

Abstract
Purpose: Interleukin-1 (IL-1) and tumor necrosis factor-alpha (TNF-α) play critical roles in mediating corneal inflammation. In this study, topical blockade of IL-1 and TNF-α, alone or in combination, was compared to conventional corticosteroid anti-inflammatory therapy in suppressing infiltration of the cornea by antigen-presenting Langerhans cells (LCs) and in promoting corneal transplant survival in a mouse model of keratoplasty.
Methods: Study drugs included topical 2% IL-1 receptor antagonist (IL-1Ra), 1.5% soluble TNF-α receptor (sTNFR), and 1% prednisolone phosphate (Pred), all formulated in hyaluronic acid vehicle. Fifty eyes of BALB/c mice were used for LC studies where the numbers of LCs were determined 1 week after electrocautery to the corneal surface or transplantation of C57BL/6 corneas. Additionally, 65 BALB/c mice received corneal allografts and were randomized to receive one of the following for 8 weeks: (1) IL-1Ra, (2) sTNFR, (3) Pred, (4) combined IL-1Ra and Pred, or (5) vehicle alone.
Results: Mean suppression of LC infiltration after electrocautery or transplantation was 67% and 71%, respectively, for IL-1Ra, 40% and 62% for sTNFR, 70% and 72% for sTNFR+IL-1Ra, and 77% and 78% for Pred alone. Rejection rates were 15% for IL-1Ra (P = .01), 38% for sTNFR (P = .1), 17% for Pred (P = .02), and 7% for combined IL-1Ra+Pred (P = .002) as compared to 69% for the vehicle-treated group. IL-1Ra and Pred, but not sTNFR, significantly inhibited post-transplantation neovascularization.
Conclusions: Topical IL-1Ra and prednisolone are comparable in their capacity to promote graft survival. sTNFR therapy, though effective, has much lower efficacy as compared to IL-1Ra or Pred. Combination IL-1Ra and steroid therapy offers only minimal added efficacy over either agent used alone.
INTRODUCTION
Corneal grafting, or penetrating keratoplasty, is the most common form of tissue transplantation; indeed, more corneal transplants are performed each year than all other forms of transplantation combined. In the United States alone, nearly 34,000 cases are performed annually. In uncomplicated first grafts, the 2-year survival rate under cover of local immune suppression, afforded by corticosteroid therapy, is over 85% to 90%.1,2 Although topical corticosteroid therapy is fraught with many side effects, including elevation of intraocular pressure and glaucoma, infection, and stromal thinning, it is still remarkable that topical therapy can lead to such extraordinary rates of success that can be achieved in other solid grafts only with profound systemic immune suppression. This high rate of success has been related to various features of the cornea and ocular microenvironment that together account for its so-called immune-privileged status.3,4 However, many corneal grafts are still rejected, and immune rejection is by far the leading cause of corneal graft failure.1,5 Inflammation in the corneal graft bed with attendant neovascularization is by far the leading tissue characteristic that heralds a high risk of rejection to a transplant.6,7 Unfortunately, neovascularization is a ubiquitous element of corneal pathology that accompanies a vast array of traumatic, inflammatory, infectious, and toxic insults.8 Grafts placed into “high-risk” beds with neovascularization exhibit rejection rates that increase to well over 50% to 90% even with maximal local and systemic immune suppression.6
MECHANISMS OF CORNEAL ALLOREJECTION
Recently, several comprehensive reviews of the immunobiology of corneal transplantation have been published in the literature,1,3,5,9 summarizing the large body of experimental evidence establishing that corneal graft rejection is mediated principally by CD4+ T cells.10–12 Research has shown, however, that activation of alloreactive T cells absolutely requires mobilization of antigen-presenting cells, for without the activity of these cells, the host remains ignorant of the presence of the transplant,13–17 leading to a state of “immunologic ignorance.” The process of corneal transplant immunity can be conceptually and functionally separated into an “afferent” (sensitization) arm and an “efferent” (effector) arm. In this context, the infiltration of the graft by antigen-presenting cells is a critical facet of the sensitization (or afferent) arm of the immune response. Once antigen-presenting cells pick up, process, and present graft (allo) antigens to host T cells, these cells expand into clones of effector cells that can then target the transplant.3,4 The expression or the efferent phase of the response is synonymous with the process of attacking the graft, and here, too, similar to the sensitization phase, local tissue factors can facilitate (or hinder) the process. And in the context of the effector phase, the degree of neovascularization is directly correlated with the efficiency with which T cells can target the transplant.3,7,18,19
Antigen-Presenting Cell Function and Mobilization in Allosensitization
The population of bone marrow—derived antigen-presenting cells that function in the cornea and ocular surface comprises diverse subsets of CD45+ cells with differing ontogeny and cell surface characteristics, including monocytic CD11b+ cells that primarily reside in the stroma, and CD11c+ dendritic cells that reside in the epithelium.20,21 Chief among the antigen-presenting cells of the ocular surface are major histocompatibility complex (MHC) class II+ Langerhans cells (LCs) that reside under normal uninflamed conditions in the limbal area but are capable of readily infiltrating the cornea in response to various inflammatory insults (including transplantation and infection) to initiate T cell—mediated immune responses.1,9,14,22–26 As such, LCs play a critical role in sensitizing the host to the various foreign antigens borne by the graft.27,28
Corneal Neovascularization and Expression of Immunity
Recruitment of inflammatory and immune cells to any tissue site, including the cornea, represents the net functional effect of adhesion molecules and chemokines that function at the level of the vascular endothelium as well as in the tissue matrix.29,30 Recent research has unraveled the many steps in the generation of a cell-mediated immune response in the cornea and has related these to specific signals (eg, chemokines) that provide directional information to leukocytes as well as factors that promote cell adhesion (eg, integrins).31–36 It is critical to emphasize, however, that the leukocytes that mediate immuno-inflammatory responses are derived from the intravascular compartment, and in that context there is no tissue factor that has a more pronounced effect on the facility with which inflammation can express itself than the overall surface area of the blood vessels in contact with the tissue. It is likely for this reason that the preponderance of immune and inflammatory conditions of the cornea present in the corneal periphery and perilimbal area. As such, the growth of blood vessels in the cornea can have a major impact on the immune status of the cornea.7,18,30
MOLECULAR REGULATION OF CORNEAL INFLAMMATION
Cellular responses are induced, regulated, and terminated by molecular mechanisms. Myriad mechanisms exist that regulate virtually every step in the induction and expression of corneal immunity.37 The divergence or distinction in the cellular and molecular pathways that promote inflammation increases the further “downstream” one goes in the immunopathogenic mechanisms at play in a particular pathology. The corollary is that early steps in induction of immunity share common pathways, and in the case of the cornea, as is the case in many tissues, activation of a few critical nuclear transcription factors (eg, NF-κB) is critical for the early steps involved in transplant-mediated rejection.2 The principal molecular mediators of these key signaling pathways are the “master” proinflammatory molecules interleukin-1 (IL-1) and tumor necrosis factor-alpha (TNF-α).29,30,38–42
Role of IL-1 in Corneal Immunity and Inflammation
The critical role of MHC class II+ LC migration into the cornea was defined above. It is now known that this migration/recruitment is regulated principally by the cytokines IL-1 and TNF-α. IL-1 is a potent proinflammatory cytokine produced primarily by monocytes and macrophages, but also by resident corneal cells.43,44 The high expression of IL-1 (along with TNF-α) is one of the early hallmarks of corneal transplant–related inflammation.42 This cytokine has a wide range of activities, including mediation of the acute phase response, chemotaxis and activation of inflammatory cells and antigen-presenting cells, and stimulation of neovascularization.45,46 IL-1 receptor antagonist (IL-1Ra) is a naturally occurring IL-1 isoform, produced by the same cells that synthesize IL-1, that undergoes high-affinity binding to IL-1 receptor I but shows no agonist activity.47–49 It has been shown that intracorneal injection of IL-1 induces centripetal migration of peripheral LCs.24,50 Conversely, neutralizing the activity of IL-1 by topical administration of IL-1Ra has been shown to suppress antigen-presenting cell migration in the cornea51 and prolong the survival of corneal allografts in the mouse.52
However, significant unanswered questions remain in relation to IL-1 modulation in the cornea. First, it has been shown that IL-1 mediation of antigen-presenting cell/LC migration in the cornea is regulated in large part through TNF-α-mediated signaling,24 begging the question of the comparative efficacy of TNF-α vs IL-1 blockade on suppressing corneal inflammation. Second, it is important to determine whether there is any additive or synergistic role to combined IL-1 and TNF-α blockade in suppressing corneal LC infiltration. And finally, the most clinically relevant question is how IL-1 blockade compares to the clinical “gold-standard,” prednisolone 1%, the most potent topical anti-inflammatory.
Role of TNF-α in Corneal Immunity and Inflammation
TNF-α mediates a large number of proinflammatory and immunoregulatory functions, such as up-regulation in the expression of adhesion and costimulatory molecules, activation of neutrophils, induction of chemokine secretion, and activation of the NF-κB signal transduction pathway.45,53 In the cornea, significant expression of TNF-α by the corneal resident cells can be induced by inflammatory stimuli.54 TNF-α activity is regulated by two distinct receptors, the type I receptor (p55; TNFR-I) and the type II receptor (p75; TNFR-II), which have largely homologous extracellular domains but distinct intracellular domains that can mediate discrete cellular responses.55 TNFR-I is believed to be the principal receptor through which many of the proinflammatory activities of TNF-α are mediated.56,57 The bioactivity of TNF-α can be dramatically suppressed by soluble TNFR-I, which binds free ligand and prevents ligation of the membrane-bound receptors.58 Similar to IL-1, administration of TNF-α by intracorneal injection not only induces migration of LCs into the central cornea but also leads to a marked increase in the number of recruited LCs at the corneal limbus, which serves as a potential reservoir for corneal LCs. In gene-targeted knockout mice lacking TNFR-I or TNFR-II, the migratory response of LCs to thermal cautery or cytokine stimulation is profoundly attenuated.24 The molecular mechanisms that mediate TNF-α induced effects on LCs have recently been elucidated. Specifically, it has been suggested that TNF-α-mediated signaling leads to up-regulation of select chemokines (eg, RANTES, MIP-1 β) and chemokine receptors (eg, CCR5) that effect LC recruitment to the cornea.25,59 Conversely, it has been shown that blockade of TNF-α function, through either deletion of the TNFR-I38 or administration of soluble TNFR-I,60 can promote acceptance of minor H- (but not MHC)-incompatible corneal transplants in mice.
However, critical and clinically relevant questions remain in relation to topical TNF-α blockade. Since sTNFR potentially suppresses binding of both TNF receptors, and concomitant deletion of both TNFR genes in “double knockout” animals has been shown to have no discernible effect on combined MHC and minor H-disparate grafts38 (the most common allograft setting faced in the clinical setting), the question remains as to whether topical sTNFR administration can promote survival of fully disparate allografts. Second, given the central role of TNF-α in inflammation and its implicated role in angiogenesis, how does topical TNF blockade compare with corticosteroid therapy? Lastly, given the distinct and yet to some extent overlapping functions of TNF-α and IL-1, how do IL-1- and TNF-α-blocking strategies compare to one another in modulating corneal inflammation?
Given these unanswered questions, the aim in this study was to compare selective topical blockade of IL-1 and TNF-α, alone or in combination, to conventional corticosteroid anti-inflammatory therapy in mouse models of corneal inflammation and fully (MHC and multiple minor H) mismatched corneal transplantation.
METHODS
MICE AND ANESTHESIA
Eight- to 10-week-old BALB/c (H-2d) and C57BL/6 (H-2b) mice were purchased (Taconic, Germantown, New York). All animals were treated according to the Association for Research in Vision and Ophthalmology Statement for the Use of Animals in Ophthalmic and Vision Research, and all protocols had received approval from the Institutional Animal Use and Care Committee. Each animal was deeply anesthetized with an intramuscular injection of 3 to 4 mg ketamine and 0.1 mg xylazine prior to all surgical procedures.
PHARMACOLOGIC STRATEGY
Topical preparations were applied to murine eyes starting on the day of the procedure and 3 times daily thereafter in a masked fashion until day of enucleation. The study medications were prepared as follows: 20 mg/mL of recombinant IL-1 receptor antagonist (IL-1Ra; Amgen Inc, Thousand Oaks, California) in sodium hyaluronic acid (HA) in phosphate-buffered saline (PBS); 15 mg/mL of soluble TNF-α receptor I (sTNFR; Amgen) in HA; prednisolone phosphate 1% prepared in HA vehicle. Vehicle-treated animals received 0.2% sodium hyaluronate only. For optimal stability of the sTNFR, a high-molecular-weight (30 kDa) polyethylene glycol (PEG) molecule was attached to the sTNFR; the product, PEG sTNFRI, was then constituted at 15 mg/mL to optimize its stable formulation in solution.
THERMAL CAUTERY OF THE CORNEAL SURFACE
Mice were anesthetized and placed under the operating microscope. Using the tip of a hand-held cautery device, 5 burns were applied to the central 50% of the cornea to induce centripetal LC migration.61 Twenty-five mice were randomly divided into 5 groups (N = 5 each) receiving the following formulations topically 3 times daily: IL-1Ra, sTNFR, Pred, combination of IL-1Ra and sTNFR, and vehicle alone. All treatments were started immediately following surgery and applied throughout the study period. At 1 week after cauterization, which correlates with the significant LC migration response in this model,51 corneas were harvested and LC enumeration was performed as detailed below.
ORTHOTOPIC CORNEAL TRANSPLANTATION
The mouse model of corneal transplantation has been described previously.52,62,63 Briefly, each BALB/c recipient (N = 90) was deeply anesthetized with an intraperitoneal injection of ketamine and xylazine, as described above, prior to all surgical procedures. The central 2 mm of the donor (C57BL/6) allogeneic cornea was excised with curved Vannas microsurgical scissors after it was briefly indented and partially trephined. The donor button was then secured in the recipient graft bed with 8 interrupted 11-0 nylon sutures (Sharpoint; Vanguard, Houston, Texas). Antibiotic ointment was applied to the corneal surface and the eyelids were closed for 24 hours with an 8-0 nylon tarsorrhaphy. All grafted eyes were examined after 72 hours; no grafts were excluded from analysis because of technical difficulties. Transplant sutures were removed in all cases on day 5. Twenty-five mice were randomly divided into 5 groups (N = 5 each) receiving 5 μL of the following formulations topically 3 times daily: IL-1Ra, sTNFR, Pred, combination of IL-1Ra and sTNFR, and vehicle alone, and were prospectively followed for 1 week, at which point they were sacrificed and the eyes enucleated (well before any sign of rejection) for LC enumeration as detailed below. The remaining 65 mice were randomized into the following groups: IL-1Ra (N=13), sTNFR (N=13), Pred (N=12), combination of IL-1Ra and Pred (N=14), and vehicle alone (N=13); they continued to receive treatment 3 times daily with the topical formulations.
LANGERHANS CELLS ENUMERATION
Major histocompatibility complex class II+ (murine Iad+) LCs were enumerated in whole corneal epithelial sheets by use of indirect immunofluorescence assay, as described previously.51,52 Briefly, at 1 week following corneal stimulation (with cautery) or transplantation, murine eyes were collected and the corneas were dissected. Corneas were placed in 20 mM ethylenediaminetetraacetic acid (EDTA) buffer and incubated for 30 to 40 minutes at 37°C, followed by removal of the epithelium in toto, and washed in PBS at room temperature. The cornea was then fixed with 95% ethanol prior to washing and incubation with 1:20 diluted primary anti-murine Iad antibody for 45 minutes at 37°C. The tissue was then washed in PBS and incubated with a FITC-labeled goat anti-mouse secondary antibody for 30 minutes at 37°C. Negative controls either bypassed this step or were incubated with antibody specific for an unrelated MHC epitope. Sections were then mounted on slides and examined under the fluorescent microscope with a square ocular grid where Langerhans cells were enumerated.
EVALUATION AND SCORING OF ORTHOTOPIC CORNEAL TRANSPLANTS
Grafts were evaluated by slit-lamp biomicroscopy and scored for opacification for 8 weeks; a longer period of follow-up is not needed in this model since the vast majority of the rejection episodes occur in weeks 3 to 5 post-transplantation.16 A previously described and standardized scoring system64-66 was used to measure the degree of opacification in the graft between 0 and 5+: 0 = clear and compact graft; 1+ = minimal superficial opacity; 2+ = mild deep (stromal) opacity with pupil margin and iris vessels visible; 3+ = moderate stromal opacity with only pupil margin visible; 4+ = intense stromal opacity with the anterior chamber visible; and 5+ = maximal corneal opacity with total obscuration of the anterior chamber. Grafts with an opacity score of 2+ or greater after 3 weeks were considered rejected; grafts with an opacity score of 3+ or greater at 2 weeks that never cleared were also regarded as rejected. The growth of neovascularization was graded between 0 and 8 as described previously based on the degree of centripetal ingrowth and quandrantic involvement of the new vessels in the graft bed.52
STATISTICAL ANALYSIS
Comparison of the mean numbers of infiltrating LCs and mean levels of post-transplant neovascularization between the different treatment protocols was made using the Student t test. Kaplan-Meier survival curves were constructed for all treatment modalities; the log-rank test for equality of survival was then performed to compare the rates of graft survival in the different groups over the entire follow-up period. All P values of < .05 were deemed significant.
RESULTS
LC INFILTRATION IN RESPONSE TO CAUTERY
To measure the mobilization of MHC class II+ LCs into the corneal tissue after a non-antigen-specific acute stimulation, corneal cautery was applied to the corneal surface—a procedure with a highly predictable migration of limbal LCs into the central areas of the cornea as previously described.24,51,61 The critical contribution of IL-151 and TNF-α24 to this process had been established but neither (i) the relative efficacy of topical IL-1 vs TNF-α blockade in suppressing this response, nor (ii) the relative efficacy of these strategies, alone or in combination, as compared to topical steroid, had been established. Accordingly, 25 mice were randomized into 5 groups (N = 5 each) receiving the following formulations: IL-1Ra, sTNFR, Pred, combination of IL-1Ra and sTNFR, and vehicle alone; treatments were started immediately following surgery and applied throughout the study period. At 1 week, corneas were harvested and LC enumeration was performed as detailed above. Results from this study are summarized in Figure 1. Application of IL-1Ra, sTNFR, combination of sTNFR and IL-1Ra, and Pred alone led to 67% (P = .01), 40% (P = .04), 70% (P = .01), and 77% (P = .001) decrease in LC migration as compared to vehicle treatment alone. Prednisolone alone had a significantly greater efficacy than sTNFR (P = .03), but not greater than IL-1Ra (P = .4). There was no additive effect to suppression of LC migration by addition of sTNFR to IL-1Ra.
FIGURE 1.
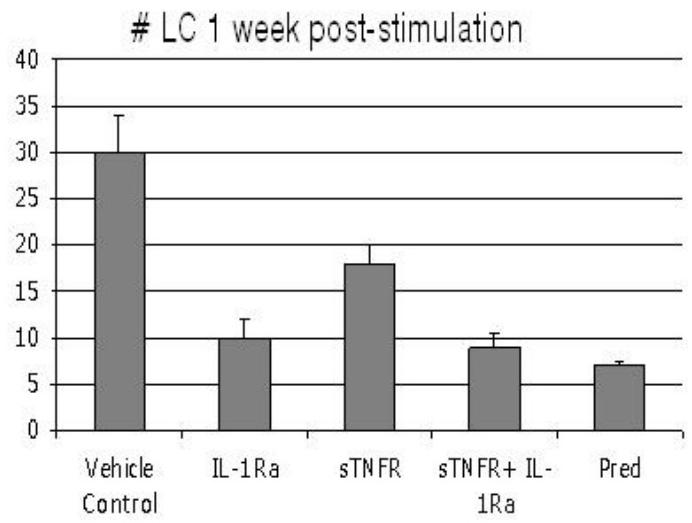
Number of major histocompatibility complex class II+ Langerhans cells (LCs) in the cornea 1 week following cauterization, based on treatment modality. Topical application of IL-1Ra (P = .01), sTNFR (P = .04), sTNFR + IL-1Ra (P = .01), and Pred (P = .001) leads to significant reduction in number of infiltrating LCs as compared to vehicle-treated controls. Topical Pred alone leads to a suppression level significantly greater than that observed for sTNFR (P = .03) but not compared to IL-1Ra (P = .4). Bars represent standard error of the mean.
LC INFILTRATION IN RESPONSE TO CORNEAL TRANSPLANTATION
Infiltration of host LCs into the graft plays a critical role in picking up graft-derived antigens, and hence in allosensitization.13-15 Given the importance of this process to the immunopathogenesis of graft rejection, the 4 treatment modalities (IL-1Ra, sTNFR, sTNFR + IL-1Ra, and Pred) were compared to one another and to vehicle-treated controls in suppressing migration of host MHC class II+ (Iad+) LCs into the allograft 1 week after transplantation. This early time period was selected because it is well before allorejection (which typically occurs after 3 weeks), which itself can affect migration of leukocytes into the graft tissue due to the resultant inflammation. By focusing at the early time period, it is possible to focus on factors that affect only the sensitization arm of alloimmunity. Accordingly, 25 BALB/c recipients of C57BL/6 allografts were randomized into 5 groups (N = 5 each); 4 groups received the specified formulations and 1 group served as a control treated with HA vehicle alone (Figure 2). Mean suppression of LC infiltration was 71% for IL-1Ra, 62% for sTNFR, 72% for sTNFR+IL-1Ra, and 78% for Pred alone, all representing significant reductions in host LC infiltration into the graft as compared to the vehicle-treated controls. No significant difference was observed between the treatment modalities. The summary of the percent mean decreases in LC infiltration in the non-antigen-specific cautery and transplant (antigen)-specific LC infiltration is shown in Figure 3. In both cases, sTNFR provided no additional effects than those observed with IL1-Ra alone.
FIGURE 2.
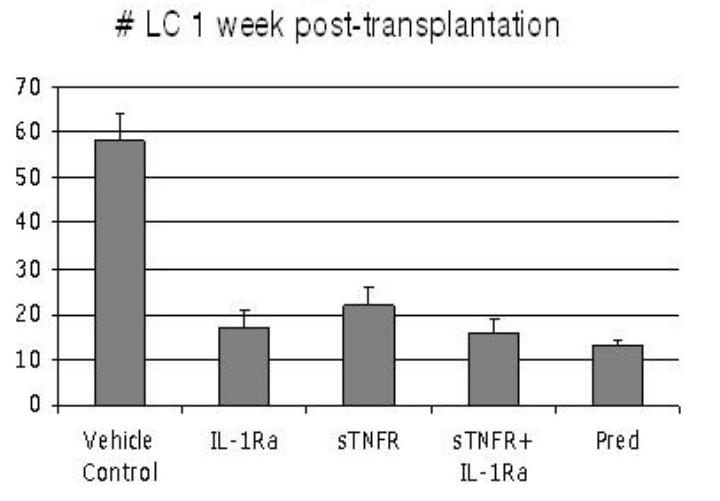
Number of limbic host–derived major histocompatibility complex class II+ Langerhans cells (LCs) in the grafted cornea 1 week after transplantation, based on treatment modality. Topical application of IL-1Ra (P = .01), sTNFR (P = .02), sTNFR + IL-1Ra (P = .01), and Pred (P = .01) leads to significant reduction in number of infiltrating LCs as compared to vehicle treatment alone. Bars represent standard error of the mean.
FIGURE 3.
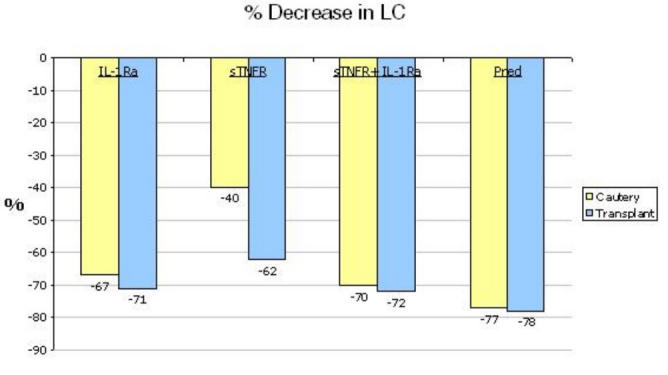
Mean percent decrease in Langerhans cell (LC) recruitment to cornea in two models of corneal inflammation (cauterization and transplantation) based on treatment modality. In the aggregate, IL-1Ra and steroid therapy have comparable efficacy, which is greater than that observed for sTNFR.
TRANSPLANT OPACIFICATION AND REJECTION
The grading scheme for opacification of murine corneal grafts used in this paper has been used widely by different groups after standardization and validation,52,62,63,65,67,68 including correlation with transplant cellular infiltration (which requires enucleation and sacrifice of hosts). This measure, procured biomicroscopically, can therefore provide significant information about the degree of graft inflammation. Typically, a transient degree of graft opacity is seen in the early postoperative period and clears in the following weeks unless rejection sets in. Based on the data above, suggesting the higher efficacy of IL-1Ra and Pred in suppressing ingress of host immune cells into the graft, in this set of experiments 65 mice were randomized into 5 groups to additionally test whether there is any additive effect of IL-1Ra and corticosteroid therapy in the transplant setting. These groups were: IL-1Ra (N = 13), sTNFR (N = 13), Pred (N = 12), combination of IL-1Ra and Pred (N = 14), and vehicle alone (N = 13). Figure 4 summarizes the graft opacity data, by treatment group, for the first several weeks after transplantation. Opacity scores were all within the same range by week 1, except for the combined treatment of Pred and IL-1Ra, which demonstrated the least acute postoperative change as compared to the other groups (P = .04). By the fourth postoperative week, all treatment groups demonstrated comparable and significant or near-signficant (IL-1Ra, P = .02; sTNFR, P = .08; Pred, P = .001; combined IL-1Ra and Pred, P = .05) suppression in opacity scores as compared to the vehicle-treated controls.
FIGURE 4.
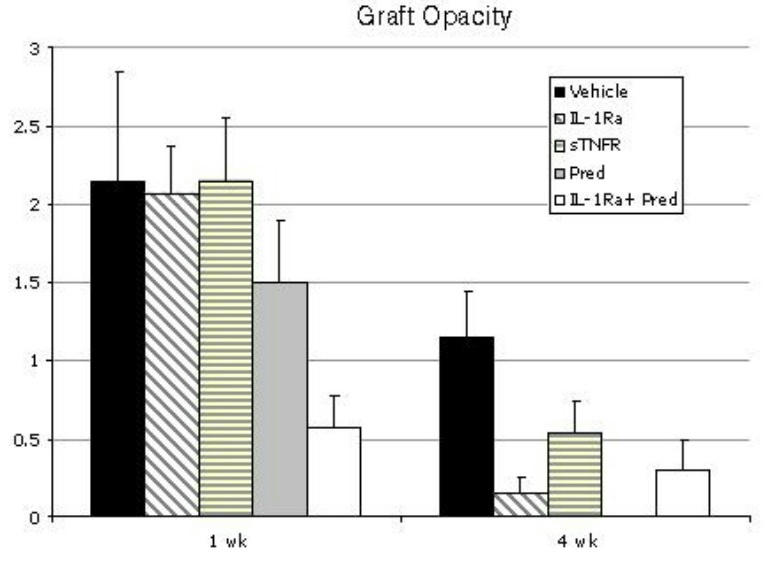
Mean corneal transplant opacity scores among 5 different treatment groups at 1 and 4 weeks postkeratoplasty. At 1 week, maximal suppression in graft opacity is observed with combined IL-1Ra and Pred therapy. By 4 weeks, all treatment regimens demonstrate significantly lower opacity scores as compared to the vehicle-treated group. Bars represent standard error of the mean.
Graft opacity levels, once beyond a threshold level of 2+, as defined in the “Methods” section, represent transplant rejection.68 Figure 5 summarizes rejection rates for the 4 experimental and one vehicle-treated groups at 4 and 8 weeks after transplantation—the latter period representing the end of the follow-up period, since graft rejection beyond this period is rare in the mouse.16 Figure 6 demonstrates the precise levels of opacity for all grafts for all treatment regimens over time. At 4 weeks, 38% of vehicle-treated grafts had succumbed to rejection as compared to 23% among the sTNFR-treated group (P = .1). Treatment with IL-1Ra, Pred, or a combination of IL-1Ra and Pred led to a significant decrease in rejection rates to 8%, 0%, and 8%, respectively. Similarly, by the end of the follow-up period at 8 weeks, close to two-thirds (69%) of the vehicle-treated eyes experienced graft rejection, whereas treatment with sTNFR decreased this rate by 45% to a total of only 38% overall. Treatment with IL-1Ra or Pred decreased the rate seen in sTNFR-treated eyes by over 50%, leading to rejection rates of 15% and 17%, respectively, and the combination of IL-1Ra and Pred demonstrated the lowest rejection rate; only 1 of 14 grafts (7%) was rejected in this group. The most robust method to evaluate transplant survival is by Kaplan-Meier analysis (Figure 7), which takes into account all time points for the different treatment regimens. This analysis revealed that, statistically, the most significant reduction in rejection rates was seen with IL-1Ra (P = .01) or Pred (P = .02) therapy; combined IL-1Ra and Pred therapy led to a profound increase in survival (P = .002). sTNFR therapy, though associated with a measurable and nearly one-half reduction in rejection rates, was not associated with a statistically significant suppression (P = .1), likely due to sample size.
FIGURE 5.
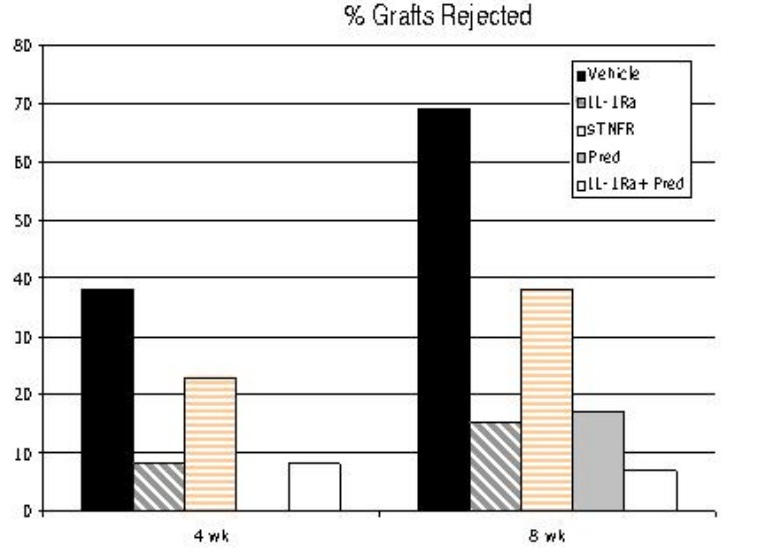
Rejection rates at 4 and 8 weeks of follow-up for corneal allografts, based on treatment regimen. By completion of the 8-week follow-up period, maximal (nearly 90%) reduction in rejection is seen with combined IL-1Ra and Pred therapy (7%) as compared to administration of vehicle alone (69%).
FIGURE 6.

Opacity scores for all grafts over 8-week follow-up period: Top left, Vehicle treatment (N = 13); Top middle, sTNFR treatment (N = 13); Top right, IL-1Ra treatment (N = 13); Bottom left, Pred treatment (N = 12); and Bottom right, Pred and IL-1Ra (N = 14
FIGURE 7.

Kaplan-Meier survival curves for all treatment regimens as compared to vehicle treatment alone. Treatment with IL-1Ra (P = .01), combined IL-1ra and Pred (P = .002), and Pred (P = .02) demonstrate higher efficacy in suppressing graft rejection as compared to sTNFR (P = .1) treatment.
POST-TRANSPLANTATION NEOVASCULARIZATION
Development of corneal neovascularization is both a consequence and cause of pronounced corneal inflammation and may have significant implications for the survival of corneal allografts.6,7,18,63,69 Therefore, corneal neovascularization was measured for the duration of the follow-up period for all treatment groups. Figure 8 demonstrates mean neovascularization scores at the early, mid, and late phases (weeks 1, 4, and 8, respectively) of the study based on treatment regimen. At week 1, combined treatment with IL-1Ra and Pred was associated with minimal neovascularization, with IL-1Ra and sTNFR having comparable effects. In contrast, by 8 weeks IL-1Ra was associated with less neovascularization as compared to vehicle alone or sTNFR (P = .1). Maximal angiostatic effect, however, was observed with Pred and with combined IL-1Ra and Pred. Since outliers can shift the mean appreciably, and yet minimal neovascularization responses may have limited biological/clinical consequences for the transplant, an analysis was done of neovascularization responses >2—that is, ingrowth of blood vessels beyond the graft-host interface (Figure 9). The data reveal that in the aggregate, while in the early postoperative period all treatments show similar angiostatic efficacy in preventing robust neovascularization responses that can threaten the graft, by 8 weeks maximal suppression of neovascularization was seen only with IL-1Ra and/or Pred therapy. Figure 10 summarizes the data in terms of percent reduction in post-transplantation neovascularization over the 8-week period based on the treatment regimen. IL-1Ra alone (38%), Pred alone (63%), and combined IL-1Ra and Pred (59%) demonstrated far higher efficacy in neovascularization suppression than the modest (16%) suppressive effect of sTNFR. Figure 11 provides the neovascularization scores for all the grafts over the 8-week follow-up period, based on treatment regimen and rejection status, demonstrating a strong correlation between neovascularization growth and rejection. The significant majority (eg, 100% in vehicle-treated, 50% in IL-1Ra-treated, and 83% for sTNFR-treated groups) of grafts with a neovascularization score of >2 were rejected by the end of the follow-up period.
FIGURE 8.
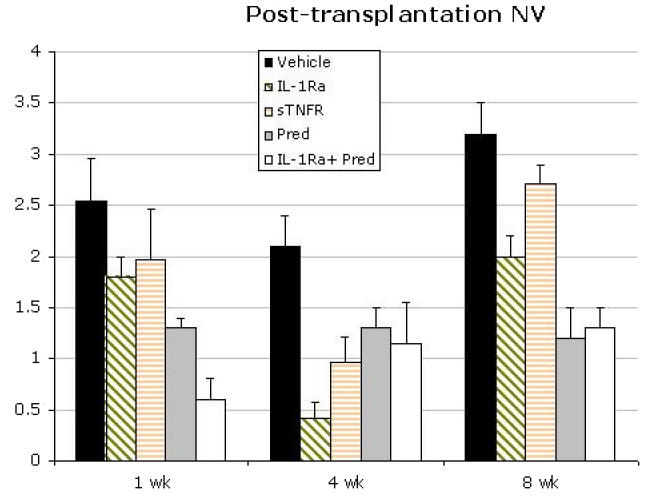
Mean scores for posttransplantation neovascularization (NV) at weeks 1, 4, and 8 based on treatment regimen. Overall, maximal angiostatic effect is seen with Pred alone and combined IL-1Ra and Pred therapy. Bars represent standard error of the mean.
FIGURE 9.
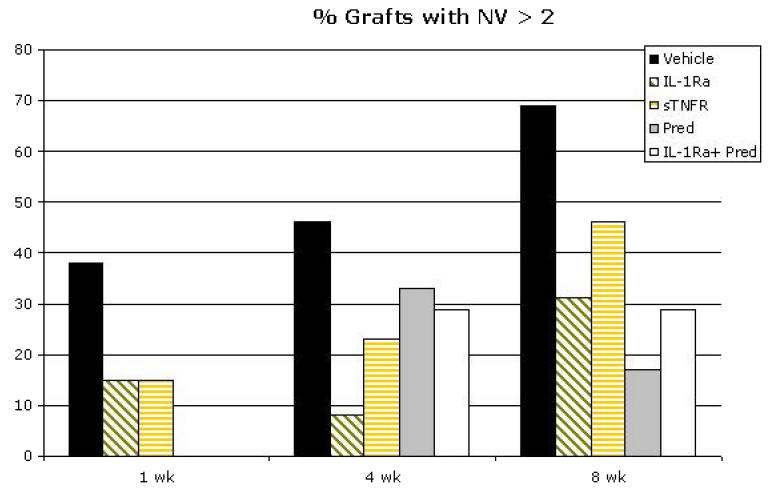
Percent of corneal transplants with neovascularization (NV) score >2 at 1, 4, and 8 weeks postgrafting based on treatment regimen. Overall, although in the early postoperative period all treatments show angiostatic efficacy, by 8 weeks maximal suppression of NV as compared to the vehicle-treated eyes is seen with IL-1Ra and/or Pred therapy.
FIGURE 10.

Percent reduction in posttransplantation neovascularization (NV) over 8 weeks based on treatment regimen. IL-1Ra alone (38%), Pred alone (63%), and combined IL-1Ra and Pred (59%) demonstrate higher efficacy in NV suppression, whereas the suppressive effect (16%) of sTNFR is very modest.
FIGURE 11.

Neovascularization (NV) scores for all grafts over 8-week follow-up period, based on treatment regimen and rejection status: Top left, Vehicle treatment (N = 13); Top middle, sTNFR treatment (N = 13); Top right, IL-1Ra treatment (N = 13); Bottom left, Pred treatment (N = 12); and Bottom right, Pred and IL-1Ra (N = 14).
DISCUSSION
Corneal blindness from inflammatory conditions, such as from infection (eg, trachoma, corneal ulceration) or from trauma or vitamin A deficiency (xerophthalmia), is extremely common, representing one of the most common causes of visual impairment worldwide, affecting several million persons. Corneal transplantation often represents the only, and last, recourse available for restoring sight for the many people blind from corneal opacification.1,3 Indeed, today, keratoplasty represents the most common form of solid tissue transplantation in the United States. However, the generally good graft outcomes have tended to overshadow the significant numbers of graft recipients whose transplants reject. In addition to the significant personal and economic costs associated with transplant rejection, host immune reactions frequently lead to more rejection episodes against the same or a future graft.6,9 The cumulative problem of corneal transplant rejection is reflected by the fact that regrafting is increasingly becoming a leading indication for corneal transplantation in large eye centers. This problem is felt nowhere more acutely than in high-risk keratoplasty, where the diseased corneal bed receiving the graft is further compromised by inflammation and corneal neovascularization that can abrogate the eye’s normal immune-privileged state and lead to fulminate graft rejections.6,7,69,70
The remarkable fact remains, however, that more than 60 years after the widespread adoption of corneal transplantation in the United States, a period that has witnessed very significant advances in eye banking and microsurgical techniques, little has changed in the medical management of this procedure. The currently available pharmaceutical armamentarium for corneal transplant survival is principally composed of corticosteroids. The introduction of these agents into the field of ophthalmology remains arguably the single most significant factor in the advances in corneal transplant surgery over the last 4 decades. Nevertheless, beyond their well-known serious complications,71 corticosteroids show widely variable efficacy in preventing ultimate immunogenic graft failure, and this is particularly the case in high-risk keratoplasty.1,6,69 The current studies were conducted to directly compare the efficacy of a potent corticosteroid, prednisolone phosphate 1%, with topical antagonists to the proinflammatory cytokines IL-1 and TNF-α that have shown efficacy in suppressing these cytokines’ activities on the ocular surface. Specifically, as described in the “Introduction,” whereas suppression of these cytokines (either by gene deletion or topical blockade) has been shown to be effective in various facets of corneal inflammation,40,52 their relative efficacy in relation to one another, or to the clinical standard of corticosteroid therapy, was yet to be demonstrated. Moreover, particularly in the case of TNF antagonism, the results of TNFR-I/p55 vs TNFR-II/p75 receptor blockade in relation to promoting graft survival have been highly variable,38 leaving open the question in relation to the efficacy of concomitant blockade of both receptors (eg, by a soluble receptor, as in sTNFR) in fully mismatched allografts that are disparate at multiple MHC and minor H loci. The experiments presented herein were conducted to address these questions.
The end points in this study, LC migration into the cornea, post-keratoplasty neovascularization, and loss of graft clarity (opacification), were chosen since they are cardinal factors that ultimately determine the fate of a transplant. Langerhans cells are the principal antigen-presenting cells of the cornea and ocular surface,13–15 whose principal activity resides in their serving as the “sentinels” of the immune system. As such, their mobilization as immunocompetent MHC class II+ antigen-presenting cells from the limbus, where they normally reside, into the cornea represents a fundamentally critical aspect in the induction of corneal immunity. Conversely, suppression of their migration in and out of the cornea can be a potent tool for immune modulation and promotion of immune quiescence.9,17,22 In the first set of experiments, a non-antigen-specific inflammatory stimulus (cauterization) was applied to the ocular surface and the ingress of LCs was quantified under different experimental conditions in which IL-1 alone, TNF-α alone, or combined IL-1 and TNF-α blockade were compared to topical corticosteroid therapy in their ability to arrest migration of class II+ LCs into the cornea. The data (Figure 1) demonstrated that, overall, prednisolone and IL-1Ra were most effective in suppressing LC mobilization, with the efficacy of sTNFR being nearly half that of prednisolone alone. Interestingly, there was no additive effect to suppression of LC migration by addition of sTNFR to IL-1Ra.
In the second set of experiments, the infiltration of host-derived limbic LCs into the allograft, before any demonstrable allorejection, was measured under different experimental conditions. Since the migration of these cells into the graft plays a critical role in picking up transplant-derived antigens,13–15 it was hypothesized that interventions that retard or suppress the ingress of these cells into the transplant can attenuate the alloimmune response. Consequently, to compare the efficacy of IL-1 vs TNF-α blockade to topical steroid therapy, either alone or in combination, the 4 treatment modalities (IL-1Ra, sTNFR, sTNFR + IL-1Ra, and Pred) were compared to one another and to vehicle-treated controls in suppressing migration of BALB/c host-derived (Iad+) LCs into the C57BL/6 allografts 1 week after transplantation; the differing immunogenetics of the 2 strains allowed for distinguishing the source of the cells. All treatment modalities were highly effective in suppressing LC migration (Figure 2), with a trend toward higher efficacy with IL-1 blockade or steroid therapy as compared to TNF-α blockade; however, this was not significant. Similar to the nontransplant stimulus (cautery) model, sTNFR administration provided no additive effect over that observed with IL-1Ra alone.
Dekaris and coworkers24 demonstrated that IL-1-mediated recruitment of LCs into the cornea is largely mediated by signaling through the TNFR-I/p55 receptor. Additionally, Yamagami and coworkers25 have recently demonstrated that the chemokine signaling for CCR5-mediated recruitment of corneal LCs is regulated by TNF-α. Accordingly, one would have predicted that TNF-α blockade would be at least as effective as IL-1 blockade in suppressing mobilization of LCs, whereas data presented herein, in the aggregate (Figure 3) do not support this assertion. It is not possible, based on the experimental data presented here, to definitively address why IL-1 blockade was at least as effective as, if not more effective than, TNF-α blockade in suppressing LC mobilization. However, several possibilities exist. First, it is important to recall that the main source for the bone marrow–derived leukocytes that infiltrate the cornea and ocular surface is the intravascular compartment.37 In this context, factors that regulate cell egress from the limbal intravascular compartment into the tissue matrix (eg, integrins, selectins, vascular permeability factors) can have a profound influence on inflammatory and immune cell mobilization into the cornea, independent of chemotactic signaling alone,30 which appears to be largely under the direction of TNF-α. For example, Zhu and Dana29 demonstrated that overexpression of ICAM-1, an important cell adhesion factor and member of the immunoglobulin superfamily of gene products, by the limbal vascular endothelial cells is critical for early infiltration of the cornea by bone marrow–derived cells, and that ICAM-1 expression is principally under the regulation of IL-1. Similarly, overexpression of matrix metalloproteinase (MMP) is required for matrix remodeling, a critical step in cell proliferation and migration. In this regard, too, it is known that MMP, including collagenase, gene expression in many tissues,46 including the cornea,72 is under the regulation of IL-1. Hence, it is certainly plausible that IL-1 antagonism is at least as effective as TNF-α inhibition on regulating LC infiltration into the cornea.
The conclusion drawn from the LC data is that IL-1Ra and Pred are most effective in suppressing ingress of cells into the cornea and that there is no measurable added efficacy by combining sTNFR and IL-1Ra therapy (Figure 3). Based on this, the following formulations were tested in an orthotopic model of corneal transplantation: IL-1Ra, sTNFR, Pred, combination of IL-1Ra and Pred, and vehicle alone, allowing to test not only the relative efficacy of sTNFR and IL-1Ra in a validated in vivo model, but to additionally test whether there is any additive effect of IL-1Ra and corticosteroid therapy in suppressing transplantation-related allorejection and neovascularization. The data are presented in detail in Figures 5 through 7 but may be essentially summarized as follows: Treatment with sTNFR cut the rejection rates seen in vehicle-treated eyes by nearly one-half. However, treatment with IL-1Ra or Pred cut the rejection rates seen with sTNFR by another one-half, meaning that IL-1Ra or Pred can reduce overall rejection rates of control eyes by nearly 75%, with no significant difference between these 2 agents in efficacy (P = .9). Remarkably, the combination of IL-1Ra and Pred demonstrated the lowest rejection rate, only 7%, although this was not statistically different from IL-1Ra alone (P = .53) or Pred alone (P = .49).
As outlined in the “Introduction,” the afferent (sensitization) and efferent (effector) arms of the alloimmune system are largely defined by the robustness by which T cells are sensitized to graft antigens by antigen-presenting cells, and the facility by which these effector cells can target the graft tissue, respectively. And a critical tissue parameter that determines the facility of cellular trafficking to an avascular inflamed tissue (cornea) is neovascularization, since the endothelialized channels, in a milieu that is normally devoid of vessels, serve as efficient conduits for host immune effector elements. Furthermore, the vascular endothelial cells can play a critical role in the recruitment and activation of cellular effectors by up-regulating adhesion and costimulatory molecules.30,73 However, neovascularization is a ubiquitous element of corneal pathology that may accompany a vast array of infectious, inflammatory, traumatic, and toxic insults to the cornea,7,74,75 and for this reason the cornea is at risk of immune-mediated destruction in many of these conditions. Unfortunately, it has been shown in both human18 and mouse68,70 settings that corneal transplantation itself can, as a result of surgical trauma and inflammation, induce neovascularization. Consequently, it is a central tenet of corneal transplant immunobiology and ophthalmologic practice that strategies that can reduce the neovascularization response (angiostasis) can also improve transplant survival.69 The data presented here provide a direct comparison in the efficacy of selective IL-1 vs TNF-α antagonism and standard angiostatic therapy with Pred in countering post-transplant neovascularization. As summarized in Figure 10, maximal angiostatic effect was seen with Pred or combined Pred and IL-1Ra therapy. Efficacy of IL-1Ra alone measured, overall, two-thirds that seen with Pred therapy, but considerably more than that seen with sTNFR administration at most of the time points measured (Figures 8 and 9). The close relationship between the degree of neovascularization and alloreactivity is reflected in the fact that a majority of grafts with neovascularization scores >2 were rejected (Figure 11).
The most proximal molecular mediator of angiogenesis is vascular endothelial growth factor (VEGF).9,19,76 Since VEGF also promotes significant vascular leakage, it is implicated as an important vascular factor in inflammatory conditions.76 Indeed, neutralization of VEGF has been shown to prolong corneal allograft survival, associated with suppressed cellular infiltration, in both the rat77 and mouse19 models of keratoplasty. These observations are germane to commonly seen corneal inflammatory disorders, since development of corneal neovascularization is both a consequence and a cause of pronounced corneal inflammation.6,7,63,69 At the molecular level, the close link between angiogenesis and inflammation is explained by the fact that VEGF receptors (eg, VEGFR-II/kdr) and ligands (eg, VEGF, VEGF-C) are up-regulated by proinflammatory cytokines, principally IL-1, and to a lesser extent by TNF-α.78 This would explain why topical application of IL-1Ra and sTNFR, as has been demonstrated here, could suppress the angiogenic response in the cornea. These data are in accord with observations made by Biswas and coworkers,79 in which IL-1Ra was shown to suppress VEGF levels in a mouse model of herpetic stromal keratitis, leading to pronounced suppression of viral keratitis–associated neovascularization. The appreciably higher angiostatic effect of IL-1Ra as compared to sTNFR in the study presented herein is also in accord with observations made by Coxon and coworkers80 in which IL-1Ra, but not sTNFR, significantly suppressed basic fibroblast growth factor- and VEGF-induced corneal neovascularization. The precise molecular mechanisms that would explain the apparent differential effect of IL-1 vs TNF-α antagonism on angiogenesis are not fully understood. First, as indicated above, the preponderance of evidence suggests a tight link between IL-1 and VEGF/VEGFR expression. Second, many of the integrins implicated in angiogenesis are in turn regulated by IL-1.81 Third, it should be recalled that TNF-α is a highly pleiotropic cytokine, whose function is in part regulation of inflammation through mediating cell death. Specifically, both TNF-α and other members of the TNF superfamily of gene products, including TNF-related apoptosis-inducing ligand (TRAIL), can induce vascular endothelial cell death; hence, blockade of their function may in part inhibit the normal mechanisms by which vascular growth is controlled.82 In contrast, virtually all known functions of IL-1 are limited to promoting inflammation and vascular sprouting. Indeed, it is plausible that many of the anti-inflammatory effects of corticosteroids are due to inhibition of IL-1 and downstream signal transduction pathways, such as NF-κB, stimulated by IL-1.46
Current immunosuppressive drugs used to prevent or treat corneal graft rejection in humans include corticosteroids and occasionally systemic immunosuppressive agents. However, corticosteroids are only variably effective in the prevention or treatment of corneal graft rejection, and their long-term use, as is often the norm in postkeratoplasty cases, is often complicated by sustained increased intraocular pressure, glaucoma, and cataracts, and more infrequently by opportunistic infections.69,71 Although several investigators have been strong proponents of systemic immunosuppressive use in corneal transplantation,83,84 their efficacy and optimal dosing are far from clear, owing to a virtually total lack of randomized masked prospective trials. Moreover, systemic immunosuppressives are fraught with myriad risks as they induce nonspecific suppression of both acquired and innate immunity.10,83,84 Development of efficacious molecular strategies that target critical pathways in transplant immunity and rejection can potentially circumvent many of these concerns. The data presented here indicate that topical blockade of IL-1, in an experimental mouse model, shows efficacy levels in suppressing immune cell infiltration, neovascularization, and graft opacification, comparable to that seen with prednisolone phosphate 1%. Much additional work needs to be done to evaluate long-term efficacy and toxicity profiles of IL-1Ra therapy, not only in transplantation but possibly in other immune and inflammatory conditions of the cornea and ocular surface. Moreover, it would be important to know whether cytokine blockade with administration of these therapies starting just prior to surgery can even provide greater efficacy than initiation of therapy on the day of surgery as performed in these studies. Finally, from a preclinical development standpoint, it would be important to compare topical IL-1Ra administration with other IL-1 antagonism strategies, such as inhibitory RNA, anti-sense, and antibody-based technologies, in their capacity to inhibit IL-1-mediated functions.
ACKNOWLEDGMENTS
Funding/Support: This study was supported in part by grants RO1 EY-12963 and R21 EY-015738 from the National Institutes of Health, the Eye Bank Association of America, the W. Clement Stone Scholar fund, the Department of Defense, and a Physician-Scientist Award from Research to Prevent Blindness.
Financial Disclosure: The author has been a recipient of research funding from Amgen for this work but has no intellectual property rights over the technologies presented herein at this time.
Other Acknowledgments: The author wishes to thank the contribution of the following individuals for the conduct of the study and collection of data: Suning Zhu, MD, Iva Dekaris, MD, PhD, and Ying Qian, MD, PhD. Statistical consultation was provided by Debra Schaumberg, ScM, MPH.
REFERENCES
- 1.Rocha G, Deschenes J, Rowsey JJ. The immunology of corneal graft rejection. Crit Rev Immunol. 1998;18:305–325. doi: 10.1615/critrevimmunol.v18.i4.20. [DOI] [PubMed] [Google Scholar]
- 2.Qian Y, Dana MR. Molecular mechanisms of immunity in corneal allotransplantation and xenotransplantation. Expert Rev Mol Med. 2001;3:1–21. doi: 10.1017/S1462399401003246. [DOI] [PubMed] [Google Scholar]
- 3.Streilein JW. Immunobiology and immunopathology of corneal transplantation. Chem Immunol. 1999;73:186–206. doi: 10.1159/000058746. [DOI] [PubMed] [Google Scholar]
- 4.Streilein JW. Immunoregulatory mechanisms of the eye. Prog Retin Eye Res. 1999;18:357–370. doi: 10.1016/s1350-9462(98)00022-6. [DOI] [PubMed] [Google Scholar]
- 5.Niederkorn JY. The immune privilege of corneal allografts. Transplantation. 1999;67:1503–1508. doi: 10.1097/00007890-199906270-00001. [DOI] [PubMed] [Google Scholar]
- 6.Mader TH, Stulting RD. The high-risk penetrating keratoplasty. Ophthalmol Clin North Am. 1991;4:411–426. [Google Scholar]
- 7.Dana MR, Streilein JW. Loss and restoration of immune privilege in eyes with corneal neovascularization. Invest Ophthalmol Vis Sci. 1996;37:2485–2494. [PubMed] [Google Scholar]
- 8.Epstein RJ, Stulting RD, Hendricks RL, Harris DM. Corneal neovascularization. Pathogenesis and inhibition. Cornea. 1987;6:250–257. doi: 10.1097/00003226-198706040-00004. [DOI] [PubMed] [Google Scholar]
- 9.Dana MR, Qian Y, Hamrah P. Twenty-five year panorama of corneal immunology: emerging concepts in the immunopathogenesis of microbial keratitis, peripheral ulcerative keratitis, and corneal transplant rejection. Cornea. 2000;19:625–643. doi: 10.1097/00003226-200009000-00008. [DOI] [PubMed] [Google Scholar]
- 10.He YG, Ross J, Niederkorn JY. Promotion of murine orthotopic corneal allograft survival by systemic administration of anti-CD4 monoclonal antibody. Invest Ophthalmol Vis Sci. 1991;32:2723–2728. [PubMed] [Google Scholar]
- 11.Ayliffe W, Alam Y, Bell EB, McLeod D, Hutchinson IV. Prolongation of rat corneal graft survival by treatment with anti-CD4 monoclonal antibody. Br J Ophthalmol. 1992;76:602–606. doi: 10.1136/bjo.76.10.602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Pleyer U, Milani JK, Dukes A, et al. Effect of topically applied anti-CD4 monoclonal antibodies on orthotopic corneal allografts in a rat model. Invest Ophthalmol Vis Sci. 1995;36:52–61. [PubMed] [Google Scholar]
- 13.Ross J, He YG, Pidherney M, Mellon J, Niederkorn JY. The differential effects of donor versus host Langerhans cells in the rejection of MHC-matched corneal allografts. Transplantation. 1991;52:857–861. doi: 10.1097/00007890-199111000-00020. [DOI] [PubMed] [Google Scholar]
- 14.Niederkorn JY. Effect of cytokine-induced migration of Langerhans cells on corneal allograft survival. Eye. 1995;9:215–218. doi: 10.1038/eye.1995.42. [DOI] [PubMed] [Google Scholar]
- 15.Yamada J, Dana MR, Zhu S-N, Alard P, Streilein JW. Interleukin 1 receptor antagonist suppresses allosensitization in corneal transplantation. Arch Ophthalmol. 1998;116:1351–1357. doi: 10.1001/archopht.116.10.1351. [DOI] [PubMed] [Google Scholar]
- 16.Yamagami S, Dana MR, Tsuru T. Draining lymph nodes play an essential role in alloimmunity generated in response to high-risk corneal transplantation. Cornea. 2002;21:405–409. doi: 10.1097/00003226-200205000-00014. [DOI] [PubMed] [Google Scholar]
- 17.Chen L, Hamrah L, Cursiefen C, et al. Vascular endothelial growth factor receptor-3 (VEGFR-3) mediates induction of immunity to corneal transplants. Nature Med. 2004;10:813–815. doi: 10.1038/nm1078. [DOI] [PubMed] [Google Scholar]
- 18.Dana MR, Schaumberg DA, Kowal VO, et al. Corneal neovascularization after penetrating keratoplasty. Cornea. 1995;14:604–609. [PubMed] [Google Scholar]
- 19.Cursiefen C, Chen L, Jackson D, Dana MR, Streilein JW. Inhibition of hemangiogenesis and lymphangiogenesis after normal-risk corneal transplantation by neutralizing VEGF promotes graft survival. Invest Ophthalmol Vis Sci. 2004;45:2666–2673. doi: 10.1167/iovs.03-1380. [DOI] [PubMed] [Google Scholar]
- 20.Hamrah P, Huq SO, Liu Y, Zhang Q, Dana MR. Corneal immunity is mediated by heterogeneous population of antigen-presenting cells. J Leukoc Biol. 2003;74:172–178. doi: 10.1189/jlb.1102544. [DOI] [PubMed] [Google Scholar]
- 21.Dana MR, Hamrah P. Corneal antigen-presenting cells: function and identity. In: Zierhut M, Stern ME, Sullivan DA, editors. Immunology of the Lacrimal Gland, Tear Film, and Ocular Surface. London: Taylor and Francis Group; 2005. pp. 141–149. [Google Scholar]
- 22.He YG, Niederkorn JY. Depletion of donor-derived Langerhans cells promotes corneal allograft survival. Cornea. 1996;15:82–89. [PubMed] [Google Scholar]
- 23.Lechler RI, Batchelor JR. Restoration of immunogenicity to passenger cell-depleted kidney allografts by the addition of donor strain dendritic cells. J Exp Med. 1982;155:31–41. doi: 10.1084/jem.155.1.31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Dekaris I, Zhu SN, Dana MR. TNF-alpha regulates corneal Langerhans cell migration. J Immunol. 1999;162:4235–4239. [PubMed] [Google Scholar]
- 25.Yamagami S, Hamrah P, Miyamoto K, et al. CCR5 chemokine receptor mediates recruitment of MHC class II-positive Langerhans cells in the mouse corneal epithelium. Invest Ophthalmol Vis Sci. 2005;46:1201–1207. doi: 10.1167/iovs.04-0658. [DOI] [PubMed] [Google Scholar]
- 26.Dana R. Corneal antigen presentation: molecular regulation and functional implications. Ocul Surf. 2005;3(4 Suppl):S169–172. doi: 10.1016/s1542-0124(12)70248-3. [DOI] [PubMed] [Google Scholar]
- 27.Gillette TE, Chandler JW, Greiner JV. Langerhans cells of the ocular surface. Ophthalmology. 1982;89:700–711. doi: 10.1016/s0161-6420(82)34737-5. [DOI] [PubMed] [Google Scholar]
- 28.Jager MJ, Gregerson DS, Streilein JW. Regulators of immunological responses in the cornea and the anterior chamber of the eye. Eye. 1995;9:241–246. doi: 10.1038/eye.1995.47. [DOI] [PubMed] [Google Scholar]
- 29.Zhu S-N, Dana MR. Expression of cell adhesion molecules on limbal and neovascular endothelium in corneal inflammatory neovascularization. Invest Ophthalmol Vis Sci. 1999;40:1427–1434. [PubMed] [Google Scholar]
- 30.Luster AD. Chemokines—chemotactic cytokines that mediate inflammation. N Engl J Med. 1998;338:436–445. doi: 10.1056/NEJM199802123380706. [DOI] [PubMed] [Google Scholar]
- 31.Whitcup SM, Nussenblatt RB, Price FW, Jr, Chan CC. Expression of cell adhesion molecules in corneal graft failure. Cornea. 1993;12:475–480. doi: 10.1097/00003226-199311000-00003. [DOI] [PubMed] [Google Scholar]
- 32.Philipp W. Leukocyte adhesion molecules in rejected corneal allografts. Graefes Arch Clin Exp Ophthalmol. 1994;232:87–95. doi: 10.1007/BF00171669. [DOI] [PubMed] [Google Scholar]
- 33.Larkin DF, Calder VL, Lightman SL. Identification and characterization of cells infiltrating the graft and aqueous humour in rat corneal allograft rejection. Clin Exp Immunol. 1997;107:381–391. doi: 10.1111/j.1365-2249.1997.279-ce1171.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.He Y, Mellon J, Apte R, Niederkorn JY. Effect of LFA-1 and ICAM-1 antibody treatment on murine corneal allograft survival. Invest Ophthalmol Vis Sci. 1994;35:3218–3225. [PubMed] [Google Scholar]
- 35.Yamagami S, Obata H, Tsuru T, Isobe M. Suppression of corneal allograft rejection after penetrating keratoplasty by antibodies to ICAM-1 and LFA-1 in mice. Transplant Proc. 1995;27:1899–1900. [PubMed] [Google Scholar]
- 36.Zhu SN, Yamada J, Streilein JW, Dana MR. ICAM-1 deficiency suppresses host allosensitization and rejection of MHC-disparate corneal transplants. Transplantation. 2000;69:1008–1013. doi: 10.1097/00007890-200003150-00061. [DOI] [PubMed] [Google Scholar]
- 37.McDermott AM, Perez V, Huang AJ, et al. Pathways of corneal and ocular surface inflammation: a perspective from the Cullen symposium. Ocul Surf. 2005;3:S131–138. doi: 10.1016/s1542-0124(12)70238-0. [DOI] [PubMed] [Google Scholar]
- 38.Yamada J, Streilein JW, Dana MR. Role of tumor necrosis factor receptors TNFR-1 (p55) and TNFR-II (p75) in corneal transplantation. Transplantation. 1995;68:944–949. doi: 10.1097/00007890-199910150-00008. [DOI] [PubMed] [Google Scholar]
- 39.Tang HL, Cyster JG. Chemokine up-regulation and activated T cell attraction by maturing dendritic cells. Science. 1999;284:819–822. doi: 10.1126/science.284.5415.819. [DOI] [PubMed] [Google Scholar]
- 40.Yamagami S, Isobe M, Tsuru T. Characterization of cytokine profiles in corneal allograft with anti-adhesion therapy. Transplantation. 2000;69:1655–1659. doi: 10.1097/00007890-200004270-00022. [DOI] [PubMed] [Google Scholar]
- 41.Yamagami S, Miyazaki D, Ono SJ, Dana MR. Differential chemokine gene expression after corneal transplantation. Invest Ophthalmol Vis Sci. 1999;40:2892–2897. [PubMed] [Google Scholar]
- 42.Zhu S, Dekaris I, Duncker G, Dana MR. Early expression of proinflammatory cytokines interleukin-1 and tumor necrosis factor-alpha after corneal transplantation. J Interferon Cytokine Res. 1999;19:661–669. doi: 10.1089/107999099313811. [DOI] [PubMed] [Google Scholar]
- 43.Niederkorn JY, Peeler JS, Mellon J. Phagocytosis of particulate antigens by corneal epithelial cells stimulates interleukin-1 secretion and migration of Langerhans cells into the central cornea. Reg Immunol. 1989;2:83–90. [PubMed] [Google Scholar]
- 44.Staats HF, Lausch RN. Cytokine expression in vivo during murine herpetic stromal keratitis. Effect of protective antibody therapy. J Immunol. 1993;151:277–283. [PubMed] [Google Scholar]
- 45.Le J, Vilcek J. Tumor necrosis factor and interleukin 1: cytokines with multiple overlapping biological activities. Lab Invest. 1987;56:234–248. [PubMed] [Google Scholar]
- 46.Dinarello CA, Wolff SM. The role of interleukin-1 in disease. N Engl J Med. 1993;328:106–113. doi: 10.1056/NEJM199301143280207. [DOI] [PubMed] [Google Scholar]
- 47.Eisenberg SP, Evans RJ, Arend WP, et al. Primary structure and functional expression from complementary DNA of a human interleukin-1 receptor antagonist. Nature. 1990;343:341–346. doi: 10.1038/343341a0. [DOI] [PubMed] [Google Scholar]
- 48.Hannum CH, Wilcox CJ, Arend WP, et al. Interleukin-1 receptor antagonist activity of a human interleukin-1 inhibitor. Nature. 1990;343:336–340. doi: 10.1038/343336a0. [DOI] [PubMed] [Google Scholar]
- 49.Granowitz EV, Clark BD, Mancilla J, Dinarello CA. Interleukin-1 receptor antagonist competitively inhibits the binding of interleukin-1 to the type II interleukin-1 receptor. J Biol Chem. 1991;266:14147–14150. [PubMed] [Google Scholar]
- 50.Van Klink F, Leher H, Jager MJ, Alizadeh H, Taylor W, Niederkorn JY. Systemic immune response to Acanthamoeba keratitis in the Chinese hamster. Ocul Immunol Inflamm. 1997;5:235–244. doi: 10.3109/09273949709085064. [DOI] [PubMed] [Google Scholar]
- 51.Dana MR, Dai RP, Zhu SN, Yamada J, Streilein JW. Interleukin-1 receptor antagonist suppresses Langerhans cell activity and promotes ocular immune privilege. Invest Ophthalmol Vis Sci. 1998;39:70–77. [PubMed] [Google Scholar]
- 52.Dana MR, Yamada J, Streilein JW. Topical interleukin-1 receptor antagonist promotes corneal transplant survival. Transplantation. 1997;63:1501–1507. doi: 10.1097/00007890-199705270-00022. [DOI] [PubMed] [Google Scholar]
- 53.Eigler A, Sinha B, Hartmann G, Endres S. Taming TNF: strategies to restrain this proinflammatory cytokine. Immunol Today. 1997;18:487–492. doi: 10.1016/s0167-5699(97)01118-3. [DOI] [PubMed] [Google Scholar]
- 54.Sekine-Okano M, Lucas R, Rungger D, et al. Expression and release of tumor necrosis factor-alpha by explants of mouse cornea. Invest Ophthalmol Vis Sci. 1996;37:1302–1310. [PubMed] [Google Scholar]
- 55.Peschon JJ, Torrance DS, Stocking KL, et al. TNF receptor-deficient mice reveal divergent roles for p55 and p75 in several models of inflammation. J Immunol. 1998;160:943–952. [PubMed] [Google Scholar]
- 56.Kondo S, Sauder DN. Tumor necrosis factor (TNF) receptor type 1 (p55) is a main mediator for TNF-alpha-induced skin inflammation. Eur J Immunol. 1997;27:1713–1718. doi: 10.1002/eji.1830270718. [DOI] [PubMed] [Google Scholar]
- 57.Bazzoni F, Beutler B. The tumor necrosis factor ligand and receptor families. N Engl J Med. 1996;334:1717–1725. doi: 10.1056/NEJM199606273342607. [DOI] [PubMed] [Google Scholar]
- 58.McComb J, Gould T, Chlipala E, et al. Antiarthritic activity of soluble tumor necrosis factor receptor type I forms in adjuvant arthritis: correlation of plasma levels with efficacy. J Rheumatol. 1999;26:1347–1351. [PubMed] [Google Scholar]
- 59.Yamagami S, Hamrah P, Zhang Y, Liu Q, Huq S, Dana MR. Early ocular chemokine gene expression and leukocyte infiltration after high-risk corneal transplantation. Mol Vis. 2005;11:632–640. [PubMed] [Google Scholar]
- 60.Qian Y, Dekaris I, Yamagami S, Dana MR. Topical soluble tumor necrosis factor receptor type I suppresses ocular chemokine gene expression and rejection of allogeneic corneal transplants. Arch Ophthalmol. 2000;118:1666–1671. doi: 10.1001/archopht.118.12.1666. [DOI] [PubMed] [Google Scholar]
- 61.Williamson JS, DiMarco S, Streilein JW. Immunobiology of Langerhans cells on the ocular surface. I. Langerhans cells within the central cornea interfere with induction of anterior chamber associated immune deviation. Invest Ophthalmol Vis Sci. 1987;28:1527–1532. [PubMed] [Google Scholar]
- 62.Sano Y, Streilein JW, Ksander BR. Detection of minor alloantigen-specific cytotoxic T cells after rejection of murine orthotopic corneal allografts: evidence that graft antigens are recognized exclusively via the “indirect pathway. Transplantation. 1999;68:963–970. doi: 10.1097/00007890-199910150-00011. [DOI] [PubMed] [Google Scholar]
- 63.Sano Y, Ksander BR, Streilein JW. Fate of orthotopic corneal allografts in eyes that cannot support anterior chamber-associated immune deviation induction. Invest Ophthalmol Vis Sci. 1995;36:2176–2185. [PubMed] [Google Scholar]
- 64.Goslings WR, Yamada J, Dana MR, et al. Corneal transplantation in antibody-deficient hosts. Invest Ophthalmol Vis Sci. 1999;40:250–253. [PubMed] [Google Scholar]
- 65.Yamada J, Kurimoto I, Streilein JW. Role of CD4+ T cells in immunobiology of orthotopic corneal transplants in mice. Invest Ophthalmol Vis Sci. 1999;40:2614–2621. [PubMed] [Google Scholar]
- 66.Ksander BR, Sano Y, Streilein JW. Role of donor-specific cytotoxic T cells in rejection of corneal allografts in normal and high-risk eyes. Transpl Immunol. 1996;4:49–52. doi: 10.1016/s0966-3274(96)80034-7. [DOI] [PubMed] [Google Scholar]
- 67.Joo CK, Pepose JS, Stuart PM. T-cell mediated responses in a murine model of orthotopic corneal transplantation. Invest Ophthalmol Vis Sci. 1995;36:1530–1540. [PubMed] [Google Scholar]
- 68.Sonoda Y, Streilein JW. Orthotopic corneal transplantation in mice—evidence that the immunogenetic rules of rejection do not apply. Transplantation. 1992;54:694–704. doi: 10.1097/00007890-199210000-00026. [DOI] [PubMed] [Google Scholar]
- 69.The collaborative corneal transplantation studies (CCTS) Effectiveness of histocompatibility matching in high-risk corneal transplantation. The Collaborative Corneal Transplantation Studies Research Group. Arch Ophthalmol. 1992;110:1392–1403. [PubMed] [Google Scholar]
- 70.Sano Y, Ksander BR, Streilein JW. Murine orthotopic corneal transplantation in high-risk eyes: rejection is dictated primarily by weak rather than strong alloantigens. Invest Ophthalmol Vis Sci. 1998;38:1130–1138. [PubMed] [Google Scholar]
- 71.Raizman M. Corticosteroid therapy of eye disease. Fifty years later. Arch Ophthalmol. 1996;114:1000–1001. doi: 10.1001/archopht.1996.01100140208016. [DOI] [PubMed] [Google Scholar]
- 72.West-Mays JA, Strissel KJ, Sadow PM, Fini ME. Competence for collagenase gene expression by tissue fibroblasts requires activation of an interleukin 1 alpha autocrine loop. Proc Natl Acad Sci. 1995;92:6768–6772. doi: 10.1073/pnas.92.15.6768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Annunziato F, Galli G, Romagnani P, et al. Chemokine receptors and other surface molecules preferentially associated with human Th1 or Th2 cells. Microbes Infect. 1999;1:103–106. doi: 10.1016/s1286-4579(99)80021-5. [DOI] [PubMed] [Google Scholar]
- 74.Klintworth GK, Burger PC. Neovascularization of the cornea: current concepts of its pathogenesis. Int Ophthalmol Clin. 1983;23:27–39. doi: 10.1097/00004397-198302310-00005. [DOI] [PubMed] [Google Scholar]
- 75.Epstein RJ, Stulting RD. Corneal neovascularization induced by stimulated lymphocytes in inbred mice. Invest Ophthalmol Vis Sci. 1987;28:1505–1513. [PubMed] [Google Scholar]
- 76.Aiello LP. Vascular endothelial growth factor and the eye: biochemical mechanisms of action and implications for novel therapies. Ophthalmic Res. 1997;29:354–362. doi: 10.1159/000268033. [DOI] [PubMed] [Google Scholar]
- 77.Yatoh S, Kawakami Y, Imai M, et al. Effect of a topically applied neutralizing antibody against vascular endothelial growth factor on corneal allograft rejection of rat. Transplantation. 1998;66:1519–1524. doi: 10.1097/00007890-199812150-00016. [DOI] [PubMed] [Google Scholar]
- 78.Kuldo JM, Ogawara KI, Werner N, et al. Molecular pathways of endothelial cell activation for (targeted) pharmacological intervention of chronic inflammatory diseases. Curr Vasc Pharmacol. 2005;3:11–39. doi: 10.2174/1570161052773898. [DOI] [PubMed] [Google Scholar]
- 79.Biswas PS, Banerjee K, Zheng M, Rouse BT. Counteracting corneal immunoinflammatory lesion with interleukin-1 receptor antagonist protein. J Leukoc Biol. 2004;76:868–875. doi: 10.1189/jlb.0504280. [DOI] [PubMed] [Google Scholar]
- 80.Coxon A, Bolon B, Estrada J, et al. Inhibition of interleukin-1 but not tumor necrosis factor suppresses neovascularization in rat models of corneal angiogenesis and adjuvant arthritis. Arthritis Rheum. 2002;46:2604–2612. doi: 10.1002/art.10546. [DOI] [PubMed] [Google Scholar]
- 81.Hynes RO. A reevaluation of integrins as regulators of angiogenesis. Nat Med. 2002;9:918–921. doi: 10.1038/nm0902-918. [DOI] [PubMed] [Google Scholar]
- 82.Cantarella G, Risuglia N, Delleva R, et al. Trail inhibits angiogenesis stimulated by VEGF expression in human glioblastoma cells. Br J Cancer. 2006;94:1428–1435. doi: 10.1038/sj.bjc.6603092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Barraquer J. Immunosuppressive agents in penetrating keratoplasty. Am J Ophthalmol. 1985;100:61–64. doi: 10.1016/s0002-9394(14)74983-9. [DOI] [PubMed] [Google Scholar]
- 84.Rehany U, Waisman M. Suppression of corneal allograft rejection by systemic cyclosporine-A in heavily vascularized rabbit corneas following alkali burns. Cornea. 1994;13:447–453. doi: 10.1097/00003226-199409000-00013. [DOI] [PubMed] [Google Scholar]