

Abstract
Hematopoietic stem cell transplantation (HSCT) requires conditioning treatments such as irradiation, which leads to a severely delayed recovery of T cell immunity and constitutes a major complication of this therapy. Currently, our understanding of the mechanisms regulating thymic recovery is limited. It is known that a subpopulation of bone marrow (BM)–derived thymic immigrant cells and the earliest intrathymic progenitors express the FMS-like tyrosine kinase 3 (Flt3) receptor; however, the functional significance of this expression in the thymus is not known. We used the BM transplant model to investigate the importance of Flt3 ligand (FL) for the regeneration of the T cell compartment. We show that FL is expressed in the adult mouse thymus on the surface of perivascular fibroblasts. These cells surround the proposed thymic entry site of Flt3 receptor–positive T cell progenitors. After irradiation, perivascular FL expression is up-regulated and results in an enhanced recovery of thymic cellularity. Thymic grafting experiments confirm an intrathymic requirement for FL. Collectively, these results show that thymic stromal cell–mediated FL–Flt3 receptor interactions are important in the reconstitution of thymopoiesis early after lethal irradiation and HSCT, and provide a functional relevance to the expression of the Flt3 receptor on intrathymic T cell progenitors.
Total body irradiation is part of the conditioning regimen necessary for hematopoietic stem cell transplantation (HSCT), which constitutes a life-saving therapy for several malignant and nonmalignant diseases. However, total body irradiation results in severe thymic atrophy and an associated peripheral T cell hypoplasia that may take up to 3 yr from which to fully recover. During this time of compromised immunity, HSCT recipients are at an increased risk of opportunistic infections and tumor relapse. Although the initial homeostatic expansion of transferred donor T cells may contribute to the peripheral T cell compartment, the recovery of a diverse repertoire of naive T cells is strictly dependent on efficient thymic function (1).
The thymus is composed of both lymphoid and nonlymphoid cells. The latter constitute the thymic stroma and comprise thymic epithelial cells (TECs) and nonepithelial cells such as fibroblasts and endothelium. Combined, thymic stromal cells play an essential role in T cell development, as they provide microenvironments that facilitate the entry of progenitor cells into the thymus, their intrathymic migration, and differentiation via well-defined stages to mature T cells (2,3).
As the thymus does not contain hematopoietic cells with a self-renewing potential, continued thymic T cell production necessitates a constant immigration of bloodborne precursor cells. Although proposed to originate from BM-resident precursor cells devoid of lineage-specific markers but positive for Sca1 and c-kit (lin−Sca1+c-kit+ [LSK]), the specific differentiation stage of precursors that enter the thymus at the corticomedullary junction remains controversial (4). Immigration of T cell progenitors constitutes an active process, which necessitates the presence of P-selectin within the thymic microenvironment, and engagement of chemokine receptors such as CXCR4 and CCR9 and adhesion molecules CD44 and integrin α4 on T cell progenitors (4). However, neither the identity nor the spatial position of stromal cells that express the respective cognate molecules to these receptors have been precisely defined. Once within the thymus, the newly immigrated T cell precursor cells are termed early thymic progenitors (ETPs), phenotypically characterized as lin−c-kit+CD25−. ETPs undergo extensive in situ expansion and differentiation that eventually give rise to functionally competent and phenotypically mature T cells (4, 5). This process is critically dependent on the stromal microenvironment for the expression of Notch ligands (6, 7), IL-7 (8, 9), and stem cell factor (10).
FMS-like tyrosine kinase 3 (Flt3) is expressed on BM and peripheral blood (PB) progenitor cells that can give rise to T cells under physiological conditions (11, 12). Flt3 is also expressed on a subpopulation of ETPs, and it is thought that Flt3+ ETPs represent the earliest defined intrathymic progenitors (13). It is currently unknown whether Flt3 expression by intrathymic T cell progenitors has a functional significance. Flt3 ligand (FL) is expressed in the BM, where it stimulates hematopoiesis (14); however, a role for FL in the thymus has not previously been defined. Mice deficient in the expression of either Flt3 or FL display defects in very early T cell development (11, 13) comparable to mice rendered deficient for P-selectin glycoprotein ligand 1 and for CXCR4. Hence, the similarities in phenotypes between these different mouse strains, as well as the expression pattern of Flt3, suggest that FL may also play a role in early T cell development.
FL has previously been proposed to contribute to hematopoietic recovery in the context of HSCT, as its serum concentrations are greatly increased after myeloablative conditioning in both mice and humans (14). However, the systemic provision of FL in either mice or humans has provided inconclusive findings regarding its role in thymic T cell recovery. FL strongly enhances the circulating stem cell number in humans (15), but no discernable effect on thymic function was reported. FL given systematically to mice increased thymic output (16, 17), which was possibly but not necessarily the consequence of an expansion of BM-derived T cell progenitors. The recruitment of LSK cells to PB appears to be unaffected by the lack of FL in FL−/− mice, yet Flt3 contributed to the overall efficiency of early thymopoiesis; however, no evidence of intrathymic Flt3/FL signaling was reported (13). Therefore, the biological significance of FL expression outside the BM microenvironment, if any, is still largely undefined.
In this report, we investigated a role for FL expression in the recovery of thymopoiesis after irradiation and BM transplantation (BMT) in mice. We found that FL was expressed by thymic perivascular fibroblasts at low levels under steady-state conditions, and FL was rapidly up-regulated on these cells after irradiation. This higher availability of FL correlated with an enhanced recovery of thymic cellularity.
RESULTS AND DISCUSSION
FL is expressed around thymic blood vessels at low levels in wild-type mice and at high levels in Flt3 receptor–deficient (Flt3−/−) mice
The Flt3 receptor is expressed on T cell progenitors in the BM and on a subset of ETPs considered to represent the most immature intrathymic progenitors (13). These findings prompted us to examine the thymus for the nature and spatial organization of the cells that express the corresponding FL. For this purpose, adult B6 and Flt3−/− mice were first analyzed for expression of FL under steady-state conditions. Using immunohistochemistry (IHC), FL expression was detected in both mouse strains on perivascular fibroblasts (ER-TR7+ and MTS15+) that are positioned at the corticomedullary junction between endothelial (CD31+) and TEC layers (cytokeratin 5+; Fig. 1, A and B). Wild-type B6 mice had a low expression of FL, whereas Flt3−/− mice displayed higher concentrations of FL.
Figure 1.

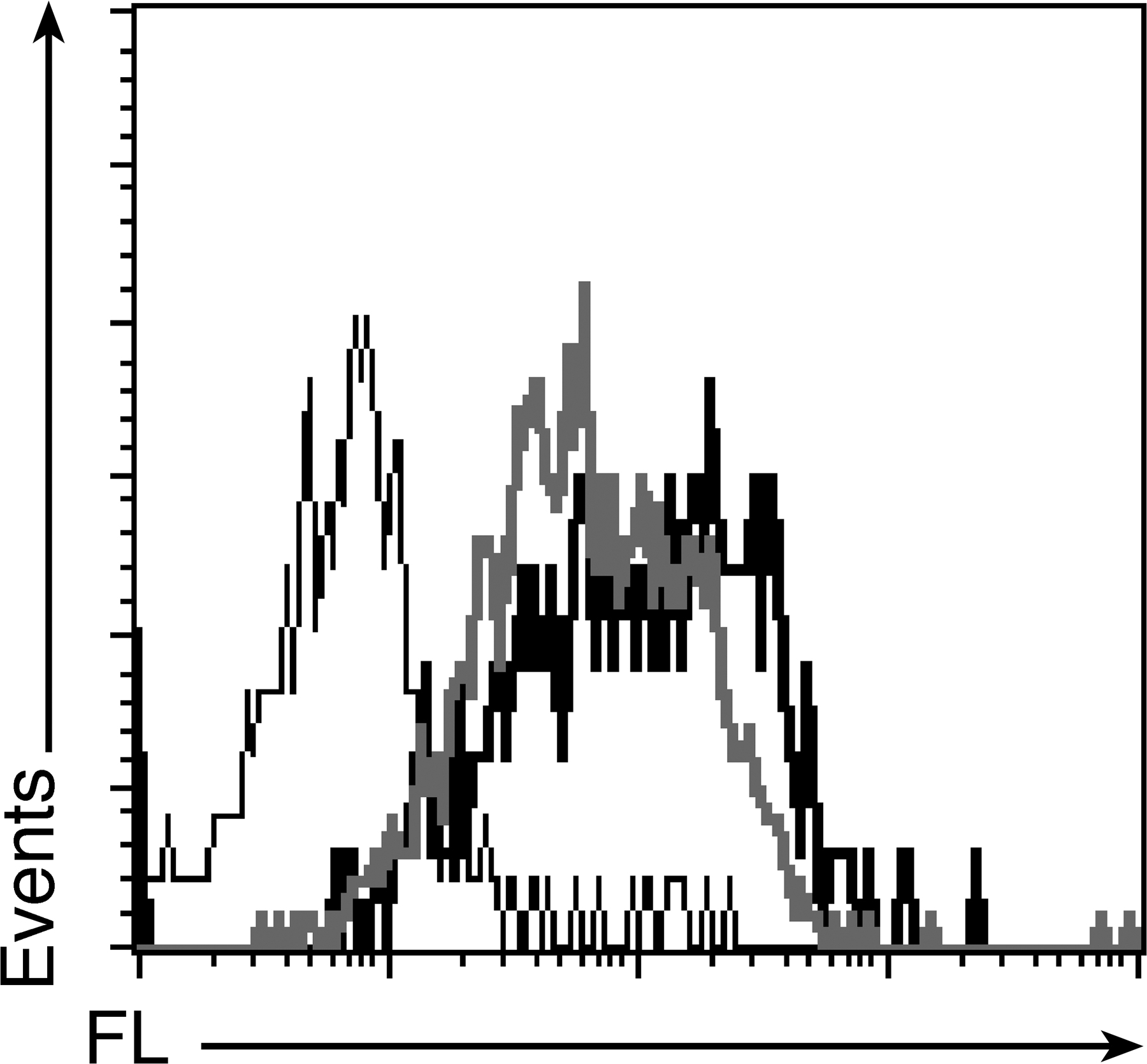
FL is expressed at low levels by thymic perivascular fibroblasts in the B6 mouse and at high levels in the Flt3−/− mouse. (A) IHC of B6, FL−/−, and Flt3−/− mouse thymi. (top) FL only (green). (bottom) FL and ER-TR7 (red) to visualize blood vessels. Bars, 60 μM. (B) IHC of Flt3−/− mouse thymus. (top) FL alone (green). (bottom) FL and staining with a secondary antibody (red), as indicated. MTS15 recognizes perivascular fibroblasts, CD31 endothelial cells, and cytokeratin 5 (K5) medullary TECs. Similar results were obtained with at least five mice for each genotype. Bars, 60 μM. (C) FCM analysis of FL expression on thymic stromal cell subsets of B6, Flt3−/−, and FL−/− mice. TECs are defined as CD45−EpCAM+, nonepithelial stromal cells as CD45−EpCAM−, and thymic fibroblasts as MTS15+CD45−EpCAM− (MTS15+). FL expression (bolded line) and isotype (continuous line) are shown. Numbers in plots indicate the MFIR (MFI of FL/MFI of isotype control). All experiments were repeated at least three times with cells purified from different mice (all aged 8 wk).
As FL can be retained intracellularly (14), we determined whether thymic FL observed by IHC is cell-surface expressed by flow cytometry (FCM) in Flt3−/− mice. As shown in Fig. 1 C, FL was expressed on the surface of non-TEC stromal cells (CD45− epithelial cell adhesion molecule [EpCAM]−) and, specifically, on perivascular fibroblasts (MTS15+). Gating of stromal populations is shown in Fig. S1 (available at http://www.jem.org/cgi/content/full/jem.20072065/DC1). These findings closely paralleled IHC results. An identical FL expression pattern was also observed in the B6 thymus at a lower intensity, with a mean fluorescence intensity ratio (MFIR) of 1.8 compared with 15 on MTS15+ cells from Flt3−/− mice (Fig. 1 C), underscoring the physiological expression of FL by non-TEC stromal cells, including perivascular fibroblasts.
FL expression is up-regulated on thymic stromal cells after irradiation
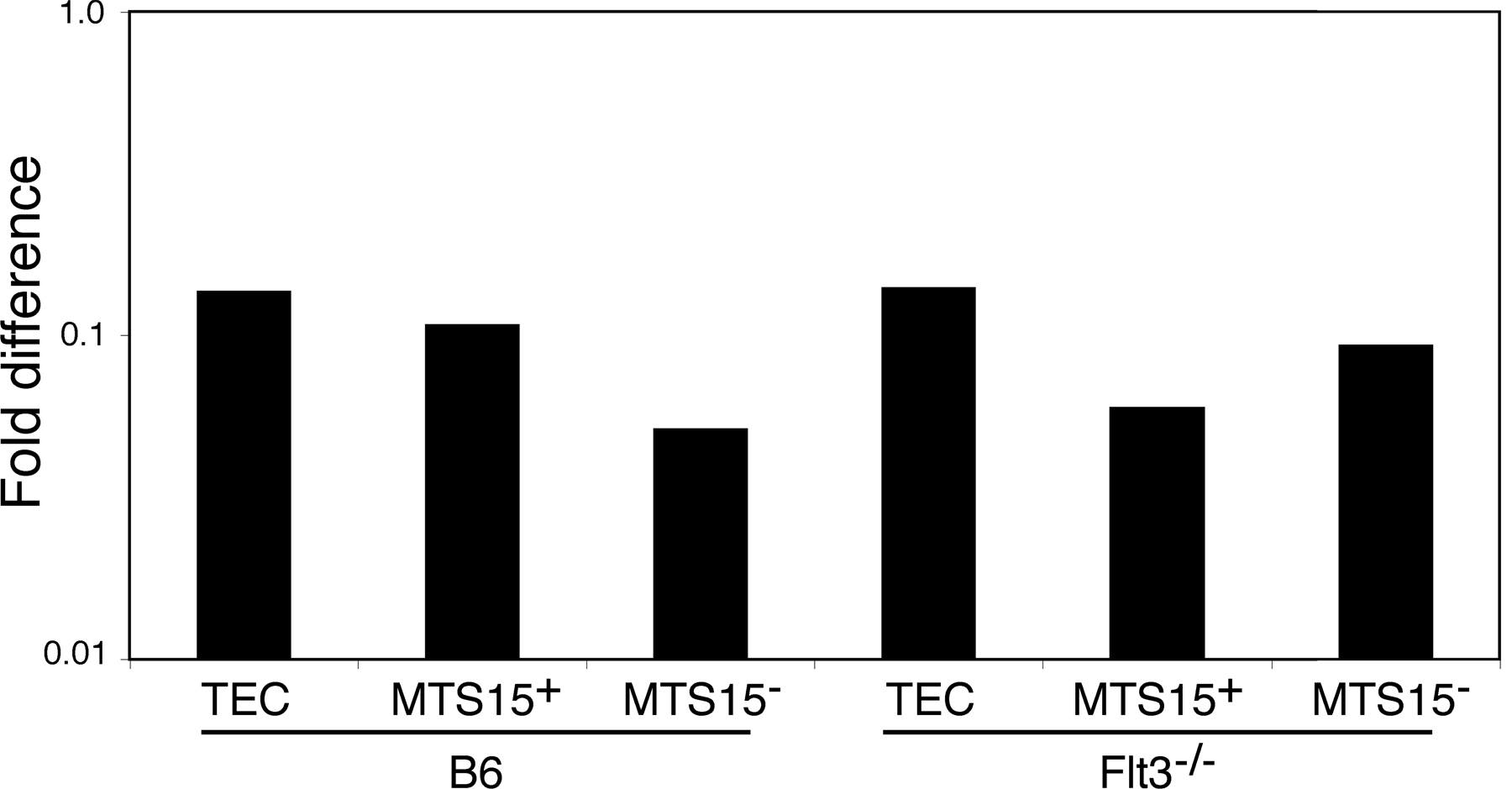
Myeloablative irradiation largely depletes the thymus of lymphoid cells and also damages the cellular composition and organization of thymic stroma (18). To assess the impact of irradiation on FL expression, thymic stromal cells from sublethally or lethally irradiated B6 mice were analyzed by IHC and FCM. According to IHC, 4 d after sublethal irradiation FL expression was up-regulated around thymic blood vessels, displaying an expression pattern and intensity similar to that of Flt3−/− mice (compare Fig. 2 [A–D] with Fig. 1 [A and B]). FCM revealed increased concentrations of FL on the surface of non-TEC stromal cells (CD45−EpCAM−), particularly MTS15+ fibroblasts after both sublethal and lethal irradiation (Fig. 2 E). The up-regulation of FL on B6 thymic stromal cells reached a peak 4–5 d after sublethal irradiation and returned to pretreatment values by day 7 (Fig. 2 F). The transient increase in FL expression in B6 mice correlated with the period of decreased thymic cellularity (Fig. 2 G). The constitutively high level of FL expression observed in Flt3−/− mice did not increase after irradiation (Fig. S2, available at http://www.jem.org/cgi/content/full/jem.20072065/DC1). FL mRNA was found not to increase after irradiation in stromal cells of both B6 and Flt3−/− mice, supporting a posttranslational control of FL expression (Fig. S3). Collectively, these results show that FL is expressed in B6 mice at low levels on the surface of thymic stromal cells, including perivascular fibroblasts, and that this expression is up-regulated after lethal and sublethal irradiation.
Figure 2.

FL is up-regulated by perivascular fibroblasts in the B6 mouse after irradiation. (A–D) IHC analysis of FL expression in the B6 mouse 4 d after sublethal irradiation. (top) FL alone (green). (bottom) FL with a secondary antibody (red), as indicated. Bars, 60 μM. (A) MTS15, (B) ER-TR7, (C) CD31, and (D) K5. Similar results were obtained with at least 5 mice for each genotype. (E) FCM analysis of FL expression on B6 thymic stromal cells, untreated and at 4 d after sublethal or lethal irradiation, as indicated. Thymic populations are defined as in Fig. 1. FL expression (bolded line) and isotype (continuous line) are shown. Numbers in plots indicate MFIR. All experiments were repeated at least three times with cells purified from different mice (all aged 8 wk). (F) Temporal expression of FL on thymic stroma of the B6 mouse after sublethal irradiation. Plots show FL expression on MTS15+ nonepithelial stromal cells at the time points indicated after irradiation. FL expression (bolded line) and isotype (continuous line) are shown. Numbers in plots indicate MFIR. Similar results were obtained from at least three different mice at each time point indicated. (G) Thymus cellularity in B6 mice after sublethal irradiation at the time points indicated after irradiation.
Thymic stromal FL expression enhances the recovery of thymopoiesis after irradiation
To investigate a specific effect of FL expression by thymic stromal cells on thymopoiesis, BM chimeras were generated in which transplanted wild-type B6 precursor cells developed in irradiated B6, Flt3−/−, or FL−/− recipients. The presence of donor-derived cells (CD45.1+) was analyzed in the different recipients (CD45.2+) at several time points after BMT.
At 10 d after conditioning and BMT, both Flt3−/− and the FL−/− recipient mice had comparable levels of donor-derived LSK cells in the BM and PB, which were significantly lower than in transplanted B6 mice (Fig. S4 A, available at http://www.jem.org/cgi/content/full/jem.20072065/DC1). The frequency of donor-type lineage-negative Flt3+ cells was also significantly reduced in both FL−/− and Flt3−/− mice (Fig. S4 B) despite a high level of soluble FL in the sera of Flt3−/− mice (not depicted). This may be caused by a lower engraftment efficiency in the Flt3−/− mouse or to a reduced bioavailability of the soluble compared with the membrane-bound form of FL, as has been suggested previously for FL (19) and is the case with the closely related c-kit ligand (20).
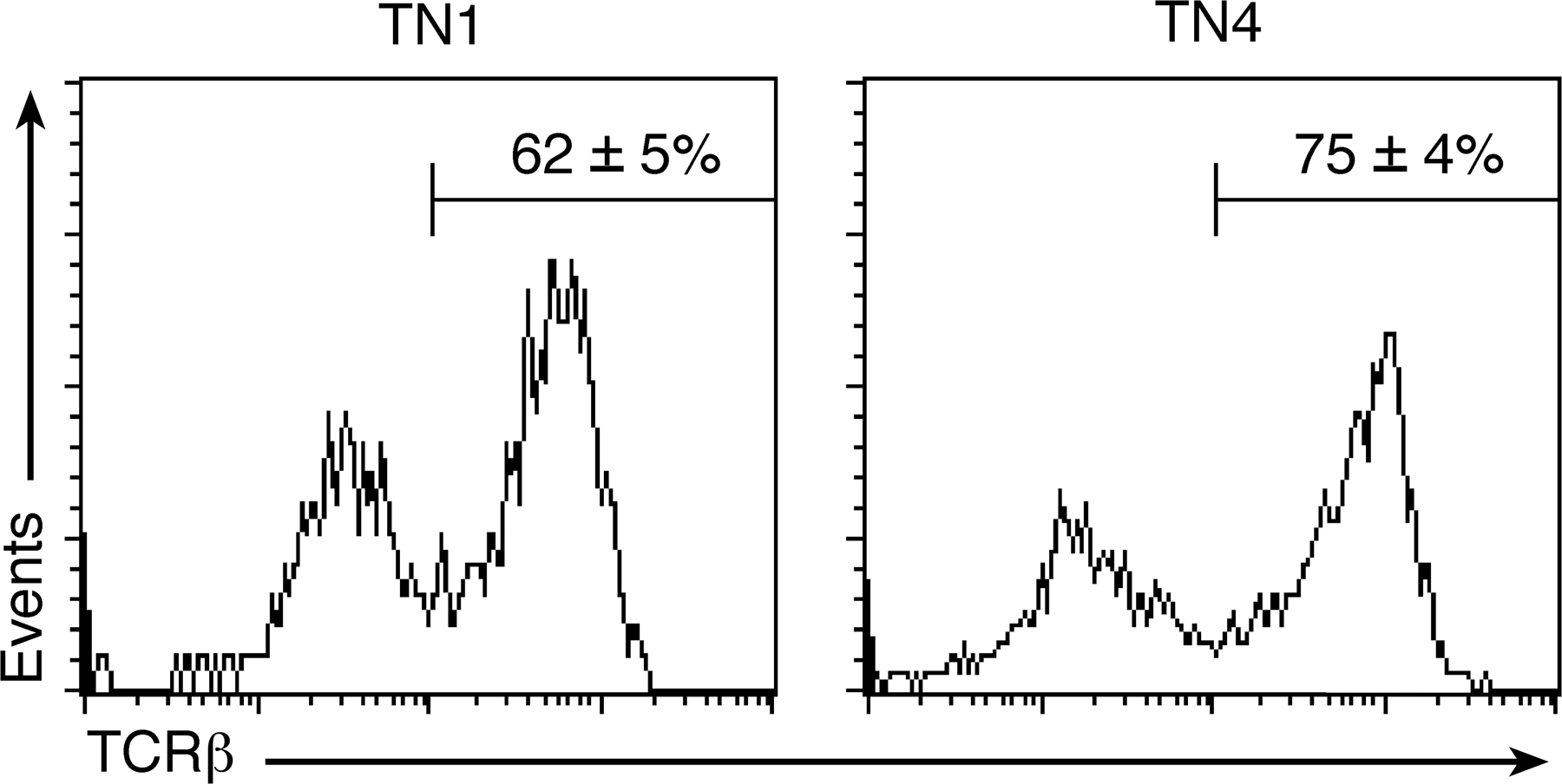
To determine whether cells entering the thymus after irradiation and BMT may be directly affected by the presence or absence of FL, donor-derived intrathymic progenitors were assessed for Flt3 receptor expression at 7 and 10 d after BMT. It was found that 17 ± 2% of ETPs and 14 ± 1% of triple-negative (TN) 1 cells expressed the Flt3 receptor and, thus, could respond to FL expressed by thymic stromal cells (Fig. 3 A).
Figure 3.

Enhanced thymic reconstitution after lethal irradiation is dependent on thymic stromal FL expression. (A) Flt3 receptor expression on TN cell populations 7 and 10 d after lethal irradiation and BMT. Flt3 expression (black line) and isotype (gray line) are shown. Numbers in plots indicate the Flt3+ cell percentage ± SEM. (B–E) Reconstitution of thymus with donor-derived cells after irradiation and BMT at the time points indicated. (B) Total donor-derived cells. (C) Cell numbers of donor-derived TN subpopulation. (D) ETP cell numbers. (E) iTCR expression on TN4 cells at 7 and 10 d after BMT. iTCR expression (bolded line) and isotype (continuous line) are shown. Numbers in the plots indicate the iTCR-positive cell percentage ± SEM. n = 5 in all experiments. Error bars represent SEM. Statistics are based on an unpaired t test analysis. *, P < 0.05; **, P < 0.005; ***, P < 0.0005.
At 7 and 10 d after irradiation and BMT, the number of donor-derived thymocytes was significantly higher in Flt3−/− mice and significantly lower in FL−/− mice (Fig. 3 B). At 14 d, the total number of donor-derived cells in the thymus was similar for all experimental groups, indicating that the effect of FL is evident very early after transplantation.
To determine whether the donor-derived cells detected in the thymus are committing to the T cell lineage, we analyzed TN populations based on a lack of lineage markers and a sequential change in CD44 and CD25 expression (21). At 7 d after irradiation and BMT, all donor-derived cells in the thymus were either of the TN1 (CD45.1+ Lin− CD44+CD25−) or TN4 (CD45.1+ Lin− CD44−CD25−) phenotype, regardless of the genetic background of the recipients (Fig. 3 C). This distribution pattern may be caused by a rapid progression of transplanted cells to the TN4 stage or entry of cells that have already attained a TN2/TN3 phenotype (CD45.1+ Lin− CD44+CD25+ and CD45.1+ Lin− CD44−CD25+, respectively) outside the thymus (22). The number of TN4 cells on day 7 was significantly lower in FL−/− mice and significantly higher in Flt3−/− mice. To confirm that cells gated as TN4 are indeed committed T cell progenitors, we examined intracellular TCRβ (iTCR) expression. We found that 56 ± 4% of the TN4 cells at day 7 and 28 ± 5% at day 10 after BMT in the B6 mouse express iTCR (Fig. 3 E). Additionally, TN1 and TN4 donor-derived cells were FCM purified from mice 7 d after BMT and cultured on an OP9–delta-like 1 (OP9-DL1). Both populations underwent growth and differentiation, and up-regulated TCRβ (Fig. S5, available at http://www.jem.org/cgi/content/full/jem.20072065/DC1).
10 d after conditioning and BMT, a small number of double-positive cells (CD4+CD8+) were detected in all groups (Fig. S6, available at http://www.jem.org/cgi/content/full/jem.20072065/DC1), presumably developing from TN4 cells observed at day 7. At day 10, Flt3−/− mice had significantly more donor TN cells of all subpopulations when compared with either FL−/− or B6 hosts, whereas FL−/− mice had significantly fewer (Fig. 3 C). At day 14, FL−/− mice continued to show a decrease in TN1 and TN2 numbers compared with the other two mouse groups. It should be noted that FL has been proposed to act in an autocrine manner (11), as wild-type progenitor cells can be induced to express FL; however, these donor cells are unable to compensate for the lack of stromal-expressed FL in the FL-deficient environment.
Donor-derived ETPs (CD45.1+ Lin−c-kit+CD25− cells) could not be identified in any experimental mouse group at day 7 after irradiation and BMT. The donor-derived TN1 cells seen at day 7 do not correspond to ETPs but may represent non–T lineage progenitors. At later time points, at 10 and 14 d, significantly more ETPs could be detected in Flt3−/− and B6 recipients when compared with FL−/− hosts (Fig. 3 D).
To visualize the intrathymic location of transplanted cells, mice of all three strains were lethally irradiated and transplanted with T cell–depleted syngeneic GFP+ BM cells. IHC analysis at day 10 after treatment demonstrated that in all groups, donor-derived cells were found in MTS15+ regions (Fig. 4). In accordance with FCM analysis (Fig. 3), more GFP+ cells were seen in irradiated B6 and Flt3−/− recipients than in FL−/− hosts. As newly immigrated thymic progenitors reside in the corticomedullary junction for up to 10 d (23), this association infers that immigrating progenitors are maintained in physical and functional contact with thymic stromal cells expressing FL.
Figure 4.

Analysis of GFP+ cells after irradiation and BMT. IHC of B6, FL−/−, and Flt3−/− thymi 10 d after lethal irradiation and BMT of T cell–depleted syngeneic GFP+ BM cells. Donor-derived GFP+ cells are shown in green. MTS15 staining is in red. (bottom) Enlarged views of the areas indicated by boxes in the top panels. Bars ,120 μM.
The present report demonstrates the functional relevance of the expression of the Flt3 receptor on thymic progenitors colonizing the thymus after irradiation, as these cells may respond to FL expression by thymic stromal cells. Moreover, these findings also provide an explanation for the10-fold reduction observed in the number of TN1 and TN2 cells in unirradiated Flt3−/− and FL−/− mice (11, 13), indicating that FL may also play a role in thymopoiesis under physiological conditions.
Intrathymic FL enhances thymocyte cell number
To test whether FL stimulates the proliferation and/or differentiation of intrathymic T cell precursors independent of immigration into the thymus, B6 CD45.1+ BM LSK cells were directly injected into a single lobe of sublethally irradiated B6 and FL−/− mice. 8 d later, there were at least 2-fold more donor-origin cells in the B6 thymus and a 5.7-fold increase in cells that had committed to the T cell lineage (i.e., TN3 cells; Fig. 5, A and B). When injections were performed using thymic-derived ETPs, a similar 2.8-fold increase in donor-derived cell number and a 2.5-fold increase in T lineage–committed cells was observed in the B6 as compared with the FL−/− mice (Fig. 5, C and D). These results indicate that intrathymic expression of FL enhances the proliferation of T cell precursors and/or their commitment to the T cell lineage.
Figure 5.

Intrathymic FL expression increases donor-derived cell numbers. CD45.1+ LSK cells (A and B) or ETPs (C and D) were injected intrathymically into B6 (CD45.2+) and FL−/− (CD45.2+) thymi after sublethal irradiation. (A and C) Total donor-derived cell numbers 8 d after intrathymic injection (CD45.1+ gated). (B and D) Numbers of donor-derived cells that had progressed to the TN3 stage of development (CD45.1+ gated). n = 8 in all experiments. (E–I) B6 and FL−/− E15 lobes were grafted under opposite kidney capsules in FL −/− hosts and allowed to engraft for 3 wk. (E) Gross anatomy of kidneys engrafted with B6 or FL−/− lobes (four lobes per kidney) 10 d after lethal irradiation and BMT with GFP+ BM cells. (F) FCM analysis of FL expression on MTS15+ fibroblasts in B6 lobes 4 d after lethal irradiation. FL expression (bolded line) and isotype (continuous line) are shown. Number in the plot indicates MFIR. (G–I) Donor-derived (GFP+) cell numbers 10 d after lethal irradiation and BMT in grafted thymic lobes. (G) Total donor-derived cells. (H and I) Donor-derived thymic subpopulation cell numbers. n = 6 in all experiments. Error bars represent SEM. *, P < 0.05; **, P < 0.005; ***, P < 0.0005.
The importance of an FL-mediated expansion of T cell progenitors has been suggested previously by fetal thymic organ studies (24) and by the inability of Flt3−/− progenitors to compete with wild-type cells in the generation of mature T cells in a wild-type mouse, even when transplanted at a fourfold excess (25). Intrathymic injection experiments do not, however, preclude a role for FL as a chemoattractant for the immigration of cells into the thymus or for attracting ETPs to their proposed niche once these cells have gained access to the thymus.
To definitively show an intrathymic requirement for FL in thymopoiesis after BMT, we performed thymic grafting experiments in which embryonic day 15 (E15) B6 and FL−/− thymic lobes were grafted under opposite kidney capsules of the same FL−/− host. In this setting, only the grafted B6 thymic stroma can express FL. Lobes were allowed to mature, and mice were lethally irradiated before BMT. B6 lobes were visually larger than FL−/− lobes (Fig. 5 E) and up-regulated FL on the surface of MTS15+ fibroblasts after irradiation (Fig. 5 F). Furthermore, 10 d after BMT, B6 lobes had a threefold increase in donor-derived cell numbers compared with FL−/− grafted lobes (Fig. 5 G). All thymocyte populations from double-negative to single-positive cells were significantly affected by the thymic stromal expression of FL (Fig. 5 H). Notably, there was a 10-fold increase in ETPs in B6 when compared with FL−/− lobes (Fig. 5 I), demonstrating that expression of FL by the thymic stroma has a significant effect on thymopoiesis in the absence of any effect FL may have on the BM.
Given the number of progenitor cells entering the thymus under steady-state conditions and after irradiation is very low (26), as well as the massive expansion these cells then undergo as ETPs, it can be postulated that the interaction of thymic progenitors expressing the Flt3 receptor with stromal FL drives an increase in thymic cellularity that can translate to a marked increase in thymic output.
This report used irradiated recipient mice, as this parallels the clinical situation of conditioning before HSCT performed routinely to treat hematological malignancies. Although irradiation does not appear to change the kinetics of early T cell development (23), previous studies have demonstrated that the thymic microenvironment is altered after this treatment. Although ETPs are dependant on stem cell factor (10) and IL-7 (8) production by the thymic microenvironment for their survival and proliferation, IL-7 production is decreased in TECs after irradiation (unpublished data) (18). In contrast, FL expression is increased even after sublethal doses of irradiation, suggesting that Flt3+ progenitors may undergo preferential expansion in this setting.
Chemokine receptors such as CCR9 and CXCR4 and adhesion molecules such as CD44 are known to play a role in early thymopoiesis (4). In this report, we provide evidence that expression of the Flt3 receptor by thymic progenitors is also functionally important and not merely a legacy from stem cells, as is the case for many genes expressed by T cell progenitors (27). Flt3 expression allows cells to respond to stromal cell–expressed FL, which may drive the extensive expansion of thymic settling cells and leads to a significant increase in early T cell numbers.
The use of FL in immunotherapy will not be straightforward, as the administration of soluble FL does not appear sufficient to enhance thymic function after irradiation. However, we show that the localized membrane-bound expression of FL in the thymus can lead to an enhanced thymic recovery. Delayed T cell recovery after irradiation represents one of the major causes of HSCT-related mortality. Although cytokine treatments are used clinically to increase levels of other cell types, such as granulocytes, after myeloablative stress (28), no cytokine treatment has been demonstrated to increase naive T cell number without deleterious side effects. The current report identifies for the first time functionally relevant FL expression in the thymus. This provides a better understanding of thymic recovery after myeloablative stress, which could in turn translate to the design of strategies to improve thymic recovery after HSCT.
MATERIALS AND METHODS
Mice.
C57BL/6, CD45.2+ (B6), C57BL/6, CD45.1+ (CD45.1), C57BL/6-transgenic (ACTB-EGFP), Flt3 knockout CD45.2+ (Flt3−/−; provided by I. Lemischka, Princeton University, Princeton, NJ), and FL knockout CD45.2+ (FL−/−; provided by J.J. Peschon, Amgen, Seattle, WA) mice were bred and maintained under pathogen-free conditions in the animal facility of the Department of Biomedicine at the University Hospital Basel. Mice used in experiments were age and sex matched and used at ages 4–12 wk. All experiments were performed in accordance with institutional and federal regulations, and were approved by the veterinary office of the canton of Basel.
IHC.
Thymi tissue sections were prepared as previously described (29) and blocked with PBS/10% mouse serum. Primary mAbs used were anti-FL–biotin (R&D Systems), anti–ER-TR7 (provided by W. van Ewijk, Leiden University, Leiden, Netherlands), anti-CD31 (clone MTS12), anti-MTS15 (29), and anti–cytokeratin 5 (Covance). Secondary antibodies and reagents used for detection were all obtained from Invitrogen. The threshold of detection for FL was determined by analysis of anti-FL immunoreactivity on FL−/− tissue. Images were captured on a laser scanning confocal microscope system (LSM 510 Meta; Carl Zeiss, Inc.).
Cell preparations and multicolor FCM analysis.
Thymocyte single-cell suspensions were prepared by mechanically disrupting the thymus using frosted glass slides. PB was obtained from the vena cava, and BM was recovered by flushing both femurs and tibias with IMDM (Invitrogen)/2% FCS.
The following mAbs were directly coupled to fluorescein isothiocyanate, phycoerythrin, phycoerythrin-Cy7, allophycocyanin, or biotin, and were commercially obtained from BD Biosciences or eBioscience: B220, CD3, CD4, CD8, CD11b, CD11c, CD19, Gr-1, NK1.1, TCRβ, and TCRγδ (designated as lineage markers for use in TN thymocyte detection); and Sca-1, CD25, CD31 (MTS12), CD44, CD45.1, CD45.2, CD117, CD135, and EpCAM. Surface marker expression was visualized with a FACSCalibur and analyzed with CellQuest software (Becton Dickinson). LSK and ETP cells were sorted to >98% purity on a FACSAria (Becton Dickinson).
Thymic stromal cells were prepared as described previously (29), with minor alterations. In brief, thymi were dissected and digested with 0.01% (wt/vol) DNaseI (Sigma-Aldrich) and 0.15% (wt/vol) collagenase D (Cedar Lane). Dissociated cells were subsequently incubated with 10 mM PBS/EDTA, resuspended in Percoll (1.115 g/cm3), and layered with Percoll (1.065 g/cm3) and PBS on top. Gradients were centrifuged at 1,220 g at 4°C for 30 min. Low density cells were subsequently stained using mAbs to CD45, CD31, EpCAM, FL, and MTS15. FCM gating of stromal cells is shown in Fig. S1 (available at http://www.jem.org/cgi/content/full/jem.20072065/DC1).
Irradiation and congenic BMT.
Mice were irradiated sublethally with a single dose of 6 Gy or lethally with a split dose of 10 Gy, using a 137Cs source (0.91 Gy/min). Mice were subsequently intravenously grafted with 2.5 × 106 T cell–depleted, CD45.1+, or GFP+ BM cells. T cells were depleted by incubation of cells with anti-CD4 and -CD8 antibodies (our preparation) for 1 h at 4°C, followed by incubation with complement (Cedar Lane) for 1 h at 37°C.
Intrathymic injections.
B6 and FL−/− mice were sublethally irradiated (6 Gy) and intrathymically injected with 2,000 CD45.1+ LSK cells or 2,000 CD45.1+ ETP cells into a single thymic lobe, as previously described (26). For each experimental mouse, the uninjected, contralateral thymic lobe was used as control.
RT-PCR.
RNA was isolated with TRIzol (Invitrogen), and quantitative real-time RT-PCR was performed with TaqMan Universal PCR Master Mix (Applied Biosystems) and analyzed on an ABI Prism 7000 (Applied Biosystems). Primer sequences were as follows: RPL19 forward, 5′-ATCCGCAAGCCTGTGACTGT-3′; RPL19 reverse, 5′-TCGGGCCAGGGTGTTTTT-3′; FL forward, 5′-CACCTCATGTACCTTCCAGCC-3′; and FL reverse, 5′-GTCTGGACGATTCGCAGACA-3′.
OP9-DL1 seeding.
TN1 and TN4 cells were purified to a purity of >98% on a FACSAria and seeded onto an OP9-DL1 monolayer. Cells were cultured for 3 wk and analyzed by FCM.
Grafting.
Adult FL−/− mice were transplanted with B6 and FL−/− E15 thymic lobes under opposite kidney capsules, as previously described (30). Grafts were allowed to mature for 3 wk. Mice were lethally irradiated (10 Gy) and intravenously grafted with 5 × 106 T cell–depleted GFP+ BM cells. Transplants were analyzed by FCM.
Statistical analysis.
Significance was analyzed by the unpaired, one-tailed Student's t test using Prism4 software (GraphPad). We considered p-values <0.05 significant (*, P < 0.05; **, P < 0.005; ***, P < 0.0005). Error bars represent SEM values.
Online supplemental material.
FCM gating used for stromal cell analysis is shown in Fig. S1. Fig. S2 demonstrates that Flt3−/−MTS15+ fibroblasts do not increase expression of FL after irradiation. Fig. S3 shows that mRNA expression of FL by thymic stromal cells does not increase after irradiation. Fig. S4 shows reconstitution of the BM and PB by donor cells 10 d after irradiation and BMT. Fig. S5 shows TCRβ up-regulation by TN1 and TN4 cells after culture on an OP9-DL1 monolayer. Fig. S6 shows numbers of donor-derived double-positive (CD4+CD8+) cells after irradiation and BMT. Online supplemental material is available at http://www.jem.org/cgi/content/full/jem.20072065/DC1.
Supplemental Material
Acknowledgments
The authors thank A. Rolink, R. Ceredig, E. Palmer, and D. Gray for critical reading of the manuscript; H. Kohler for cell sorting; I. Lemischka (Princeton University, Princeton, NJ) for Flt3−/− mice; and J.J. Peschon (Amgen, Seattle, WA) for FL−/− mice.
This work was supported by the Swiss National Science Foundation (grant 3100-110511 to A. Wodnar-Filipowicz; grant 3100-68310.02 to G.A. Holländer), the National Institutes of Health (grant ROI-A1057477-01 to G.A. Holländer), the Roche Research Foundation (grant 2005-118 to L. Kenins), and the National Health and Medical Research Council of Australia (CJ Martin fellowship 237036 to J.W. Gill).
The authors have no conflicting financial interests.
L. Kenins and J.W. Gill contributed equally to this work.
G.A. Holländer and A. Wodnar-Filipowicz contributed equally to this work.
References
- 1.Copelan, E.A. 2006. Hematopoietic stem-cell transplantation. N. Engl. J. Med. 354:1813–1826. [DOI] [PubMed] [Google Scholar]
- 2.Gill, J., M. Malin, J. Sutherland, D. Gray, G. Hollander, and R. Boyd. 2003. Thymic generation and regeneration. Immunol. Rev. 195:28–50. [DOI] [PubMed] [Google Scholar]
- 3.Takahama, Y. 2006. Journey through the thymus: stromal guides for T-cell development and selection. Nat. Rev. Immunol. 6:127–135. [DOI] [PubMed] [Google Scholar]
- 4.Bhandoola, A., H. von Boehmer, H.T. Petrie, and J.C. Zuniga-Pflucker. 2007. Commitment and developmental potential of extrathymic and intrathymic T cell precursors: plenty to choose from. Immunity. 26:678–689. [DOI] [PubMed] [Google Scholar]
- 5.Petrie, H.T., and J.C. Zuniga-Pflucker. 2007. Zoned out: functional mapping of stromal signaling microenvironments in the thymus. Annu. Rev. Immunol. 25:649–679. [DOI] [PubMed] [Google Scholar]
- 6.Radtke, F., A. Wilson, G. Stark, M. Bauer, J. van Meerwijk, H.R. MacDonald, and M. Aguet. 1999. Deficient T cell fate specification in mice with an induced inactivation of Notch1. Immunity. 10:547–558. [DOI] [PubMed] [Google Scholar]
- 7.Rothenberg, E.V. 2007. Negotiation of the T lineage fate decision by transcription-factor interplay and microenvironmental signals. Immunity. 26:690–702. [DOI] [PubMed] [Google Scholar]
- 8.Banwell, C.M., K.M. Partington, E.J. Jenkinson, and G. Anderson. 2000. Studies on the role of IL-7 presentation by mesenchymal fibroblasts during early thymocyte development. Eur. J. Immunol. 30:2125–2129. [DOI] [PubMed] [Google Scholar]
- 9.Bolotin, E., M. Smogorzewska, S. Smith, M. Widmer, and K. Weinberg. 1996. Enhancement of thymopoiesis after bone marrow transplant by in vivo interleukin-7. Blood. 88:1887–1894. [PubMed] [Google Scholar]
- 10.Massa, S., G. Balciunaite, R. Ceredig, and A.G. Rolink. 2006. Critical role for c-kit (CD117) in T cell lineage commitment and early thymocyte development in vitro. Eur. J. Immunol. 36:526–532. [DOI] [PubMed] [Google Scholar]
- 11.Sitnicka, E., D. Bryder, K. Theilgaard-Monch, N. Buza-Vidas, J. Adolfsson, and S.E. Jacobsen. 2002. Key role of flt3 ligand in regulation of the common lymphoid progenitor but not in maintenance of the hematopoietic stem cell pool. Immunity. 17:463–472. [DOI] [PubMed] [Google Scholar]
- 12.Adolfsson, J., O.J. Borge, D. Bryder, K. Theilgaard-Monch, I. Astrand-Grundstrom, E. Sitnicka, Y. Sasaki, and S.E. Jacobsen. 2001. Upregulation of Flt3 expression within the bone marrow Lin(−)Sca1(+)c-kit(+) stem cell compartment is accompanied by loss of self-renewal capacity. Immunity. 15:659–669. [DOI] [PubMed] [Google Scholar]
- 13.Sambandam, A., I. Maillard, V.P. Zediak, L. Xu, R.M. Gerstein, J.C. Aster, W.S. Pear, and A. Bhandoola. 2005. Notch signaling controls the generation and differentiation of early T lineage progenitors. Nat. Immunol. 6:663–670. [DOI] [PubMed] [Google Scholar]
- 14.Wodnar-Filipowicz, A. 2003. Flt3 ligand: role in control of hematopoietic and immune functions of the bone marrow. News Physiol. Sci. 18:247–251. [DOI] [PubMed] [Google Scholar]
- 15.Maraskovsky, E., E. Daro, E. Roux, M. Teepe, C.R. Maliszewski, J. Hoek, D. Caron, M.E. Lebsack, and H.J. McKenna. 2000. In vivo generation of human dendritic cell subsets by Flt3 ligand. Blood. 96:878–884. [PubMed] [Google Scholar]
- 16.Fry, T.J., M. Sinha, M. Milliron, Y.W. Chu, V. Kapoor, R.E. Gress, E. Thomas, and C.L. Mackall. 2004. Flt3 ligand enhances thymic-dependent and thymic-independent immune reconstitution. Blood. 104:2794–2800. [DOI] [PubMed] [Google Scholar]
- 17.Wils, E.J., E. Braakman, G.M. Verjans, E.J. Rombouts, A.E. Broers, H.G. Niesters, G. Wagemaker, F.J. Staal, B. Lowenberg, H. Spits, and J.J. Cornelissen. 2007. Flt3 ligand expands lymphoid progenitors prior to recovery of thymopoiesis and accelerates T cell reconstitution after bone marrow transplantation. J. Immunol. 178:3551–3557. [DOI] [PubMed] [Google Scholar]
- 18.Chung, B., L. Barbara-Burnham, L. Barsky, and K. Weinberg. 2001. Radiosensitivity of thymic interleukin-7 production and thymopoiesis after bone marrow transplantation. Blood. 98:1601–1606. [DOI] [PubMed] [Google Scholar]
- 19.Buza-Vidas, N., M. Cheng, S. Duarte, H. Nozad, S.E. Jacobsen, and E. Sitnicka. 2007. Crucial role of FLT3 ligand in immune reconstitution after bone marrow transplantation and high-dose chemotherapy. Blood. 110:424–432. [DOI] [PubMed] [Google Scholar]
- 20.Miyazawa, K., D.A. Williams, A. Gotoh, J. Nishimaki, H.E. Broxmeyer, and K. Toyama. 1995. Membrane-bound Steel factor induces more persistent tyrosine kinase activation and longer life span of c-kit gene-encoded protein than its soluble form. Blood. 85:641–649. [PubMed] [Google Scholar]
- 21.Godfrey, D.I., J. Kennedy, T. Suda, and A. Zlotnik. 1993. A developmental pathway involving four phenotypically and functionally distinct subsets of CD3−CD4−CD8− triple-negative adult mouse thymocytes defined by CD44 and CD25 expression. J. Immunol. 150:4244–4252. [PubMed] [Google Scholar]
- 22.Maillard, I., B.A. Schwarz, A. Sambandam, T. Fang, O. Shestova, L. Xu, A. Bhandoola, and W.S. Pear. 2006. Notch-dependent T-lineage commitment occurs at extrathymic sites following bone marrow transplantation. Blood. 107:3511–3519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Lind, E.F., S.E. Prockop, H.E. Porritt, and H.T. Petrie. 2001. Mapping precursor movement through the postnatal thymus reveals specific microenvironments supporting defined stages of early lymphoid development. J. Exp. Med. 194:127–134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Moore, T.A., and A. Zlotnik. 1997. Differential effects of Flk-2/Flt-3 ligand and stem cell factor on murine thymic progenitor cells. J. Immunol. 158:4187–4192. [PubMed] [Google Scholar]
- 25.Mackarehtschian, K., J.D. Hardin, K.A. Moore, S. Boast, S.P. Goff, and I.R. Lemischka. 1995. Targeted disruption of the flk2/flt3 gene leads to deficiencies in primitive hematopoietic progenitors. Immunity. 3:147–161. [DOI] [PubMed] [Google Scholar]
- 26.Schwarz, B.A., A. Sambandam, I. Maillard, B.C. Harman, P.E. Love, and A. Bhandoola. 2007. Selective thymus settling regulated by cytokine and chemokine receptors. J. Immunol. 178:2008–2017. [DOI] [PubMed] [Google Scholar]
- 27.Tydell, C.C., E.S. David-Fung, J.E. Moore, L. Rowen, T. Taghon, and E.V. Rothenberg. 2007. Molecular dissection of prethymic progenitor entry into the T lymphocyte developmental pathway. J. Immunol. 179:421–438. [DOI] [PubMed] [Google Scholar]
- 28.Bradstock, K.F. 2002. The use of hematopoietic growth factors in the treatment of acute leukemia. Curr. Pharm. Des. 8:343–355. [DOI] [PubMed] [Google Scholar]
- 29.Gray, D.H., D. Tull, T. Ueno, N. Seach, B.J. Classon, A. Chidgey, M.J. McConville, and R.L. Boyd. 2007. A unique thymic fibroblast population revealed by the monoclonal antibody MTS-15. J. Immunol. 178:4956–4965. [DOI] [PubMed] [Google Scholar]
- 30.Gill, J., M. Malin, G.A. Hollander, and R. Boyd. 2002. Generation of a complete thymic microenvironment by MTS24(+) thymic epithelial cells. Nat. Immunol. 3:635–642. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}