

The universal genetic code encodes 19 amino acids and just one N-alkyl amino acid (“imino acid”), proline (Pro). This raises some interesting questions. Given that several N-alkyl amino acids are major products of prebiotic synthetic experiments and meteorite analyses, why did Nature select Pro?1 Is Pro incorporated into protein less efficiently than an amino acid by the translation apparatus? Are other N-alkyl amino acids excluded from incorporation, for example, by proofreading? Can translation be engineered to incorporate other N-alkyl amino acids efficiently? Answers bear on plans to translate mRNA templates into libraries of N-alkyl amino acid polymers for genetic selection of drug leads.2 N-Alkylation is desirable because it can confer the important pharmacological properties of protease resistance, membrane permeability, and even oral availability (e.g., cyclosporin A).
The above questions have been investigated experimentally using chemoenzymatically synthesized N-alkyl aminoacyl-transfer RNAs (tRNAs) and in vitro translation systems. Efficiencies are typically reported as the yield of product with an unnatural aminoacyl-tRNA in comparison with the control yield of 100% from all wild-type aminoacyl-tRNAs. Results can be summarized as follows. Pro was incorporated at 46% efficiency from an unnatural tRNA that could incorporate Ala at 28%.3 Pro analogues ranged in incorporation efficiencies from 43% to undetectable.3–5 N-Methyl amino acids ranged from 72% to undetectable.3,4,6,7 N-Ethyl alanine incorporation was undetectable.3 It is difficult to draw general conclusions from these results because of the wide range of reported efficiencies and the paucity of experiments. Interpretation may be complicated by the large number of variables between the experimental systems: prokaryotic versus eukaryotic, presence of competitors and editing by aminoacyl-tRNA synthetases in crude systems versus absence in synthetase-free pure systems, tRNA adaptors that are natural versus methylated natural versus unnatural, tRNA anticodons that are natural versus unnatural, different messenger RNA (mRNA) codons for incorporation, different amino acids preceding incorporation, and requirement for amino acid incorporation downstream or not.
Of the various translation systems available for comparison of amino acid analogues, a purified E. coli system free of aminoacyl-tRNA synthetases2 is highly suitable. It is well-defined, flexible, and incorporates unnatural amino acids selectively and efficiently. We thus used this system to compare the incorporation of amino/N-alkyl amino acids (Figure 1A). To examine the effects of different codons, anticodons, and tRNA bodies, we constructed two new tRNA adaptors with wild-type anticodons to compare with our original E. coli tRNAAsn-based unmodified tRNA (termed tRNAAsnB).8 They are based on two other unmodified E. coli tRNAs,9 termed tRNAPheB and tRNAAlaB (Figure 2, left). Synthetic amino/N-alkyl aminoacyl-tRNAs were assayed in a full translation cycle by programming incorporation of 3H-Val immediately downstream (Figure 1B).
Figure 1.
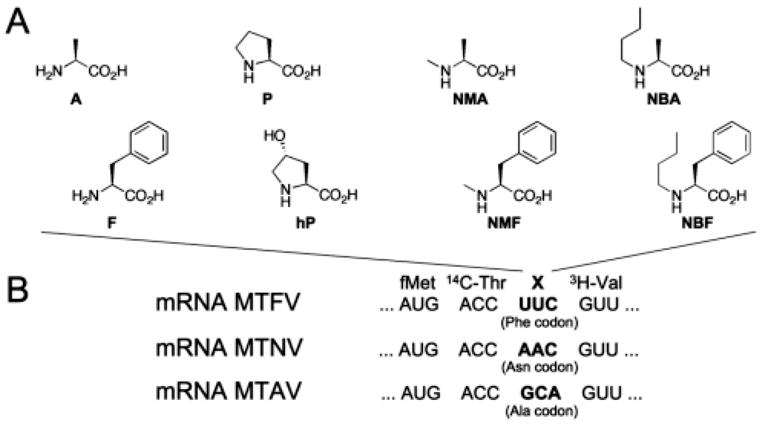
The 24 translation assays. (A) l-amino acids and N-alkyl-l-amino acids synthesized in NVOC-amino-protected form on the dinucleotide pdCpA for ligation onto tRNAminusCA transcripts (see Supporting Information). A, Ala; F, Phe; P, Pro; hP, 4-hydroxy-Pro; NMA, N-methyl-Ala; NMF, N-methyl-Phe; NBA, N-butyl-Ala; NBF, N-butyl-Phe. (B) Coding sequences of mRNAs: f, formyl; X, amino/N-alkyl amino acid from (A).
Figure 2.
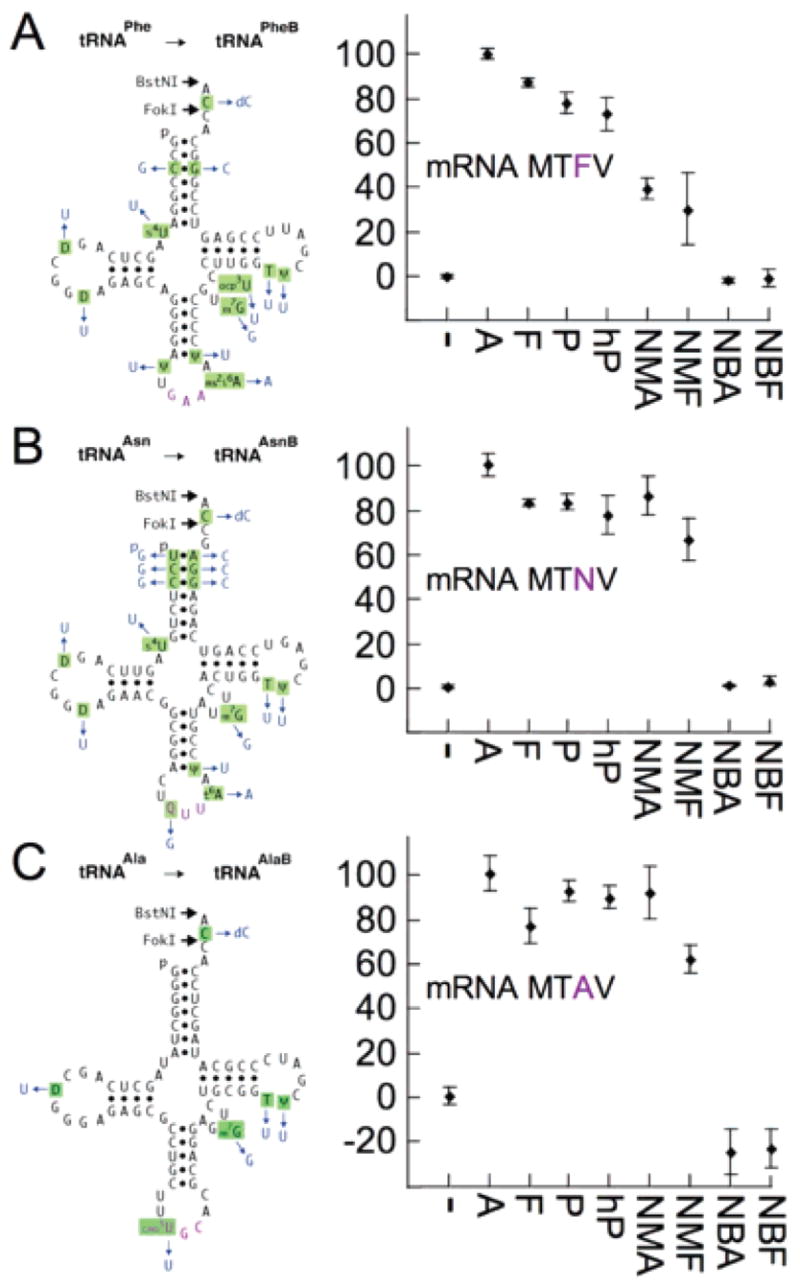
Efficiencies of incorporation of amino/N-alkyl amino acids from synthetic tRNAs into peptide tetramers in the 24 assays of Figure 1. Left: natural E. coli tRNAs (black with purple anticodons) and their synthetic unmodified counterparts (changes in blue). Right: yields of peptide tetramers in pure, synthetase-free translations incubated at 37 °C for 40 min. (−) Background counts defined by omitting cognate aminoacyl-tRNAs for the third codons. Addition of wild-type Phe-tRNAPhe in (A) gave the positive control (95 ± 5%; not plotted). Ala-tRNAPheB gave 100 ± 2%, and the other two Ala-charged unmodified tRNAs also saturated tetrapeptide synthesis (normalized to 100%). Yields with all other substrates were calculated relative to their respective Ala-tRNAs. Standard deviations of quadruplicate experiments are shown. See Supporting Information.
As a positive control, we translated mRNA MTFV into fMTFV using all-natural tRNAs precharged in vitro with all-natural amino acids by aminoacyl-tRNA synthetases. The yield of this peptide (Figure 2, legend) was essentially the same as the yields when natural Phe-tRNAPhe was substituted with our chemoenzymatically synthesized Phe-tRNAPheB or Ala-tRNAPheB (Figure 2A, A and F on the x-axis). This validated the ability of certain synthetic substrates to saturate the assay.
Ala, Phe, Pro, and 4-Hydroxy-Pro
The 12 incorporation efficiencies (Figure 2, right) ranged from quantitative to very efficient, with the cyclic secondary amino acids being comparable with the primary amino acids. All incorporation efficiencies were substantially greater than those reported for amino acids from suppressor tRNAs in crude translation systems,10 presumably due to a lack of competitors in our system.
N-Methyl Amino Acids
For N-Me-Ala, incorporation was very efficient from tRNAAsnB and tRNAAlaB but only intermediate from tRNAPheB (Figure 2, right). For N-Me-Phe, incorporation was intermediate from all three tRNAs. This demonstrates that essentially quantitative incorporation of an N-Me amino acid is possible, provided that it is matched with a suitable tRNA adaptor and used in a purified system. It has been speculated that the natural pairings of amino acids and their cognate tRNAs may be optimal for delivery by the aminoacyl-tRNA carrier protein, elongation factor Tu, and for incorporation by the ribosome.11 Our data with unmodified tRNAs, together with our earlier intermediate efficiency incorporations of these same N-Me amino acids from two other tRNAAsnB adaptors,7 indicate that the pairing is particularly important for N-Me amino acids. Results are difficult to predict: our expectations that tRNAPheB would be the best adaptor for Phe and N-Me-Phe and that tRNAAlaB would be the best adaptor for Ala and N-Me-Ala were not fully borne out. Data are consistent with the rather varied incorporation efficiencies from different tRNA bodies reported for N-Me amino acids3,4,6,7 and other amino acids.12
N-Butyl Amino Acids
Incorporations were undetectable (limit of sensitivity) 5%; Figure 2, right). This was surprising given the lack of competing cognate incorporations but is consistent with the failure to incorporate N-ethyl alanine in a crude system.3 Note that with the mRNA MTAV, measured dpm was paradoxically less than the minus cognate tRNA control. This is because the 3H-Val incorporation for the minus cognate tRNA control was unexpectedly higher than for the minus mRNA control due to missense translation, and the N-Bu aminoacyl-tRNAs inhibited missense translation by 4-fold. Translation inhibition was not nonspecific because 14C-Thr incorporation was unaffected, indicating that N-Bu aminoacyl-tRNAs can bind to the ribosomal A site. Direct evidence for codon-specific binding of an N-Bu aminoacyl-tRNA was obtained by 3 min ribosome binding assays (Figure S1 in Supporting Information).
Returning to our original questions, is Pro a privileged N-alkyl amino acid for incorporation by the translation apparatus? In support of this, three different tRNA bodies gave efficient incorporations for 12/12 amino acid/Pro/hydroxy-Pro assays but only for 4/12 incorporations for N-Me and N-Bu amino acids. It is hard to explain this preference by arguing that translation co-evolved with Pro or Pro-tRNAPro because the structures of the 20 natural amino/N-alkyl amino acids differ greatly and we used three different tRNA bodies. An alternative explanation is that Pro is more chemically reactive than N-Me amino acids and much more reactive than N-Bu amino acids. The five-membered Pro ring is less hindered than the freely rotating N-Me, which is less hindered than the larger N-Bu. Indeed, solid-phase peptide synthesis couples Pro more efficiently than N-Me amino acids and other bulky amino acid analogues,13 and Pro analogues and N-Me amino acids are the most common N-alkyl amino acids incorporated by cellular nonribosomal peptide synthetases.14 If the incorporation preferences for our purified E. coli system are in fact governed by the innate chemical reactivities of various N-alkyl amino acids, analogous preferences would be predicted for the translation machinery of every organism. Our data suggest that many N-alkyl amino acids in the cell may be excluded from incorporation into proteins by the translation apparatus, even though some can be charged onto tRNAs by aminoacyl-tRNA synthetases.15 Finally, the data support exploration of Pro analogues and N-Me amino acids, each charged on several tRNAs, as substrates for engineering ribosomal synthesis of genetically selectable libraries of protease-resistant, N-alkyl, peptide ligands.
Supplementary Material
Supporting Information Available: Figure S1 and additional experimental details. This material is available free of charge via the Internet at http://pubs.acs.org.
Acknowledgments
A.C.F.’s work began in the Department of Pathology, Brigham and Women’s Hospital, Harvard Medical School, under the exceptional mentorship of Dr. Stephen Blacklow. We thank Drs. Michael Pavlov and Måns Ehrenberg for advice and materials. This work was supported by NIH grants (to A.C.F. and V.W.C.), American Cancer Society grants (to A.C.F.), and the Swiss National Foundation (to L.G.D.).
References
- 1.Weber AL, Miller SL. J Mol Evol. 1981;17:273–84. doi: 10.1007/BF01795749. [DOI] [PubMed] [Google Scholar]
- 2.(a) Forster AC, Weissbach H, Blacklow SC. Anal Biochem. 2001;297:60–70. doi: 10.1006/abio.2001.5329. [DOI] [PubMed] [Google Scholar]; (b) Tan Z, Blacklow SC, Cornish VW, Forster AC. Methods. 2005;36:279–90. doi: 10.1016/j.ymeth.2005.04.011. [DOI] [PubMed] [Google Scholar]
- 3.Ellman JA, Mendel D, Schultz PG. Science. 1992;255:197–200. doi: 10.1126/science.1553546. [DOI] [PubMed] [Google Scholar]
- 4.(a) Chung HH, Benson DR, Schultz PG. Science. 1993;259:806–9. doi: 10.1126/science.8430333. [DOI] [PubMed] [Google Scholar]; (b) Karginov VA, Mamaev SV, An H, Van Cleve MD, Hecht SM, Komatsoulis GA, Abelson JN. J Am Chem Soc. 1997;119:8166–76. [Google Scholar]
- 5.(a) Josephson K, Hartman MCT, Szostak JW. J Am Chem Soc. 2005;127:11727–35. doi: 10.1021/ja0515809. [DOI] [PubMed] [Google Scholar]; (b) Lummis SCR, Beene DL, Lee LW, Lester HA, Broadhurst RW, Dougherty DA. Nature. 2005;438:248–52. doi: 10.1038/nature04130. [DOI] [PubMed] [Google Scholar]; (c) Choudhury AK, Golovine SY, Dedkova LM, Hecht SM. Biochemistry. 2007;46:4066–76. doi: 10.1021/bi062042r. [DOI] [PubMed] [Google Scholar]
- 6.(a) Bain JD, Wacker DA, Kuo EE, Chamberlin AR. Tetrahedron. 1991;47:2389–400. [Google Scholar]; (b) Frankel A, Millward SW, Roberts RW. Chem Biol. 2003;10:1043–50. doi: 10.1016/j.chembiol.2003.11.004. [DOI] [PubMed] [Google Scholar]; (c) Merryman C, Green R. Chem Biol. 2004;11:575–82. doi: 10.1016/j.chembiol.2004.03.009. [DOI] [PubMed] [Google Scholar]
- 7.Tan Z, Forster AC, Blacklow SC, Cornish VW. J Am Chem Soc. 2004;126:12752–3. doi: 10.1021/ja0472174. [DOI] [PubMed] [Google Scholar]
- 8.Forster AC, Tan Z, Nalam MNL, Lin H, Qu H, Cornish VW, Blacklow SC. Proc Natl Acad Sci USA. 2003;100:6353–7. doi: 10.1073/pnas.1132122100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.(a) Harrington KM, Nazarenko IA, Dix DB, Thompson RC, Uhlenbeck OC. Biochemistry. 1993;32:7617–22. doi: 10.1021/bi00081a003. [DOI] [PubMed] [Google Scholar]; (b) Ma C, Kudlicki W, Odom OW, Kramer G, Hardesty B. Biochemistry. 1993;32:7939–45. doi: 10.1021/bi00082a015. [DOI] [PubMed] [Google Scholar]
- 10.Cornish VW, Mendel D, Schultz PG. Angew Chem, Int Ed Engl. 1995;34:621–33. [Google Scholar]
- 11.LaRiviere FJ, Wolfson AD, Uhlenbeck OC. Science. 2001;294:165–8. doi: 10.1126/science.1064242. [DOI] [PubMed] [Google Scholar]
- 12.Cload ST, Liu DR, Froland WA, Schultz PG. Chem Biol. 1996;3:1033–8. doi: 10.1016/s1074-5521(96)90169-6. [DOI] [PubMed] [Google Scholar]
- 13.Humphrey JM, Chamberlin AR. Chem Rev. 1997;97:2243–66. doi: 10.1021/cr950005s. [DOI] [PubMed] [Google Scholar]
- 14.Nonribosomal peptide amino acids are listed at http://bioinfo.lifl.fr/norine/.
- 15.Hartman MCT, Josephson K, Szostak JW. Proc Natl Acad Sci USA. 2006;103:4356–61. doi: 10.1073/pnas.0509219103. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supporting Information Available: Figure S1 and additional experimental details. This material is available free of charge via the Internet at http://pubs.acs.org.