

Abstract
Caregivers of Alzheimer’s disease patients endure chronic stress associated with a decline of immune function. To assess the psychological and immunological changes of caregivers, we compared depressive symptoms, PBMC composition, in vitro activation-induced proliferation and cytokine production, and telomere length and telomerase activity of 82 individuals (41 caregivers and 41 age- and gender-matched controls). We found depressive symptoms were significantly higher in caregivers than in controls (p < 0.001). Correspondingly, caregivers had significantly lower T cell proliferation but higher production of immune-regulatory cytokines (TNF-α and IL-10) than controls in response to stimulation in vitro. We examined the impact of these changes on cellular replicative lifespan and found that caregivers had significantly shorter telomere lengths in PBMC than controls (6.2 and 6.4 kb, respectively, p < 0.05) with similar shortening in isolated T cells and monocytes and that this telomere attrition in caregivers was not due to an increase of shorter telomere possessing T cell subsets in PBMC. Finally, we showed that basal telomerase activity in PBMC and T cells was significantly higher in caregivers than in controls (p < 0.0001), pointing to an unsuccessful attempt of cells to compensate the excessive loss of telomeres in caregivers. These findings demonstrate that chronic stress is associated with altered T cell function and accelerated immune cell aging as suggested by excessive telomere loss.
The progressive deteriorating conditions of Alzheimer’s disease (AD)3 patients put a considerable emotional and physical burden on their primary caregivers. The consequence of this chronic stress on biological and immunological function has begun to be understood (1). A general physiological response to stress includes the secretion of neuroendocrine hormones and catecholamines by hypothalamic-pituitary-adrenal axes and the sympathetic nervous system, which in turn modulates functions of various types of cells, including immune cells (2, 3). It has been shown that chronic stress leads to an increase in the levels of proinflammatory cytokines and a decline of the overall immune response, resembling the findings in age-associated changes (4). However, the precise mechanisms of psychological stress on immune dysfunction and aging remain to be elucidated.
Recent studies have shown that telomeres, the end structure of chromosomes, not only maintain chromosomal stability but also regulate cellular replicative lifespan of cells (5). Incomplete terminal synthesis of the lagging strand during DNA replication results in loss of terminal telomere repeats (TTAGGG)n, and the substantial telomere shortening that occurs with successive cell division leads to cell division arrest or senescence (6). In contrast, a telomere-synthesizing enzyme, telomerase, can elongate telomeres and prevent their extensive shortening. In lymphocytes, telomere attrition has been observed with cell division in vitro and with differentiation and age in vivo, and the expression of telomerase in lymphocytes is highly regulated during development and activation (7). Naive T cells have longer telomeres than their descendent memory T cells (8, 9). Newly generated T cells are CD28+ and have longer telomeres than repeatedly stimulated T cells that lose expression of CD28 (10). The rate of telomere loss in lymphocytes appears to be dependent on the level of telomerase activity (11) and enhanced expression of telomerase is capable of minimizing telomere loss or even maintaining telomere length in actively dividing lymphocytes (12, 13).
The consequence of chronic stress on telomere length and telomerase activity of PBMC was recently reported. Higher oxidative stress, lower basal level of telomerase activity, and shorter telomere length of PBMC were found in mothers of chronically ill children (14), and shorter telomere length of PBMC in chronically stressed individuals with mood disorders (15). Although these findings provide evidence that stress may modulate cellular aging, they also raise more questions. These studies lacked parallel assessment of the function of the immune cells and therefore the impact of telomere shortening on PBMC function is unclear. In addition, as PBMC contain many types of cells and subsets of the same type of cells with different telomere lengths, it is not clear whether the shortened telomeres of PBMC were due to telomere shortening in all types of cells or an increase of shorter telomere-possessing cells. Moreover, PBMC consist of mainly noncycling resting lymphocytes that express little telomerase activity and monocytes expressing no detectable telomerase. The significance of the low level of telomerase activity difference in these resting cells is not known. In contrast, activation can dramatically induce telomerase activity in lymphocytes. Whether there is any change in activation-induced telomerase activity in lymphocytes in these stressed individuals has not been examined.
To address these questions, we designed and conducted a study of the primary caregivers (spouse and offspring) of AD patients who endure substantial psychological and physical stress and their age- and gender-matched controls. We analyzed the composition of PBMC and assessed T cell function, so that the functional and telomere length changes of these immune cells (PBMC, T cells, and monocytes) could be specifically analyzed. In addition, we compared both basal and activation-induced telomerase activity of T cells (in PBMC and isolated T cells) between caregivers and controls. Our findings suggested that chronic stress alters T cell function and accelerates T cell aging.
Materials and Methods
Information of the study subjects
Eighty-two subjects (41 caregivers and 41 controls) were part of a larger study on caregiver stress, immune function, and health (4, 16); for this report, we used frozen PBMC for all caregivers and controls in the cohort who could be individually matched on age (±5 years) and gender. Twenty-six caregivers were spouses and 15 caregivers were offspring of AD patients. The controls were age and gender matched to the caregivers. There were 11 male and 30 female subjects with a mean age of 65 ± 1 for the caregiver and control groups. The Ohio State University Biomedical Research Review Committee approved the project; all participants gave written informed consent before participation. Depressive symptoms were assessed by the Center for Epidemiological Studies Depression Scale, which has been used extensively in population studies to provide data on depression (17). The scale was administered when blood was drawn.
Isolation of PBMC, T cells, and monocytes
PBMC were isolated by centrifugation in a Lympho-Ficoll gradient and preserved at −80°C before analysis. Monocytes and T cells were isolated by a positive selection procedure using Dynabeads conjugated with CD14 and CD2, respectively (Invitrogen Life Technologies) based on the manufacturer’s instruction. Briefly, PBMC were incubated with anti-CD14 Ab-conjugated beads at the ratio of 4 beads per monocytes for 25 min; bead-bound monocytes were then isolated using a magnet. The remaining PBMC were incubated with anti-CD2 Ab-conjugated beads for 25 min, and anti-CD2-bound T cells were further purified. The yield of monocytes and T cells was counted and used in the specified experiment.
Flow cytometry analysis
The composition of PBMC was analyzed by flow cytometry with a panel of Abs against CD4 (PE), CD8 (PE Alexa Fluor 700), CD14 (PE), CD16 (allophycocyanin), CD19 (FITC), CD28 (FITC), and CD45RA (allophycocyanin) (Invitrogen Life Technologies) according to the manufacturer’s instructions. The data of four-color-stained cells were collected by FACSCalibur and analyzed by CellQuest Pro (BD Biosciences).
Stimulation of T cells in vitro
Frozen PBMCs in 90% FBS and 10% DMSO were thawed using the following procedure. First, the frozen cells were incubated in a 37°C water bath for ~1 min to raise the temperature from −80°C to 0°C. Second, the cells were transferred to a 15-ml tube and 10 ml of warm medium, RPMI 1640 with 10% FBS and 10 U/ml penicillin/10 μg/ml streptomycin (Invitrogen Life Technologies), was gradually added into the tube to raise the temperature from 0°C to 37°C in 10 min. Thawed PBMC and purified T cells were cultured at 37°C in an atmosphere of 5% CO2 for 2–5 h and then stimulated with anti-CD3/CD28 mAb-conjugated beads (Invitrogen Life Technologies) at a 1:1 cell/bead ratio. Stimulated PBMC were harvested at day 3 for cell proliferation and telomerase activity analysis.
Proliferation
PBMC were cultured at 5 × 104 cells/well in quadruplicate in 96-well flat-bottom plates in 0.2 ml of RPMI 1640 medium with 10% FBS and penicillin-streptomycin. After 48 h stimulation with anti-CD3/28 Abs, 1 μCi [3H] thymidine in 50 μl of medium was added to each well and incubated for an additional 20 h before harvest. The counts of [3H]thymidine were measured using a scintillation counter (Beckman Coulter). The [3H]thymidine counts of stimulated T cells were subtracted from the medium control and then normalized to the number of T cells in PBMC.
Cytokine measurement
PBMC were cultured at 2 × 106 cells/well in 12-well flat-bottom plates in 2.0 ml of RPMI 1640 medium supplemented with 10% FBS and penicillin-streptomycin. After 72 h of stimulation with anti-CD3/CD28, the cell supernatants were harvested and the concentration of cytokines (IL-2, IL-4, IL-6, IL-8, IL-10, GM-CSF, IFN-γ, and TNF-α) were quantified with the BioPlex protein array system according to the recommended procedure (Bio-Rad). The concentrations of cytokines from T cell-stimulated cell supernatants were normalized to the number of T cells in PBMC. Sera from these individuals were collected and the concentration of TNF-α was measured by ELISA (4).
Telomere length
Telomere length was measured by Southern blot based on methods previously described (8). In brief, genomic DNA was isolated from PBMC, T cells, and monocytes using a DNA isolation kit (Gentra Systems) and digested with HinfI and RsaI (Roche Molecular Biochemicals) (40U of each enzyme/5 μg of DNA). Digested DNA was loaded at 1 μg/well on a 0.6% agarose gel and separated by electrophoresis. The gel was dried at 65°C for 2 h, denatured, and neutralized. The hybridization was performed with a [32P]end-labeled oligonucleotide (CCCTAA)4 probe, at 43°C overnight. The gels were washed three times (5× SSC/0.1% SDS, 2× SSC/0.1% SDS, and 3.2 M tetramethylammonium chloride/0.1% SDS) at 45°C, followed by analysis with a phosphor imager (Typhoon 9410; Amersham Biosciences). Mean terminal telomere restriction fragment (TRF) was calculated as described previously (18).
Telomerase activity
Telomerase activity was measured using a modified telomeric repeats amplification protocol (TRAP), as described previously (19). In brief, cell lysates were extracted from unstimulated and stimulated PBMC and isolated T cells with ice-cold CHAPS (Calbiochem) lysis buffer at 100 μl/106 cells. The cell extracts were used for telomere synthesis and incubated at 30°C for 1 h, followed by amplification of telomere repeats under the described conditions with 35 cycles in a DNA Engine Tetrad (MJ Research). The Ts primer was conjugated with a fluorescent dye, TAMRA, which was used to visualize and quantitate the amplified telomere repeats after electrophoretic separation on a 12% polyacrylamide gel (Novex; Invitrogen Life Technologies) and visualization with a fluorescent imaging scanner (Typhoon 9410; GE Healthcare). The quantitation of telomere products was conducted by using Image-Quant software. Telomerase activity was expressed as the ratio of the intensity of telomere products and the intensity of the internal control. The level of telomerase activity was based on an equivalent number of cells.
Statistical analysis
The differences of biological parameters between the caregivers and their controls were analyzed by a paired two-tailed or one-tailed Student’s t test, and p values <0.05 were considered significant.
Results
Caregivers of AD patients exhibit higher levels of stress than their age- and gender-matched controls
Caregivers in this study were spouses or offspring of AD patients who experienced and endured chronic psychological stress. Our cohort consisted of 41 caregivers and 41 age- and gender-matched controls. Their average age was 65 ± 1 and the average time of caregiving was 5.2 ± 0.5 years. There were 11 male and 30 female subjects in each group. To determine the levels of depressive symptoms, we used the Center for Epidemiological Studies Depression Scale (17) and found that caregivers had significantly higher average levels of depressive symptoms (at least 2-fold higher, p < 0.001) than did controls (Fig. 1). This finding confirms that caregivers experience higher levels of psychological stress than their controls.
FIGURE 1.
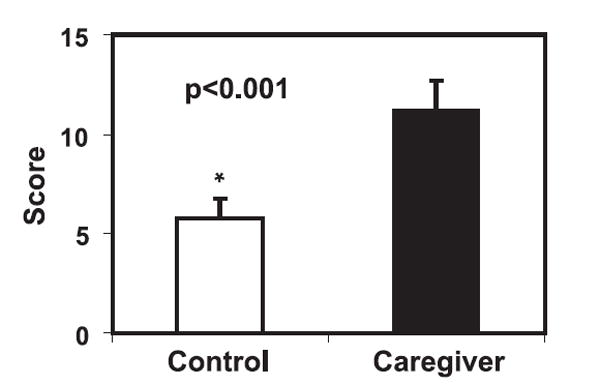
Increased psychological stress in caregivers of AD patients. The levels of depression were measured in caregivers and controls. Data are presented as mean and SEM (n = 41). Depression scores were significantly higher in caregivers than in controls as analyzed by paired Student’s t test.
Caregivers of AD patients have similar PBMC compositions but lower T cell proliferation and higher production of proinflammatory cytokines than their controls
To assess the immunological changes in caregivers, we first analyzed the composition of PBMC. As expected, there were some differences in the percentage of each type of cells (T and B cells, NK cells, and monocytes) among different individuals but the differences in the percentages were not statistically significant between caregivers and controls (Table I). We further analyzed the subsets of T cells (naive and memory, and CD28+ and CD28− T cells) and again we found no significant differences in the percentages of these T cell subsets between the two groups (Table I). To examine the functional changes of T cells, we analyzed T cell proliferation and cytokine production in response to stimulation. PBMC were stimulated with anti-CD3 plus anti-CD28 Abs for 3 days and T cell proliferation was measured by [3H]thymidine incorporation. Eight selected T cell cytokines (IL-2, IL-4, IL-6, IL-8, IL-10, GM-CSF, IFN-γ, and TNF-α) were measured from culture supernatants of stimulated T cells. We found that activation-induced T cell proliferation was significantly (p = 0.04) lower in caregivers (55,173 ± 6,736 cpm) compared with controls (72,433 ± 6,919 cpm) (Fig. 2A). In contrast, stimulated T cells produced significantly higher levels of TNF-α (p < 0.05) and IL-10 (p < 0.05) from caregivers than from controls (Fig. 2B). In parallel, we found significantly higher levels of TNF-α in serum from caregivers than from controls (p < 0.05) (Fig. 2C). The levels of IL-2, IL-4, IL-6, IL-8, GM-CSF, and IFN-γ were not significantly different between the two groups (Table II). These findings suggest that chronic stress significantly reduced the proliferative capacity of T cells, yet enhanced the production of immune-regulatory cytokines TNF-α and IL-10.
Table I.
Composition of cells in PBMC of caregivers and controlsa
| Type of Cell | Control | Caregiver | p Valueb |
|---|---|---|---|
| CD4 T cell | 32.4 ± 1.2 | 32.9 ± 1.5 | 0.40 |
| CD4+CD28− | 2.0 ± 0.3 | 2.2 ± 0.3 | 0.32 |
| CD4+CD45RA+ | 11.0 ± 0.8 | 13.3 ± 1.4 | 0.09 |
| CD8 T cell | 11.5 ± 0.8 | 13.0 ± 0.8 | 0.10 |
| CD8+CD28− | 6.3 ± 0.6 | 9.4 ± 2.0 | 0.07 |
| CD8+CD45RA+ | 5.3 ± 0.5 | 7.4 ± 1.2 | 0.07 |
| B cell (CD19+) | 8.5 ± 0.6 | 8.8 ± 0.7 | 0.35 |
| NK cell (CD16+) | 11.1 ± 0.9 | 10.2 ± 0.7 | 0.20 |
| Monocyte (CD14+) | 10.6 ± 0.9 | 10.0 ± 0.6 | 0.28 |
PBMC were analyzed by FACS and data are presented as mean ± SEM (n = 38).
Statistical analysis was done by the paired Student t test.
FIGURE 2.
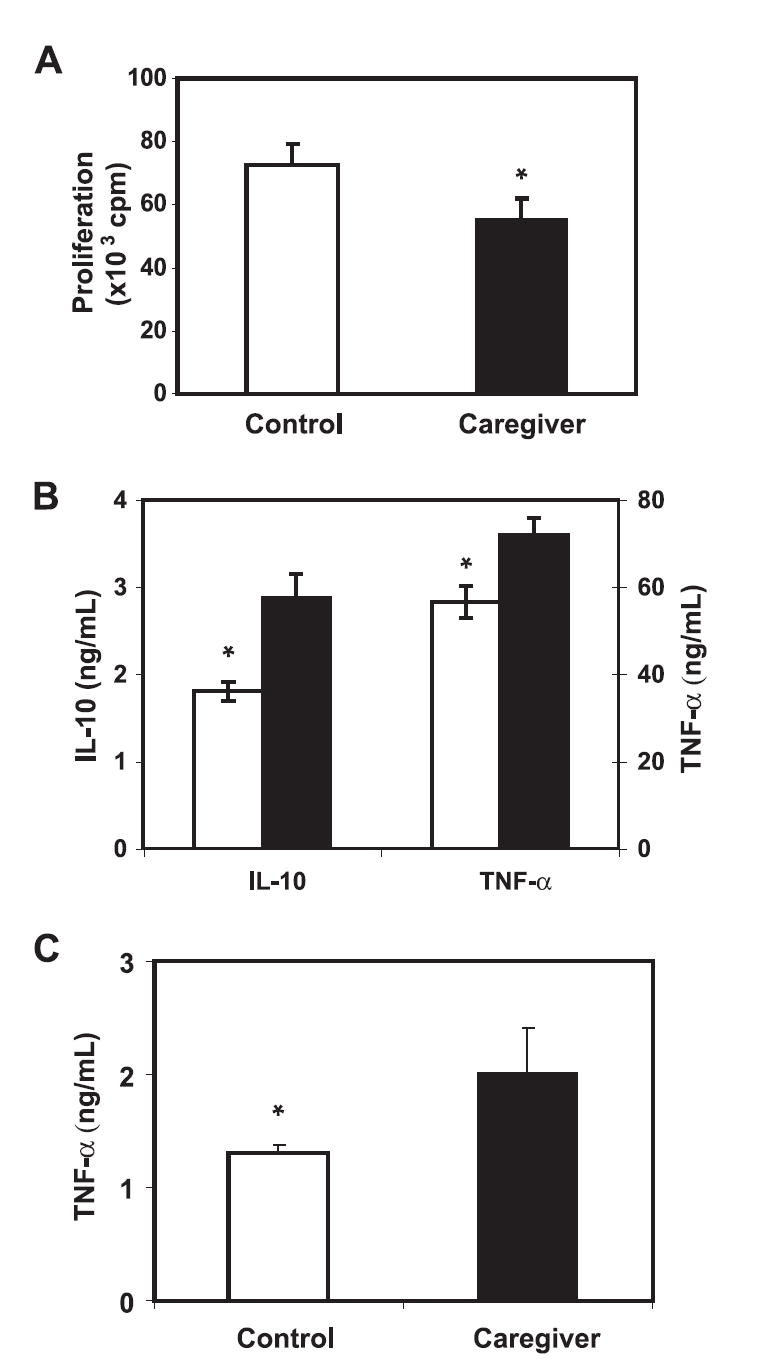
Reduced proliferation and increased proinflammatory cytokine production of T cells in caregivers. A, Activation-induced T cell proliferation is lower in caregivers than in controls. PBMC were stimulated with anti-CD3 and anti-CD28 Abs for 72 h in the presence of 1 μCi [3H]thymidine for the last 20 h. [3H]Thymidine incorporation was quantified using a liquid scintillation counter and normalized based on the number of T cells. Data are presented as mean (×103) and SEM (n = 38). B, Production of TNF-α and IL-10 are higher in caregivers than in controls. Culture supernatants of PBMC were collected after 72 h stimulation by anti-CD3 and anti-CD28 Abs, and TNF-α and IL-10 were quantified with the BioPlex protein array system. The concentrations of cytokines are presented as nanograms per milliliter. C, Higher levels of TNF-α in serum of caregivers than that of controls. Sera of 41 pairs of subsets were collected and TNF-α concentration was determined by ELISA (4). Data are presented as mean and SEM (n = 37). All statistical analyses were done by paired Student’s t test; *, p < 0.05 by Students’ t test.
Table II.
Cytokine production in stimulated T cells of caregivers and controlsa
| Type of Cytokine | Control | Caregiver | p Valueb | n |
|---|---|---|---|---|
| IL-2 | 164.7 ± 23.6 | 181.3 ± 20.6 | 0.26 | 34 |
| IL-4 | 0.2 ± 0.02 | 0.2 ± 0.02 | 0.12 | 37 |
| IL-6 | 7.5 ± 2.3 | 6.2 ± 1.8 | 0.27 | 37 |
| IL-8 | 662 ± 140 | 504 ± 70 | 0.09 | 37 |
| GM-CSF | 3.0 ± 0.4 | 3.1 ± 0.4 | 0.37 | 37 |
| IFN-γ | 15.1 ± 1.5 | 14.0 ± 1.4 | 0.25 | 37 |
The concentrations of cytokines are micrograms per milliliter. Data are presented as mean and SEM (n = 37, 34).
Statistical analysis was done by the paired Student t test.
Caregivers of AD patients have significantly shorter telomere length of PBMC than controls
We then analyzed the impact of chronic stress on the telomere length of PBMC. We found that telomere length of PBMC was generally correlated with the age of the individuals for both care-givers and their controls (Fig. 3A). The average rate of telomere shortening was 19 and 25 bp/year for both caregiver and control groups, respectively. Strikingly, the average telomere lengths of PBMC from caregivers were 6.2 ± 0.1 kb compared with 6.4 ± 0.1 kb from controls and caregivers had significantly shorter telomere lengths than controls (p < 0.05) (Fig. 3B). As the compositions of PBMC and shorter telomere possessing T cell subsets were similar between caregivers and controls, the difference of telomere length observed here in PBMC is likely reflecting telomere loss across all major cell types including T cells, B cells, and monocytes. To further confirm this, T cells and monocytes were isolated from PBMC and their telomere lengths were determined. As expected, we found that the mean telomere length of T cells from caregivers were 6.3 ± 0.2 kb compared with 6.5 ± 0.2 kb from controls, and that the mean telomere length of monocytes from caregivers were 5.7 ± 0.1 kb compared with 5.9 ± 0.1 kb from controls (Fig. 3C). Due to the limited samples of isolated cells (11 and 21 for T cell and monocyte, respectively), the p values did not reach statistical significance (0.08 and 0.12 for T cell and monocyte, respectively). On average, caregivers had shorter telomere length in PBMC (240 bp less), in T cells (261 bp less), and in monocytes (198 bp less) than those of controls. These findings suggest that excessive loss of telomere occurs in both T cells and monocytes.
FIGURE 3.
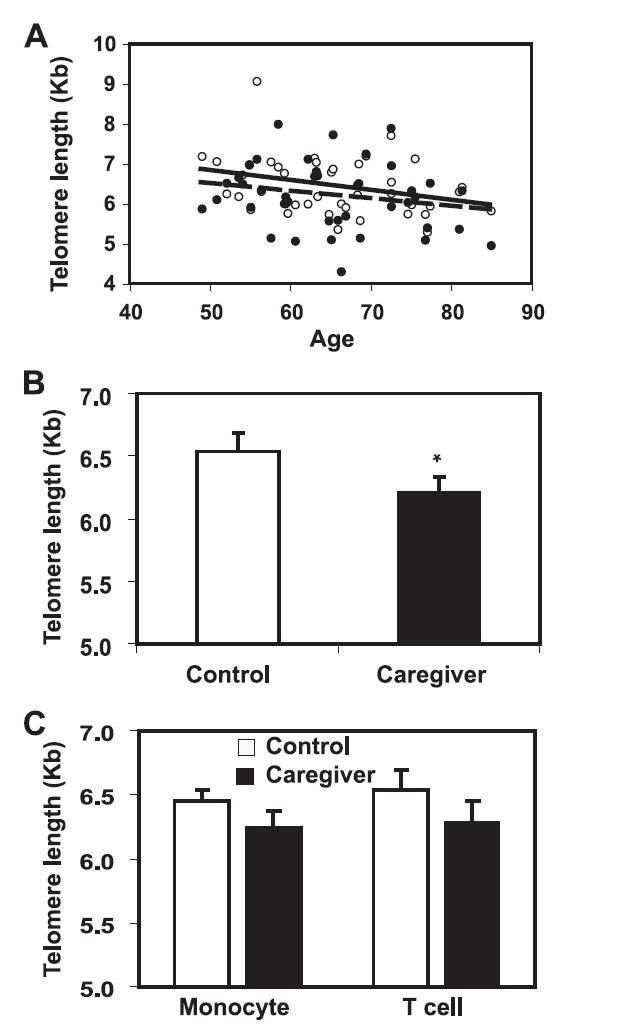
Shortened telomere length of PBMC, T cells, and monocytes in caregivers. A, Telomere length of PBMC decreases with age in caregivers and controls. Mean TRF (mTRF) length for caregivers and controls is plotted against their age. Caregivers: solid circle and dotted line; controls: open circle and solid line. B, Caregivers have shorter telomere length than controls. The mean TRF was calculated and compared between caregivers and controls. Data are presented as mean and SEM (n = 41). C, Caregivers have shorter T cell and monocyte telomere length than controls. The mean TRF was calculated and compared between caregivers and controls. Data are presented as mean and SEM (n = 21 and n = 11 in T cells and monocytes, respectively); *, p < 0.05 by Students’ t test.
Telomerase activity was significantly higher at the basal level but similar after activation of T cells from caregivers of AD patients as compared with controls
To assess whether telomere loss was related to the reduction of telomerase activity in lymphocytes, we compared both basal and activation-induced telomerase activity between caregivers and their controls. In contrast to the previous finding of lower levels of telomerase activity in mothers of sick children (14), we found a significantly higher level of telomerase activity in PBMC from caregivers than from controls (1.4 ± 0.2 and 0.6 ± 0.1, respectively, p < 0.0001) (Fig. 4A) and in T cells from caregivers than from controls (2.2 ± 0.5 and 1.5 ± 0.3, respectively, p < 0.05) (Fig. 4B). After stimulation, telomerase activity was similar in activated PBMC (Fig. 4C) and T cells (data not shown) between caregivers than in controls. The increased basal telomerase activity in PBMC and T cells may reflect the attempt of immune cells to compensate for the excessive loss of telomeres of caregivers, while the ability of activation-induced telomerase activity appears normal in caregivers.
FIGURE 4.
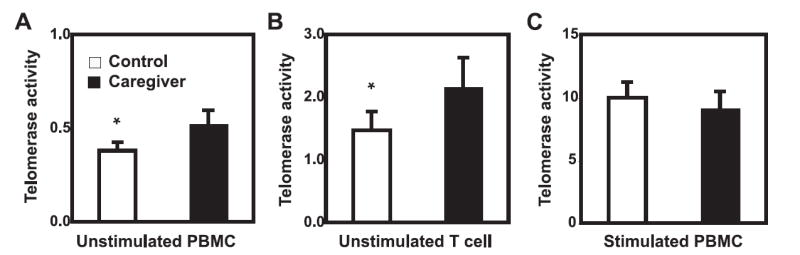
Increased basal level but no difference in activation-induced telomerase activity of PBMC and T cells in caregivers. A, Basal PBMC telomerase activity was higher in caregivers compared with controls. Telomerase activity was measured by TRAP assay and normalized to total lymphocytes. B, Basal T cell telomerase activity was higher in caregivers compared with controls. Telomerase activity was measured by TRAP assay. The activity was presented at mean and SEM (n = 12). C, Activation-induced telomerase activity was not significantly different between caregivers and controls. PBMC were stimulated in vitro with anti-CD3 plus anti-CD28 Abs for 72 h and harvested for measurement of telomerase activity. Data are presented as mean and SEM (n = 38); *, p < 0.0001 by Students’ t test.
Discussion
The findings of this study support the emerging notion that chronic psychological stress has a negative impact on immune cell function and may accelerate their aging. By parallel analyses of composition and function of immune cells and their telomere length and telomerase activity, we provide compelling evidence that chronic stress influences at least three aspects of T cell functions: 1) hyperproduction of the immune-regulatory cytokine TNF-α and inhibitory cytokine IL-10; 2) decreased activation-induced proliferation without reduction of growth-related cytokines such as IL-2 and IL-4; and 3) impaired telomere length maintenance, even with up-regulated telomerase. However, the mechanistic pathways of chronic stress that lead to the impairment of these immune cell functions remain to be elucidated.
Accumulating evidence suggests that chronic stress is associated with decline of immune function and may accelerate aging (4, 20). In this study, we found a reduction of T cell proliferation and increased production of TNF-α and IL-10 in response to in vitro stimulation in caregivers, but normal production of other cytokines such as IL-2, IL-4, IL-6, IL-8, GM-CSF, and IFN-γ (Table II). It is interesting to note that IL-10 has antioxidant function (21) and the increased expression of IL-10 in the caregivers may reflect a feedback response of T cells in reaction to increased oxidative stress. Our analysis of the inducible NO synthase also showed a correlation of increased expression of inducible NO synthase and depressive symptoms (our unpublished data). Additional experiments will be needed to elucidate the changes of oxidative stress in response to caregiving. Previous studies have reported several age-associated changes in T cells (22). However, we did not observe significant changes in the number or proportion of CD28− T cells or other age-related subsets in caregivers as compared with controls. Considering the average caregiving time of our subjects was only 5 years, it may take a longer time to observe the age-associated gross changes of T cell compositions. Together, these findings suggest, in the caregiver population studied here, there are significant functional defects of T cells but no detectable accelerated age-associated changes in T cell subpopulations.
Accelerated loss of telomeres has been reported in conditions associated with defective telomerase (23-25). Dyskeratosis congenita, a human genetic disorder which is marked by defects of telomerase RNA directly or indirectly via different genetic abnormalities, displays a progressive bone marrow failure syndrome affecting several tissues and organs (24, 25). The common feature of these affected tissues and organs is that they are highly regenerative, requiring a high rate of cell proliferation for their function. Without sufficient telomerase to compensate for telomere loss, a high rate of proliferation leads to shortened telomere length in these cells. Similar findings were also reported in telomerase-deficient mice (23, 26). Epel et al. (14) reported telomere shortening in the peripheral blood leukocytes of women who were caregivers for chronically ill children. However, it was not clear whether shortened telomeres of PBMC in the report were due to an increase of shorter telomere possessing T cells in PBMC or across the board telomere shortening in all types of cells. Our findings here demonstrate that the loss of telomeres in caregivers was not due to the increase of shorter telomere cells in PBMC. Based on the reported rates of telomere attrition in PBMC (ranging from 31 bp/year to 67 bp/year) (27, 28), the differences of 240 bp shorter could account for ~4–8 years of shortened lifetime and could be even greater if it was calculated based on the rate of 19 bp/year of this study.
Epel et al. (14) also reported a reduced basal level of telomerase activity in caregivers compared with controls and concluded that defects of telomerase activity in the caregivers may contribute to the loss of telomere. In contrast, we found here a significant increase in basal telomerase activity but not in the activation-induced levels of telomerase activity in caregivers of AD patients. It is not clear what the reason is behind the difference of these two findings. The increased basal telomerase activity in caregivers of AD patients may reflect an attempt of immune cells to compensate for excessive loss of telomeres. In this regard, caregivers in this study may be at the relative early phase of stress-induced impairment of telomere maintenance as compared with the mothers of sick children. Further studies are needed to elucidate the kinetic relationship of cell proliferation and telomerase activity in the maintenance of telomere length.
Telomerase activity is strictly regulated in human cells during development and differentiation (29), and can be positively or negatively regulated by cytokines and hormones. IL-2, IL-7, and IL-15 are among cytokines that are capable of inducing telomerase in T cells (19, 30). In contrast, IFN-α, TGF-β, and dexamethasone are capable of reducing telomerase activity in different types of cells (31-33). Although it is known that chronic stress can alter the balance of the production of hormones and cytokines, the specific hormones and/or cytokines that are responsible for regulation of telomerase in immune cells is unknown. In this study, we found that TNF-α levels were significantly higher in supernatants of activated T cells and serum from caregivers than from controls. However, the role of TNF-α in regulation of telomerase is still controversial. Akiyama et al. (34) showed that TNF-α can induce activation and nuclear translocation of telomerase in the first hour following stimulation of PBMC. But the long-term effects of TNF-α on telomerase activity are unclear, as is the identity of cell types in PBMC that are responsible for such changes. In contrast, Beyne-Rauzy et al. (35) recently reported that TNF-α inhibits human telomerase reverse transcriptase expression in myeloid cells through activation of a JNK pathway. It remains to be determined what the role of elevated TNF-α may be in regulation of telomerase activity in caregivers. We also found that IL-10 levels were significantly higher in supernatants of activated T cells from caregivers than from controls. Previous caregiver studies have found a higher expression of IL-10 in response to stress experienced by caregivers (36). However, it remains to be determined whether an increase in the immuno-suppressive cytokine, IL-10, is a counter measurement for an increase in proinflammatory cytokines or other unidentified changes in caregivers of AD patients.
It is now evident that individuals experiencing chronic stress are associated with shortened telomere length in their PBMC. However, the rate of telomere attrition in these individuals is not known. As telomere length is influenced by genetic factors and exhibits considerable polymorphisms within the population, a longitudinal analysis will be required to determine the rate of telomere attrition, changes of telomerase activity, and decline of immune function in association with the levels and duration of chronic stress of these caregivers and their controls. It is equally important to determine the physiological impact of telomerase activity and shortened telomeres on the overall function of immune cells. Further determination of how the psychological stress signals translate and influence cellular functions will bridge the mechanistic gap linking these two arenas.
Acknowledgments
We thank Richard Hodes and Mark Mattson for critical reading and comments on the manuscript. We thank Jettanong Klaewsonghram and Wai Kai Chiu for assistance with the proliferation assay, Bob Pyle for cytokine measurement, and Christa Morris for FACS analysis. We thank the Central Ohio Alzheimer’s Association for their help with recruitment.
Footnotes
A.K.D., Y.Y., H.N., Y.Z., and N.-p.W. were supported by the Intramural Research Program of the National Institute on Aging, National Institutes of Health (NIH). J.K.K.-G. and R.G. were supported in part by NIH Grant AG025732, NIH General Clinical Research Center Grant MO1-RR-0034, Comprehensive Cancer Center Core Grant CA16058, and the Gilbert and Kathryn Mitchell Endowment.
Abbreviations used in this paper: AD, Alzheimer’s disease; TRF, telomere restriction fragment; TRAP, telomeric repeats amplification protocol.
Disclosures The authors have no financial conflict of interest.
References
- 1.Glaser R, Kiecolt-Glaser JK. Stress-induced immune dysfunction: implications for health. Nat Rev Immunol. 2005;5:243–251. doi: 10.1038/nri1571. [DOI] [PubMed] [Google Scholar]
- 2.Pedersen WA, Wan R, Mattson MP. Impact of aging on stress-responsive neuroendocrine systems. Mech Ageing Dev. 2001;122:963–983. doi: 10.1016/s0047-6374(01)00250-0. [DOI] [PubMed] [Google Scholar]
- 3.Mayer EA, Fanselow MS. Dissecting the components of the central response to stress. Nat Neurosci. 2003;6:1011–1012. doi: 10.1038/nn1003-1011. [DOI] [PubMed] [Google Scholar]
- 4.Kiecolt-Glaser JK, Preacher KJ, MacCallum RC, Atkinson C, Malarkey WB, Glaser R. Chronic stress and age-related increases in the proinflammatory cytokine IL-6. Proc Natl Acad Sci USA. 2003;100:9090–9095. doi: 10.1073/pnas.1531903100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Blackburn EH. Switching and signaling at the telomere. Cell. 2001;106:661–673. doi: 10.1016/s0092-8674(01)00492-5. [DOI] [PubMed] [Google Scholar]
- 6.Hao LY, Armanios M, Strong MA, Karim B, Feldser DM, Huso D, Greider CW. Short telomeres, even in the presence of telomerase, limit tissue renewal capacity. Cell. 2005;123:1121–1131. doi: 10.1016/j.cell.2005.11.020. [DOI] [PubMed] [Google Scholar]
- 7.Hodes RJ, Hathcock KS, Weng NP. Telomeres in T and B cells. Nat Rev Immunol. 2002;2:699–706. doi: 10.1038/nri890. [DOI] [PubMed] [Google Scholar]
- 8.Weng NP, Levine BL, June CH, Hodes RJ. Human naive and memory T lymphocytes differ in telomeric length and replicative potential. Proc Natl Acad Sci USA. 1995;92:11091–11094. doi: 10.1073/pnas.92.24.11091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Rufer N, Brummendorf TH, Kolvraa S, Bischoff C, Christensen K, Wadsworth L, Schulzer M, Lansdorp PM. Telomere fluorescence measurements in granulocytes and T lymphocyte subsets point to a high turnover of hematopoietic stem cells and memory T cells in early childhood. J Exp Med. 1999;190:157–167. doi: 10.1084/jem.190.2.157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Monteiro J, Batliwalla F, Ostrer H, Gregersen PK. Shortened telomeres in clonally expanded CD28−CD8+ T cells imply a replicative history that is distinct from their CD28+CD8+ counterparts. J Immunol. 1996;156:3587–3590. [PubMed] [Google Scholar]
- 11.Weng NP, Palmer LD, Levine BL, Lane HC, June CH, Hodes RJ. Tales of tails: regulation of telomere length and telomerase activity during lymphocyte development, differentiation, activation, and aging. Immunol Rev. 1997;160:43–54. doi: 10.1111/j.1600-065x.1997.tb01026.x. [DOI] [PubMed] [Google Scholar]
- 12.Rufer N, Migliaccio M, Antonchuk J, Humphries RK, Roosnek E, Lansdorp PM. Transfer of the human telomerase reverse transcriptase (TERT) gene into T lymphocytes results in extension of replicative potential. Blood. 2001;98:597–603. doi: 10.1182/blood.v98.3.597. [DOI] [PubMed] [Google Scholar]
- 13.Roth A, Yssel H, Pene J, Chavez EA, Schertzer M, Lansdorp PM, Spits H, Luiten RM. Telomerase levels control the lifespan of human T lymphocytes. Blood. 2003;102:849–857. doi: 10.1182/blood-2002-07-2015. [DOI] [PubMed] [Google Scholar]
- 14.Epel ES, Blackburn EH, Lin J, Dhabhar FS, Adler NE, Morrow JD, Cawthon RM. Accelerated telomere shortening in response to life stress. Proc Natl Acad Sci USA. 2004;101:17312–17315. doi: 10.1073/pnas.0407162101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Simon NM, Smoller JW, McNamara KL, Maser RS, Zalta AK, Pollack MH, Nierenberg AA, Fava M, Wong KK. Telomere shortening and mood disorders: preliminary support for a chronic stress model of accelerated aging. Biol Psychiatry. 2006;60:432–435. doi: 10.1016/j.biopsych.2006.02.004. [DOI] [PubMed] [Google Scholar]
- 16.Kiecolt-Glaser JK, Marucha PT, Malarkey WB, Mercado AM, Glaser R. Slowing of wound healing by psychological stress. Lancet. 1995;346:1194–1196. doi: 10.1016/s0140-6736(95)92899-5. [DOI] [PubMed] [Google Scholar]
- 17.Radloff LS. The CES-D scale: a self-report depression scale for research in the general population. Appl Psychol Meas. 1977;1:385–401. [Google Scholar]
- 18.Harley CB, Futcher AB, Greider CW. Telomeres shorten during ageing of human fibroblasts. Nature. 1990;345:458–460. doi: 10.1038/345458a0. [DOI] [PubMed] [Google Scholar]
- 19.Li Y, Zhi W, Wareski P, Weng NP. IL-15 activates telomerase and minimizes telomere loss and may preserve the replicative life span of memory CD8+ T cells in vitro. J Immunol. 2005;174:4019–4024. doi: 10.4049/jimmunol.174.7.4019. [DOI] [PubMed] [Google Scholar]
- 20.Kiecolt-Glaser JK, Dura JR, Speicher CE, Trask OJ, Glaser R. Spousal caregivers of dementia victims: longitudinal changes in immunity and health. Psychosom Med. 1991;53:345–362. doi: 10.1097/00006842-199107000-00001. [DOI] [PubMed] [Google Scholar]
- 21.Haddad JJ, Fahlman CS. Redox- and oxidant-mediated regulation of interleukin-10: an anti-inflammatory, antioxidant cytokine? Biochem Biophys Res Commun. 2002;297:163–176. doi: 10.1016/s0006-291x(02)02094-6. [DOI] [PubMed] [Google Scholar]
- 22.Weng NP. Aging of the immune system: how much can the adaptive immune system adapt? Immunity. 2006;24:495–499. doi: 10.1016/j.immuni.2006.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Blasco MA, Lee HW, Hande MP, Samper E, Lansdorp PM, DePinho RA, Greider CW. Telomere shortening and tumor formation by mouse cells lacking telomerase RNA. Cell. 1997;91:25–34. doi: 10.1016/s0092-8674(01)80006-4. [DOI] [PubMed] [Google Scholar]
- 24.Mitchell JR, Wood E, Collins K. A telomerase component is defective in the human disease dyskeratosis congenita. Nature. 1999;402:551–555. doi: 10.1038/990141. [DOI] [PubMed] [Google Scholar]
- 25.Vulliamy T, Marrone A, Goldman F, Dearlove A, Bessler M, Mason PJ, Dokal I. The RNA component of telomerase is mutated in autosomal dominant dyskeratosis congenita. Nature. 2001;413:432–435. doi: 10.1038/35096585. [DOI] [PubMed] [Google Scholar]
- 26.Chiang YJ, Hemann MT, Hathcock KS, Tessarollo L, Feigenbaum L, Hahn WC, Hodes RJ. Expression of telomerase RNA template, but not telomerase reverse transcriptase, is limiting for telomere length maintenance in vivo. Mol Cell Biol. 2004;24:7024–7031. doi: 10.1128/MCB.24.16.7024-7031.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Slagboom PE, Droog S, Boomsma DI. Genetic determination of telomere size in humans: a twin study of three age groups. Am J Hum Genet. 1994;55:876–882. [PMC free article] [PubMed] [Google Scholar]
- 28.Iwama H, Ohyashiki K, Ohyashiki JH, Hayashi S, Yahata N, Ando K, Toyama K, Hoshika A, Takasaki M, Mori M, Shay JW. Telomeric length and telomerase activity vary with age in peripheral blood cells obtained from normal individuals. Hum Genet. 1998;102:397–402. doi: 10.1007/s004390050711. [DOI] [PubMed] [Google Scholar]
- 29.Cong YS, Wright WE, Shay JW. Human telomerase and its regulation. Microbiol Mol Biol Rev. 2002;66:407–425. doi: 10.1128/MMBR.66.3.407-425.2002. table. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Soares MV, Borthwick NJ, Maini MK, Janossy G, Salmon M, Akbar AN. IL-7-dependent extrathymic expansion of CD45RA+ T cells enables preservation of a naive repertoire. J Immunol. 1998;161:5909–5917. [PubMed] [Google Scholar]
- 31.Xu D, Erickson S, Szeps M, Gruber A, Sangfelt O, Einhorn S, Pisa P, Grander D. Interferon α down-regulates telomerase reverse transcriptase and telomerase activity in human malignant and nonmalignant hematopoietic cells. Blood. 2000;96:4313–4318. [PubMed] [Google Scholar]
- 32.Li H, Xu D, Li J, Berndt MC, Liu JP. Transforming growth factor β suppresses human telomerase reverse transcriptase (hTERT) by Smad3 interactions with c-Myc and the hTERT gene. J Biol Chem. 2006;281:25588–25600. doi: 10.1074/jbc.M602381200. [DOI] [PubMed] [Google Scholar]
- 33.Akiyama M, Hideshima T, Hayashi T, Tai YT, Mitsiades CS, MItsiades N, Chauhan D, Richardson P, Munshi NC, Anderson KC. Cytokines modulate telomerase activity in a human multiple myeloma cell line. Cancer Res. 2002;62:3876–3882. [PubMed] [Google Scholar]
- 34.Akiyama M, Yamada O, Hideshima T, Yanagisawa T, Yokoi K, Fujisawa K, Eto Y, Yamada H, Anderson KC. TNFα induces rapid activation and nuclear translocation of telomerase in human lymphocytes. Biochem Biophys Res Commun. 2004;316:528–532. doi: 10.1016/j.bbrc.2004.02.080. [DOI] [PubMed] [Google Scholar]
- 35.Beyne-Rauzy O, Prade-Houdellier N, Demur C, Recher C, Ayel J, Laurent G, Mansat-De VM. Tumor necrosis factor-α inhibits hTERT gene expression in human myeloid normal and leukemic cells. Blood. 2005;106:3200–3205. doi: 10.1182/blood-2005-04-1386. [DOI] [PubMed] [Google Scholar]
- 36.Glaser R, MacCallum RC, Laskowski BF, Malarkey WB, Sheridan JF, Kiecolt-Glaser JK. Evidence for a shift in the Th-1 to Th-2 cytokine response associated with chronic stress and aging. J Gerontol A Biol Sci Med Sci. 2001;56:M477–M482. doi: 10.1093/gerona/56.8.m477. [DOI] [PubMed] [Google Scholar]