

Abstract
Arousal from sleep is an important protective response to hypoxia that becomes rapidly depressed in active sleep (AS) when hypoxia is repeated. This study questioned whether there might also be selective depression of cardio-respiratory responses to hypoxia during AS.
Nine newborn lambs (7-22 days of age) were studied over three successive nights. The first and third nights were baseline studies (inspired oxygen fraction, Fi,O2= 0.21). During the second night, during every epoch of sleep, lambs were exposed to 60 s episodes of isocapnic hypoxia (Fi,O2= 0.10).
During quiet sleep (QS), the probability of arousal in hypoxia exceeded the probability of spontaneous arousal (P < 0.001) throughout repeated exposures to hypoxia. Similarly, there were persisting increases in ventilation (135 ± 25 %), blood pressure (3 ± 1 %) and heart rate (3 ± 1 %).
By contrast, rapid depression of all responses occurred during repetitive hypoxia in AS. Thus, the probability of arousal in hypoxia exceeded the probability of spontaneous arousal during the first 10 hypoxia exposures (P < 0.001) but not thereafter. Similarly, during the first 10 exposures to hypoxia, the changes in ventilation (88 ± 15 %) and blood pressure (5 ± 1 %) were greater than subsequent responses (P < 0.05).
We conclude that, when repeated, hypoxia rapidly becomes ineffective in stimulating protective arousal, ventilatory and blood pressure responses in AS, but not in QS. Selective depression of responses during AS may render the newborn particularly vulnerable to hypoxia in this state.
The classical reflex response to a fall in arterial oxygen tension, an increase in ventilation mediated by the peripheral chemoreceptors (Heymans & Heymans, 1927), is significantly depressed during sleep; thus arousal to the waking state becomes a critical defence against developing profound hypoxaemia in sleep (Phillipson & Sullivan, 1978). Arousal also re-establishes the powerful central drive to breathe that is intrinsic to wakefulness but which is lost during sleep (Phillipson & Sullivan, 1978). In complementary fashion, the ventilatory augmentation associated with hypoxaemia, albeit limited during sleep, may also stimulate arousal (Phillipson et al. 1978; Henderson-Smart & Read, 1979b;Jeffery & Read, 1980; Neubauer et al. 1981; Berthon-Jones & Sullivan, 1982).
Both the arousal and the ventilatory responses to hypoxaemia in sleep may be substantially altered if the hypoxaemic stress is repeated, as it is in obstructive sleep apnoea (OSA) in the adult (Davies & Stradling, 1993) and possibly in infants who die of sudden infant death syndrome (SIDS) (Rognum & Saugstad, 1991; Kahn et al. 1992). New information shows that hypoxia, though normally a potent arousing stimulus (Phillipson et al. 1978), becomes ineffective when hypoxia is repeated (Fewell & Konduri, 1989; Johnston et al. 1998). Depression of arousal is rapid in onset and confined to the active sleep (AS) state (also known as rapid-eye-movement sleep); by contrast, hypoxia remains a powerful arousing stimulus in quiet sleep (QS) (Johnston et al. 1998). As with arousal, the ventilatory response to hypoxia is lost during repeated exposures to hypoxia (Worthington et al. 1993; Waters et al. 1996) a phenomenon that is similar in some respects to the ventilatory depression that emerges during continuous hypoxia (Weil, 1994). Circulatory changes occurring during hypoxia are also powerful stimuli for arousal that may add to the arousing effects of respiratory changes (Fewell & Johnson, 1984; Horne et al. 1989, 1991), but whether these additive relationships might change with repetition of hypoxia is not known. Whether the circulatory changes themselves are modified by repeated hypoxia is also unknown, though persisting hypertension develops in response to daily episodes of hypoxia over several weeks (Fletcher et al. 1992; Brooks et al. 1997). Resolution of these issues is important, as marked oscillation of blood pressure (and possibly persisting hypertension) together with sleep disruption are characteristic features of OSA (Shepard, 1986).
The mechanisms that underlie the depressed arousal and ventilatory responses when hypoxia is repeated are not known. It is also unknown whether depression of ventilation, like depression of arousal, is similarly confined to the AS state, as previous studies have not controlled for the sleep- waking state (Worthington et al. 1993; Waters et al. 1996). It is important to establish whether depression is confined to AS as concurrent failure of ventilation and arousal may seriously add to the risk of severe hypoxaemia in AS, as this state is characterized by reduced lung gas volume, irregular breathing pattern and diminished intercostal muscle function (Henderson-Smart & Read, 1979a).
This study tested the hypothesis that cardio-respiratory responses, like the arousal response, are depressed by repetitive hypoxia during AS. We reasoned that the coincident depression of arousal and cardio-respiratory responses might represent the operation of a common mechanism. These questions were addressed by examining cardio-respiratory and arousal responses in lambs during a sequence of repeated exposures to hypoxia in an overnight study.
METHODS
Animals
Nine newborn lambs (Border-Leicester × Merino, 4.4 ± 0.3 kg, mean ±s.e.m.) were separated from their ewes within 48 h of birth, housed with another lamb and taught to feed unaided prior to surgery (Lamb Milk Replacer, Veanavite, Shepparton, Australia). Five of the lambs were used in a previous study (Johnston et al. 1998). All surgical and experimental protocols were carried out in accordance with the guidelines established by the National Health and Medical Research Council of Australia and with the approval of the Standing Committee in Ethics in Animal Experimentation of Monash University. At the completion of the study, lambs were killed by lethal overdose of anaesthetic (150 mg kg−1 sodium pentobarbitone, i.v.).
Surgical preparation
Each lamb was anaesthetized (2 % halothane in 60 % oxygen and nitrous oxide delivered via a face mask for induction) and then intubated and mechanically ventilated (1-1.5 % halothane in 60 % oxygen, balance nitrous oxide). We inserted a catheter (Tygon, o.d. 1.5 mm) non-occlusively into the axillary artery to measure arterial blood pressure and heart rate. Arterial blood was also sampled from this catheter for analysis of pH, arterial partial pressure of carbon dioxide (Pa,CO2), arterial partial pressure of oxygen (Pa,O2) and base excess corrected for a body temperature of 39.5°C (ABL500 Radiometer, Copenhagen, Denmark) during periods of normoxia and hypoxia in both sleep states. A silicon rubber balloon-tipped catheter of internal volume of less than 1 ml was inserted into the intrapleural cavity at the sixth intercostal space to record pleural pressure and thus assess changes in respiratory amplitude. Paired Teflon-coated stainless steel wire electrodes (Medwire, New York) were implanted on the left and right parietal cortex to record the electrocorticogram (ECoG), at the inner and outer canthus of the right eye to record the electro-oculogram (EOG) and in the dorsal musculature of the neck to record the nuchal electromyogram (EMGn). Electrodes were referenced to a single electrode sewn into the subcutaneous tissue of the scalp. Sleep state determinations were based upon these electrophysiological recordings. QS was defined by the presence of a high-voltage (80-100 μV peak to peak), low-frequency (1-3 Hz) ECoG, the absence of eye movements and reduced EMGn activity compared with the awake state. Arousal from this state was characterized by a change in the ECoG to low-voltage, high-frequency waves, increased EMGn activity, and opening of the eyes. AS was defined by the presence of low-voltage (10-20 μV peak to peak), predominately high-frequency waves (>3 Hz) on the ECoG, the presence of rapid eye movements and the absence of EMGn tone (Horne et al. 1989; Grant et al. 1995). Arousal was characterized by a return of tonic activity in the EMGn.
We also implanted a fenestrated tracheostomy tube (i.d. 5.0 mm, Shiley Inc., Irvine, CA, USA) into the trachea. When the fenestrated tube was capped the animal breathed normally through its upper airway. When an inner cannula was placed in the tracheostomy tube the fenestration leading to the upper airway was blocked and the lamb breathed through an external circuit. This system allowed the inspired gas mixture to be rapidly changed during study periods.
Physiological measurements
Chronic recordings were made using procedures that we have previously reported (Johnston et al. 1998). Each lamb was allowed a minimum of 72 h to recover from surgery before being studied. The lamb's cage was partitioned to prevent the lamb from turning around while still allowing the lamb to stand, lie down, move forward and backward and feed freely. The arterial catheter and intrapleural balloon were connected to calibrated strain gauge manometers (Cobe CDX III, Cobe Laboratories, Lakewood, CO, USA) and referenced to the mid-thoracic level when the animal was lying down. A pulse oximeter probe (Nellcor N200, Nellcor Inc., Hayward, CA, USA) was placed around the tail to measure arterial oxygen saturation (Sp,O2). We connected the strain gauge manometers, pulse oximeter and electrodes (ECoG, EOG and EMGn) to an amplifier and signal conditioner (Cyberamp 380, Axon Instruments). Arterial pressure, pleural pressure and Sp,O2 were low-pass filtered at 100 Hz. The electrophysiological signals were also filtered (30-100 Hz for EMGn and 0.3-40 Hz for EOG and ECoG). All signals were displayed on a thermal chart recorder (model 7758A, Hewlett-Packard, Waltham, MA, USA) and stored on computer at a sampling rate of 200 Hz (ADAC 4801A analog-digital converting board, ADAC, Woburn, MA, USA and CVSOFT Data Acquisition and Analysis Software, Odessa Computer Systems, Calgary, Canada).
Overnight sleep study
Each study consisted of three 12 h overnight sleep recordings, performed on sequential control, test and recovery nights. The studies were conducted between 20.00 and 08.00 h under controlled temperature conditions (22-25°C). To control for possible effects of the breathing circuit on sleep and arousability, the lambs were randomly assigned on the control and recovery nights to breathe either via the upper airway, or, alternatively, via a breathing circuit. A bias flow of 20 l min−1 was used in the breathing circuit and a positive end-expiratory pressure of 2 cmH2O was applied to preserve lung inflation while the upper airway was bypassed. On the test night, the lambs were exposed to 60 s of isocapnic hypoxia during every AS and QS epoch (inspired oxygen fraction, Fi,O2= 0.10; inspired carbon dioxide fraction, Fi,CO2= 0.03; in N2). The stimulus was presented in each sleep epoch after allowing a 30 s baseline period for the sleep state to be clearly established. Duplicate studies were conducted in two lambs after allowing 2-4 days recovery between studies. Thus, a total of 11 studies were conducted over an age range of 7-22 days (12 ± 2 days, mean ±s.e.m.). Duplicate cardio-respiratory, blood gas and acid- base data were averaged to give one value for each lamb, then averaged and presented as means ±s.e.m. The lambs tolerated the brief episodes of moderate hypoxia without distress, showing no lasting effects from the test procedure (as evidenced by their normal sleep on the recovery night), and gained weight (220 ± 22 g day−1) and behaved normally throughout the study.
Data analysis
Arousal from sleep
The arousing effects of the hypoxia stimulus were quantified by using probability analysis (Horne et al. 1989, 1991) to compare the probability of arousal during hypoxia and the probability of spontaneous arousal. This form of analysis allows the determination of the ‘true’ probability of arousal, free of contamination from spontaneous arousals, and also controls for temporal changes that may occur in overnight studies. The probability of arousal during hypoxia was calculated for each sleep state as the percentage of tests in which arousal occurred during the 60 s stimulus. The probability of spontaneous arousal was calculated for each sleep state during the control and recovery nights as the percentage of spontaneous arousals that occurred between 30 and 90 s of each sleep epoch (a time period that matched the period in which the hypoxia stimuli were presented). Data were stratified (Horne et al. 1989, 1991; Johnston et al. 1998) to prevent bias arising from differences in the number of sleep epochs between animals. The analysis included the first 40 sleep epochs on each of the three study nights; too few animals were exposed to a greater number of hypoxia episodes to allow further analysis. Data from the first 40 exposures were partitioned into groups of 10 in order to test the time course of responses. Differences between the probability of arousal in hypoxia and the probability of spontaneous arousal, the probability of arousal in hypoxia for sequential groups of 10 exposures to hypoxia, the probability of spontaneous arousal on the control and recovery nights, and the probability of spontaneous arousal with or without the breathing circuit were tested using the χ2 test.
Cardio-respiratory responses
All analyses were performed using analysis software (CVSOFT Data Acquisition and Analysis Software, Odessa Computer Systems, Calgary, Canada). Mean arterial blood pressure, heart rate, Sp,O2 values, respiratory amplitude (A, as assessed by pleural pressure), respiratory frequency (f) and an index of ventilation (V, the product of A and f) were determined for each sleep epoch during the test night and averaged over the 30 s period of normoxia prior to the hypoxia stimulus and over the 10 breaths immediately prior to arousal during hypoxia. For those tests which did not result in an arousal, cardio-respiratory variables were averaged over the 10 breaths that occurred just prior to the 30 s point of the hypoxia exposure, a period which corresponded to the average arousal time. Measurements of Sp,O2 were corrected for the total time delay due to the breathing circuit, the oximeter response time, and the circulation delay time to the tail (range 6-13 s; mean ±s.e.m., 9 ± 1 s).
A two-way analysis of variance for repeated measures was used to determine if differences in blood gas and acid-base status existed between sleep states and between periods of normoxia and hypoxia. A one-way analysis of variance for repeated measures was used to detect differences in the number of sleep epochs, epoch length, and total sleep time across the three study nights, and to assess the effects of repetitive hypoxia upon cardio-respiratory variables. A Student-Newman-Keuls test was used to isolate any differences detected by the analyses of variance. Student's paired t test was used to detect differences in cardio-respiratory variables between periods of normoxia and hypoxia. In all tests, P < 0.05 was considered statistically significant.
RESULTS
There were no sleep state-related differences in arterial blood gases or pH during normoxia (Table 1). During periods of hypoxia, Pa,O2 was significantly lower (P < 0.05) in both sleep states compared with normoxia. Consistent with the poorer ventilatory response to hypoxia in AS (Henderson-Smart & Read, 1979b), Pa,O2 was lower and Pa,CO2 slightly higher in this state compared with QS during exposure to hypoxia.
Table 1.
Effects of hypoxia on blood gas variables during sleep in lambs
| Quiet sleep | Active sleep | |||
|---|---|---|---|---|
| Normoxia | Hypoxia | Normoxia | Hypoxia | |
| Pa,O2 (mmHg) | 99 ± 3 | 59 ± 3* | 96 ± 3 | 48 ± 2*† |
| Pa,CO2 (mmHg) | 41 ± 2 | 40 ± 1 | 42 ± 1 | 43 ± 1† |
| pH | 7.41 ± 0.01 | 7.41 ± 0.01 | 7.40 ± 0.01 | 7.39 ± 0.01 |
| Base excess (mmol 1−1) | 1.1 ± 0.6 | 0.9 ± 0.8 | 0.9 ± 0.6 | 0.9 ± 0.6 |
Values are means ±s.e.m., n = 9 lambs. Pa,CO2, arterial partial pressure of carbon dioxide; Pa,O2, arterial partial pressure of oxygen.
P <0.05, normoxia vs. hypoxia
P <0.05, QS vs. AS.
Arousal from sleep
There were no differences in epoch number, epoch length or total sleep time according to whether the upper airway was intact or bypassed, or between the control and recovery nights in either sleep state. Accordingly, the probability of spontaneous arousal for each state was determined by pooling data from the control and recovery nights. The imposition of repetitive hypoxia in AS did not alter the number of sleep epochs or the total sleep time from values of the control and recovery nights, although the epoch length was slightly reduced (P < 0.05, Table 2). In QS, although the number of epochs remained constant, epoch length and total sleep time were significantly reduced on the test night (P < 0.05).
Table 2.
Sleep distribution data in overnight recordings
| Study night | Quiet sleep | Active sleep |
|---|---|---|
| Number of epochs | ||
| Control | 32 ± 2 | 20 ± 1 |
| Test | 31 ± 3 | 19 ± 2 |
| Recovery | 29 ± 3 | 19 ± 1 |
| Epoch length (min) | ||
| Control | 5.7 ± 0.3 | 4.2 ± 0.4 |
| Test | 4.1 ± 0.3* | 3.2 ± 0.2* |
| Recovery | 5.8 ± 0.3 | 4.2 ± 0.3 |
| Total sleep time (min) | ||
| Control | 180 ± 12 | 86 ± 12 |
| Test | 122 ± 11* | 61 ± 6 |
| Recovery | 168 ± 14 | 78 ± 7 |
Values are means ±s.e.m. (n = 11 studies).
P <0.05, control vs. test night, recovery vs. test night.
During QS, the overall probability of arousal, calculated over the total of 40 exposures to hypoxia, averaged 61 %, significantly higher than the probability of spontaneous arousal (22 %, χ2= 97.0, P < 0.001). Similarly, in AS the overall (40 exposures) probability of arousal in hypoxia (29 %) was significantly higher than the probability of spontaneous arousal (14 %, χ2= 17.4, P < 0.001). When the series of 40 exposures to hypoxia was partitioned into sequential groups of ten, the pattern of arousal probability differed significantly between sleep states. During QS, the probability of arousal in hypoxia remained significantly higher than the probability of spontaneous arousal for all the groups of 10 exposures to hypoxia (P < 0.001). Conforming with this result, there were no differences in the probability of arousal in hypoxia across the sequential groups of 10 exposures to hypoxia (P > 0.05, Fig. 1). Persistence of arousal in QS is consistent with the significant reductions in epoch length and total sleep time that were observed in this state (Table 2). By contrast, in AS only during the first 10 exposures to hypoxia was the probability of arousal significantly higher than the probability of spontaneous arousal (χ2= 20.0, P < 0.001). Subsequently, the probability of arousal in hypoxia decreased to equal the probability of spontaneous arousal. In keeping with this, the probability of arousal during the first 10 exposures to hypoxia was significantly higher than each of the subsequent exposures (P < 0.01, Fig. 1). The rapid depression of arousal conforms with the observation that total sleep time in AS was unchanged by the testing procedure (Table 2).
Figure 1. Probability of arousal from sleep and corresponding ventilatory responses during repetitive hypoxia.
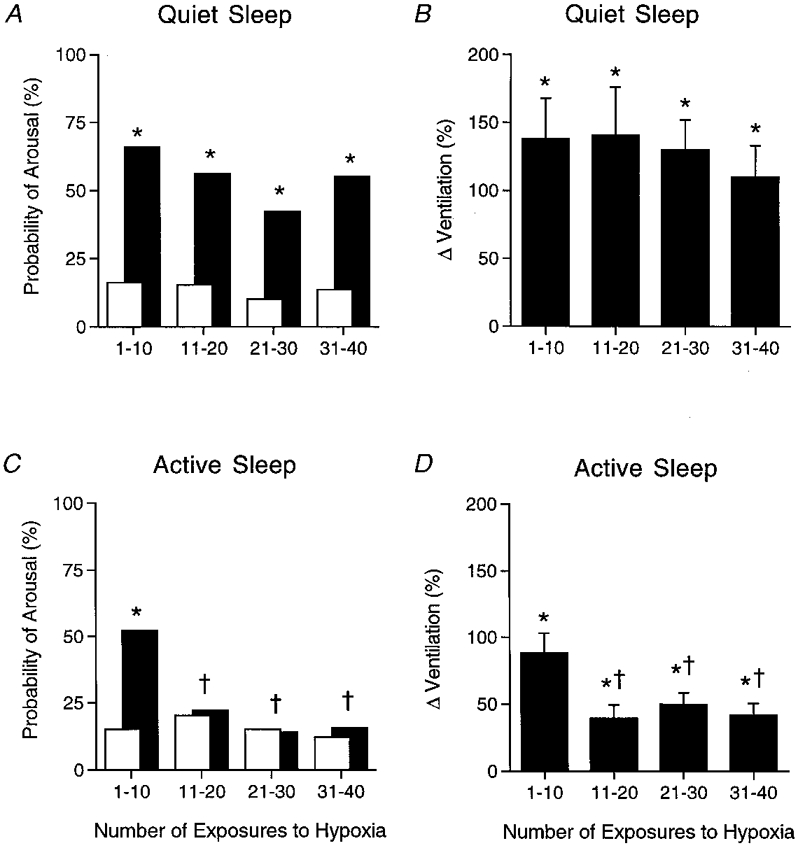
Probability of arousal from sleep during repetitive hypoxia (▪) during QS (A) and AS (C) plotted against the number of exposures to hypoxia (values for 11 studies in 9 lambs), compared with the probability of spontaneous arousal during normoxia (□). Associated ventilatory responses to repeated hypoxia (measured as a percentage change from normoxia levels) are shown for QS (B) and AS (D). Note that during QS the probability of arousal in response to hypoxia remained elevated above the spontaneous levels, whereas during AS the increment in arousal was abolished after just 10 exposures. * P < 0.05, probability of arousal during hypoxia vs. probability of spontaneous arousal; † P < 0.05, probability of arousal during hypoxia vs. probability of arousal during hypoxia for the first 10 hypoxic exposures. Similarly, there was no depression of the ventilatory response to hypoxia during QS, but in AS there was a depression of the ventilatory response after 10 exposures that paralleled the arousal depression. * P < 0.05, ventilatory response to hypoxia (percentage change from normoxia) vs. ventilation in normoxia; † P < 0.05, ventilatory response to hypoxia vs. ventilatory response to hypoxia for the first 10 exposures to hypoxia.
Cardio-respiratory responses
During QS, V averaged over the entire series of 40 exposures to hypoxia increased significantly from the level in normoxia (136 ± 25 %, P < 0.001). In AS, V also increased significantly during hypoxia (55 ± 8 %, P < 0.001), though the increase was smaller than in QS (P < 0.005). Partitioned into sequential groups of 10 exposures to hypoxia, V increased significantly in all groups compared with normoxia during both QS and AS (P < 0.05, Table 3), though the nature of the increase differed between sleep states. In QS, there were no differences in V across the sequential groups of exposures (P > 0.05, Fig. 1). The similarity of V in QS early and late in the series of exposures to hypoxia is illustrated by the respiratory recordings in Fig. 2. By contrast, in AS, there was a significant fall in the magnitude of V after the first 10 exposures to hypoxia. Thus V in the first 10 exposures significantly exceeded the values in all of the subsequent exposures (P < 0.05, Fig. 1), as illustrated by the respiratory recordings in Fig. 2.
Table 3.
Cardio-respiratory responses during repetitive hypoxia: number of exposures to hypoxia
| Quiet sleep | Active sleep | |||||||
|---|---|---|---|---|---|---|---|---|
| 1–10 | 11–20 | 21–30 | 31–40 | 1–10 | 11–20 | 21–30 | 31–40 | |
| ΔV (%) | 144 ± 30 ‡ | 140 ± 35 ‡ | 131 ± 21‡ | 146 ± 41 ‡ | 88 ± 15 ‡ | 39 ± 10†‡ | 50 ± 9 †‡ | 41 ± 9 †‡ |
| ΔA (%) | 73 ± 16‡ | 74 ± 21‡ | 68 ± 15‡ | 83 ± 29‡ | 50 ± 12‡ | 25 ± 8†‡ | 34 ± 7‡ | 30 ± 8‡ |
| Δf (%) | 37 ± 5‡ | 34 ± 4‡ | 36 ± 4‡ | 36 ± 6‡ | 20 ± 4‡ | 12 ± 3‡ | 13 ± 4‡ | 9 ± 4 |
| ΔBP (%) | 3 ± 1‡ | 3 ± 1‡ | 3 ± 1‡ | 3 ± 1‡ | 5 ± 1‡ | 0 ± 1† | 4 ± 1 | 2 ± 2 |
| ΔHR(%) | 4 ± 1‡ | 3 ± 1‡ | 2 ± 1‡ | 4 ± 1‡ | −3 ± 1 | −2 ± 2 | 0 ± 1 | −2 ± 2 |
| Sp,O2(%) | 84 ± 2‡ | 85 ± 2‡ | 87 ± 2‡ | 87 ± 2‡ | 82 ± 2‡ | 83 ± 2‡ | 81 ± 2‡ | 81 ± 3‡ |
Values are means ±s.e.m., n = 9 lambs. Values are expressed as a percentage change from normoxia for the following: ΔV, change in ventilation; ΔA, change in respiratory amplitude; Δf, change in respiratory frequency; ΔBP, change in mean arterial blood pressure; ΔHR, change in heart rate. Sp,O2, pulse oxygen saturation.
P <0.05, value vs. 1–10 exposures (ANOVA and Student-Newman-Keuls post hoc test).
P < 0.05, value vs. normoxia (ANOVA and Student-Newman-Keuls post hoc test).
Figure 2. Respiratory and blood pressure responses to repeated hypoxia in sleep.
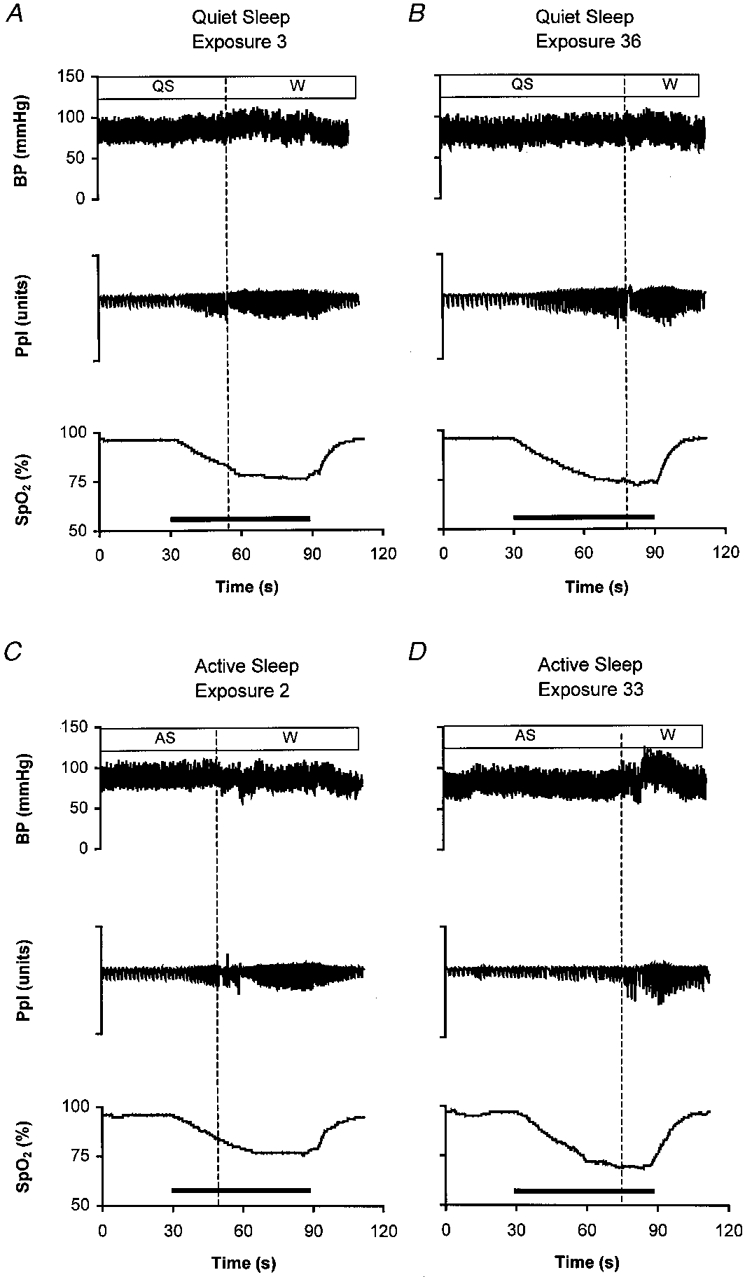
Physiological recordings in a sleeping lamb during periods of quiet sleep (QS) and active sleep (AS) during repeated exposures to hypoxia (horizontal bar). Arousal to the waking state (W) is signified by the vertical dotted line. BP, arterial blood pressure (mmHg); Ppl, pleural pressure (arbitrary units); Sp,O2, pulse oxygen saturation (as a percentage). The hypoxia stimulus, induced by rapidly changing the air in the inspiratory line to a gas mixture of Fi,O2= 0.10, Fi,CO2= 0.03 in N2, is denoted by the horizontal bar. Animals were exposed to a series of 40 repetitions of hypoxia during sleep; the number of each illustrated exposure in the series is indicated on each panel. During QS, progressive increases of ventilatory amplitude (Ppl) and BP occur up to the point of arousal. Note that the cardio-respiratory responses in QS are similar early (A) and late (B) in the series of exposures to hypoxia. During AS, by contrast, the progressive increases in ventilation and blood pressure that occur up to the point of arousal early in the series of exposures (C) are depressed during later exposures (D).
The components of V, amplitude (A) and frequency (f) examined over the entire series of exposures to hypoxia were significantly increased during hypoxia in both QS (ΔA = 71± 16 %, P = 0.003; Δf = 36± 3 %, P < 0.001) and AS (ΔA = 35± 6 %, P < 0.001; Δf = 14± 2 %, P < 0.001). These responses partitioned into sequential groups of 10 exposures to hypoxia are detailed in Table 3. For QS (in a similar pattern to V) A and f did not fall with repeated exposure to hypoxia. During AS, significantly lower values of V following exposures 1-10 were explained by decrements in A and f, though the changes reached significance (P < 0.05) only for A in the 11-20 exposure group (Table 3).
Over the entire series of repetitive exposures to hypoxia, blood pressure during QS increased from 79 ± 3 mmHg during normoxia to 82 ± 4 mmHg during hypoxia (P < 0.005), and heart rate increased from 173 ± 6 beats min−1 during normoxia to 179 ± 6 beats min−1 during hypoxia (P < 0.01). Overall, blood pressure during AS increased from 75 ± 3 mmHg during normoxia to 77 ± 3 mmHg during hypoxia (P < 0.001), and heart rate did not change (166 ± 6 beats min−1 during normoxia, 165 ± 5 beats min−1 during hypoxia, P > 0.05). When examined over sequential groups of 10 exposures to hypoxia, there was a significant increase in blood pressure and heart rate from baseline normoxia values throughout QS (P < 0.05, Table 3, Fig. 2). Moreover, the circulatory responses were of similar magnitude across the series of 10 exposures (Table 3). In marked contrast, during AS the blood pressure increase was confined to the first group of 10 exposures and there were no changes in heart rate (Table 3, Fig. 2).
DISCUSSION
This study shows that with repetition of hypoxia, cardio-respiratory responses, like the arousal response (Johnston et al. 1998), are depressed during AS. The concurrent loss of the ventilatory, cardiovascular and arousal responses appears to be the result of a powerful and rapid depression, occurring after just 10 exposures to brief, moderate hypoxia imposed during sleep. Intriguingly, this depression is confined to AS. During QS, hypoxia remains a powerful arousing stimulus and cardio-respiratory responses to hypoxia are preserved. It is possible, as we elaborate below, that the concurrent depression of arousal, ventilation and vascular responses during hypoxia may represent a modification of the AS state that is directed at oxygen conservation. Paradoxically, when hypoxia is repeated, this strategy could increase the risk of severe hypoxaemia and death during AS by also depressing the important protective response of arousal.
Our study supports the concept that arousal resulting from cardio-respiratory disturbances is poor in AS by comparison with QS. Arousal responses to a range of cardio-respiratory disturbances, including hypoxia, are significantly delayed in AS (Phillipson et al. 1978; Henderson-Smart & Read, 1979b;Jeffery & Read, 1980; Fewell & Baker, 1987; Horne et al. 1989, 1991). Additionally, arousal from AS, but not from QS, is depressed with repeated, episodic hypoxia (Johnston et al. 1998) and airway occlusion (Fewell et al. 1988; Harding et al. 1997). Moreover, an arousal decrement becomes apparent in AS very rapidly, occurring after as few as 10 exposures to hypoxia (Johnston et al. 1998).
When exposures to hypoxia are more numerous and more severe, arousal depression is also seen in QS (Fewell & Konduri, 1989). However, by contrast with AS, arousal from QS appears to be an order of magnitude less sensitive to depression, as arousal decrements occur only after as many as 100 episodes of severe hypoxia, possibly in association with global cerebral depression (Fewell & Konduri, 1989). Arousal depression also occurs in both states following 1-2 weeks exposure to repeated airway obstruction. However, this arousal depression is explained by sleep disruption rather than hypoxia per se (Brooks et al. 1997).
Sleep deprivation and fragmentation can impair arousal and ventilatory responses to respiratory stimuli (Bowes et al. 1980; Brooks et al. 1997). However, the depression we observed in the present study cannot be attributed to disruption of sleep as the total time spent in AS on the test night was not affected by the imposition of hypoxic testing, though individual AS epochs were slightly shortened. The absence of any compensating rebound of AS on the recovery night suggests further that AS on the test night was largely undisturbed. In any case, the mild disruption of QS (reduced epoch length and total sleep time) was not associated with any diminution in the arousal response to hypoxia in that state.
Continuous hypoxia is well known to depress ventilation, especially in the newborn in which depression emerges rapidly after 1 min of exposure to moderate hypoxia (Rigatto, 1984; Cohen et al. 1997). The effects of repetitive hypoxia, mimicking OSA, are less certain as ventilatory depression has been observed with exposure to repeated, severe hypoxia (Fi,O2= 0.06) but not moderate hypoxia (Fi,O2= 0.10) (Waters et al. 1996). Similarly, Worthington et al. (1993) observed ventilatory depression after 6-8 h exposure to severe hypoxia (Fi,O2= 0.08) repeated at 1 min intervals. Comparison with the present study is complicated as sleep state was not measured in the previous studies of repetitive hypoxia. Importantly, in our study depression of ventilation was selective for sleep state, occurring only in AS. Failure to distinguish sleep state may have obscured a ventilatory depression in AS at moderate levels of hypoxia in the study of Waters et al. (1996).
Characteristically the awake newborn on exposure to hypoxia exhibits small transient increases in blood pressure against a background of significant vasodilatation that occurs in association with compensating increases in heart rate and cardiac output. Subsequently, there is a progressive decrease in heart rate, cardiac output, blood pressure and whole body oxygen consumption (Sidi et al. 1983). Uncertainty exists on the usual cardiovascular response to hypoxia during sleep. We observed significant differences in the responses according to sleep state and the number of repeated exposures to hypoxia. In QS, there was elevation of heart rate and blood pressure that was sustained over the series of exposures. By contrast, in AS, blood pressure increased only in the first 10 exposures and there were no increases of heart rate in this sleep state. Previous studies of hypoxia in the newborn lamb have reported conflicting data on blood pressure and heart rate responses during sleep in the newborn (Fewell et al. 1984; Fewell & Baker, 1987; Fewell & Konduri, 1988). A conflict is also evident in the understanding of whether sleep-related differences exist in baroreflex gain (see Horne et al. 1989). Although these conflicts require clarification, the consistency of the sleep-related differences that we have observed in each of arousal, ventilation and blood pressure, together with their parallel depression during AS, suggests that cardiovascular responses to hypoxia are significantly different in this sleep state compared with wakefulness and QS, with the most prominent difference being the short-lived increment in arterial pressure that disappears with repeated hypoxia.
Acute hypoxaemia is a complex stress that involves integrated adjustments in the respiratory and cardiovascular systems that vary with species, age and the degree of arterial hypoxaemia and its duration (Sidi et al. 1983; Daly, 1986; Weil, 1994). Tachycardia and peripheral vasodilatation that are characteristic of arterial hypoxaemia are to a considerable extent secondary to ventilatory stimulation (Daly, 1986). Overriding these responses there is an overall impairment in AS of integrated circulatory adjustments (Lenzi et al. 1987; Franzini, 1990) and reflex ventilatory stimulation (Henderson-Smart & Read, 1979b;Phillipson & Bowes, 1986; Campbell et al. 1998). The increase in ventilation, blood pressure and heart rate that occurred during QS in our experiments show that, in this state, the responses are consistent with previous concepts which were developed without regard to the state of sleep or wakefulness. Moreover, the poorer ventilatory responses overall, and the absence of tachycardia that we observed during hypoxia in AS are in keeping with known modulations of the classical responses by the imposition of this sleep state. We now report that there is a further depression of ventilation and blood pressure responses in AS that occurs in parallel with the depression of arousal during repeated hypoxia, an observation that has not previously been recognized (Fewell et al. 1988; Harding et al. 1997; Johnston et al. 1998).
Decline in the input from thoracic mechanoreceptors may be particularly important in the depression of arousal as the ventilatory response to hypoxia was high early in the series of exposures and declined significantly thereafter. These receptors normally have a powerful arousing influence when breathing is stimulated (Yasuma et al. 1991). Declines in blood pressure responses in AS possibly arise from declining ventilatory responses to hypoxia (Daly, 1986). That blood pressure responses were depressed after 10 hypoxia exposures uniquely in AS is suggested by three lines of evidence. First, in AS it is only in exposures 1-10 that blood pressure was elevated compared with normoxia. Second, in AS blood pressure responses in exposures 11-20, 21-30 and 31-40 were lower than in exposures 1-10, though this reached significance only for the 11-20 group. That the difference was not significant thereafter, though the tendency remained evident, we would ascribe to the small magnitude of the responses and the well-known variability of AS. Finally, the pattern in AS is in marked contrast to the findings in QS, where blood pressure remained elevated throughout the series of hypoxia exposures, illustrating the distinct nature of the response in AS. Though blood pressure changes were small compared with those occurring in the ventilatory response, baroreceptors appear to have a powerful input to the reticular activating system in the newborn (Horne et al. 1989, 1991) and could have been significant in terms of the overall loss of arousability.
Previously it has been proposed that simultaneous decreases in breathing and metabolic rate represent an appropriate, life-preserving strategy of the newborn mammal faced with hypoxia (Mortola et al. 1989; Weil, 1994). It has also been speculated that by diminishing oxygen requirements when faced with an oxygen deficit the newborn may increase its tolerance of hypoxia in a manner analogous to that of diving and hibernating mammals (Weil, 1994). Operation of this mechanism could be particularly effective in AS as in this state whole body and cerebral utilization of oxygen is elevated compared with QS (Stothers & Warner, 1984; Franzini, 1992). However, the newborn is susceptible to developing more severe hypoxaemia in AS because of reduced lung volume, irregular breathing pattern and diminished intercostal muscle function (Henderson-Smart & Read, 1979a), making arousal from this sleep state a critical defence against developing severe hypoxaemia. The results of our study suggest that impairment of arousal with simultaneous depression of the ventilatory response to hypoxia would exaggerate the extent of desaturation and, paradoxically, expose the newborn to a life-threatening hypoxaemia in AS.
In summary, arousal from AS in response to repeated challenges with hypoxia was depressed after only 10 exposures, while arousal from QS remained unaffected. There was a parallel and sleep state-specific depression of the ventilatory and blood pressure responses to hypoxia, implying that there may be a similar, albeit as yet unknown, mechanism causing depression in both the behavioural and cardio-respiratory systems. The sleep state-specific depression of all these responses suggests that AS is a state of great vulnerability for the sleeping newborn.
Acknowledgments
This work was supported by the National Health and Medical Research Council of Australia, the Sudden Infant Death Research Foundation, Victoria, Australia, the Monash Research Foundation for Mothers and Babies, the National Health and Medical Research Council of Australia, and the Alberta Heritage Foundation for Medical Research. The valuable comments of Dr Philip Berger and technical support of Mrs Jennene Wild and Mr Vojta Brodecky are gratefully acknowledged.
References
- Berthon-Jones M, Sullivan CE. Ventilatory and arousal responses to hypoxia in sleeping humans. American Review of Respiratory Disease. 1982;125:632–639. doi: 10.1164/arrd.1982.125.6.632. [DOI] [PubMed] [Google Scholar]
- Bowes G, Woolf GM, Sullivan CE, Phillipson EA. Effect of sleep fragmentation on ventilatory and arousal responses of sleeping dogs to respiratory stimuli. American Review of Respiratory Disease. 1980;122:899–908. doi: 10.1164/arrd.1980.122.6.899. [DOI] [PubMed] [Google Scholar]
- Brooks D, Horner RL, Kimoff J, Kozar LF, Render-Teixeira CL, Phillipson EA. Effect of obstructive sleep apnea versus sleep fragmentation on responses to airway occlusion. American Journal of Respiratory and Critical Care Medicine. 1997;155:1609–1617. doi: 10.1164/ajrccm.155.5.9154865. [DOI] [PubMed] [Google Scholar]
- Campbell AJ, Bolton DPG, Taylor BJ, Sayers RM. Responses to an increasing asphyxia in infants: effects of age and sleep state. Respiration Physiology. 1998;112:51–58. doi: 10.1016/s0034-5687(98)00008-5. [DOI] [PubMed] [Google Scholar]
- Cohen G, Malcolm G, Henderson-Smart D. A comparison of the ventilatory response of sleeping newborn lambs to step and progressive hypoxaemia. The Journal of Physiology. 1997;503:203–214. doi: 10.1111/j.1469-7793.1997.203bi.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daly M, de B. Interactions between respiration and circulation. In: Cherniack NS, Widdicombe JG, editors. Handbook of Physiology, section 3, The Respiratory System, Control of Breathing. part 2. II. Bethesda, MD, USA: American Physiological Society; 1986. pp. 529–594. chap. 16. [Google Scholar]
- Davies RJO, Stradling JR. Acute effects of obstructive sleep apnoea. British Journal of Anaesthesia. 1993;71:725–729. doi: 10.1093/bja/71.5.725. [DOI] [PubMed] [Google Scholar]
- Fewell JE, Baker SB. Arousal from sleep during rapidly developing hypoxemia in lambs. Pediatric Research. 1987;22:471–477. doi: 10.1203/00006450-198710000-00023. [DOI] [PubMed] [Google Scholar]
- Fewell JE, Johnson P. Acute increases in blood pressure cause arousal from sleep in lambs. Brain Research. 1984;311:259–265. doi: 10.1016/0006-8993(84)90088-x. [DOI] [PubMed] [Google Scholar]
- Fewell JE, Konduri GG. Repeated exposure to rapidly developing hypoxemia influences the interaction between oxygen and carbon dioxide in initiating arousal from sleep in lambs. Pediatric Research. 1988;24:28–33. doi: 10.1203/00006450-198807000-00008. [DOI] [PubMed] [Google Scholar]
- Fewell JE, Konduri GG. Influence of repeated exposure to rapidly developing hypoxaemia on the arousal and cardiopulmonary response to developing hypoxaemia in lambs. Journal of Developmental Physiology. 1989;11:77–82. [PubMed] [Google Scholar]
- Fewell JE, Williams BJ, Hill DE. Sleep does not affect the cardiovascular response to alveolar hypoxia in lambs. Journal of Developmental Physiology. 1984;6:401–405. [PubMed] [Google Scholar]
- Fewell JE, Williams BJ, Szabo JS, Taylor BJ. Influence of repeated upper airway obstruction on the arousal and cardiopulmonary response to upper airway obstruction in lambs. Pediatric Research. 1988;23:191–195. doi: 10.1203/00006450-198802000-00013. [DOI] [PubMed] [Google Scholar]
- Fletcher EC, Lesske J, Behm R, Miller CC, III, Stauss H, Unger T. Carotid chemoreceptors, systemic blood pressure, and chronic episodic hypoxia mimicking sleep apnea. Journal of Applied Physiology. 1992;46:1081–1085. doi: 10.1152/jappl.1992.72.5.1978. [DOI] [PubMed] [Google Scholar]
- Franzini C. The control of the peripheral circulation during sleep. In: Mancia M, Marini G, editors. The Diencephalon and Sleep. New York: Raven Press; 1990. pp. 343–353. [Google Scholar]
- Franzini C. Brain metabolism and blood flow during sleep. Journal of Sleep Research. 1992;1:3–16. [Google Scholar]
- Grant DA, Davidson TL, Fewell JE. Automated scoring of sleep in the neonatal lamb. Sleep. 1995;18:439–445. doi: 10.1093/sleep/18.6.439. [DOI] [PubMed] [Google Scholar]
- Harding R, Jakubowska AE, McCrabb GJ. Arousal and cardio-respiratory responses to airflow obstruction in sleeping lambs: effect of sleep state, age, and repeated obstruction. Sleep. 1997;20:693–701. doi: 10.1093/sleep/20.9.693. [DOI] [PubMed] [Google Scholar]
- Henderson-Smart DJ, Read DJC. Reduced lung volume during behavioural active sleep in the newborn. Journal of Applied Physiology. 1979a;46:1081–1085. doi: 10.1152/jappl.1979.46.6.1081. [DOI] [PubMed] [Google Scholar]
- Henderson-Smart DJ, Read DJC. Ventilatory responses to hypoxaemia during sleep in the newborn. Journal of Developmental Physiology. 1979b;1:195–208. [PubMed] [Google Scholar]
- Heymans J-F, Heymans C. Sur les modifications directes et sur la régulation réflexe de l'activité du centre respiratoire de la tête isolée du chien. Archives Internationales de Pharmacodynamie et de Therapie. 1927;33:272–370. [Google Scholar]
- Horne RSC, Berger PJ, Bowes G, Walker AM. Effect of sinoaortic denervation on arousal responses to hypotension in newborn lambs. American Journal of Physiology. 1989;256:H434–440. doi: 10.1152/ajpheart.1989.256.2.H434. [DOI] [PubMed] [Google Scholar]
- Horne RSC, Berger PJ, De Preu ND, Walker AM. Arousal responses to hypertension in lambs: effect of sinoaortic denervation. American Journal of Physiology. 1991;260:H1283–1289. doi: 10.1152/ajpheart.1991.260.4.H1283. [DOI] [PubMed] [Google Scholar]
- Jeffery HE, Read DJC. Ventilatory responses of newborn calves to progressive hypoxia in quiet and active sleep. Journal of Applied Physiology. 1980;48:892–895. doi: 10.1152/jappl.1980.48.5.892. [DOI] [PubMed] [Google Scholar]
- Johnston RV, Grant DA, Wilkinson MH, Walker AM. Repetitive hypoxia rapidly depresses arousal from active sleep in newborn lambs. The Journal of Physiology. 1998;510:651–659. doi: 10.1111/j.1469-7793.1998.651bk.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kahn A, Groswasser J, Rebuffat E, Sottiaux M, Blum D, Foerster M, Franco P, Bochner A, Alexander M, Bachy A, Richard P, Verghote M, Le Polain D, Wayenberg JL. Sleep and cardio-respiratory characteristics of infant victims of sudden death: a prospective case-control study. Sleep. 1992;15:287–292. doi: 10.1093/sleep/15.4.287. [DOI] [PubMed] [Google Scholar]
- Lenzi P, Cianci T, Guidalotti PL, Leonardi GS, Franzini C. Brain circulation during sleep and its relation to extracerebral hemodynamics. Brain Research. 1987;415:14–20. doi: 10.1016/0006-8993(87)90264-2. [DOI] [PubMed] [Google Scholar]
- Mortola JP, Rezzonico R, Lanthier C. Ventilation and oxygen consumption during acute hypoxia in newborn mammals: a comparative analysis. Respiration Physiology. 1989;78:31–43. doi: 10.1016/0034-5687(89)90140-0. [DOI] [PubMed] [Google Scholar]
- Neubauer JA, Santiago TV, Edelman NH. Hypoxic arousal in intact and chemodenervated sleeping cats. Journal of Applied Physiology. 1981;51:1294–1299. doi: 10.1152/jappl.1981.51.5.1294. [DOI] [PubMed] [Google Scholar]
- Phillipson EA, Bowes G. Control of breathing during sleep. In: Cherniack NS, Widdicombe JG, editors. Handbook of Physiology, section 3, The Respiratory System, Control of Breathing. part 2. II. Bethesda, MD, USA: American Physiological Society; 1986. pp. 649–689. chap. 19. [Google Scholar]
- Phillipson EA, Sullivan CE. Arousal: the forgotten response to respiratory stimuli. American Review of Respiratory Disease. 1978;118:807–809. doi: 10.1164/arrd.1978.118.5.807. [DOI] [PubMed] [Google Scholar]
- Phillipson EA, Sullivan CE, Read DJC, Murphy E, Kozar LF. Ventilatory and arousal responses to hypoxia in sleeping dogs. Journal of Applied Physiology. 1978;44:512–520. doi: 10.1152/jappl.1978.44.4.512. [DOI] [PubMed] [Google Scholar]
- Rigatto H. Control of ventilation in the newborn. Annual Review of Physiology. 1984;46:661–674. doi: 10.1146/annurev.ph.46.030184.003305. [DOI] [PubMed] [Google Scholar]
- Rognum TO, Saugstad OD. Hypoxanthine levels in vitreous humor: evidence of hypoxia in most infants who died of sudden infant death syndrome. Pediatrics. 1991;87:306–310. [PubMed] [Google Scholar]
- Shepard JW., Jr . Hemodynamics in obstructive sleep apnea. In: Fletcher EC, editor. Abnormalities of Respiration during Sleep: Diagnosis, Pathophysiology, and Treatment. Orlando, FL, USA: Grune & Stratton; 1986. pp. 39–61. [Google Scholar]
- Sidi D, Kuipers JRG, Teitel D, Heymann MA, Rudolph AM. Developmental changes in oxygenation and circulatory responses to hypoxemia in lambs. American Journal of Physiology. 1983;245:H674–682. doi: 10.1152/ajpheart.1983.245.4.H674. [DOI] [PubMed] [Google Scholar]
- Stothers JK, Warner RM. Thermal balance and sleep state in the newborn. Early Human Development. 1984;9:313–322. doi: 10.1016/0378-3782(84)90076-8. [DOI] [PubMed] [Google Scholar]
- Waters KA, Beardsmore CS, Paquette J, Meehan B, Cote A, Moss IR. Respiratory responses to rapid-onset, repetitive vs. continuous hypoxia in piglets. Respiration Physiology. 1996;105:135–142. doi: 10.1016/0034-5687(96)00046-1. [DOI] [PubMed] [Google Scholar]
- Weil JV. Ventilatory response to CO2 and hypoxia after sustained hypoxia in awake cats (editorial) Journal of Applied Physiology. 1994;76:2251–2252. doi: 10.1152/jappl.1994.76.6.2251. [DOI] [PubMed] [Google Scholar]
- Worthington JM, Hedner JA, Sullivan CE. Depressed hypoxic ventilatory responsiveness after prolonged intermittent hypoxia in the rat. In: Walker AM, McMillen C, editors. Second SIDS International Conference and First SIDS Global Strategy Meeting. New York: Perinatology Press; 1993. pp. 238–242. National SIDS Council. [Google Scholar]
- Yasuma F, Kozar LF, Kimoff RJ, Bradley TG, Phillipson EA. Interaction of chemical and mechanical respiratory stimuli in the arousal response to hypoxia in sleeping dogs. American Review of Respiratory Disease. 1991;143:1274–1277. doi: 10.1164/ajrccm/143.6.1274. [DOI] [PubMed] [Google Scholar]