical substitution has previously been described in GJA3 (connexin 46) leading to autosomal dominant nuclear cataract without microcornea.
Conclusions
This is a novel mutation identified in the first transmembrane domain (M1) of GJA8. These findings further expand the mutation spectrum of connexin 50 (Cx50) in association with congenital cataract and microcornea.
Introduction
Congenital cataract is one of the common causes of visual impairment and childhood blindness. Its incidence is estimated to be 2.2-2.49 per 10,000 live births [1,2]. Wide clinical and genetic variability has been observed. Nearly one-third of the cases show a positive family history of which autosomal dominant inheritance is the most common [3]. Congenital cataract can occur either as an isolated anomaly, in association with other ocular anomalies, or as a component of multi-systemic disorder. Microcornea-cataract syndrome (OMIM 116150) is characterized by the association of congenital cataract and microcornea without any other systemic anomaly or dysmorphism. Mutations in CRYAA (OMIM 123580), CRYBB1 (OMIM 600929), GJA8 (OMIM 600897), and MAF (OMIM 177075) have been reported in families affected with cataract-microcornea syndrome [4-7].
The eye lens is an avascular structure, and intercellular transport of small biomolecules of less than 1 kDa is mediated through connexins (Cx) that encode gap junction channel proteins [8,9]. In humans, at least 20 connexins classified into three families have been identified [10,11]. Three of these connexins belong to the α-connexin family and are expressed in the lens, connexin 43 (Cx43, GJA1) in the epithelial cells, connexin 46 (Cx46, GJA3), and Cx50 (GJA8) in lens fibers [12-15]. Lens fibers are connected to the epithelial cells via gap junctions and are dependent on a metabolically active epithelium for maintenance of the intracellular ionic conditions necessary to prevent precipitation of crystallins and hence cataract formation [16]. Cx46 and Cx50 are responsible for joining the lens cells into a functional syncytium; in addition, Cx50 is also important for lens growth [17].
We encountered a three-generation Indian family with bilateral congenital cataract at the Dr. Daljit Singh Eye Hospital, Amritsar, India. The appearance of the cataract was reminiscent of a jellyfish. All affected individuals showed microcornea in association with congenital cataract. Upon sequence analysis of the candidate genes, CRYAA, CRYBB1, MAF, GJA3, and GJA8, we identified a heterozygous c.134G→C change in GJA8 that resulted in the substitution of a highly conserved tryptophan by serine at codon 45 (p.W45S). This change co-segregated completely with the disease phenotype.
Methods
Family description
The index case, a 12-year-old child, was diagnosed with bilateral cataract. The family history revealed five affected members in three generations (Figure 1). A detailed ophthalmologic examination, which included slit lamp examination and photography of the affected lenses, was performed on seven members of the family and revealed that five members were bilaterally affected (four had a history of cataract extraction in childhood) and two individuals were unaffected.
Figure 1.
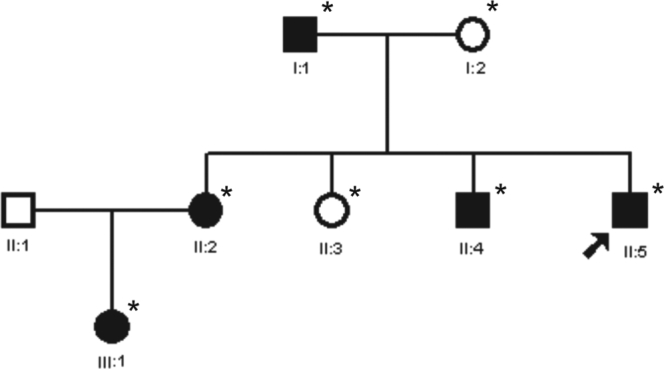
Pedigree of a family with individuals affected by congenital cataract and microcornea. The pedigree of an autosomal dominant congenital cataract (ADCC) family with affected individuals (indicated as filled circles and squares) in three generations is shown. All the affected members showed microcornea in association with congenital cataract. The asterisk indicates those individuals who underwent ophthalmologic examinations and were genetically investigated. The proband (II:5) is indicated with an arrow.
Mutation analysis
Informed consent was obtained from each individual studied. This study was approved by the Ethics Review Board of the Guru Nanak Dev University, consistent with the provisions of the Declaration of Helsinki. Blood was drawn and DNA was isolated by standard methods. Mutation screening was performed in the exonic regions of candidate genes, CRYAA (GenBank NM_000394), CRYBB1 (NM_001887), MAF (NM_001031804), GJA3 (GenBank NM_021954), and GJA8 (GenBank NM_005267), localized at 21q22.3, 22q11.2-q12.2, 16q23.1, 13q11, and 1q21, respectively. The coding regions and exon-intron boundaries of the candidate genes were amplified using previously published primer sequences [4,5,18-20]. Initially, genomic DNA from two affected and one unaffected individual was amplified. Upon identification of a nucleotide substitution in GJA8 in tested affected individuals, three more affected and one unaffected individual’s DNA were tested. Amplification was performed in 25 μl reactions containing 50 ng genomic DNA, 10 pmoles of each forward and reverse primer, 200 μM dNTP, 10X polymerase chain reaction (PCR) buffer, 1.5 mM MgCl2, and 0.25 U Taq DNA polymerase (AmpliTaq Gold; Applied Biosystems, Foster City, CA). To amplify GC-rich amplicons, 5% DMSO was used in the reaction. Amplification conditions consisted of an initial denaturation step at 95 °C for 5 min followed by 35 cycles consisting of a denaturation step at 95 °C for 10 s, optimal annealing step for 30 s at temperatures ranging between 53 °C-61 °C for different amplicons, and an extension at 72 °C for 45 s followed by a final extension step at 72 °C for 10 min. PCR products were purified using a PCR products purification kit (QIAquick; Qiagen, Valencia, CA) and sequenced bidirectionally with ABI BigDyeTM Terminator Cycle Sequencing Ready Reaction Kit version 3.1 (Applied Biosystems) as described elsewhere [18]. Sequencing results were assembled and analyzed using the SeqMan II program of the Lasergene package (DNA STAR Inc., Madison, WI).
Results
Phenotype description
The lens opacity appeared axial, extending from the anterior capsule to the posterior capsule. At the anterior end, the round opacity was about 2 mm in diameter and placed slightly eccentrically toward 10 o'clock. There appeared to be a 1 mm x 1/2 mm blunt projection extending from the anterior end on the nasal side. On the temporal side, there was a fan-like opacity, which tends to hide the deeper structure of the opacity, comprising about a dozen finger-like projections radiating in all directions (Figure 2). The cataracts in this family had a jellyfish-like appearance, different from the coralliform cataract, which has a more delicate and sparse structure and axial branches that are sharp and thin. All five affected individuals also had microcornea, their corneal diameter was less than 10 mm in both horizontal and vertical meridians (average normal diameters are 12.6 mm and 11.7 mm, respectively [21]. Apart from congenital cataract and microcornea, no other ocular anomalies such as microphthalmia, amblyopia, strabismus, or glaucoma were detected in any of these affected members.
Figure 2.

Photograph (three-dimensional lens) of a patient taken through a slit lamp. The lens opacity is axial, extending from the anterior capsule to the posterior capsule. At the anterior end, there is a round opacity about 2 mm in diameter, placed slightly eccentrically toward 10 o'clock. There is a 1 mm x 1/2 mm blunt projection on the nasal side. On the temporal side, there seems to be a fan-like opacity, which tends to hide the deeper structure of the opacity, comprising about a dozen finger-like projections going in all directions. The cataract appears like a jellyfish.
Mutation screening
Bidirectional sequencing of the coding region of GJA8 in the five affected (I:1, II:2, II:4, II:5, and III:1) and two unaffected individuals (I:2 and II:3) showed a heterozygous change, G>C (), at position 134 (c.134G→C) from the translation start site in the affected individuals. The alteration was not seen in any of the unaffected family members tested nor in the 108 unrelated control subjects (216 chromosomes) from the same Northern Indian population (data not shown) but was confirmed in all affected individuals. This nucleotide substitution replaces an evolutionarily highly conserved tryptophan with serine at amino acid position 45 (p.W45S) in the first α-helical transmembrane domain 1 (M1) of connexin 50.
Discussion
In the present study, we report a novel p.W45S substitution in the connexin 50 polypeptide associated with jellyfish-like cataract and microcornea. The p.W45S substitution is likely to cause disease since it segregated with the disorder and was not detected either in any of the tested unaffected family members nor in the 108 unrelated controls. The tryptophan-45 in connexin 50 is predicted to lie within the first transmembrane domain (M1) and is well conserved in different species (Figure 3B). Furthermore, the mutation is adjacent to a previously reported mutation, p.V44E, also associated with cataract and microcornea [6]. Recently, Ponnam et al. [22] have identified a frameshift mutation, p.T203fs, associated with autosomal recessive total cataract, microcornea, and microphthalmia in one of two affected siblings.
Figure 3.

DNA sequence of a part of GJA8 from an unaffected and an affected individual with jelly-fish like cataract followed by a multiple alignment of partial amino acid sequences of connexin 50 from different species and of connexin 46 from homo sapiens. A: DNA sequence analysis of a part of GJA8 from an unaffected and an affected individual are displayed in the electropherograms (forward strand; individuals II:3 and II:5, respectively). The wild-type G in the sequence of the unaffected individual and the heterozygous c.134G→C change resulting in substitution of tryptophan-45 by serine (p.W45S) in the affected individual’s sequence are indicated by arrows. F indicates the portion of GJA8 sequence, the forward strand, in the unaffected and affected individuals. B: A multiple alignment of partial amino acid sequences of connexin 50 from different species and of connexin 46 from Homo sapiens is shown. The alignment data indicate that tryptophan at position 45 (indicated by an arrow) is highly conserved in different species in connexin 50 and also in connexin 46 in Homo sapiens.
Different mutations have been detected in GJA8 in association with congenital cataract and significant interfamilial phenotypic variability has been observed. The phenotypes in most of the cases that have mutations in GJA8 have been described as zonular nuclear pulverulent cataract with dust-like opacities. The cataract phenotype in the present family differs from these because there are no “pulverized” dust-like opacities in the lens. Diverse mutational mechanisms like a dominant negative effect for p.P88S and p.P88Q [20,23,24] or a loss of function for p.D47A, p.G22R, and p.R23T [25-27] have been proposed. Functional implications of these mutations may account for the phenotypic differences.
Interestingly, Ma et al. [28] have reported an identical substitution, p.W45S, in another connexin polypeptide, connexin 46 (Cx46), in a Chinese family with 10 members over three generations affected with nuclear cataract but without microcornea. This indicates an essential role for tryptophan-45 in Cx46 and Cx50 in maintaining eye lens transparency.
This is also illustrated in connexin knockout and knockin mice. While Cx46 knockout mice develop nuclear cataracts only, Cx50 knockout mice show nuclear cataracts in combination with smaller lenses. This is associated with delayed fiber cell maturation and reduced epithelial cell proliferation. Interestingly, in knockin Cx46 mice, where connexin 46 is expressed under the endogenous Cx50 promoter, the lenses are transparent but smaller, which suggests that connexin 46 cannot substitute for connexin 50 in lens growth [29].
In summary, we describe a novel, heterozygous p.W45S mutation in Cx50 co-segregating completely with congenital cataract and microcornea in a three-generation Indian family. The phenotype observed in this family showed marked differences to the previously reported phenotypes linked with GJA8. As a result, the possibility that variants in other genes involved in maintaining lens transparency, development, and growth thus determining the cataract phenotype cannot be excluded.
Acknowledgments
We wish to thank the patients and their relatives for their cooperation. We are grateful to Professor Digweed for critically reading the manuscript. This work was in part supported by grant number DBT/BT/IC/71/89/Pt from the Department of Biotechnology sanctioned to J.R.S. and INI 331 from Bundesministerium für Bildung und Forschung to K.S., and SR/FT/L-91 from the Department of Science and Technology to V.V.
References
- 1.Rahi JS, Dezateux C, British Congenital Cataract Interest Group. Measuring and interpreting the incidence of congenital ocular anomalies: lessons from a national study of congenital cataract in the UK. Invest Ophthalmol Vis Sci. 2001;42:1444–8. [PubMed] [Google Scholar]
- 2.Wirth MG, Russell-Eggitt IM, Craig JE, Elder JE, Mackey DA. Aetiology of congenital and paediatric cataract in an Australian population. Br J Ophthalmol. 2002;86:782–6. doi: 10.1136/bjo.86.7.782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Vanita ,, Singh JR, Singh D. Genetic and segregation analysis of congenital cataract in the Indian population. Clin Genet. 1999;56:389–93. doi: 10.1034/j.1399-0004.1999.560507.x. [DOI] [PubMed] [Google Scholar]
- 4.Vanita V, Singh JR, Hejtmancik JF, Nürnberg P, Hennies HC, Singh D, Sperling K. A novel fan-shaped cataract-microcornea syndrome caused by a mutation of CRYAA in an Indian family. Mol Vis. 2006;12:518–22. [PubMed] [Google Scholar]
- 5.Willoughby CE, Shafiq A, Ferrini W, Chan LL, Billingsley G, Priston M, Mok C, Chandna A, Kaye S, Heon E. CRYBB1 mutation associated with congenital cataract and microcornea. Mol Vis. 2005;11:587–93. http://www.molvis.org/molvis/v11/a69/ [PubMed] [Google Scholar]
- 6.Devi RR, Vijayalakshmi P. Novel mutations in GJA8 associated with autosomal dominant congenital cataract and microcornea. Mol Vis. 2006;12:190–5. [PubMed] [Google Scholar]
- 7.Hansen L, Eiberg H, Rosenberg T. Novel MAF mutation in a family with congenital cataract-microcornea syndrome. Mol Vis. 2007;13:2019–22. [PubMed] [Google Scholar]
- 8.Kumar NM, Gilula NB. The gap junction communication channel. Cell. 1996;84:381–8. doi: 10.1016/s0092-8674(00)81282-9. [DOI] [PubMed] [Google Scholar]
- 9.Bruzzone R, White TW, Paul DL. Connections with connexins: the molecular basis of direct intercellular signaling. Eur J Biochem. 1996;238:1–27. doi: 10.1111/j.1432-1033.1996.0001q.x. [DOI] [PubMed] [Google Scholar]
- 10.Evans WH, Martin PEM. Gap junctions: structure and function. Mol Membr Biol. 2002;19:121–36. doi: 10.1080/09687680210139839. review. [DOI] [PubMed] [Google Scholar]
- 11.Willecke K, Eiberger J, Degen J, Eckardt D, Romualdi A, Guldenagel M, Deutsch U, Sohl G. Structural and functional diversity of connexin genes in the mouse and human genome. Biol Chem. 2002;383:725–37. doi: 10.1515/BC.2002.076. [DOI] [PubMed] [Google Scholar]
- 12.Goodenough DA. The crystalline lens. A system networked by gap junctional intercellular communication. Semin Cell Biol. 1992;3:49–58. doi: 10.1016/s1043-4682(10)80007-8. [DOI] [PubMed] [Google Scholar]
- 13.Musil LS, Beyer EC, Goodenough DA. Expression of the gap junction protein connexin 43 in embryonic chick lens: molecular cloning, ultrastructural localization, and post-translational phosphorylation. J Membr Biol. 1990;116:163–75. doi: 10.1007/BF01868674. [DOI] [PubMed] [Google Scholar]
- 14.Paul DL, Ebihara L, Takemoto LJ, Swenson KI, Goodenough DA. Connexin 46, a novel lens gap junction protein, induces voltage-gated currents in nonjunctional plasma membrane of Xenopus oocytes. J Cell Biol. 1991;115:1077–89. doi: 10.1083/jcb.115.4.1077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.White TW, Bruzzone R, Goodenough DA, Paul DL. Mouse Cx50, a functional member of the connexin family of gap junction proteins, is the lens fiber protein MP70. Mol Biol Cell. 1992;3:711–20. doi: 10.1091/mbc.3.7.711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Simon AM, Goodenough DA. Diverse functions of vertebrate gap junctions. Trends Cell Biol. 1998;8:477–83. doi: 10.1016/s0962-8924(98)01372-5. [DOI] [PubMed] [Google Scholar]
- 17.White TW. Unique and redundant connexin contributions to lens development. Science. 2002;295:319–20. doi: 10.1126/science.1067582. [DOI] [PubMed] [Google Scholar]
- 18.Vanita V, Singh D, Robinson PN, Sperling K, Singh JR. A novel mutation in the DNA-binding domain of MAF at 16q23.1 associated with autosomal dominant “cerulean cataract” in an Indian family. Am J Med Genet A. 2006;140:558–66. doi: 10.1002/ajmg.a.31126. [DOI] [PubMed] [Google Scholar]
- 19.Mackay D, Ionides A, Kibar Z, Rouleau G, Berry V, Moore A, Shiels A, Bhattarcharya S. Connexin 46 mutations in autosomal dominant congenital cataract. Am J Hum Genet. 1999;64:1357–64. doi: 10.1086/302383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Shiels A, Mackay D, Ionides A, Berry V, Moore A, Bhattacharya S. A missense mutation in the human connexin50 gene (GJA8) underlies autosomal dominant “zonular pulverulent” cataract, on chromosome 1q. Am J Hum Genet. 1998;62:526–32. doi: 10.1086/301762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Pepose JS, Ubels JL. The cornea. In: Hart WM, editor. Adler’s physiology of the eye. Clinical application. 9th ed. St. Louis: Mosby Year Book; 1992. p. 29–70. [Google Scholar]
- 22.Ponnam SPG, Ramesha K, Tejwani S, Ramamurthy B, Kannabiran C. Mutation of the gap junction protein alpha 8 (GJA8) gene causes autosomal recessive cataract. J Med Genet. 2007;44:e85. doi: 10.1136/jmg.2007.050138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Pal JD, Berthoud VM, Beyer EC, Mackay D, Shiels A, Ebihara L. Molecular mechanism underlying a Cx50-linked congenital cataract. Am J Physiol. 1999;276:C1443–6. doi: 10.1152/ajpcell.1999.276.6.C1443. [DOI] [PubMed] [Google Scholar]
- 24.Arora A, Minogue PJ, Liu X, Reddy MA, Ainsworth JR, Bhattacharya SS, Webster AR, Hunt DM, Ebihara L, Moore AT, Beyer EC, Berthoud VM. A novel GJA8 mutation is associated with autosomal dominant lamellar pulverulent cataract: further evidence for gap junction dysfunction in human cataract. J Med Genet. 2006;43:e2. doi: 10.1136/jmg.2005.034108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Xu X, Ebihara L. Characterization of a mouse Cx50 mutation associated with the No2 mouse cataract. Invest Ophthalmol Vis Sci. 1999;40:1844–50. [PubMed] [Google Scholar]
- 26.Chang B, Wang X, Hawes NL, Ojakian R, Davisson MT, Lo WK, Gong XA. Gja8 (Cx50) point mutation causes an alteration of alpha 3 connexin (Cx46) in semi-dominant cataracts of Lop10 mice. Hum Mol Genet. 2002;11:507–13. doi: 10.1093/hmg/11.5.507. [DOI] [PubMed] [Google Scholar]
- 27.Willoughby CE, Arab S, Gandhi R, Zeinali S, Arab S, Luk D, Billingsley G, Munier FL, Heon E. A novel GJA8 mutation in an Iranian family with progressive autosomal dominant congenital nuclear cataract. J Med Genet. 2003;40:e124. doi: 10.1136/jmg.40.11.e124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ma Z, Zheng J, Yang F, Ji J, Li X, Tang X, Yuan X, Zhang X, Sun H. Two novel mutations of connexin genes in Chinese families with autosomal dominant congenital nuclear cataract. Br J Ophthalmol. 2005;89:1535–7. doi: 10.1136/bjo.2005.075184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Gong X, Cheng C, Xia C. Connexins in lens development and cataractogenesis. J Membr Biol. 2007;218:9–12. doi: 10.1007/s00232-007-9033-0. [DOI] [PubMed] [Google Scholar]