

Abstract
Following transplantation of green fluorescent protein (GFP)-labeled bone marrow (BM) into irradiated, wild-type Sprague-Dawley rats, propagated GFP+ cells migrate to adipose tissue compartments. In order to determine the relationship between GFP+ BM-derived cells and tissue resident GFP− cells on the stem cell population of adipose tissue, we conducted detailed immunohistochemical analysis of chimeric whole fat compartments and subsequently isolated and characterized adipose-derived stem cells (ASCs) from GFP+ BM chimeras. In immunohistochemistry, a large fraction of GFP+ cells in adipose tissue were strongly positive for CD45 and smooth muscle actin, and evenly scattered around the adipocytes and blood vessels, while all CD45+ cells within the blood vessels were GFP+. A small fraction of GFP+ cells with mesenchymal marker CD90 also existed in the perivascular area. Flow cytometric and immunocytochemical analyses showed that cultured ASCs were CD45−/CD90+/CD29+. There was a significant difference in both the cell number and phenotype of the GFP+ ASCs in two different adipose compartments, the omental (abdominal) and the inguinal fat (subcutaneous) pad; a significantly higher number of GFP−/CD90+ cells were isolated from the subcutaneous depot as compared to the abdominal depot. The in vitro adipogenic differentiation of the ASCs was achieved; however, all cells that had differentiated were GFP−. Based on phenotypical analysis, GFP+ cells in adipose tissue in this rat model appear to be of both hematopoietic and mesenchymal origin; however, infrequent isolation of GFP+ ASCs and their lack of adipogenic differentiation suggest that the contribution of BM to ASCs generation might be minor.
Keywords: Adipose, chimera, stem cell, GFP
INTRODUCTION
The distribution of cells within adipose tissue has been investigated, with the primary cell type present in adipose tissue being adipocytes. However, as fat is a vascularized tissue, other cell types that reside in the adipose tissue compartment include endothelial cells, macrophages, smooth muscle cells, and fibroblasts. The determination of a population of cells other than adipocytes was first identified in the mid 1970’s. In 1975, Stiles et al documented a morphologically different phenotype of cells in human adipose tissue [1]. In 1976, Dardick et al determined that these cells can differentiate into adipocytes [2]. For the next two decades, several groups began studying these cells, termed ‘preadipocytes’ [3–9]. It was in 2001 when the plasticity of these cells were published by Zuk et al [10]. Initial reports demonstrated the mesenchymal plasticity of the cells, followed by several reports on the transdifferentiation of these seemingly mesenchymal stem cells into both ectodermal and endodermal phenotypes [11–16]. The identification of a stem cell population in adipose tissue has lead to an extensive and rapid growth of research in the fields of regenerative medicine and adipose biology.
While the plasticity and potential of the stem cells derived from the adipose tissue compartment remain under intense scrutiny, the origin of the stem cells is unclear. One possibility is that the cells are derived from the bone marrow. Recently, Crossno et al reported that progenitor cells originating from the bone marrow can contribute to an increase in adipocyte number [17]. Their in vivo mouse study demonstrated that exposure to a high-fat diet or treatment with the thiazolidinedione rosiglitazone for three weeks promoted the trafficking of circulating bone marrow-derived progenitor cells into adipose tissue. This was evidenced by the appearance of green fluorescent protein (GFP)+ multilocular adipocytes.
In order to test the hypothesis that adipose-derived stem cells (ASCs) are derived from the bone marrow, we identified a population of GFP positive as well as GFP− cells from the adipose compartments of BM chimeric rats. There was migration of the GFP+ cells to two different adipose compartments (the omental and the inguinal fat pad), as identified by flow cytometry and immunohistochemistry. In addition to whole adipose tissue analysis, ASCs were isolated from the two different adipose compartments and cultured to passage three. Our results indicate that GFP+ cells in adipose tissue in this rat model appear to be of both hematopoietic and mesenchymal origin; however, infrequent isolation of GFP+ ASCs and their lack of adipogenic differentiation suggest that the contribution of BM to ASCs generation may be a rare event.
Materials and Methods
In vivo model
GFP-transgenic and Wild Type (WT) Sprague-Dawley (SD) rats were obtained from Japan SLC, Inc. (Hamamatsu, Japan). The expression of GFP was under the control of the cytomegalovirus enhancer and the chicken β-actin promoter derived from an expression vector, pCAGGS [18,19]. Animals were maintained in laminar flow cages in a specific pathogen-free animal facility at the University of Pittsburgh. All procedures in this experiment were performed according to the guidelines of the Council on Animal Care at the University of Pittsburgh and the National Research Council’s Guide for the Humane Care and Use of Laboratory Animals.
Bone marrow cells (BMCs) were obtained from 8–12 week old GFP rats by flushing the tibias and femurs with RPMI 1640 (Invitrogen, Carlsbad, CA), supplemented with 25 mM HEPES buffer, 2 mM L-glutamine, and 10 μg/ml gentamicin (all from Life Technologies, Grand Island, NY). Flow cytomeric analysis showed that 69.7 ± 5.1% of GFP BMCs were GFP-positive, while ~30% did not express GFP. Similar observation was reported in the BMCs from the GFP mouse [20]. Furthermore, the majority of GFP+ BMCs were CD45+, while the GFP− BMCs were CD45−.
Unfractionated BMCs (2 ×108 cells/animal) with >95% viability in trypan blue exclusion test were intravenously injected into WT rat recipients after 9.5 Gy whole body irradiation (137Cs source). Tacrolimus (0.5 mg/kg/day, a gift from Astellas Pharma Inc., Tokyo, Japan) was given for 7 days after BMC infusion. Successful creation of GFP chimera was confirmed by detecting GFP+ peripheral mononuclear blood cells (PBMCs) with flow cytometry in the blood. The percentage of GFP+ PBMCs in WT recipients quickly increased and reached >95% by 100 days [21]. At 116 to 402 days after transplantation, subcutaneous and intra-abdominal fat tissues were obtained from 26 GFP radiation chimeras and used in this study.
Cell characterization using immunofluorescence microscopy
For immunocytochemistry analysis, cultured ASCs were fixed and processed for immunofluorescence as described [22] using the antibodies listed in Table 1. For immunohistochemistry, adipose tissue was removed from GFP+ bone marrow chimeric rats and immersion fixed in 2% paraformaldehyde in PBS for several hours. Fixed tissue was stored in PBS at 4°C until processed. Tissue was then cut into approximately 1 mm3 pieces and stained in situ. Tissue was mixed on a Nutator (Becton-Dickinson, Franklin Lakes, NJ) during all procedures and rinses were 15 minutes each. Tissue pieces were rinsed three times in PBS, rinsed three times in PBS containing 0.5% BSA, 0.15% glycine (PBG Buffer). Tissue was permeabilized in PBG buffer containing 0.1% Triton X-100 (PBG) for 2 hours at room temperature, then blocked in 5% non-immune goat serum in PBGT buffer (PBG with 0.05% Triton X-100) for 2 hours at room temperature. Primary antibodies (see Table 1) diluted in PBGT buffer were added to tissue overnight at 4°C. Tissue was rinsed 4 times in PBGT buffer then fluorescently tagged secondary and phalloidin (Table 1), diluted in PBGT buffer, were added to the sections overnight at 4°C. Tissue was rinsed three times in PBGT buffer, three times in PBS, then nuclei were stained using 0.01% Hoechst dye (bis Benzimide) in PBS for 10 minutes. Following a wash in PBS, tissue was coverslipped using gelvatol (23 g poly(vinyl alcohol) 2000, 50 ml glycerol, 0.1% sodium azide to 100 ml PBS) and viewed on a Fluoview 1000 confocal microscope (Olympus, Center Valley, PA). Confocal stacks were processed using Imaris (Bitplane, St. Paul, MN) image analysis software.
Table 1.
Antibodies and counterstains used in this study
| Cells | Epitope | 1° Antibody, Clone, Source, Dilution | 2° Antibody, Source, Dilution |
|---|---|---|---|
| Endothelial Cells | CD31/Platelet-Endothelial Cell Adhesion Molecule | IF: Mouse anti-rat CD31 (Serotec) 1:100 FACS: Mouse anti-rat CD31(Chemicon) 1:100 | IF: Goat anti-mouse Cy3(Jackson ImmunoLabs) 1:1000; FACS: Goat anti- mouse PE (Caltag) 1:100 |
| Endothelial Cells/BM Stromal Cells | CD106/VCAM | IF & FACS: PE-conjugated Mouse anti- rat CD106 | IF: Goat anti-mouse Cy3(Jackson ImmunoLabs) 1:1000 |
| Mesenchymal Stem Cells | CD29/Beta-1 Integrin | IF& FACS: Biotin-conjugated CD29 (BD PharMingen) 1:100 | IF: Streptaviden Cy3(Jackson ImmunoLabs) 1:1000; FACS: Streptaviden PE (Cedarlane) 1:500 |
| Mesenchymal Stem Cells | CD90/Thy-1 | IF & FACS: PE-conjugated CD90 (BD PharMingen) 1:100 | IF: Streptaviden Cy3 (Jackson ImmunoLabs) 1:1000 |
| Endothelial Cell Precursors/Mesenchymal Stem Cells | CD133 | IF & FACS: Rabbit anti-CD133(GeneTex, Inc.) 1:100 | IF: Goat ant-rabbit Cy3(Jackson ImmunoLabs) 1:1000; FACS: Mouse ant- rabbit PE (SouthernBiotech) 1:100 |
| Leukocytes/Hematopoietic cells | CD45/Commo n Leukocyte Antigen | IF: Mouse anti-rat CD45, OX1 (Serotec) 1:100 FACS: Mouse anti-rat CD45 conjugated to PE, (BDPharMingen) 1:100 | IF: Goat anti-mouse Cy3 or Cy5 (Jackson ImmunoLabs) 1:1000 |
| Smooth Muscle Cells/pericytes | Smooth Muscle Actin | IF: Mouse anti-SMA conjugated to Cy3(Sigma) 1:250. | |
| GFP | Green Fluorescent Protein | TEM ImmunoGold: rabbit anti-GFP(Abcam) 1:100 | TEM ImmunoGold: Goat anti-rabbit 5 nm gold(Amersham) 1:25. |
| Counterstain | Nucleus | Hoechst 33258: Bis-benzimidizole(Sigma) 10 μg/ml | |
| Counterstain | F-actin | Phalloidin-Alexa 647 or Rhodamine- Phalloidin (Invitrogen) 1:250 |
IF: Immunofluorescence; FACS: Fluorescence activated cell sorting.
Vendor Source Addresses: Abcam: Cambridge, MA; Amersham: Piscataway, NJ; BD PharMingen, San Diego, CA; Caltag Laboratories: Burlingame, CA; Cedarlane: Ontario, Canada; Chemicon international, Temecula, CA; GeneTex: San Antonio, TX; Invitrogen, Carlesbad CA; Jackson ImmunoLabs, West Grove, PA; Serotec, Raleigh, NC; Sigma: St. Louis, MO; SouthernBiotech: Birmingham, AL
Cell characterization using electron microscopy
Adipose tissue pieces (1 mm3) were immersion fixed in 2.5% glutaraldehyde in PBS overnight, then processed as described [23]. For immunoelectron microscopy, tissue pieces were fixed in 2% paraformaldehyde in PBS for 2 hours then dehydrated and embedded in gelatin capsules with LRWhite acrylic resin (SPI Supplies, West Chester, PA) and cured at 60°C following the manufacturer’s directions. Ultrathin sections (70 nm) were labeled with anti-GFP antibody (Table 1) as described [22]. Sections were viewed on JEM 1210 or 1011 transmission electron microscope (JEOL, Peabody, MA).
Isolation and culture of rat adipose-derived stem cells (ASCs)
Adipose tissue was excised from both the omentum (i.e., abdominal) and the inguinal fat pad (i.e., subcutaneous) of naïve (e.g., non-chimeric, GFP+) and chimeric rats, and underwent enzymatic digestion by 0.075% collagenase II (Worthington Biochemical Corp., Lakewood, NJ) in Hank’s Balanced Salt Solution for 60 minutes at 37°C with shaking. Digested tissue was filtered and centrifuged, and erythrocytes were removed by treatment with erythrocyte lysis buffer. The remaining cells were transferred to tissue culture flasks with Dulbecco Modified Eagle Medium (DMEM; Gibco/Invitrogen Corp., Grand Island, NY) plus supplement F12 (Gibco/Invitrogen), and, after an attachment period of six hours, non-adherent cells were removed by a phosphate buffered solution (PBS; Gibco/Invitrogen) wash. Attached cells were cultured in DMEM/F12 media, supplemented with 10% fetal bovine serum (FBS; Gibco/Invitrogen), 0.1 μM dexamethasone (Sigma-Aldrich, Co., St. Louis, MO), 1% penicillin-streptomycin (Gibco/Invitrogen), and 1.25 mg/L amphotericin B (Gibco/Invitrogen), and expanded in vitro until passage three.
Adipocytic differentiation and histochemical assays
ASCs (passage three) were seeded into six-well tissue culture plates (5×105 cells/well) and grown until subconfluence (1–2 days) in DMEM/F12. Medium was then replaced with adipogenic or control media. Adipogenic medium (AM) consisted of DMEM/F12, supplemented with 33 μM Biotin, 0.5 μM insulin, 17 μM D-Pantothenic Acid, 0.2 nM dexamethasone, 1 μM ciglitazone, 0.2 nM T3, and 10 mg/L transferrin. Control medium (CM) was identical to the DMEM/F12 used during the expansion stage. After cells were grown to 100% confluency, adipogenic induction media with IBMX (540 μM) was added for 2 days. Then for 12 days, the media was changed every 48 hrs with adipogenic media. Cells were washed with PBS w/EDTA twice, and fixed with 10% buffered formalin for 10 minutes. Cells were then washed with distilled H2O twice, and stained with Oil Red O (20 mL of stock solution consisting of 30 mg Oil Red O powder in 60 ml 2-propanol (0.5%). Next, 13.3 mL H2O was added and the cells were incubated for 30 minutes at room temperature. Cells were washed with H2O to remove debris. The resultant positive red stain was evaluated via light microscopy.
Cell surface expression using flow cytometry
Cultured ASCs (passage three) were analyzed by flow cytometry for their surface marker expression. Antibodies used in this study are listed in Table 1. For flow cytometry, cultured ASCs were washed and incubated with monoclonal antibodies at 4°C for 30 minutes. After 3 washes with PBS, ASCs were further incubated for 30 minutes with secondary antibodies as needed. Stained cells were fixed in 1% paraformaldehyde and analyzed on a LSR II (BD Biosciences, San Jose, CA), and data were analyzed using FACSDiva software (BD Biosciences). Isotype-matched non-specific antibodies were used for the control.
Statistical Analysis
Unless otherwise specified, the results are reported as mean ± standard deviation. T-tests were conducted to assess differences among treatment groups. Statistical significance was set at a p-value less than or equal to 0.05.
Results
Immunohistochemistry of Rat Adipose Tissue
Whole fat tissue from the chimera was assessed for the markers described in Table 1 using immunofluorescence microscopy. Results indicate GFP+ cells were evenly scattered around the 70–100 μm adipocytes (Figure 1a). Most GFP+ cells had a mesenchymal morphology and were observed to be smooth muscle actin (SMA) positive. The SMA signal in the GFP+ cells was equivalent to that observed in the pericytes surrounding the blood vessels. In Figure 1b, partial confocal stack showed a GFP+ adipocyte in the BM chimera. The GFP+ cytoplasmic labeling of the positive adipocyte surrounds the lipidic portion of the cell. This is a rare event and was observed only once in 6 different chimeric animals examined. GFP+ blood cells were also identified within the F-actin+ blood vessels (Figure 1c). Transmission electron microscopy revealed that the typical GFP+ cell within the adipose tissue integrated between adipocytes, possessed large quantities of rough endoplasmic reticulum (Figure 1d). Immuno-TEM analysis of LRWhite acrylic embedded chimeric adipose tissue indicated that these cells were GFP+ when stained for the GFP protein (Figure 1e).
Figure 1. Evaluation of the distribution and ultrastructure of bone marrow-derived GFP+ cells within the fat of radiation chimera rat 165 days following bone marrow transplantation.
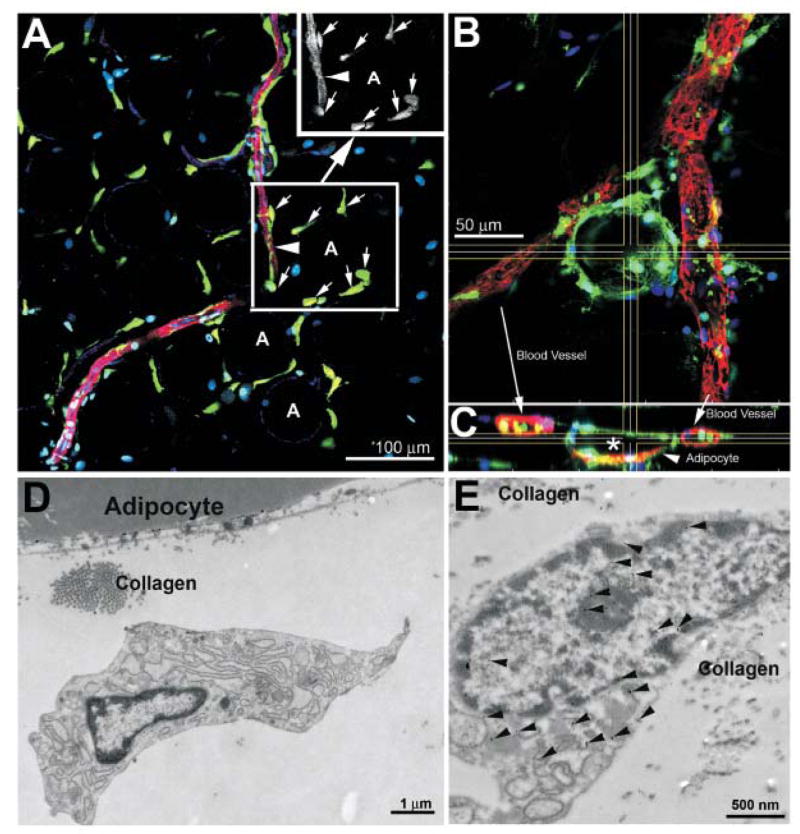
Panel 1A. Tissue was stained for SMA (red), F-actin (blue) and the nucleus (Hoechst’s dye, cyan) in addition to collecting the endogenous fluorescence of GFP (green). Single slice confocal of the tissue shows GFP+ cells were evenly scattered around the 70–100 μm adipocytes (some labeled A as reference). Most GFP+ cells were observed to be SMA positive (arrows) as more clearly shown in the black and white rendition of the red channel in the Panel A insert. The SMA signal in the GFP+ cells was equal to that observed in the pericytes surrounding the blood vessels (arrowhead).
Panel 1B. Partial confocal stack showing a GFP+ adipocyte in the BM chimera. This is a rare event and was observed only once in 6 chimeric animals examined (tissue counterstained for F-actin (rhodamine phalloidin, red) and nucleus (Hoechst’s dye, blue). GFP signal is cytoplasmic and does not partition into the lipidic inclusion of the adipocyte.
Panel 1C. Expanded XY projection from Panel B at axes delineated by the horizonal and vertical lines, indicating the GFP+ blood cells within the F-actin+ blood vessels (arrows). The GFP+ cytoplasmic labeling of the positive adipocyte surrounds the lipidic portion of the cell (*).
Panel 1D. Transmission electron micrograph of the typical GFP+ cell within the adipose tissue, showing these cells, integrated between adipocytes, possess large quantities of rough endoplasmic reticulum.
Panel 1E. Immuno-TEM analysis of LRWhite acrylic embedded chimeric adipose tissue indicated that these cells were GFP+ when stained for the GFP protein (arrows indicate secondary antibody 5 nm gold particles).
Bone Marrow-derived ASCs in Radiation Chimera
Whole adipose tissue from chimeric rats was stained for CD90 and counterstained F-actin (Figure 2). Areas within the tissue show variation in colocalization of CD90 with the GFP BM marker; both GFP−/CD90+ cells. The lower bottom panel in Figure 2 indicates black and white rendition of red channel. The right top panel in Figure 2 shows an area without apparent colocalization of CD90 and GFP signal.
Figure 2.

Adipose tissue from GFP+ chimeric rats was stained for CD90 (Cy3, red) and counterstained F-actin (blue). Areas within the tissue show variation in colocalization of CD90 with the GFP BM marker. Left top panel, arrows indicate dual signal, arrowhead indicates GFP−/CD90+ cell. Lower bottom panel indicates black and white rendition of red channel. Right top panel shows an area without apparent colocalization of CD90 and GFP signal.
Additionally, the whole adipose tissue from GFP+ chimeric rats was stained for CD45 and counterstained for F-actin (Figure 3). It is evident that the majority of cells are both GFP+/CD45+, while a much smaller number of cells are GFP+/CD45−. In Figure 3, it is also evident that there are a few GFP−/CD45+ cells in this field.
Figure 3.

Adipose tissue from GFP+ chimeric rats was stained for CD45 (Cy5, blue) and counterstained F-actin (red). Left top panel shows single optical slice confocal image and right panel shows the black and white rendition of the Cy5 channel, accentuating the CD45+ cells. Arrows indicate the cells that are both GFP+/CD45+, arrowheads indicate the cells that are GFP+/CD45−. In this panel there are also a few GFP−/CD45+ cells in this field with this phenotype. The bottom panels are higher magnification of the top panels.
Adipose-derived Stem Cell Isolation
Collagenase digestion of rat adipose tissue yielded three morphologically distinct populations of cells after culturing until passage three. One population demonstrated a fibroblast-like morphology, similar to ASCs described by Zuk et al [10]. These cells exhibited rapid in vitro expansion. A second, smaller population was GFP+. A third population resembled endothelial cells. Figure 4a depicts the three populations in a light micrograph, and Figure 4b is the corresponding GFP+ image, with only one population of the cells labeled positive for GFP.
Figure 4.

a) Light micrograph of isolated cells from adipose tissue at passage 3, b) corresponding image with GFP+, c) Light micrograph of adipose stem cells that had undergone adipogenesis, and d) corresponding image with GFP+.
Oil Red O Staining
Cells cultured in AM exhibited lipid inclusion after 8 days. Figure 4c depicts a representative positive Oil Red O lipid stain of ASCs cultured in AM for 2 weeks, and Figure 4d depicts the corresponding GFP+ image. No cells that had differentiated into adipocytes were GFP+.
Surface Marker Expression of ASCs
ASCs were isolated and cultured from subcutaneous and abdominal fat tissues obtained from GFP+ radiation chimeras at 116–402 days after bone marrow transplantation. Surface marker expression of cultured ASCs was analyzed by flow cytometry. In the naïve (e.g, GFP+, non-chimera) rats, nearly all ASCs obtained from subcutaneous and abdominal fat tissues of naïve animals were CD45− and CD29+. The majority of the ASCs also expressed CD90 but they did not express CD31, CD106, or CD133 (Figure 5a). Subsequently, CD90 and CD29 were used to determine the origin of ASCs in experiments of GFP radiation chimera. Mitchell et al demonstrated that ASCs are positive for both CD90 and CD29, with expression increasing to >90% at passage 4 [24]. The findings were confirmed with immunocytochemistry of ASCs (Figure 5b).
Figure 5.
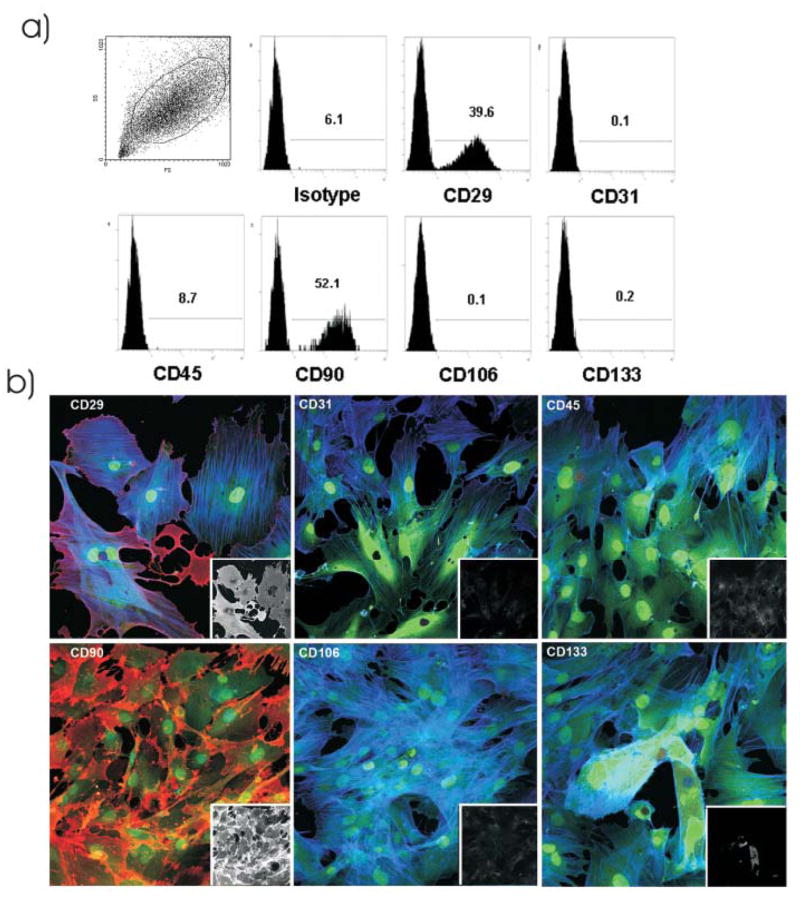
Surface markers expression of cultured, passage three ASCs from naïve GFP+ rats. a) Representative plots of ASCs obtained by culturing the subcutaneous and abdominal fat tissues obtained from naïve animals. Flow cytometric analysis showed that cultured ASCs expressed CD29 and CD90. b) Cells expressed CD29 and CD90 (red) in vitro, while the majority of these cells were negative for CD31, CD45, CD106 and CD133. Blue is F-actin stain and green is endogenous GFP signal. Inserts are the black and white rendition of the red channel for each of these markers.
In the GFP chimeras, flow cytometry showed different levels of CD90 expression depending on the origin of the fat; although ASCs derived from both subcutaneous and abdominal fat tissues expressed CD29, ASCs from the abdominal fat showed significantly lower levels of CD90 expression than those from the subcutaneous fat (Figure 6a). The majority of cultured ASCs obtained from GFP chimeras were GFP negative; however, there was a small population (~5%) of GFP expressing CD90+ and/or CD29+ ASCs (Figure 6b). Interestingly, cultured GFP+ ASCs tended to express low levels of CD90 and CD29, compared to GFP− ASCs. The finding was not different among fat tissues obtained from GFP chimeras at 116–402 days of bone marrow transplantation.
Figure 6.

Surface markers expression of cultured, passage three ASCs from GFP+ radiation chimera. a) ASCs obtained from fat tissues of radiation chimera were analyzed by flow cytometry. CD29 and/or CD90 expressing ASCs were mostly GFP negative; however, small numbers of GFP positive bone marrow-derived ASC were detected. Of note, ASCs from the abdominal fat and subcutaneous fat differ in the expression of CD90. Data is mean ± SD of 3 experiments, * = p ≤ 0.05. b) Representative scattergrams showing GFP+ CD29+/CD90+ ASCs. The intensity of the expression of CD29 and CD90 on GFP+ ASCs is lower than that expressed on GFP− ASCs. The abdominal and subcutaneous fats were obtained from radiation chimera at 161 days after bone marrow transplantation.
Discussion
The mesenchymal stem cell population from adipose tissue has recently been examined in great detail, yet the origin of adipose-derived stem cells is currently unknown. Adipose-derived adult stem cells (ASCs) are easy to obtain and have demonstrated plasticity in differentiating to cells of mesenchymal lineage [11,25–30]. Cells derived from discarded human fat have many properties that make them ideal for tissue engineering applications, such as immunocompatibility, multi-potency, abundancy, amenable to gene therapy, and capability of self-renewal. For example, Hedrick’s group has reported that processed lipoaspirate contains a heterogeneous population of cells that exhibit a fibroblast-like morphology and have the capacity to differentiate into adipocytes, osteoblasts, and chondrocytes [25]. Gimble and Guilak have also reported the differentiation potential of ASCs [26]. Others have shown the potential of ASCs to differentiate into adipocytes, [25] osteoblasts, [27] chondrocytes, [28] and cardiomyocytes [29].
The migration of stem cells after transplantation has been studied previously, including ten Hove et al, who determined that donor-derived cells (from blood) were found in liver tissue specimens after allogeneic stem cell transplantation in nine female patients [31]. Migration of stem cells into the liver had previously been shown to be a rare event. For example, in a study of sex-mismatched liver transplant recipients, Ng et al reported that while recipient cells constituted up to 50% of all cells in the liver allograft, most cells demonstrated macrophage/Kupffer cell differentiation, with only 1.6% showing hepatocytic differentiation [32]. In 2002, Körbling et al reported that recipients of peripheral blood stem cells showed donor-derived hepatocytes up to 7% in liver tissue specimens endothelium by bone-marrow-derived cells [33]. Recently, Nakao et al reported that infused bone marrow stem cells engraft into both the allogeneic marrow environment and syngeneic extra-marrow environment (intestinal graft) [34].
Bone marrow-derived mesenchymal stem cells have been used to promote engraftment of peripheral mononuclear blood cells in transplanted animals [35–41]. In 2005, Kim et al reported the examination of adipose-derived mesenchymal stem cells to promote engraftment of hematopoietic stem cell transplantation in NOD/SCID mice, indicating that ASCs could be used as an alternative to BMSCs [35]. Planat-Benard et al demonstrated that mouse adipose-derived stem cells, human adipose-derived stem cells, and even de-differentiated adipocytes all induce significant neovascularization when injected into the muscles of an ischemic mouse hind limb model (immunodeficient strains were used with human donor cells) [42].
While the potential of ASCs is exciting, it is important to determine the origin of ASCs, as these cells could potentially cure hematopoietic malignancies that are as yet untreatable. We have observed a GFP+ population of cells residing in the adipose tissue compartments after GFP+ bone marrow transplantation. We conducted detailed immunohistochemical analysis of chimeric whole fat compartments and subsequently isolated, characterized and cultured ASCs from GFP+ BM chimeras. The immunohistochemistry analysis of fat tissue revealed that a large fraction of GFP+ cells in adipose tissue were strongly positive for CD45 and smooth muscle actin (Figure 1). The CD45+/SMA+ cells were evenly scattered around the adipocytes and blood vessels, while CD45+ leukocytes within the vessels were GFP+ (Figure 3). A small fraction of GFP+/CD90+ cells also existed in the perivascular area (Figure 2) but their presence was highly variable within the tissue.
In addition to whole fat analysis, ASCs were isolated, characterized and cultured from two different adipose tissue compartments: the omental (abdominal) and the inguinal fat (subcutaneous) pad. Flow cytometric and immunocytochemical analyses showed that cultured ASCs were CD45−/CD90+/CD29+ (Figure 5). A significantly higher number of GFP−/CD90+ cells were isolated from the subcutaneous depot as compared to the abdominal depot (Figure 6). Furthermore, the in vitro adipogenic differentiation of ASCs was achieved; however, all cells that had differentiated were GFP− (Figure 4).
Based on phenotypical analysis, GFP+ cells in adipose tissue in this rat model appear to be of both hematopoietic and mesenchymal origin; however, infrequent isolation of GFP+ ASCs and their lack of adipogenic differentiation suggest that the contribution of BM to ASC generation is a rare event. This could also be due to the initial transplantation of a bone marrow cell population with a limited mesenchymal stem cell population or engraftment. Numerous previous studies have used the bone marrow transplantation model to examine the differentiation of BMCs into the parenchyma of various tissues; however, the bone marrow stroma and mesenchymal stem cells have been shown to remain host phenotype after experimental and clinical bone marrow transplantation, [43–46] suggesting limited engraftment capability of donor BMCs into host bone marrow stroma compartment after transplantation. Nevertheless, both hematopoietic and mesenchymal GFP+ cells were identified in the adipose tissue of radiation chimeras in this study, and the result may indicate the differentiation of hematopoietic stem cells into mesenchymal cell populations. Thus, although improved engraftment of BMCs into mesenchymal compartment might increase BM-derived ASCs, the study demonstrates that in spite of plasticity of hematopoietic stem cells, they appear to have limited contribution to ASC generation. Future studies to further understand the cell behavior after migration and the effect of body mass index include treating the rats with a high fat diet. This study provides further insight in both the frequency of ASCs in two different rat depots and also the stem cell migration into the adipose tissue compartments.
Conclusions
This data increases our understanding of the origin of mesenchymal stem cells residing in adipose tissue by providing further insight into the migration of bone marrow cells into adipose tissue compartments. Both hematopoietic and mesenchymal stem cells were identified as migrating from the bone marrow into both adipose compartments; however, infrequent isolation of GFP+ ASCs and their lack of adipogenic differentiation suggest that the contribution of BM to ASCs generation might be minor. This could be correlated to the initial population of mesenchymal stem cells in the transplanted bone marrow cells. It was also found that the GFP− mesenchymal stem cell population was significantly higher in the subcutaneous depot as compared to the abdominal depot, indicating the inguinal fat pad may be a better source of ASCs. While in this rat study, the presence of a small population (~5%) of GFP+ ASCs suggests that ASCs could be recruited from the bone marrow, if necessary, a significant GFP+ mesenchymal stem cell population originating from the bone marrow was not identified. Further studies can be conducted to identify alternative ASC origins, such as pericytes.
Acknowledgments
The authors gratefully acknowledge Dr. Masaru Okabe (Osaka University, Japan) for providing GFP rats, and Dr. Fengli Guo, Mr. Marc Rubin and Ms. Mara Sullivan for electron microscopy technical assistance. This work was supported by CA76541 to DBS.
References
- 1.Stiles J, Francendese A, Masoro E. Influence of age on size and number of fat cells in the epididymal depot. Am J Physiol. 1975;229:1531–1568. doi: 10.1152/ajplegacy.1975.229.6.1561. [DOI] [PubMed] [Google Scholar]
- 2.Dardick I, Poznanski W, Waheed I, et al. Ultrastructural observations on differentiating human preadipocytes cultured in vitro. Tissue Cell. 1976;8:561–571. doi: 10.1016/0040-8166(76)90013-6. [DOI] [PubMed] [Google Scholar]
- 3.Hausman GJ, Richardson RL. Newly recruited and pre-existing preadipocytes in cultures of porcine stromal-vascular cells: morphology, expression of extracellular matrix components, and lipid accretion. J Anim Sci. 1998;76:48–60. doi: 10.2527/1998.76148x. [DOI] [PubMed] [Google Scholar]
- 4.Niesler CU, Siddle K, Prins JB. Human preadipocytes display a depot-specific susceptibility to apoptosis. Diabetes. 1998;47:1365–1368. doi: 10.2337/diab.47.8.1365. [DOI] [PubMed] [Google Scholar]
- 5.Maslowska MHSA, MacLean LD, Cianflone K. Regional differences in triacylglycerol synthesis in adipose tissue and in cultured preadipocytes. Journal of Lipid Research. 1993;34:219–228. [PubMed] [Google Scholar]
- 6.Kirkland JL, Hollenberg CH, Gillon WS. Age, anatomic site, and the replication and differentiation of adipocyte precursors. Am J Physiol. 1990;258:C206–10. doi: 10.1152/ajpcell.1990.258.2.C206. [DOI] [PubMed] [Google Scholar]
- 7.Tchkonia T, Giorgadze N, Pirtskhalava T, et al. Fat depot origin affects adipogenesis in primary cultured and cloned human preadipocytes. Am J Physiol Regul Integr Comp Physiol. 2002;282:R1286–1296. doi: 10.1152/ajpregu.00653.2001. [DOI] [PubMed] [Google Scholar]
- 8.Hausman GJ, Richardson RL. Adipose tissue angiogenesis. J Anim Sci. 2004;82:925–934. doi: 10.2527/2004.823925x. [DOI] [PubMed] [Google Scholar]
- 9.Hausman G, Hausman D. Search for the preadipocyte progenitor cell. J Clin Invest. 2006;116:3103–3106. doi: 10.1172/JCI30666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zuk PA, Zhu M, Mizuno H, et al. Multilineage Cells from Human Adipose Tissue: Implications for Cell-Based Therapies. Tissue Engineering. 2001;7:211–228. doi: 10.1089/107632701300062859. [DOI] [PubMed] [Google Scholar]
- 11.Safford KM, Hicok KC, Safford SD, et al. Neurogenic differentiation of murine and human adipose-derived stromal cells. Biochem Biophys Res Commun. 2002;294:371–379. doi: 10.1016/S0006-291X(02)00469-2. [DOI] [PubMed] [Google Scholar]
- 12.Seo M, Suh S, Bae Y, et al. Differentiation of human adipose stromal cells into hepatic lineage in vitro and in vivo. Biochem Biophys Res Commun. 2005;328:258–264. doi: 10.1016/j.bbrc.2004.12.158. [DOI] [PubMed] [Google Scholar]
- 13.Taléns-Visconti R, Bonora A, Jover R, et al. Human mesenchymal stem cells from adipose tissue: Differentiation into hepatic lineage. Toxicol In Vitro. 2007;21:324–329. doi: 10.1016/j.tiv.2006.08.009. [DOI] [PubMed] [Google Scholar]
- 14.Tholpady SS, Katz AJ, Ogle RC. Mesenchymal stem cells from rat visceral fat exhibit multipotential differentiation in vitro. Anat Rec. 2003;272A:398–402. doi: 10.1002/ar.a.10039. [DOI] [PubMed] [Google Scholar]
- 15.Kang SK, Lee DH, Bae YC, et al. Improvement of Neurological Deficits by Intracerebral Transplantation of Human Adipose-Derived Stromal Cells After Cerebral Ischemia in Rats. Experimental Neurology. 2003;183:355–366. doi: 10.1016/s0014-4886(03)00089-x. [DOI] [PubMed] [Google Scholar]
- 16.Safford KM, Safford SD, Gimble JM, et al. Characterization of Neuronal/Glial Differentation of Murine Adipose-Derived Adult Stromal Cells. Experimental Neurology. 2004;187:319–328. doi: 10.1016/j.expneurol.2004.01.027. [DOI] [PubMed] [Google Scholar]
- 17.Crossno J, Majka S, Grazia T, et al. Rosiglitazone promotes development of a novel adipocyte population from bone marrow-derived circulating progenitor cells. J Clin Invest. 2006;116:3220–3228. doi: 10.1172/JCI28510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Okabe M, Ikawa M, Kominami K, et al. ‘Green mice’ as a source of ubiquitous green cells. FEBS Lett. 1997;407:313–319. doi: 10.1016/s0014-5793(97)00313-x. [DOI] [PubMed] [Google Scholar]
- 19.Ito T, Suzuki A, Imai E, et al. Bone marrow is a reservoir of repopulating mesangial cells during glomerular remodeling. J Am Soc Nephrol. 2001;12:2625–2635. doi: 10.1681/ASN.V12122625. [DOI] [PubMed] [Google Scholar]
- 20.Kawakami N, Sakane N, Nishizawa F, et al. Green fluorescent protein-transgenic mice: immune functions and their application to studies of lymphocyte development. Immunol Lett. 1999;70:165–171. doi: 10.1016/s0165-2478(99)00152-2. [DOI] [PubMed] [Google Scholar]
- 21.Toyokawa H, Nakao A, Stolz D, et al. 3D-confocal structural analysis of bone marrow-derived renal tubular cells during renal ischemia/reperfusion injury. Lab Invest. 2006;86:72–82. doi: 10.1038/labinvest.3700363. [DOI] [PubMed] [Google Scholar]
- 22.Stolz D, Zamora R, Vodovotz Y, et al. Peroxisomal localization of inducible nitric oxide synthase in rat hepatocytes. Hepatology. 2002;36:81–93. doi: 10.1053/jhep.2002.33716. [DOI] [PubMed] [Google Scholar]
- 23.Wack KE, Ross MA, Zegarra V, et al. Sinusoidal ultrastructure evaluated during the revascularization of regenerating rat liver. Hepatology. 2001;33:363–378. doi: 10.1053/jhep.2001.21998. [DOI] [PubMed] [Google Scholar]
- 24.Mitchell J, McIntosh K, Zvonic S, et al. Immunophenotype of human adipose-derived cells: temporal changes in stromal-associated and stem cell-associated markers. Stem Cells. 2006;24:376–385. doi: 10.1634/stemcells.2005-0234. [DOI] [PubMed] [Google Scholar]
- 25.Zuk PA, Zhu M, Ashjian P, et al. Human adipose tissue is a source of multipotent stem cells. Mol Biol Cell. 2002;13:4279–4295. doi: 10.1091/mbc.E02-02-0105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gimble JM, Guilak F. Differentiation potential of adipose derived adult stem (ADAS) cells. Curr Top Dev Biol. 2003;58:137–160. doi: 10.1016/s0070-2153(03)58005-x. [DOI] [PubMed] [Google Scholar]
- 27.Dragoo JL, Choi JY, Lieberman JR, et al. Bone induction by BMP-2 transduced stem cells derived from human fat. J Orthop Res. 2003;21:622–629. doi: 10.1016/S0736-0266(02)00238-3. [DOI] [PubMed] [Google Scholar]
- 28.Awad HA, Halvorsen YD, Gimble JM, et al. Effects of transforming growth factor beta1 and dexamethasone on the growth and chondrogenic differentiation of adipose-derived stromal cells. Tissue Eng. 2003;9:1301–1312. doi: 10.1089/10763270360728215. [DOI] [PubMed] [Google Scholar]
- 29.Nathan S, Das De S, Thambyah A, et al. Cell-based therapy in the repair of osteochondral defects: a novel use for adipose tissue. Tissue Eng. 2003;9:733–744. doi: 10.1089/107632703768247412. [DOI] [PubMed] [Google Scholar]
- 30.Ashjian PH, Elbarbary AS, Edmonds B, et al. In vitro differentiation of human processed lipoaspirate cells into early neural progenitors. Plast Reconstr Surg. 2003;111:1922–1931. doi: 10.1097/01.PRS.0000055043.62589.05. [DOI] [PubMed] [Google Scholar]
- 31.ten Hove W, Verspaget H, Barge R, et al. Liver chimerism after allogeneic blood stem cell transplantation. Transplant Proc. 2007;39:231–236. doi: 10.1016/j.transproceed.2006.10.022. [DOI] [PubMed] [Google Scholar]
- 32.Ng I, Chan K, Shek W, et al. High frequency of chimerism in transplanted livers. Hepatology. 2003;38:989. doi: 10.1053/jhep.2003.50395. [DOI] [PubMed] [Google Scholar]
- 33.Korbling M, Katz R, Khanna A, et al. Hepatocytes and epithelial cells of donor origin in recipients of peripheral-blood stem cells. N Engl J Med. 2002;346:738–746. doi: 10.1056/NEJMoa3461002. [DOI] [PubMed] [Google Scholar]
- 34.Nakao A, Toyokawa H, Kimizuka K, et al. Simultaneous bone marrow and intestine transplantation promotes marrow-derived hematopoietic stem cell engraftment and chimerism. Blood. 2006;108:1413–1420. doi: 10.1182/blood-2006-02-004341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kim S, Cho H, Kim Y, et al. Human adipose stromal cells expanded in human serum promote engraftment of human peripheral blood hematopoietic stem cells in NOD/SCID mice. Biochem Biophys Res Commun. 2005;329:25–31. doi: 10.1016/j.bbrc.2005.01.092. [DOI] [PubMed] [Google Scholar]
- 36.Angelopoulou M, Novelli E, Grove J, et al. Cotransplantation of human mesenchymal stem cells enhances human myelopoiesis and megakaryocytopoiesis in NOD/SCID mice. Exp Hematol. 2003;31:413–420. doi: 10.1016/s0301-472x(03)00042-0. [DOI] [PubMed] [Google Scholar]
- 37.Maitra B, Szekely E, Gjini K, et al. Human mesenchymal stem cells support unrelated donor hematopoietic stem cells and suppress T-cell activation. Bone Marrow Transplant. 2004;33:597–604. doi: 10.1038/sj.bmt.1704400. [DOI] [PubMed] [Google Scholar]
- 38.Laughlin M, Barker J, Bambach B, et al. Hematopoietic engraftment and survival in adult recipients of umbilical-cord blood from unrelated donors. N Engl J Med. 2001;344:1815–1822. doi: 10.1056/NEJM200106143442402. [DOI] [PubMed] [Google Scholar]
- 39.Nolta J, Hanley M, Kohn D. Sustained human hematopoiesis in immunodeficient mice by cotransplantation of marrow stroma expressing human interleukin-3: analysis of gene transduction of long-lived progenitors. Blood. 1994;83:3041–3051. [PubMed] [Google Scholar]
- 40.Anklesaria P, Kase K, Glowacki J, et al. Engraftment of a clonal bone marrow stromal cell line in vivo stimulates hematopoietic recovery from total body irradiation. Proc Natl Acad Sci U S A. 1987;84:7681–7685. doi: 10.1073/pnas.84.21.7681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Almeida-Porada G, Porada C, Tran N, et al. Cotransplantation of human stromal cell progenitors into preimmune fetal sheep results in early appearance of human donor cells in circulation and boosts cell levels in bone marrow at later time points after transplantation. Blood. 2000;95:3620–3627. [PubMed] [Google Scholar]
- 42.Planat-Benard V, Silvestre JS, Cousin B, et al. Plasticity of human adipose lineage cells toward endothelial cells: physiological and therapeutic perspectives. Circulation. 2004;109:656–663. doi: 10.1161/01.CIR.0000114522.38265.61. [DOI] [PubMed] [Google Scholar]
- 43.Simmons P, Przepiorka D, Thomas E, et al. Host origin of marrow stromal cells following allogeneic bone marrow transplantation. Nature. 1987;328:429–432. doi: 10.1038/328429a0. [DOI] [PubMed] [Google Scholar]
- 44.Rieger K, Marinets O, Fietz T, et al. Mesenchymal stem cells remain of host origin even a long time after allogeneic peripheral blood stem cell or bone marrow transplantation. Exp Hematol. 2005;33:605–611. doi: 10.1016/j.exphem.2005.02.004. [DOI] [PubMed] [Google Scholar]
- 45.Cilloni D, Carlo-Stella C, Falzetti F, et al. Limited engraftment capacity of bone marrow-derived mesenchymal cells following T-cell-depleted hematopoietic stem cell transplantation. Blood. 2000;96:3637–3643. [PubMed] [Google Scholar]
- 46.Pozzi S, Lisini D, Podesta M, et al. Donor multipotent mesenchymal stromal cells may engraft in pediatric patients given either cord blood or bone marrow transplantation. Exp Hematol. 2006;34:934–942. doi: 10.1016/j.exphem.2006.03.007. [DOI] [PubMed] [Google Scholar]