

Abstract
I measured the strength of interaction between a marine herbivore and its growing resource over a realistic range of absolute and relative abundances. The herbivores (hermit crabs: Pagurus spp.) have slow and/or weak functional and numerical responses to epiphytic diatoms (Isthmia nervosa), which show logistic growth in the absence of consumers. By isolating this interaction in containers in the field, I mimicked many of the physical and biological variables characteristic of the intertidal while controlling the densities of focal species. The per capita effects of consumers on the population dynamics of their resource (i.e., interaction strength) were defined by using the relationship between hermit crab density and proportional change in the resource. When this relationship is fit by a Weibull function, a single parameter distinguishes constant interaction strength from one that varies as a function of density. Constant interaction strength causes the proportion of diatoms to fall linearly or proportionally as hermit crab density increases whereas per capita effects that increase with density cause an accelerating decline. Although many mathematical models of species interactions assume linear dynamics and invariant parameters, at least near equilibrium, the per capita effects of hermit crabs on diatoms varied substantially, apparently crossing a threshold from weak to strong when consumption exceeded resource production. This threshold separates a domain of coexistence from one of local extinction of the resource. Such thresholds may help explain trophic cascades, resource compensation, and context-dependent interaction strengths, while indicating a way to predict trophic effects, despite nonlinearities, as a function of vital rates.
Keywords: herbivory, alternative states, community ecology, productivity, consumption
To understand the dynamics of complex communities, ecologists have begun to quantify food web links by their interaction strengths (1–7). Interaction strength represents the per capita effect of one species on another’s population growth (8–11) or density (3, 4). The concept of interaction strength is useful to assess functional redundancy (12–14), community stability (9, 15, 16), and indirect effects (10, 11) and provides insight into the arrangement of food webs in terms of many weak and a few strong links (3, 4, 17). Furthermore, to the extent that per capita effects are constant, they can be multiplied by species densities to make predictions about total effects (2).
The assumption that pairs of species have a single constant strength of interaction arises from two sources. First, theory generally considers interactions to occur near system equilibrium, at which effects on population growth rate can be approximated by linear functions involving constant per capita effects (9–11). Second, when interaction strengths are measured empirically, experiments tend to be carried out at one time and place and involve a comparison between cases where a species is removed vs. present at a single “natural” density (3). Consequently, only a single average value is determined for interaction strength.
Yet abundant evidence shows that interactions are not linear and that, instead, interaction strengths vary over a range of observed conditions. For instance, in trophic cascades, consumer dynamics are decoupled from resource dynamics, perhaps because of subsidies from other sources (18, 19). Consumer densities remain sufficiently high that the resource cannot recover, and trophic effects remain strong even though a lower consumer density might have no detectable effect on the resource (20). Similarly, herbivores may have qualitatively different effects on plants at high and low densities. Despite detrimental effects of overgrazing, moderate herbivory may improve plant performance if consumers remove tissue that would otherwise succumb to self-thinning or if they recycle nutrients that promote plant growth (21–25). Pest outbreaks may occur because a population suddenly escapes consumers that previously had been able to keep it in check (26–28). Conversely, if consumers become seasonally abundant, the point at which consumption exceeds resource production has been termed the “catastrophe date” because the resource rapidly disappears (29, 30). Ecologists are increasingly recognizing alternative community “states,” each of which is persistent but marked by distinct controlling interactions (31–34). Phenomenologically, alternative community states imply that effects of a species at one density cannot necessarily be scaled linearly with density to predict effects at another density.
Direct measurements of interaction strength usually are carried out within a limited range of conditions (e.g., densities, food web structures, times of year). Nevertheless, consumer interaction strength often is found to vary with density (4–7, 35, 36). Furthermore, variable per capita effects commonly are observed in competition studies, when frequency and density both influence the competition coefficient (37, 38). These experiments and the nonlinear phenomena described above caution against an assumption of constant per capita interaction strength. If measured interaction strengths are valid only within a particular context, they cannot be used to assess food web global stability or to extrapolate species’ impacts from one situation to another. However, an alternative to case-by-case assessment of interaction strength is to focus on the mechanisms that cause interaction strength to change. This paper presents the first experimental determination of pairwise trophic interaction strength in which the objective is to assess variation as a function of density. A potential mechanism, resulting in variable per capita effects, is the shift in relative magnitudes of the vital rates of the intertidal herbivore and alga involved in the interaction.
METHODS
In the focal pairwise interaction, hermit crabs (Pagurus spp.) feed on a large (0.3-mm) chain-forming diatom (Isthmia nervosa). The diatom is epiphytic on a variety of red algal species, at least one of which (Odonthalia floccosa) suffers from heavy diatom loads and thus may receive indirect benefits from diatom consumers (39). On Tatoosh Island, WA (48°24′N, 124°44′W), these species occupy rocky, low intertidal benches and tidepools that receive some protection from the direct force of western swell (39). Based on censuses from 1992 to 1996, diatoms can occur naturally at densities of 0–140 mg/cm of Odonthalia, and hermit crabs can occur naturally at 0–0.1 cm−1 of Odonthalia.
I studied these interactions in field microcosms—9 × 9 × 12-cm containers that were anchored in tidepools to reduce dislodgement by waves. Constructed of translucent plastic with 1-mm mesh on the top and two sides, the containers reduced light (down 10%) and water flow and required removal of accumulated sediment and periphyton every 2 weeks. However, in other respects, they mimicked tidepool systems: Trials spanned the range of natural densities of Isthmia and Pagurus, and containers also housed microalgae, small crustaceans, worms, limpets, and snails.
Within these containers, I assessed the ability of different densities of hermit crabs to reduce diatom cover. Containers were placed in three tidepools, and various combinations of consumer and resource were initiated on 15 occasions. All trials occurred during four summers (April to September 1993–1996) and ran for 2–4 weeks. The number of containers (=trials) within tidepools varied among dates because of the presence of other treatments not considered here and damage to some containers. ANOVA was used to check for an effect of month or tidepool on the experimental results before assuming that all dates and tidepools were equivalent.
In total, 162 containers included hermit crabs and 188 did not. These latter “control” containers served to determine whether diatoms declined for reasons unrelated to hermit crabs and provided information on population growth rates of diatoms at different initial densities. Also, 98 trials were run in which a piece of epiphytized Odonthalia was placed with an unepiphytized plant to assess the ability of Isthmia to colonize new substrate.
Odonthalia with epiphytic diatoms was mounted on epoxy putty bases and attached to upended screws on the floor of the container. Diatom biomass was estimated from host size (total length of shoots, which were unbranched) and cover score [on a scale of 0 (none) to 6 (maximum), where the relationship between cover score and biomass is known (39)]. A large host plant with a low cover score can have the same diatom biomass as does a small host with higher diatom cover. All hermit crab densities were standardized relative to initial diatom biomass.
Considerable controversy exists concerning the correct way to measure interaction strength (1–6, 9, 16). Instead of choosing among metrics, I examined the outcome of the experiment directly, in terms of proportional change in diatom biomass. Conceptually, it is critical to distinguish interaction strength from the relationship between hermit crab density and final diatom biomass after the 2-week experiment. Constant interaction strength could result in a linear, declining relationship (until diatoms are gone) if consumers feed with constant yield or in a negative exponential relationship if consumers feed with constant effort. Given sampling error, these two sorts of relationships may be difficult to distinguish (although I used the negative exponential as a null model). In contrast, constant interaction strength cannot generate a threshold in the relationship between consumer density and final resource abundance. Specifically, if per capita effects increase with hermit crab density, then fewer diatoms will remain at the end of the experiment than would be expected from a proportional or absolute extrapolation of hermit crab effects.
I described the relationship between hermit crab density (x, normalized for initial diatom biomass) and proportion of diatoms remaining (y) with a Weibull function (40), which can accommodate a threshold. The parameter a (a > 1) defines the steepness of a sigmoid decline in diatoms remaining, and b defines the hermit crab density where the total effect on the resource is changing most rapidly (=threshold):
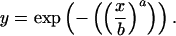 |
1 |
The Weibull function is a negative exponential when a = 1. When a > 1, however, hermit crabs have effects on diatoms that increase faster than hermit crab density. At high values of a, two qualitatively different outcomes emerge: Either diatoms persist at high densities, or they are completely eliminated. Thus, the statistical test for nonconstant interaction strength is to determine how likely it is that a > 1. I developed likelihood profiles for a and b, considering sparse (<45 mg⋅cm− 1 of host) and dense (>70 mg⋅cm− 1 of host) diatoms separately. Likelihoods were calculated as
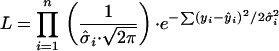 |
2 |
where n is the number of trials, y is the actual proportion of diatoms remaining, and ŷ is the value expected from the Weibull function (41). After negative logarithms are taken on each side of equation (2), the likelihood is seen to be related inversely to the sum of squared deviances divided by variance (lower deviance implies higher likelihood).
Although both variables in the experiment (i.e., normalized hermit crab density and proportion of diatoms remaining after 2 weeks) were measured with error, likelihood methods are well developed only for cases involving error in dependent variables (42, 43). I assumed error only in y, which should be conservative, and estimated the variance directly for each trial. The trials were first ordered by their average rank in hermit crab density and proportion of diatoms remaining. Beginning with the sixth point, and sliding across the data set until reaching the sixth from the end, means and SE of proportion remaining were bootstrapped (44) by using each point and five on either side. The first and last five points were assumed to have the same error associated with them as did the sixth point from its end of the ordered list. Points with no variance (due to the large number of cases in which diatoms either did not change or disappeared altogether) were assumed to have the minimum SE calculated for other points. This bimodality of outcome (i.e., either all or none of the diatoms remained) is itself an indication that total effect on the resource did not change incrementally with hermit crab density. Bootstrapped SEs (σ̂i/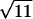 ) then were incorporated into the likelihood function.
) then were incorporated into the likelihood function.
Feeding rates of hermit crabs were measured directly in the laboratory over 1–2 days. Trials occurred in April (n = 16), May (n = 3), and September (n = 8) 1993. Initial biomass of Isthmia (on Odonthalia) ranged over an order of magnitude (100 to >1,000 mg wet weight). Diatoms were blotted dry and weighed before and after being placed with a single hermit crab. To determine feeding rates, biomass loss in containers with hermit crabs was compared with controls lacking consumers. Nineteen controls were available, and most lost mass over 1–2 days. Daily rate of biomass loss increased with initial biomass (loss = −0.00014·initial biomass − 0.00298, adj. r2 = 0.63, F = 32.2, P < 0.0001); the scatter (residuals) around this relationship describes the uncertainty in controls. For Isthmia with hermit crabs, total biomass loss stems from a combination of the expected loss of diatoms not due to hermit crabs (based on the regression), the uncertainty in this loss (based on residuals), and consumption. For each feeding trial, total biomass loss was first adjusted by the regression and then by each of the 19 residuals, giving 19 estimates of feeding rate. Bootstrap samples (200) of n = 19 were chosen randomly with replacement from this set and were used to calculate a mean feeding rate and SE for the hermit crab in that trial.
RESULTS AND DISCUSSION
Per capita effects of hermit crabs on diatom population dynamics were not constant across density, in contrast to expectations based on linear dynamics and invariant parameter values. In the containers used in this experiment, an increase in hermit crab density first had little impact on diatoms but then caused a steep decline in the proportion remaining after 2 weeks (Fig. 1). Fewer hermit crabs were required to eliminate a given diatom biomass when diatoms were distributed densely on a small host than when distributed sparsely on a larger one. Fig. 1 shows maximum likelihood relationships between hermit crab density and proportion of diatoms remaining, modeled as a Weibull function. Fig. 2A shows the likelihood profile as a is varied, assuming b is unfixed (allowed to vary to give the highest likelihood for a particular a). The important result is that a is likely to be >1, which introduces a threshold into the relationship between consumer density and resource remaining. For sparse diatoms, there are two peaks in the likelihood function, but the higher value of a essentially creates a step in the function at a slightly lower threshold hermit crab density. In Fig. 2B, the threshold density b is varied as a is left unfixed. The most likely threshold value when diatoms are dense (b = 0.0028) occurs at a lower hermit crab density than when diatoms are sparse (b = 0.0110).
Figure 1.

Maximum likelihood relationships between hermit crab (Pagurus) density and the proportion of their diatom (Isthmia) resource remaining after 2 weeks in field-deployed containers. The relationship was fit by using a Weibull function (Eq. 1). (A) Diatoms distributed sparsely on host Odonthalia flocossa (<45 mg⋅cm−1). (B) Diatoms distributed densely on their host (>70 mg⋅cm−1). Circled points were considered outliers and were not used for model fitting.
Figure 2.

Likelihood profiles for parameters in the Weibull function used to describe the relationship between hermit crab density and proportion of diatoms remaining. Likelihood ratios normalize all values relative to the maximum likelihood. (A) Threshold steepness (a) when b is unfixed. (B) Threshold hermit crab density (b) when a is unfixed. The cutoff values for 95% confidence intervals around the maximum likelihood occur at a likelihood ratio of 0.15.
In the absence of hermit crabs, diatoms generally maintained or increased their cover. Of 188 control containers, <30% declined in diatom score, and <7% declined more than one score. Average final relative to initial biomass was 0.93 (SE = 0.87–1.00, back-transformed from ln-transformed values). When controls were grouped by year in 2-month intervals to generate sample sizes of >5 per group, there was no effect of date, tidepool, or date × tidepool on the outcome of the trial (ANOVA: F6, 168 = 1.47, P = 0.19; F2, 168 = 2 .05, P = 0.13; F11, 168 = 1.61, P = 0.10).
In the presence of hermit crabs, the outcome of the experiment tended toward one of two states: little change in the resource and coexistence of plant and herbivore or rapid and complete elimination of diatoms. Yet, 19 trials with sparse diatoms (36%) and 49 trials with dense diatoms (45%) showed intermediate diatom cover at the end of 2 weeks. The short duration of the experiment was chosen because, in 28 trials run for 4 weeks, >85% showed no change in outcome over the final 2 weeks. However, it is possible that, in some trials, hermit crabs had insufficient time to drive diatoms to their final state. Another explanation for intermediate outcomes is higher diatom productivity at lower densities, particularly for trials that began with dense diatoms.
This sort of threshold between coexistence and local resource extinction was recognized >2 decades ago (45), and the dynamics go back much further (46, 47). Population abundance is augmented by recruitment and reduced by harvest. Slight increases in recruitment can cause a species to cross an “escape threshold” and shift to a higher population density. Alternatively, a slight decrease in population growth rate (or increase in harvest rate) may cause the population to cross an “extinction threshold,” an unstable equilibrium that leads to a population crash. The salient conditions producing thresholds are: (i) harvest rates exceed population growth rates at low resource densities; and (ii) population growth rates exceed harvest rates at higher resource densities. These conditions require a weak coupling of consumer dynamics to changes in the resource (19, 48).
Hermit crab dynamics are coupled weakly to diatoms, which might be the reason that small changes in the density of the consumer apparently cause qualitatively different outcomes for the resource. Because hermit crab density varies spatially as a function of shell supply (49–51), hermit crabs are unlikely to respond numerically to changes in food resources. Furthermore, hermit crab feeding rates were constant over a wide range of diatom abundances in the laboratory (Fig. 3). The hermit crab functional response rises rapidly and saturates early, which should allow feeding rates to exceed diatom production when diatoms are rare. Thus, per capita effects may vary with hermit crab density because consumers only affect the resource if their consumption exceeds production, at which point, because consumption stays high despite declining resource biomass, diatoms can be eliminated rapidly.
Figure 3.

Feeding rates of Pagurus hirsutiusculus on Isthmia nervosa at different initial biomasses (wet weight). Error bars represent SEs, which were generated from 200 bootstrap samples of the differences in weight change between 19 controls and each hermit crab trial. Based on analysis of covariance, daily feeding rate is higher for large than for medium and small hermit crabs (F = 6.95, P = 0.005), but initial diatom biomass has no effect on feeding rate (F = 2.20, P = 0.15). Open circles, large hermit crabs; x, medium; dark points, small.
Diatom population growth is nonlinear, which may cause per capita effects of hermit crabs at a given density (standardized to diatom biomass) to differ depending on whether diatoms are distributed sparsely or densely. For 136 controls with no decline in diatoms, Isthmia showed logistic growth (Fig. 4), perhaps because sparse diatoms have greater access to light and nutrients that allow high division rates. The most dramatic examples of diatom growth in field-deployed containers came from the 98 trials in which unepiphytized Odonthalia was inoculated with diatoms. Of these, 90 hosted diatoms within 2 weeks. Thus, per capita trophic effects may be weaker on sparse than on dense diatoms because sparse diatoms have higher division rates and so can sustain higher consumption before declining. Two alternative states arise for diatoms, in which they may be limited by consumers at low diatom abundances and self-limited when higher.
Figure 4.

Productivity of Isthmia nervosa inside containers (mg⋅d−1 on each cm of host Odonthalia; mean ± SE). Sample sizes are presented above each mean value. Although the graph excludes trials in which diatoms declined, incorporating those trials simply shifts the relationship between diatom biomass and productivity lower, rather than changing its shape. The second order regression using mean productivity is significant (y = −0.042 + 0.038x − 2.646·10−4x2; F = 8.725, P = 0.0348, n = 7). Division rates (r) also were measured in the laboratory (October 8–14, 1994, 16 h light:8 h dark, 10–15°C, in autoclaved seawater with added Provasoli’s Enrichment Solution and metasilicate). This (maximum) division rate is shown by the dashed line: r = 0.075 div⋅d− 1 ± 0.0079 SE, n = 33.
The interaction between Pagurus and Isthmia is characterized by the absence of an equilibrium (because hermit crab densities do not depend on diatoms) and by the presence of nonlinearities because diatoms undergo intraspecific competition. These are precisely the conditions that violate assumptions of local stability analyses and should cause constant interaction strength to fail.
Based on cases cited in the introduction, it seems clear that the marine intertidal is not unusual in harboring interactions marked by thresholds between alternative states. This scenario is likely to be common in a whole range of systems where generalist consumers are decoupled from the abundance of a specific resource. And even under these conditions, we require some means by which to quantify and communicate the strength of biological interactions. Is the only option, then, to define interaction strength so that it applies within a restricted context? I would suggest, instead, that we focus on the boundaries of potential trophic effects. These boundaries are set in two ways. At one end, maximum per capita impact follows from rapid removal of a nongrowing resource and therefore should be related to feeding and encounter rates. Alternative resources and consumer interference can reduce feeding and therefore interaction strength. Per capita effects also may decline as a resource is depleted, either because consumer densities are high and the total effect must be divided among them or because results are recorded after long intervals and the total effect must be divided over this time. At the other end of the spectrum, minimum total and per capita effects arise when the resource can compensate for biomass loss, that is, at high resource productivity. Only if the ratio of consumption rate to production rate exceeds 1.0 will consumers be able to reduce their resource. Otherwise, the resource should grow and persist at the same or higher level, and trophic effects will be minimized. These minimal trophic effects will be most common when resource population growth is high, which can be determined, for example, by λ in matrix models (52). For a resource such as Isthmia, maximum productivity is simple to define as the division rate of diatoms on a known amount of substrate.
Density-dependent interaction strengths introduced by nonequilibrium, nonlinear dynamics actually may reinforce shifts in complex communities over space and time. A small change in the abundance of one species may result in qualitative shifts in another. For hermit crabs and diatoms, a decoupling of consumer from resource dynamics and self-limitation of the resource apparently create a threshold in the trophic interaction over realistic relative abundances. In fact, density-dependent interaction strength can arise in numerous ways, many of which generate thresholds (31, 45, 53). The assumption of constant interaction strength fails because, even when a description of links in a food web applies widely, considerable local variation in species’ densities may exist. The challenge now lies in discerning the mechanisms determining interaction strength so that trophic dynamics become more predictable. Central to these predictions are the relative magnitudes of resource productivity and rates of consumption.
Acknowledgments
I thank the Makah Indian Nation and US Coast Guard for allowing work on Tatoosh Island. While examining thresholds, I benefited greatly from discussions with D. T. Haydon and from inspiration lingering in C. S. Holling’s old University of British Columbia office. This paper is based on material submitted in partial fulfillment of a Ph.D. at the University of Washington, Seattle. Funding for this research was provided through a National Science Foundation predoctoral fellowship, the ARCS Foundation, and National Science Foundation Grant OCE91–15760 to R. T. Paine.
References
- 1.Laska M S, Wootton J T. Ecology. 1998;79:461–476. [Google Scholar]
- 2.Wootton J T. Ecol Monogr. 1997;67:45–64. [Google Scholar]
- 3.Paine R T. Nature (London) 1992;355:73–75. [Google Scholar]
- 4.Navarrete S A, Menge B A. Ecol Monogr. 1996;66:409–429. [Google Scholar]
- 5.Menge B A, Daley B, Wheeler P A. In: Food Webs. Polis G A, Winemiller K O, editors. New York: Chapman & Hall; 1996. pp. 258–274. [Google Scholar]
- 6.Menge B A, Berlow E L, Blanchette C A, Navarrete S A, Yamada S B. Ecol Monogr. 1994;64:249–286. [Google Scholar]
- 7.Fagan W F, Hurd L E. Ecology. 1994;75:2022–2032. [Google Scholar]
- 8.MacArthur R. Trans Conn Acad Arts Sci. 1972;44:177–188. [Google Scholar]
- 9.May R M. Stability and Complexity in Model Ecosystems. Princeton, NJ: Princeton Univ. Press; 1973. [PubMed] [Google Scholar]
- 10.Yodzis P. Ecology. 1988;69:508–515. [Google Scholar]
- 11.Bender E A, Case T J, Gilpin M E. Ecology. 1984;65:1–13. [Google Scholar]
- 12.Steneck R S, Dethier M N. Oikos. 1994;69:476–495. [Google Scholar]
- 13.Walker B H. Cons Biol. 1992;6:18–23. [Google Scholar]
- 14.Lawton J H, Brown V K. In: Biodiversity and Ecosystem Function. Shulze E-D, Mooney H A, editors. Berlin: Springer; 1993. pp. 255–270. [Google Scholar]
- 15.Moore J C, de Ruiter P C, Hunt H W. Science. 1993;261:906–908. doi: 10.1126/science.261.5123.906. [DOI] [PubMed] [Google Scholar]
- 16.de Ruiter P C, Neutel A-M, Moore J C. Science. 1995;269:1257–1260. doi: 10.1126/science.269.5228.1257. [DOI] [PubMed] [Google Scholar]
- 17.Cornell H V, Lawton J H. J Anim Ecol. 1992;61:1–12. [Google Scholar]
- 18.Bustamante R H, Branch G M, Eekhout S. Ecology. 1995;76:2314–2329. [Google Scholar]
- 19.Polis G A, Hurd S D. In: Food Webs. Polis G A, Winemiller K O, editors. New York: Chapman & Hall; 1996. pp. 275–285. [Google Scholar]
- 20.Paine R T. J Anim Ecol. 1980;49:667–685. [Google Scholar]
- 21.Hik D S, Jeffries R L. J Ecol. 1990;78:180–195. [Google Scholar]
- 22.McNaughton S J, Milchunas D G, Frank D A. Ecology. 1996;77:974–977. [Google Scholar]
- 23.Sterner R W. Science. 1986;231:605–607. doi: 10.1126/science.231.4738.605. [DOI] [PubMed] [Google Scholar]
- 24.Williams S L, Carpenter R C. J Phycol. 1990;26:36–40. [Google Scholar]
- 25.Bergquist A M, Carpenter S R. Ecology. 1986;67:1351–1360. [Google Scholar]
- 26.Hanski I, Otronen M. Oikos. 1985;44:165–174. [Google Scholar]
- 27.Royama T. Ecol Monogr. 1984;54:429–462. [Google Scholar]
- 28.Wallner W E. Annu Rev Entomol. 1987;32:317–340. [Google Scholar]
- 29.Hairston N G, Munns W R. Am Nat. 1984;123:733–751. [Google Scholar]
- 30.Burkhardt S, Lehman J T. Limnol Oceanogr. 1994;39:1007–1019. [Google Scholar]
- 31.Knowlton N. Am Zool. 1992;32:674–682. [Google Scholar]
- 32.Sutherland J P. Am Nat. 1974;108:859–873. [Google Scholar]
- 33.Sutherland J P. Am Nat. 1990;136:270–275. [Google Scholar]
- 34.van de Koppel J, Rietkerk M, Weissing F J. Tr Ecol Evol. 1997;12:352–356. doi: 10.1016/s0169-5347(97)01133-6. [DOI] [PubMed] [Google Scholar]
- 35.Berlow E L, Navarrette S A. J Exp Mar Biol Ecol. 1997;214:195–229. [Google Scholar]
- 36.Berlow E L. Ecol Monogr. 1997;67:435–460. [Google Scholar]
- 37.Antonovics J, Kareiva P. Proc R Soc Lond B. 1988;319:601–613. doi: 10.1098/rstb.1988.0068. [DOI] [PubMed] [Google Scholar]
- 38.Pfister C A. Am Nat. 1995;146:271–291. [Google Scholar]
- 39.Ruesink J L. J Phycol. 1998;34:29–38. [Google Scholar]
- 40.Hastings N A J, Peacock J B. Statistical Distributions. London: Butterworths; 1975. [Google Scholar]
- 41.Hilborn R, Mangel M. The Ecological Detective. Princeton, NJ: Princeton Univ. Press; 1997. [Google Scholar]
- 42.Pascual M A, Kareiva P. Ecology. 1996;77:337–349. [Google Scholar]
- 43.Walters C J, Ludwig D. Can J Fish Aq Sci. 1981;38:704–710. [Google Scholar]
- 44.Efron B, Tibshirani R. Science. 1991;253:390–395. doi: 10.1126/science.253.5018.390. [DOI] [PubMed] [Google Scholar]
- 45.Holling C S. Annu Rev Ecol Syst. 1973;4:1–23. [Google Scholar]
- 46.Beverton R J H, Holt S J. On the Dynamics of Exploited Fish Populations. London: H. M. Stationery Office; 1957. [Google Scholar]
- 47.Ricker W E. J Fish Res Bd Can. 1954;11:559–623. [Google Scholar]
- 48.Polis G A, Holt R D, Menge B A, Winemiller K O. In: Food Webs. Polis G A, Winemiller K O, editors. New York: Chapman & Hall; 1996. pp. 435–460. [Google Scholar]
- 49.Abrams P A. Oecologia. 1987;72:233–247. doi: 10.1007/BF00379274. [DOI] [PubMed] [Google Scholar]
- 50.Abrams P A. Oecologia. 1987;72:248–258. doi: 10.1007/BF00379275. [DOI] [PubMed] [Google Scholar]
- 51.Vance R R. Ecology. 1972;53:1062–1074. [Google Scholar]
- 52.Caswell H. Matrix Population Models. Sunderland, MA: Sinauer; 1989. [Google Scholar]
- 53.Dyer M I. J Appl Ecol. 1975;12:719–726. [Google Scholar]