

Abstract
The contractile properties and ATPase activity of skinned human skeletal muscle fibres from vastus lateralis were examined. Fibre types were resolved from single fibre segments by SDS-polyacrylamide gel electrophoresis. ATPase activity was determined by enzymatic coupling of ATP resynthesis to the oxidation of NADH.
The partitioning of ATPase activity into (a) calcium-activated activity due to actomyosin (AM) interaction, (b) calcium-activated activity of the sarcoplasmic reticular (SR) calcium pump, and (c) basal (calcium independent) activity was investigated by comparing ATP utilization before and after exposure of the preparations for 30 min to a solution containing 0.5 % Triton X-100, which effectively abolished the SR ATPase activity.
Partitioning of ATPase activity was also determined by measuring ATP utilization and force at different concentrations of butanedione monoxime (BDM), which inhibits AM interaction.
The results obtained with Triton X-100 and BDM were similar. At saturating Ca2+ concentrations and 20 °C, the AM, SR and basal ATPase activities per litre cell volume (±s.e.m.) amounted to 46 ± 4, 51 ± 4 and 19 ± 2 μm s−1 in type I fibres (n = 21), 139 ± 14, 69 ± 8 and 30 ± 3 μm s−1 in type IIA fibres (n = 25), 137 ± 22, 175 ± 28 and 26 ± 8 μm s−1 in type IIA/B fibres (n = 4) and 108 ± 13, 169 ± 42 and 32 ± 8 μm s−1 in type IIB fibres (n = 2).
These results indicate that ATP utilization for SR Ca2+ pumping in fast fibres is considerably larger than in slow fibres. The SR ATPase activity in human muscle represents a considerable fraction of the total (AM + SR + basal) ATPase activity.
The major energy-consuming processes in active skeletal muscle are the release and uptake of calcium ions by the sarcoplasmic reticulum (SR) and the actomyosin (AM) interaction, which results in force development. The chemical energy for these processes is provided by the hydrolysis of adenosine 5′-triphosphate (ATP). It is well established that marked differences exist between fibre types with regard to the rates of energy expenditure associated with the SR and AM ATPase. The variation in SR ATPase is most likely to be due to differences of density in SR Ca2+ pumps between different fibre types (Zubrzycka-Gaarn et al. 1982). The difference in the isometric AM ATPase activity between fibre types originates from differences between myosin isoforms (see for a review, Reggiani et al. 2000).
Little is known about the fibre type dependence of energy expenditure in human muscle fibres. Improvements in 31P NMR spectroscopy have made it possible to estimate ATP consumption during twitch stimulation and during maximal voluntary contraction in skeletal muscle in vivo (Boska, 1991; Blei et al. 1993a,b; Turner et al. 1993). However, the large heterogeneity in fibre properties and the variability in fibre type distribution make it very difficult to determine the differences in ATP consumption at the cellular level. Using permeabilized muscle fibres obtained from biopsies from healthy volunteers, Stienen et al. (1996) showed that considerable differences exist in myofibrillar ATPase activity (and isometric tension cost) between fibres of different type. However, so far nothing is known about the partitioning of the SR and AM ATPase activity in human skeletal muscle fibres.
The present study aimed to determine the maximum rates of ATP consumption for the SR Ca2+ pump and for the actomyosin interaction in single human skeletal muscle fibres during isometric contraction and their fibre type dependence. For this purpose single fibres were used which were made permeable by means of saponin treatment. Saponin permeabilizes the surface membrane of the fibres but does not affect the SR Ca2+ pump (Launikonis & Stephenson, 1997). Since the non-ionic detergent Triton X-100 effectively destroys the SR, we compared the ATPase activity before and after Triton X-100 treatment in order to quantify the AM and SR ATPase activity in the same preparations.
Another method which can be employed in skinned preparations for this purpose is based on the action of 2,3-butanedione monoxime (BDM) on the actomyosin interaction. BDM inhibits force production as well as AM ATPase activity (Higuchi & Takemori, 1989). It has previously been used to study the SR and AM ATPase (Stienen et al. 1995) and the force-dependent and force-independent heat production (Buschman et al. 1996) in muscle fibres from Xenopus. Several studies, however, demonstrated a dose-dependent effect of BDM on the shape of the Ca2+ transients, both in cardiac (e.g. Gwathmey et al. 1992) and in skeletal muscle (Fryer et al. 1988; Horiuti et al. 1988) which might interfere with these determinations. BDM has the advantage over Triton X-100 in that its action is reversible but because it might interfere with Ca2+ handling, we decided to employ both methods.
The results presented here give information on the partitioning of the major energy-requiring processes during muscle contraction, which is essential for understanding the differences in efficiency of energy transduction in human muscle tissue. A preliminary abstract of part of this work has previously been published (Szentesi et al. 1999).
METHODS
The methods used in this study are a combination of those described previously (Bottinelli et al. 1994; Stienen et al. 1995, 1996). The methods used are briefly described below and details are given for the experimental protocols followed.
Muscle biopsies and permeabilization of fibres
The experiments conformed with the Declaration of Helsinki. The local ethics committees approved the study and the subjects gave informed consent. Muscle biopsies were taken from two healthy male volunteers (24 and 46 years) who had no previous history of muscular or neuromuscular disease. These biopsies were taken under local anaesthesia with Xylocaine, from the distal portion of vastus lateralis muscle using the needle biopsy technique (Bergstrom, 1962). The needle (outer diameter 5 mm) was inserted at an oblique angle to maximize the length of the fibre segments. The biopsies (wet weight about 50 mg) were divided into bundles of about 50 fibres each and placed in storage solution to which glycerol was added (50 % v/v). The bundles were stored at -20 °C for up to 1 month.
On the day of an experiment one bundle of fibres was placed in a dissection chamber containing storage solution at a temperature of 5-6 °C. A number of segments of single fibres of about 5 mm in length were isolated. Thereafter, the fibres were permeabilized by saponin by incubating them in storage solution containing 50 μg ml−1 saponin for 30 min. At the end of the incubation period, the solution was replaced by a relaxing solution without saponin. In a few cases (n = 5), fresh fibres were mechanically skinned, by peeling off the surface membrane with sharp forceps, immediately after obtaining the biopsy. This was done to assess the possible effects of saponin treatment and storage in glycerol on SR ATPase activity.
A few experiments were performed on saponin-skinned soleus fibres from rat. One Wistar rat was anaesthetised by sodium pentobarbitone (0.3 ml, i.p.) and killed by exsanguination. The soleus muscle was excised quickly and superficial fibres were isolated and treated as was the case for the human fibres.
Solutions
The storage solution contained (mm): Na2ATP 3, magnesium acetate 5, EGTA 5, potassium dihydrogen phosphate 5, potassium propionate 150, dithiothreitol (DTT) 1. Three different bathing solutions were used during the experiments: a relaxing solution, a pre-activating solution with low EGTA concentration and an activating solution. The composition of the basic solutions is given in Table 1. The pCa (-log10[Ca2+]) of the activating solutions was 4.4. In addition, all solutions contained 5 mm caffeine to prevent the inhibition of the SR ATPase activity by Ca2+ inside the SR. Activating solutions containing BDM (5, 10, 20 and 40 mm) were obtained by appropriate mixing of activating solutions without and with 40 mm BDM (solid) added. The pH of the solutions was adjusted to pH 7.1 with potassium hydroxide.
Table 1.
Basic composition of the solutions
| Relaxing | Pre-activating | Activating | |
|---|---|---|---|
| MgCl2 | 8.31 | 7.78 | 7.63 |
| Na2ATP | 5.79 | 5.79 | 5.86 |
| PEP | 10 | 10 | 10 |
| EGTA | 20 | 0.5 | — |
| HDTA | — | 19.5 | — |
| Ca2+-EGTA | — | — | 20 |
| K-Prop | 34.7 | 35.8 | 35.7 |
Concentrations are given in millimolar. PEP: phosphoenol pyruvate (Roche); HDTA: hexamethylene diamine-tetraacetate (Fluka). All solutions contained in addition 4 mg ml−1 pyruvate kinase (500 U ml−1),0.24 mg ml−1 lactic dehydrogenase (870 U ml−1), 5 mM sodium azide, 10 μM oligomycin B, 0.8 mM NADH and 0.2 mM p1, p5-di(adenosin-5′)pentaphosphate (Roche) and 5 mM caffeine. Potassium propionate (K-Prop) was added to adjust ionic strength to 200 mM. Ca2+-EGTA was made by dissolving equimolar amounts of CaCO3 and EGTA. The free Mg2+ and MgATP concentrations were, respectively, 1 and 5 mM. The solutions contained 100 mM N,N-bis[2-hydroxyethyl]-2-aminoethanesulfonic acid (Bes). The pH was adjusted to 7.1 with potassium hydroxide. Various additions were performed depending on the type of experiment as described in Methods. All chemicals were obtained from Sigma unless otherwise stated.
Experimental set-up
The apparatus used to measure the ATPase activity was as described previously (Glyn & Sleep, 1985; Stienen et al. 1990). It consisted of several temperature-controlled troughs in which the fibre could be immersed. Temperature was maintained at 20 ± 0.5 °C. The fibre segments were mounted, by means of aluminium T-clips (Goldman & Simmons, 1984), between a force transducer (AE801, Sensonor, Horten, Norway) and a fixed hook. Sarcomere length was measured in relaxing solution by means of He-Ne laser diffraction and was adjusted to 2.5 μm. During the actual measurement of ATPase activity, the preparation was kept in a small trough with glass windows having a volume of 30 μl. Hydrolysis of ATP inside the fibre was linked to the oxidation of NADH. The decline in NADH concentration was measured photometrically via the absorption at 340 nm of near-UV light from a 75 W xenon light source that passed beneath the fibre. The rate of ATP hydrolysis was derived from the slope of the absorbance signal relative to the baseline found before and after immersion of the fibre in the measuring chamber. Calibration of the absorbance signal was carried out after each recording by injecting a known amount of ADP into the measuring chamber via a stepper motor-controlled pipette.
Experimental protocol
During measurements, the fibre segment was incubated in relaxing solution for 3 min, in pre-activating solution for about 3 min, and in activating solution until a steady force level was attained, and was then transferred back into the relaxing solution. After the first contraction, sarcomere length was readjusted to 2.5 μm, if necessary, and the horizontal and vertical diameter and length of the fibre segment were measured by means of a dissection microscope. Cross-sectional area and fibre volume were calculated assuming an elliptical cross-section. The results were corrected for deterioration in force and ATPase activity by linear interpolation between control values. After the first control recording, one or several activations were recorded using activating solutions which contained different BDM concentrations. These measurements were followed by a maximally activating control measurement without BDM. In several cases, the preparation was subsequently incubated for 30 min in dissecting solution containing 0.5 % Triton X-100 and the control and BDM recordings were repeated. To determine the basal ATPase activity, the ATPase measurement was performed while the fibre was immersed in relaxing solution (pCa 9). In some cases cyclopiazonic acid (CPA, 10 μm) as a specific inhibitor of the SR calcium pump was applied. The experiments were terminated when isometric force during the control measurements was < 75 % of the force of the first activation. ATPase activities were normalized to fibre volume and force was normalized to cross-sectional area of the fibres.
Myosin isoform identification
Myosin composition was determined by means of polyacrylamide gel electrophoresis (PAGE) after denaturation in sodium dodecyl sulphate (SDS), using a method similar to that described by Danieli-Betto et al. (1986). After each successful series of force measurements, the fibre segment (typically 2 mm long) was placed in a small test-tube filled with 10 μl of solution with the following composition: 62.5 mm Tris-HCl, 2.3 % (w/v) SDS, 10 % glycerol and 5 % (v/v) mercaptoethanol (Laemmli, 1970), and after solubilization for at least 2 h at room temperature, stored at -70 °C. A small fraction of the solution, typically 2 μl, was analysed with SDS-PAGE. Gel dimensions were: width, 18 cm; height, 16 cm (4 cm stacking gel, 12 cm running gel); and thickness, 0.75 mm. The acrylamide/bisacrylamide ratio was 37.5:1. Total acrylamide was 4 and 6 % in the stacking and running gel, respectively. The stacking gel buffer contained 0.4 % (w/v) SDS and 0.5 m Tris-HCl (pH 6.8), and the separating gel buffer contained 0.4 % SDS, 1.5 m Tris-HCl (pH 8.8) and 35 % (v/v) glycerol. The electrode buffer contained 25 mm Tris, 192 mm glycine and 0.1 % SDS (pH 8.3). Samples were run for 22-24 h at constant voltage (100 V) at 20 °C. The gels were stained with silver.
Data and statistical analysis
Force and ATPase activity were recorded with a pen recorder and after analog-to-digital conversion by a personal computer at a sampling rate of 10 Hz. Data values are given as means ±s.e.m. of n experiments. Student’s t test and one-way analysis of variance (ANOVA) were used to assess statistical significance of the differences between fibre types. A paired t test was used to assess statistical significance of the differences in data before and after Triton treatment. A probability of less than 0.05 was considered statistically significant. The effect of BDM on isometric force and ATPase activity was approximated by means of the following relation:
| (1) |
where, YBDM represents the isometric force or ATPase activity in the presence of BDM, Y0 is the value in control conditions (without BDM) and Y∞ is the residual level at infinite BDM concentration normalized to Y0. EC50 is the BDM concentration where the inhibition is 50 %. The values derived from the curve fits are given ± standard error of the estimate.
RESULTS
ATPase activity and force development in different fibre types
Force development and ATPase activity during isometric contraction were studied in 52 fibre segments obtained from human vastus lateralis muscle. All fibres were characterized on the basis of MHC composition (see Fig. 1) and divided into four groups: type I (or slow), 21 fibres; type IIA, 25 fibres; type IIA/B (showing coexistence of fast IIA and IIB MHC isoforms), 4 fibres; and type IIB, 2 fibres. In the literature the human MHC isoform IIB is also called IIX. The dimensions of the fibre segments were measured at 2.5 μm sarcomere length. The mean dimensions obtained were independent of fibre type (P < 0.05). The mean values for all preparations were: segment length, 2.5 ± 0.1 mm; width, 104 ± 3 μm; and depth, 113 ± 3 μm.
Figure 1. Electrophoretic analysis of the myosin heavy chain composition of human fibre segments from vastus lateralis muscle.
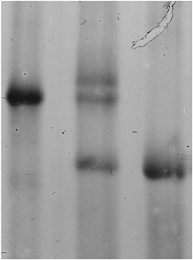
The figure shows examples from a type IIA fibre (lane 1), a type I fibre (lane 3) and from a laboratory standard which contained a mixture of fibres with almost equal amounts of type I, IIA and IIB myosin heavy chains (lane 2).
The overall means of the total ATPase activity (±s.e.m.) per litre cell volume and the mean maximal force per cross-sectional area (tension) measured at the beginning of the experiments in type IIA/B, IIB, I and IIA fibres are given in the last four columns of Table 2. The differences in maximal tension between fibre types were not significant (P < 0.05). The differences in total ATPase activity between fibre types were highly significant. The results from the type IIB fibres were not included in the statistical comparison because this group contained only two fibres. The resting or basal ATPase activity was measured in relaxing solution (pCa 9). The overall mean values obtained in each fibre type are given in Table 2. Basal ATPase activity corresponded to 16 ± 2, 13 ± 1, 8 ± 2 and 12 ± 4 % of the total ATPase activity in type I, IIA, IIA/B and IIB fibres, respectively. This relative basal activity in type I fibres was significantly higher (P < 0.05) than in type IIA and IIA/B fibres.
Table 2.
Overview of the fibre type dependence of the total, AM, SR, basal ATPase isometric tension and tension cost
| Triton X-100 BDM | BDM + Triton | |||||||
|---|---|---|---|---|---|---|---|---|
| I | IIA | I | IIA | IIA/IIB | IIB | I | IIA | |
| Number of fibres | 5 | 6 | 20 | 23 | 4 | 2 | 21 | 25 |
| Total ATPase (μM s−1) | 119 ± 5¶ | 306 ± 44¶* | 112 ± 5 | 227 ± 18 | 332 ± 43 | 309 ± 46 | 111 ± 5†‡ | 224 ± 16§ |
| AM ATPase (μM s−1) | 58 ± 6 | 207 ± 35* | 43 ± 4 | 132 ± 14 | 137 ± 22 | 108 ± 13 | 46 ± 4†‡ | 139 ± 14 |
| SR ATPase (μM s−1) | 41 ± 5 | 75 ± 13 | 54 ± 4 | 66 ± 8 | 175 ± 28 | 169 ± 42 | 51 ± 4‡ | 69 ± 8§ |
| Basal ATPase (μM s−1) | 20 ± 2¶ | 29 ± 7¶ | 19 ± 2 | 31 ± 4 | 26 ± 8 | 32 ± 8 | 19 ± 2 | 30 ± 3 |
| Isometric tension (kN m−2) | 93 ± 9¶ | 73 ± 9¶ | 90 ± 6 | 75 ± 5 | 74 ± 11 | 54 ± 8 | 87 ± 7 | 76 ± 4 |
| Tension cost (μM s−1 m2 kN−1) | 0.70 ± 0.12 | 2.88 ± 0.4* | 0.52 ± 0.05 | 1.84 ± 0.20 | 1.69 ± 0.04 | 1.88 ± 0.18 | 0.56 ± 0.05†‡ | 1.93 ± 0.19 |
Values are means ± S.E.M. derived from the individual measurements in each fibre segment. Values for BDM IIA/IIB and IIB and for BDM + Triton I and IIA (the last 4 columns) are means for each fibre type. Note that in some fibres both Triton and BDM were used. In this case the mean values from both determinations were used to calculate the mean values per fibre type. Tension cost was defined as the isometric AM ATPase activity divided by force. Type IIA/B and IIB fibres were present only in the BDM series.
Measured before Triton X-100 treatment.
Significant difference between Triton and BDM series
significant difference between type I and type IIA in overall mean
significant difference between type I and type IIA/B in overall mean
significant difference between type IIA and type IIA/B in overall mean.
To assess the effects of saponin treatment on the ATPase and force values, a comparison was made between the results of five mechanically skinned fibres (4 of type IIA, 1 of type IIB) with those of the saponin-skinned fibres. Total ATPase was 278 ± 13 and 240 μm s−1, basal ATPase was 47 ± 5 and 43 μm s−1 and isometric tension was 68 ± 7 and 43 kN m−2 in type IIA and IIB fibres, respectively. The differences between mechanically skinned and saponin-treated IIA fibres were not statistically significant. Therefore in the overall results and in the statistical evaluations (summarized in Table 2), the data from mechanically and saponin-skinned preparations were pooled.
Discrimination between AM and SR ATPase activity by Triton X-100 treatment
The method used to determine the AM and SR ATPase activity is illustrated in Fig. 2. In this figure ATPase activity and force development are shown for a saponin-treated IIA type fibre before and after Triton X-100 treatment. Before Triton X-100 treatment, the ATPase activity measured reflects the total (AM + SR + basal) ATPase activity while after Triton X-100 treatment the (AM + basal) ATPase activity remains. Therefore, provided that the basal ATPase activity before and after Triton treatment is the same, subtraction of the two values yields the SR ATPase activity.
Figure 2. Force and ATPase activity in a type IIA fibre.
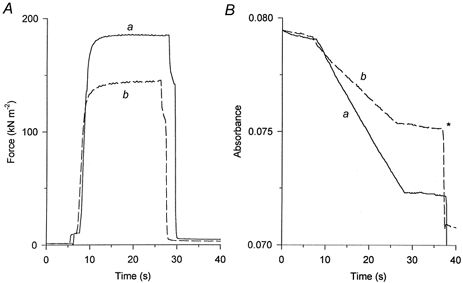
A, force development; B, NADH absorbance. Continuous traces (a) were obtained before while dashed traces (b) were recorded after abolishing the SR ATPase activity of the fibre with Triton X-100. The fibre was activated by transferring it from the preactivating solution (pCa 9) into the activating solution (pCa 4.4), and relaxed, at the end of the measurement, by transferring it back into relaxing solution (pCa 9). At the asterisk a calibration of the absorbance signal was carried out which corresponded to 0.5 nmol of ATP hydrolysed. The zero level in the absorbance signal was arbitrarily chosen. Fibre diameters measured in two perpendicular directions: 90 and 130 μm; fibre segment length: 2.3 mm.
The ATPase activity and maximal isometric force were determined before and after Triton exposure in 11 fibres. In this experimental group only type I and type IIA fibres were present. As can be seen in Fig. 2, the ATPase activity was reduced substantially (to 59.7 %) after Triton X-100 treatment while force development was reduced less (to 79.5 %). The total ATPase activity during this series of experiments in saponin-skinned type I (n = 5) and type IIA fibres (n = 6) expressed per litre cell volume amounted to 119 ± 5 and 306 ± 44 μm s−1, respectively. On average the maximum calcium-activated ATPase activity decreased to 58.1 ± 5.3 and 72.9 ± 3.2 % of the maximum measured before the Triton X-100 treatment, while force decreased to 83.3 ± 2.3 and 78.2 ± 2.7 %(n = 5 and 6) in type I and type IIA fibres, respectively.
In nine fibre segments, the basal ATPase activity was determined by incubating the fibre in relaxing solution (pCa 9) at 20 °C. Before Triton X-100 exposure the mean basal ATPase activity found in saponin-skinned type I (n = 4) and type IIA (n = 5) fibres was 20 ± 2 and 29 ± 7 μm s−1, respectively. These values were not significantly different from the basal ATPase activity (16 ± 2 and 24 ± 6 μm s−1 in type I and type IIA fibres, respectively) found after Triton X-100 treatment. Hence it can be concluded that basal ATPase activity before and after Triton was the same. This implies, as noted above, that the SR ATPase activity can be obtained from the difference in ATPase activity before and after Triton treatment. Using this value, the partitioning between the AM, SR and basal ATPase activities can be calculated (Table 2; columns 2 and 3).
BDM dependence of the force and ATPase activity
The effect of BDM on force and ATPase activity was investigated in 49 fibres both before and after Triton X-100 treatment up to a BDM concentration of 40 mm. The mean values of the total ATPase activity and the basal ATPase activity expressed per litre cell volume as well as the mean maximal tension before the Triton treatment in type I, IIA, IIA/B and IIB fibres are given in Table 2. The effect of BDM on isometric force and ATPase activity in a type IIA fibre is illustrated in Fig. 3. It can be seen that BDM (40 mm, in this example) caused a considerable reduction in force (by 87.2 %) and ATPase activity (by 56.3 %) compared to the control measurement.
Figure 3. Effect of BDM on force and ATPase activity in a saponin-treated type IIA fibre.
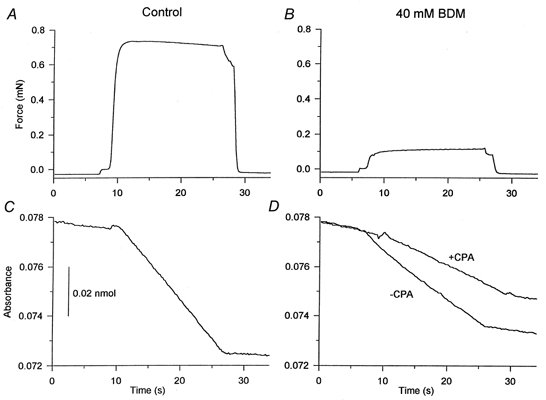
A and B, force; C and D, NADH absorbance. In A and C, the results are shown under control conditions. In B and D, the results are shown in the presence of 40 mm BDM. The absorbance traces marked + or -CPA were carried out in the presence and absence of 10 μm CPA, respectively. CPA causes a marked reduction in the ATPase activity, whereas the force recordings in the presence and absence of CPA were identical. Fibre diameters measured in two perpendicular directions: 100 and 160 μm; fibre segment length: 3.0 mm.
This figure also shows the effect of 10 μm CPA, a specific inhibitor of the SR ATPase activity. The ATPase activity in the presence of 40 mm BDM was markedly depressed by CPA (22.5 % of the total ATPase activity in control), indicating that a large fraction of the total ATPase activity, which remained when force production had been reduced, was due to SR ATPase activity. The force was identical before and after CPA treatment in the presence of BDM (11.7 % of the control). It can be noted that CPA was very effective because the fraction of ATPase activity remaining in CPA corresponds well with the fraction of the force remaining in 40 mm BDM when the basal ATPase activity is taken into account.
As Fig. 4A shows, active force was reduced on average to about 15 % in the presence of 40 mm BDM. The effect of BDM on isometric force before and after Triton X-100 treatment was the same and independent of fibre type. In contrast, the depressing effect of BDM on ATPase activity (Fig. 4B) after Triton was larger than before. This indicates that Triton X-100 removes a BDM-insensitive component of the total ATPase activity, namely the ATPase associated with Ca2+ pumping (the SR ATPase). Note that in this figure the calcium-activated ATPase activity is plotted, i.e. the total – basal ATPase activity, to facilitate the determination of the SR ATPase activity.
Figure 4. BDM dependence of isometric force (A) and ATPase activity (B).
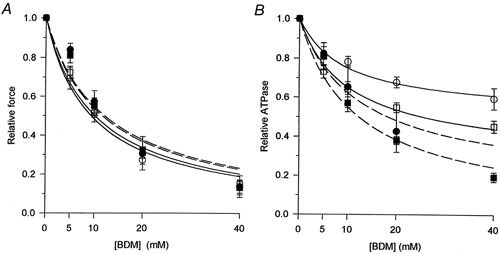
Open symbols, before treatment by Triton X-100; filled symbols, after Triton. Circles, type I fibres; squares, type IIA fibres. Error bars, s.e.m., shown only when larger than the symbol size. Continuous and dashed lines, least square fit of eqn (1) to the means before and after the Triton treatment, respectively. The effect of BDM on isometric force was half-maximal at a concentration of 9.4 ± 1.0, 10.1 ± 1.0, 12.1 ± 2.5 and 11.7 ± 2.0 mm in type I and IIA fibres before and after the Triton treatment, respectively.
Force and the ATPase activity could be fitted by eqn (1) (Methods). In order to limit the number of variables in the fit procedure, we assumed in the case of force that the value at infinite BDM concentration (Y∞) was equal to zero while in the case of ATPase activity, the EC50 was considered equal to the value obtained from the fit on force. It was found that the effect of BDM on the isometric force before and after Triton exposure was the same in different fibre types. The EC50 values were 9.4 ± 1.0 and 10.1 ± 1.0 mm in saponin-treated type I and IIA fibres, respectively. The EC50 values after Triton treatment were very similar (12.1 ± 2.5 and 11.7 ± 2.0 mm, in type I and IIA fibres, respectively). The values for Y∞, the residual ATPase activity at infinite [BDM] or zero force, were 51.8 ± 1.7 % in type I and 29.9 ± 1.8 % in type IIA fibres. These values reflect the relative contribution of the SR ATPase activity to the total ATPase activity.
The effects of BDM are also illustrated in Fig. 5. Here the relative ATPase activity before and after Triton treatment, shown in Fig. 4, is replotted as a function of the relative force. A linear relationship between force and ATPase activity was found before as well as after Triton X-100 treatment. The linearity of both force-ATPase relationships suggests that the SR ATPase activity is not influenced by BDM in the concentration range up to 40 mm.
Figure 5. The relationship between isometric force and ATPase activity at different BDM concentrations before and after Triton X-100 treatment.
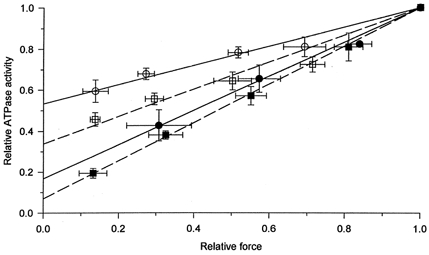
The results of experiments at the same BDM concentrations were normalized to the values in the absence of BDM and than averaged. Open symbols, before Triton X-100 treatment; filled symbols, after Triton treatment. Circles, type I fibres; squares, type IIA fibres. In the figure BDM concentrations in increasing order (0, 5, 10, 20 and 40 mm) correspond to ATPase activities in decreasing order. Error bars, s.e.m. values. The continuous and dashed lines were obtained by fitting a straight line (y = a(x– 1) + 1) to the individual data points before and after the Triton X-100 exposure, respectively.
To determine the SR ATPase activity, we fitted a linear function to the data points. The y-axis intercept of the fitted line (the ATPase activity at zero force) in saponin-skinned type I and IIA fibres was 53.2 ± 2.0 and 33.9 ± 3.9 %, respectively. After Triton treatment, it was only 16.7 ± 3.0 and 6.9 ± 0.9 % of the calcium-activated ATPase activity measured in the absence of BDM in type I and IIA fibres, respectively. The intercepts after Triton treatment were not significantly different from zero. In 23 fibres only one BDM concentration (20 mm) was used. The results obtained in these fibres were not significantly different from the overall mean values.
Fibre type dependency of the AM, SR, basal ATPase activity and tension cost
The mean results of all type I and IIA fibres are given in the last two columns in Table 2. This table also shows the mean values of tension cost, defined as the ratio between isometric AM ATPase activity and tension, for each fibre type, calculated from the individual values obtained in each fibre.
The data were obtained from two biopsies from different subjects. Significant differences (≈33 %) were found in the total, AM and SR ATPase activity in type IIA fibres between the two biopsies, but there were no significant differences in total, AM and SR ATPase activity in type I fibres and in total isometric force in both fibre types.
Figure 6 shows the partitioning of the ATPase activities obtained for the different fibre types, i.e. the mean values from both methods for type I and type IIA fibres, as well as those for type IIA/B and IIB fibres obtained by BDM treatment. It can be seen that the basal ATPase is a relatively small fraction of the total ATPase activity and that the SR and AM ATPase co-vary to some extent.
Figure 6. Fibre type dependency of the basal, SR and AM ATPase activity.
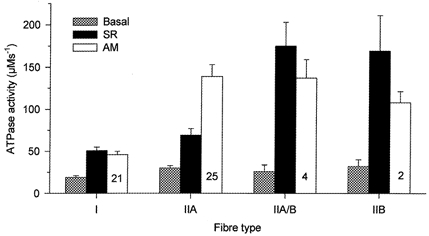
Mean data from all experiments (Table 2, last 4 columns). The numbers of fibres in each group are shown inside the bars.
Rate of SR Ca2+-ATPase and ATP turnover per myosin head in different fibre types
Assuming that the density of myosin heads which participate during contraction is 200 μm (see Glyn & Sleep, 1985), the AM ATPase activity corresponds to a rate of ATP turnover per myosin head of 0.23, 0.70, 0.69 and 0.54 s−1 in type I, IIA, IIA/B and IIB fibres, respectively. In intact human skeletal muscle fibres the SR Ca2+ pump content is about 50 μm (Sturk et al. 1998). In skinned fibres, the volume is increased osmotically by about a factor of 1.44 (Larsson & Moss, 1993), which reduces the concentration of Ca2+ pumps to about 35 μm. Assuming a stoichiometry of the Ca2+ pump in the SR of 2 Ca2+ per ATP, the SR ATPase rate corresponds to a rate of Ca2+ uptake per SR Ca2+ pump (at 20 °C) of about 7 s−1 (range 3-10 s−1) in human fibres.
DISCUSSION
This study gives a description of the maximal SR ATPase activity in different types of human skeletal muscle fibres. In type I and type IIA fibres, the predominant fibre types in human vastus lateralis, two different methods were employed to determine the partitioning of the AM, SR and basal ATPase activity (Triton and BDM treatment). The two methods yielded similar results but it has to be noted that some uncertainty is present in the determinations using Triton X-100. The isometric force development after Triton treatment was smaller than before Triton treatment. If Triton would also reduce the AM ATPase activity, our estimates of the SR ATPase activity in these experiments would be somewhat too large. However, Triton was used in only 20 % of the experiments. Thus this uncertainty has little impact on the mean values presented. Caution should be exerted when extrapolating our values to body temperature because the temperature dependence of the SR ATPase in human fibres is not known. It should be noted, however, that in the range between 20 and 35 °C, the temperature coefficient of mammalian SR ATPase (Inesi et al. 1973) is very similar to the temperature coefficient of human AM ATPase found between 20 and 30 °C (Stienen et al. 1996). This would imply that their ratio would be fairly temperature independent.
The SR ATPase activity was about 50 % of the calcium-activated ATPase activity in type I, IIA/B and IIB fibres and about 30 % in type IIA fibres. Since force development and the AM and SR ATPase activity was studied in the same preparations under identical conditions, these results give information about the relationship between activation and contractile processes at the cellular level. Furthermore, the values obtained are relevant under physiological circumstances because the composition of the bathing solutions used for the skinned fibres was similar to the composition of the cytosol.
Comparison with previous results
The rate of ATP turnover and tension in skinned human fibres after Triton X-100 found in this study are ≈25 % smaller than the values at 20 °C of Stienen et al. (1996). However, the values obtained for tension cost did not differ significantly, except those in type I fibres. Since ATP turnover and tension are both expressed per cross-sectional area, we consider it likely that the origin of this difference resides in the fact that in the current study the diameter of the fibres was measured before Triton treatment, while in the previous study the diameter was measured after Triton treatment. Triton dissolves the SR and removes the mitochondria (Miller et al. 1985) and hence may cause a decrease in cross-sectional area of the fibres. It can be noted that in saponin-skinned soleus muscle fibres from the rat we found that the mean cross-sectional area of the fibres (±s.e.m.) after Triton treatment was reduced by 23 ± 3 %(n = 3) of the area before Triton treatment. However, because of the difference in tension cost in type I fibres between the two studies, differences among subjects might be involved as well.
The range in SR ATPase activity expressed as a fraction of the total ATPase activity found in this study (0.3-0.6) is very similar to that found in biochemical and heat measurements in other species. For instance, Infante et al. (1964), Smith (1972), Homsher et al. (1972), Burchfield & Rall (1986), Stienen et al. (1996) and Rome & Klimov (2000) found values in the range from 0.25 to 0.4.
The activity of the SR Ca2+ pumps at 20 °C is about 7 s−1. Taking the temperature dependence of the SR Ca2+ pump into account (e.g. Stienen et al. 1996), this rate agrees well with the initial rate of Ca2+ uptake found in mammalian SR vesicles, about 10 s−1 at 24-25 °C (Inesi & Scarpa, 1972), and with the rate found by Feher et al. (1998) in muscle homogenates from rat EDL (30 s−1 at 37 °C). This implies that the activity of the SR Ca2+ pump obtained in our study is similar to that found in in vitro studies.
The reduction in force after Triton treatment (≈20 %) is larger than the reduction found in our laboratory in Xenopus fibres (≈5 %). This indicates that human fibres appear to be more sensitive to Triton treatment than Xenopus fibres.
BDM dependence of the AM and SR ATPase activity
Our results show that isometric force and AM ATPase activity vary in proportion at different BDM concentrations. The BDM dependencies were such that both in type I and type IIA fibres depression was half-maximal at 10 mm BDM. In a previous study (Stienen et al. 1995) it was found that BDM had similar effects on isometric force and AM ATPase activity in skinned muscle fibres of Xenopus laevis. Fryer et al. (1988) found reductions in force development to 46.6 ± 2.8 % in fast and 60.1 ± 2.0 % in slow fibres at 10 mm BDM, which were significantly different. It can be concluded that the BDM sensitivity in human fibres is very similar to the sensitivity in rat fibres but that it does not depend significantly on fibre type. From caged-ATP experiments Kagawa et al. (1995) concluded that BDM inhibits the Ca2+-regulated attachment of the cross-bridges and force generation of the attached cross-bridges. Our experiments indicate that the tension cost (i.e. the ratio of AM ATPase activity and isometric tension) is independent of the BDM concentration. In a two-state cross-bridge model, tension cost is proportional to the rate of detached cross-bridges (e.g. Brenner, 1988). Therefore our results are in agreement with a predominant effect of BDM on the apparent rate of cross-bridge attachment.
Several effects of BDM on calcium handling by the sarcoplasmic reticulum are reported in the literature. A direct reduction of calcium release by BDM has previously been reported (Fryer et al. 1988; Horiuti et al. 1988). Horiuti et al. (1988) also reported an inhibiting effect of BDM on calcium uptake into the SR. De Armas et al. (1998) showed that BDM modified the delayed (Qγ) component of intramembraneous charge movement in frog muscle and in this way indirectly reduced Ca2+ release from the SR. The linear relationship between total ATPase activity and force found in this study indicates that the maximum activity of the SR calcium pump is not affected by up to 40 mm BDM.
Fibre type dependence of the AM and SR ATPase activity
The maximal actomyosin (AM) ATPase activity per litre cell volume (±s.e.m.) at 20 °C in type I fibres amounted to 46 ± 4 μm s−1. The AM ATPase activity in type IIA fibres amounted to 139 ± 14 μm s−1, i.e. about a factor of 3 larger than in slow twitch fibres. The SR ATPase activity in type I fibres was 51 ± 4 μm s−1 while in type IIA fibres amounted to 69 ± 8 μm s−1. This indicates that it is somewhat larger in IIA fibres than in type I fibres. In the mixed type IIA/B fibres and in type IIB fibres the SR ATPase activities were 175 ± 28 and 169 ± 42 μm s−1, respectively, i.e. about a factor of three larger than in type I fibres. This difference in SR ATPase activity is intermediate between the sixfold difference in Ca2+-ATPase content found between fast and slow fibres from rabbit (Leberer & Pette, 1986) and the 1.8-fold difference found in chicken (Gauthier & Hobbs, 1986). The human fibres appear to differ from rabbit in that the SR ATPase activity in type IIA and IIA/B fibres differs significantly.
Our results indicate that type IIA can be considered as an intermediate fibre type. Its SR ATPase activity is near that of type I, but its AM ATPase activity resembles that of IIA/B and IIB fibres. A previous study from our laboratory (Stienen et al. 1995) revealed differences in AM ATPase activity between slow- and fast-twitch fibres from Xenopus, while the SR ATPase activities in these two fibre types were fairly similar. It seems plausible to attribute the differences (or the lack thereof) between fibres of different type to expression levels of SR Ca2+ pumps (Zubrzycka-Gaarn et al. 1982). The range of the SR ATPase activity expressed as a fraction of the total ATPase activity ranged from 0.3 in slow type I fibres to 0.6 in fast type IIB fibres. Although we consider this difference meaningful it can be noted that the overall co-variation found between the energy requirements for the activation and contraction processes in animal studies (Homsher & Kean, 1978) also applies to human muscle fibres.
The activity of the SR Ca2+ pump in human fibres at body temperature
The values for the SR ATPase activity obtained range between 50 μm s−1 in type I fibres and about 175 μm s−1 in type IIA/B fibres. These values can be used to estimate the maximum rate of ATP consumption during isometric contractions at muscle temperature (35 °C) in vivo. In skinned fibres, the volume is increased osmotically by about a factor of 1.44. Our values were obtained at 20 °C. The temperature-dependent increase in SR ATPase activity for an increase in temperature by 10 °C amounts to a factor of ≈3 (e.g. Stienen et al. 1995). Taking the difference in volume and in temperature into account it can be estimated that the maximum SR ATPase in vivo should increase by a factor of 4.3 and range between 215 and 750 μm s−1.
Acknowledgments
We thank Dr László Csernoch for valuable discussions. This study was supported, in part, by the Netherlands Organisation for Scientific Research (NWO) and Hungarian Research Grant (OTKA 030246). P.S. holds a Bolyai fellowship.
References
- Bergstrom J. Muscle electrolytes in man. Scandinavian Journal of Clinical Laboratory Investigation. 1962;14(suppl.):68. [Google Scholar]
- Blei ML, Conley KE, Kushmerick MJ. Separate measurements of ATP utilization and recovery in human skeletal muscle. Journal of Physiology. 1993a;465:203–222. doi: 10.1113/jphysiol.1993.sp019673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blei ML, Conley KE, Odderson IR, Esselman PC, Kushmerick MJ. Individual variation in contractile cost and recovery in human skeletal muscle. Proceedings of the National Academy of Sciences of the USA. 1993b;90:7396–7400. doi: 10.1073/pnas.90.15.7396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boska M. Estimating the ATP cost of force production in the human gastrocnemius-soleus muscle group using 31P MRS and 1H MRI. NMR in Biomedicine. 1991;4:173–181. doi: 10.1002/nbm.1940040404. [DOI] [PubMed] [Google Scholar]
- Bottinelli R, Canepari M, Reggiani C, Stienen GJM. Myofibrillar ATPase activity during isometric contraction and isomyosin composition in rat single skinned muscle fibres. Journal of Physiology. 1994;481:663–675. doi: 10.1113/jphysiol.1994.sp020472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brenner B. Effect of Ca2+ on cross-bridge turnover kinetics in skinned single rabbit psoas fibres: implications for regulation of muscle contraction. Proceedings of the National Academy of Sciences of the USA. 1988;85:3265–3269. doi: 10.1073/pnas.85.9.3265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burchfield DM, Rall JA. Temperature dependence of the crossbridge cycle during unloaded shortening and maximum isometric tetanus in frog skeletal muscle. Journal of Muscle Research and Cell Motility. 1986;7:320–326. doi: 10.1007/BF01753652. [DOI] [PubMed] [Google Scholar]
- Buschman HPJ, Elzinga G, Woledge R. The effect of the level of activation and shortening velocity on energy output in type 3 muscle fibres from Xenopus laevis. Pflügers Archiv. 1996;433:153–159. doi: 10.1007/s004240050261. [DOI] [PubMed] [Google Scholar]
- Danieli-Betto D, Zerbato E, Betto R. Type I, IIa and IIb myosin heavy chain electrophoretic analysis of rat muscle fibres. Biochemical and Biophysical Research Communications. 1986;138:981–987. doi: 10.1016/s0006-291x(86)80592-7. [DOI] [PubMed] [Google Scholar]
- De Armas R, González S, Brum G, Pizarro G. Effects of 2,3-butanedione monoxime on excitation-contraction coupling in frog twitch fibres. Journal of Muscle Research and Cell Motility. 1998;19:961–977. doi: 10.1023/a:1005409121660. [DOI] [PubMed] [Google Scholar]
- Feher JJ, Waybright TD, Fine ML. Comparison of sarcoplasmic reticulum capabilities in toadfish (Opsanus tau) sonic muscle and rat fast twitch muscle. Journal of Muscle Research and Cell Motility. 1998;19:661–674. doi: 10.1023/a:1005333215172. [DOI] [PubMed] [Google Scholar]
- Fryer MW, Neering IR, Stephenson DG. Effects of 2,3-butanedione monoxime on the contractile activation properties of fast- and slow-twitch rat muscle fibres. Journal of Physiology. 1988;407:53–75. doi: 10.1113/jphysiol.1988.sp017403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gauthier GF, Hobbs AW. Freeze-fractured sarcoplasmic reticulum in adult and embryonic fast and slow muscles. Journal of Muscle Research and Cell Motility. 1986;7:122–132. doi: 10.1007/BF01753413. [DOI] [PubMed] [Google Scholar]
- Glyn H, Sleep J. Dependence of adenosine triphosphatase activity of rabbit psoas muscle fibres and myofibrils on substrate concentration. Journal of Physiology. 1985;365:259–276. doi: 10.1113/jphysiol.1985.sp015770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldman YE, Simmons RM. Control of sarcomere length in skinned muscle fibres of Rana temporaria during mechanical transients. Journal of Physiology. 1984;350:497–518. doi: 10.1113/jphysiol.1984.sp015215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gwathmey JK, Hajjar RJ, Solaro RJ. Contractile deactivation and uncoupling of crossbridges. Circulation Research. 1992;69:1280–1292. doi: 10.1161/01.res.69.5.1280. [DOI] [PubMed] [Google Scholar]
- Higuchi H, Takemori S. Butanedione monoxime suppresses contraction and ATPase activity of rabbit skeletal muscle. Journal of Biochemistry. 1989;105:638–643. doi: 10.1093/oxfordjournals.jbchem.a122717. [DOI] [PubMed] [Google Scholar]
- Homsher E, Kean CJ. Skeletal muscle energetics and metabolism. Annual Review of Physiology. 1978;40:93–131. doi: 10.1146/annurev.ph.40.030178.000521. [DOI] [PubMed] [Google Scholar]
- Homsher E, Mommaerts WFHM, Ricchiuti NV, Wallner A. Activation heat, activation metabolism and tension-related heat in frog semitendinosus muscles. Journal of Physiology. 1972;220:601–625. doi: 10.1113/jphysiol.1972.sp009725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horiuti K, Higuchi H, Umazume Y, Konishi M, Okazaki O, Kurihara S. Mechanism of action of 2,3-butanedione 2-monoxime on contraction of frog skeletal muscle fibres. Journal of Muscle Research and Cell Motility. 1988;9:156–164. doi: 10.1007/BF01773737. [DOI] [PubMed] [Google Scholar]
- Inesi G, Millman M, Eletr S. Temperature-induced transitions of function and structure in sarcoplasmic reticulum membranes. Journal of Molecular Biology. 1973;81:483–504. doi: 10.1016/0022-2836(73)90518-4. [DOI] [PubMed] [Google Scholar]
- Inesi G, Scarpa A. Fast kinetics of adenosine triphosphate dependent Ca2+ uptake by fragmented sarcoplasmic reticulum. Biochemistry. 1972;11:356–359. doi: 10.1021/bi00753a008. [DOI] [PubMed] [Google Scholar]
- Infante AA, Klaupiks D, Davies RE. Length, tension and metabolism during short isometric contractions of frog sartorius muscles. Biochimica et Biophysica Acta. 1964;88:215–217. doi: 10.1016/0926-6577(64)90171-8. [DOI] [PubMed] [Google Scholar]
- Kagawa K, Horiuti K, Yamada K. BDM compared with Pi and low Ca2+ in the cross-bridge reaction initiated by flash photolysis of caged ATP. Biophysical Journal. 1995;69:2590–2600. doi: 10.1016/S0006-3495(95)80130-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227:680–681. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- Larsson L, Moss RL. Maximum velocity of shortening in relation to myosin isoform composition in single fibres from human skeletal muscles. Journal of Physiology. 1993;472:595–614. doi: 10.1113/jphysiol.1993.sp019964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Launikonis BS, Stephenson DG. Effects of saponin treatment on the sarcoplasmic reticulum of rat, cane toad and crustacean (yabby) skeletal muscle. Journal of Physiology. 1997;504:425–437. doi: 10.1111/j.1469-7793.1997.425be.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leberer E, Pette D. Immunochemical quantification of sarcoplasmic reticulum Ca-ATPase, of calsequestrin and of parvalbumin in rabbit skeletal muscles of defined fibre composition. European Journal of Biochemistry. 1986;156:489–496. doi: 10.1111/j.1432-1033.1986.tb09607.x. [DOI] [PubMed] [Google Scholar]
- Miller DJ, Elder HY, Smith GL. Ultrastructural and X-ray microanalytical studies of EGTA- and detergent-treated heart muscle. Journal of Muscle Research and Cell Motility. 1985;6:525–540. doi: 10.1007/BF00711913. [DOI] [PubMed] [Google Scholar]
- Reggiani C, Bottinelli R, Stienen GJM. Sarcomeric myosin isoforms: fine tuning of a molecular motor. News in Physiological Sciences. 2000;15:26–33. doi: 10.1152/physiologyonline.2000.15.1.26. [DOI] [PubMed] [Google Scholar]
- Rome LC, Klimov AA. Superfast contractions without superfast energetics: ATP usage by the SR-Ca2+ pumps and crossbridges in toadfish swimbladder muscle. Journal of Physiology. 2000;526:279–298. doi: 10.1111/j.1469-7793.2000.t01-1-00279.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith ICH. Energetics of activation in frog and toad muscle. Journal of Physiology. 1972;220:583–599. doi: 10.1113/jphysiol.1972.sp009724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stienen GJM, Kiers JL, Bottinelli R, Reggiani C. Myofibrillar ATPase activity in skinned human skeletal muscle fibres: fibre type and temperature dependence. Journal of Physiology. 1996;493:299–307. doi: 10.1113/jphysiol.1996.sp021384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stienen GJM, Roosemalen MCM, Wilson MGA, Elzinga G. Depression of force by phosphate in skinned skeletal muscle fibres of the frog. American Journal of Physiology. 1990;259:C349–357. doi: 10.1152/ajpcell.1990.259.2.C349. [DOI] [PubMed] [Google Scholar]
- Stienen GJM, Zaremba R, Elzinga G. ATP utilization for calcium uptake and force production in skinned muscle fibres of Xenopus laevis. Journal of Physiology. 1995;482:109–122. doi: 10.1113/jphysiol.1995.sp020503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sturk A, Lehmann-Horn F, Melzer W. Voltage-dependent calcium release in human malignant hyperthermia muscle fibres. Biophysical Journal. 1998;75:2402–2410. doi: 10.1016/S0006-3495(98)77684-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szentesi P, Zaremba R, Stienen GJM. ATP utilization for calcium uptake and force production in skinned human skeletal muscle fibres. Journal of Muscle Research and Cell Motility. 1999;20:93. [Google Scholar]
- Turner DL, Jones DA, McIntyre DB, Newham DJ. ATP turnover measured by 31P-magnetic resonance spectroscopy in human adductor pollicis muscle during isometric and shortening contractions. Journal of Physiology. 1993;459:154P. [Google Scholar]
- Zubrzycka-Gaarn E, Korczak B, Osinska H, Sarzala MG. Studies on sarcoplasmic reticulum from slow-twitch muscle. Journal of Muscle Research and Cell Motility. 1982;3:191–212. doi: 10.1007/BF00711942. [DOI] [PubMed] [Google Scholar]