

Abstract
In keeping with the ‘muscular wisdom hypothesis’, many studies have documented that the firing rate of the majority of motor units decreased during fatiguing isometric contractions. The present study investigated whether the application of periodic muscle vibration, which strongly activates muscle spindles, would alter the modulation of motor unit firing rate during submaximal fatiguing isometric contractions.
Thirty-three motor units from the lateral head of the triceps brachii muscle were recorded from 10 subjects during a sustained isometric 20 % maximal voluntary contraction (MVC) of the elbow extensors. Vibration was interposed on the contraction for 2 s every 10 s. Twenty-two motor units were recorded from the beginning of the fatigue task. The discharge rate of the majority of motor units remained constant (12/22) or increased (4/22) with fatigue. Six motor units demonstrated a reduction in discharge rate that later returned toward initial values; these motor units had higher initial discharge rates than the other 16 motor units.
In a second series of experiments, four subjects held a sustained isometric 20 % MVC for 2 min and then vibration was applied as above for the remainder of the contraction. In this case, motor units initially demonstrated a decrease in firing rate that increased after the vibration was applied. Thus muscle spindle disfacilitation of the motoneurone pool may be associated with the decline of motor unit discharge rate observed during the first 2 min of the contraction.
In a third set of experiments, seven subjects performed the main experiment on one occasion and repeated the fatigue task without vibration on a second occasion. Neither the endurance time of the fatiguing contraction nor the MVC torque following fatigue was affected by the application of vibration. This finding calls into question the applicability of the muscular wisdom hypothesis to submaximal contractions.
The ‘muscular wisdom hypothesis’, coined by Marsden and colleagues (1983), proposed that the discharge rate of motor units slowed during sustained maximal voluntary contractions (MVCs) to minimize fatigue. Many studies have found that the firing rate of the majority of motor units decreased even during sustained submaximal isometric contractions (Person & Kudina 1972; Garland et al. 1994, 1997). However, Bigland-Ritchie et al. (1986) used tungsten microelectrodes to record a population response of motor units to a fatiguing submaximal contraction in quadriceps muscle. Although they found an increase in mean motor unit firing rate with fatigue, recruitment of new motor units could have accounted for the higher rates.
The aforementioned uncertainty in interpretation can be avoided by tracking single motor units throughout the fatiguing contraction. When motor units were followed throughout a fatiguing task, few motor units were found to demonstrate a decline in discharge rate when regular dynamic movements interrupted the isometric fatigue task (Miller et al. 1996; Griffin et al. 1998). Thus, a decline in motor unit firing rate is not an invariable feature of fatigue (see also Macefield et al. 2000). This raises the question as to the mechanisms underlying the decline in motor unit firing rate during isometric contractions as opposed to dynamic contractions. As muscle spindles are more active during dynamic than static contractions (Edin & Vallbo, 1990), it is possible that the repeated activation of muscle spindles during the dynamic contractions served to maintain motor unit discharge rates during fatiguing arm movements.
A reduction in Ia afferent activity was observed during the first minute of a sustained isometric 30 % MVC of the human dorsiflexor muscles (Macefield et al. 1991). Bongiovanni & Hagbarth (1990) found that muscle vibration (a potent stimulus for muscle spindles) did not increase the firing rate of motor units in non-fatigued muscle but vibration did augment force, EMG and motor unit firing rate during fatiguing isometric MVCs. However, prolonged periods (1–2 min) of vibration deepened the reduction in motor unit firing rate (Bongiovanni et al. 1990). Thus continuous high frequency spindle activation (as with vibration), or continuous low frequency spindle activation (as with isometric contraction) may cause a decrease in spindle support to the motoneurone pool. Brief periods of muscle stretch during movement may prevent muscle spindle disfacilitation, and consequently the decline in motor unit discharge rate in a dynamic fatigue protocol.
The purpose of this experiment was to investigate the effect of brief periodic muscle spindle activation on the motor unit discharge rate during sustained submaximal isometric contractions of the triceps brachii muscle. We hypothesized that if the fatigue-related modulation of motor unit firing rate during submaximal contractions could be prevented by vibration and if the muscular wisdom hypothesis applies to submaximal contractions, then the resultant fatigue should be exacerbated. Consequently, as a secondary objective, this experiment sought to determine the influence of any modulation of motor unit firing rate on force production during fatigue. A brief report of this work has appeared in abstract form (Griffin et al. 1999).
METHODS
Subjects
A total of 17 subjects (ages 21–45 years, 4 female) with no history of metabolic or neuromuscular disorder volunteered to participate in this study and signed a written consent form. The University Ethics Committee approved all experimental procedures, in accordance with the Helsinki declaration. Ten subjects (ages 21–40 years, 2 female) took part in the main experiment.
Experimental protocol
Subjects were seated in a chair with their right shoulder in 90 deg of flexion and the right forearm in front of them with the palm facing inward. The elbow was positioned at a 90 deg angle and secured with straps. Movement of the elbow was prevented by rigid supports at the elbow and the shoulder. The wrist was placed in a metal cuff mounted on a force transducer that measured elbow extension force (JR3, Inc., Woodland, CA, USA). A vibrator (110 Hz, 3 mm amplitude; T. Eaton Co., Toronto, Canada) was mounted in a device that held the vibrator securely against the distal tendon of the right triceps brachii muscle. A pulse generator (A.M.P.I., Jerusalem, Israel) controlled the onset and duration of muscle vibration. A subcutaneous fine-wire electrode (Formvar-coated stainless steel, 50 μm diameter; California Fine Wire Company, Grover Beach, CA, USA) was inserted with a hollow needle over the lateral head of the right triceps brachii muscle (Enoka et al. 1988). Surface EMG electrodes (0.8 cm in diameter) were placed 2 cm apart, longitudinally over the muscle bellies of the triceps brachii and biceps brachii muscles of the right arm.
At the beginning of the experiment, subjects performed two or three practice MVCs (with 3–5 min rest) to familiarize themselves with the protocol and to provide feedback on their performance. The contraction was considered to be maximal if two consecutive performances did not differ by more than 5 %. Following this practice session, two MVCs of the elbow extensors were recorded, the largest of which was considered to be the peak MVC torque. Five isometric ramp contractions (2 N m s−1) were performed to determine the threshold torque of motor unit recruitment. Subjects were instructed to hold an isometric contraction of the elbow extensors at 20 % MVC for as long as possible and were provided with verbal encouragement. Visual feedback of the force was displayed on a screen placed in front of the subject. While holding the 20 % MVC, the triceps brachii muscle was vibrated for 2 s every 10 s. Subjects were told not to adjust the voluntary force while the muscle was vibrated; that is, they were to continue contracting as they were in the previous 8 s. If the vibration induced an involuntary change in the force, subjects were instructed to return the force to the 20 % MVC target when the vibration was over. After the fatigue task, a narrow ischaemic cuff placed around the upper arm near the axilla was inflated to 300 mmHg for 3 min to prevent recovery during the performance of two post-fatigue MVCs and five ramp contractions.
In a second series of experiments, four subjects who participated in the main experiment (23–40 years, 1 female) held a sustained 20 % MVC for 2 min and then vibration was applied for 2 s every 10 s. This experiment was designed to observe the natural behaviour of the motor units to isometric fatigue and then identify any differences that occurred in discharge rate resulting from the application of vibration.
A third set of experiments was performed to investigate the impact of the application of vibration on the development of fatigue. Seven additional subjects (24–45 years, 3 female) performed the main experimental protocol on one occasion and repeated the sustained 20 % MVC fatigue task without vibration on a second occasion. The same subjects performed both experiments in order to control for the effects of individual differences such as gender and strength. The two fatigue tests were separated by at least 48 h and the order was alternated between subjects. Subjects did not know the results from the first test when they performed the second one. Only the force and triceps brachii surface EMG were recorded during these experiments. Thus, ramp contractions were not performed in the third experiment.
Data analysis
All data were recorded on VHS tape using a PCM interface (A. R. Vetter Corp., Rebersburg, PA, USA). The subcutaneous EMG was filtered at 10–10 000 Hz. Surface EMGs were recorded with a band-pass filter of 10–2000 Hz. The fatigue task was divided into a holding phase of 8 s and vibration phase of 2 s in duration.
The surface EMG recordings were digitized with a sampling rate of 4000 Hz, full-wave rectified and integrated over each holding phase. The mean integrated surface EMG was calculated for the first five holding phases and the five holding phases just before the endurance time. The force data were digitized with a sampling rate of 100 Hz and averaged separately for the holding phases and the vibration phases. The augmentation of force produced by vibration was calculated as the difference between the vibration and holding phases. The endurance time of the subject was defined as the time at which the holding force decreased by 2 % MVC or the coefficient-of-variation of the force within the holding phase reached 5 %.
Motor unit potentials were analysed offline with a template-matching algorithm (SPS-8701 waveform discrimination system, sampling rate 90 kHz; Signal Processing Systems, Prospect, SA, Australia) and post-processed further with a waveform subtraction algorithm (SPS-postproc). The threshold torque for motor unit recruitment was averaged over the five ramp contractions. Motor unit activity during the fatigue task was analysed separately during the 2 s vibration and the 8 s holding phases. The discharge rate of the motor unit during the first 8 s holding phase was considered to be the initial discharge rate. Examination of the interspike intervals (ISI) within each 8 s holding phase revealed a mean change of only 3.3 ms within each phase. Thus, the discharge rate within each 8 s holding phase was very stable. In seven experiments, the vibration produced artifacts in the subcutaneous EMG recording and thus motor unit discharge rate during the vibration phase could not be analysed in those experiments.
Statistical analysis
For each motor unit, the mean ISI for each vibration and holding phase was plotted separately. Subsequently, an autocorrelation time-series analysis was used to assess for changes over time. If the analysis indicated a change in the duration of motor unit ISIs over time, the changes were evaluated further by first dividing the duration of the experiment into thirds. The mean motor units ISIs in each third were compared using one-way ANOVA and post-hoc Bonferroni tests. Accordingly, the motor units were divided into groups depending on whether the motor units exhibited an increase, a decrease or no change in discharge rate.
Student's paired t tests were used to compare the MVC torque and the threshold torque for motor unit recruitment, before and after the fatigue task. Paired t tests were used to assess changes between the first 50 s and the last 50 s of the fatigue task in the integrated surface EMG. Endurance times for the fatiguing 20 % MVC isometric contractions with and without vibration were compared using paired t tests. Unless otherwise stated, the data in the text are presented as means ±s.d.
RESULTS
Muscle force and fatigue
Subjects held a mean force of 20.3 ± 1.5 % MVC for an endurance time of 4.7 ± 1.5 min. The presence of muscle fatigue was evidenced by a decrease in the peak MVC torque by 30.9 ± 11.7 % (55.7 ± 19.0 to 38.7 ± 11.6 N m) and an increase in the surface EMG of the triceps brachii muscle by 75.1 ± 65.8 %. The surface EMG of the biceps brachii muscle showed less than 2 % increase above the baseline and was considered to be negligible.
Muscle force during vibration
During the first 2 s application of vibration, the force increased significantly by 2.2 ± 1.5 % MVC above the 20 % MVC target force. The force augmentation during the vibration phase reduced to baseline values by the end of the task. Figure 1 displays the force and surface EMG for one subject. The force augmentation induced by vibration gradually declines, as does the ability of the subject to produce smooth sustained forces. The surface EMG remains fairly constant until it increases near the endurance limit.
Figure 1. Force and surface EMG during a representative experiment.
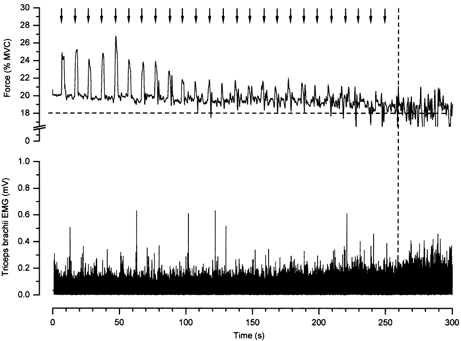
The elbow extension force (top) and the surface EMG from triceps brachii muscle (bottom) for an entire contraction in one subject are displayed. Arrows indicate the onset of the vibration (2 s in duration) and the initial augmentation of force associated with the vibration. The criteria for determining endurance time are displayed with dashed lines. At the endurance limit, the force in the holding phase dropped below 18 % MVC. Note the increase in surface EMG near the endurance limit.
Motor unit recruitment
The activity of 33 motor units from the 10 subjects was recorded. Twenty-two motor units were active from the beginning of the task and 11 were newly recruited as the muscle fatigued. Before fatigue, the threshold torque for motor unit recruitment ranged from 4.4 to 36.6 % of the prefatigue MVC. The mean threshold torque for motor unit recruitment before fatigue was 12.6 ± 5.1 % MVC for motor units active from the beginning of the task and 26.7 ± 7.3 % MVC for motor units newly recruited during the fatigue task. Two of the newly recruited motor units were not recruited during the ramps. Overall, the mean threshold torque for motor unit recruitment declined significantly by 50.1 ± 30.9 % (from 16.6 ± 8.3 to 8.2 ± 5.6 % prefatigue MVC) following fatigue. An example of a motor unit that was recruited initially during vibration and then became active during the holding phase is depicted in Fig. 2. Before becoming continuously active during the holding phase, the motor unit fired intermittently (Fig. 2B).
Figure 2. Force and motor unit recordings as fatigue progresses.
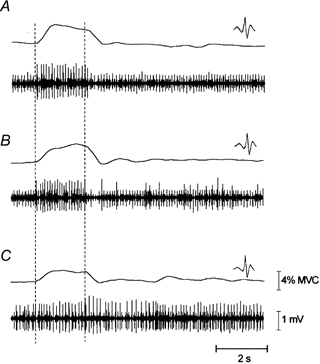
The muscle force and triceps brachii subcutaneous EMG for three trials of one subject during the fatigue task are displayed. The periods of vibration are indicated with dashed lines. The force during the vibration is higher at the beginning of the task (A, 30 s into the contraction) and progressively reduces as the muscle fatigues (C, 5 min of contraction). The waveform of the large amplitude motor unit is displayed. This unit is initially active only during the vibration phase (A), becomes intermittently active during the holding phase (B, 2.5 min of contraction) and is then continuously active during both the vibration and holding phase (C).
Motor unit discharge rates during holding phase
The majority (16/22) of the motor units that were active from the beginning of the task demonstrated either an increase (4/22) or no significant change (12/22) in discharge rate during the fatigue task (Fig. 3). Only six motor units were found to have a decrease in discharge rate; even these returned to higher values by the end of the fatigue task. The mean firing rate of the motor units are presented in Fig. 3, as a percentage of the initial discharge rate. Motor units are grouped according to the discharge behaviour revealed by autocorrelation time-series analysis and subsequent one-way ANOVA. The 16 motor units that demonstrated no change or an increase in discharge rate by the end of the fatigue task had significantly higher prefatigue recruitment thresholds (13.9 ± 4.1 vs. 10.7 ± 2.7 % MVC) and lower initial firing frequencies (10.9 ± 2.4 vs. 13.1 ± 1.9 Hz) than the six units that displayed a decline.
Figure 3. Modulation of motor unit discharge rate during the main experiment.
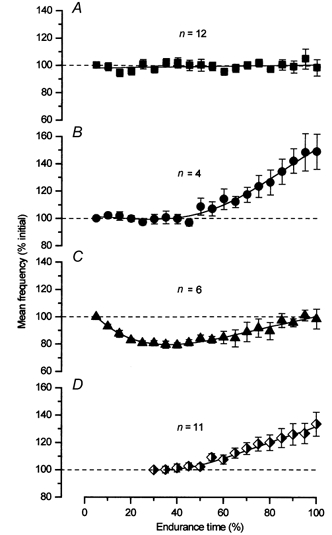
The mean ±s.e.m. firing frequencies of all motor units throughout the task are grouped according to the type of modulation in discharge rate during fatigue. The data for each motor unit are normalized to its initial firing frequency. Panels A, B and C represent motor units that were active from the beginning of the contraction; panel D represents the units that were recruited during the fatigue task. The majority of the units that were active from the beginning (12/22) did not demonstrate a change in discharge rate with fatigue (A). Four motor units demonstrated an increase in discharge rate (B), while six were found to decrease and then return to baseline (C). The newly recruited units (D) showed an increase in discharge rate (9/11).
Eleven motor units were recruited during the fatigue task (Fig. 3D). These motor units started to discharge with low initial firing rates (7.8 ± 1.7 Hz) and 9/11 motor units exhibited an increase in discharge rate over time. No newly recruited motor units demonstrated a decline in discharge rate. Note that the recruitment of additional motor units occurred at approximately the same time as the firing rate of motor units in panels B and C increased.
Motor unit discharge rate during vibration
Seven motor units could be followed during the 2 s vibration periods. All these motor units were active in the holding phases as well; 5/7 were active in the holding phase from the beginning of the contraction and 2/7 were recruited during the holding phase at about 40 % of endurance time. The discharge rate of the motor units during the vibration phase paralleled that found in the holding phase. That is, if the motor unit discharge rate remained relatively constant in the holding phase, it did so in the vibration phase as well. This behaviour is depicted in Fig. 4 with two motor units from different subjects. In each panel, the discharge rate of the same motor unit is plotted over time during the 2 s vibration periods and during the 8 s holding phases. It is interesting to note that in one subject the discharge rate was higher during vibration than during the holding phase, yet in the other subject, it was lower. The torque augmentation that occurred during vibration was different in these two subjects. In the experiment depicted in the top panel, the vibration resulted in a 4 % MVC torque augmentation, whereas in the bottom experiment, the augmentation was only 1.2 % MVC. Overall, 5/7 motor units demonstrated higher discharge rates in vibration than in the holding phase. The torque augmentation in these experiments ranged from 4–6 % MVC. The 2/7 motor units that demonstrated lower discharge rates in vibration than in the holding phase were from the same subject who exhibited the smallest amount of torque augmentation from these experiments. Consistent with the small torque augmentation, there was no evidence of recruitment of additional motor units during the vibration phase in that subject.
Figure 4. Discharge rate of two motor units during the vibration and holding phases.
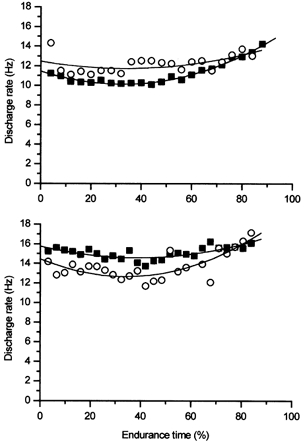
Discharge rates of single motor units in two different experiments during the 2 s vibration (○) and 8 s holding phase (▪) are presented. The motor unit displayed in the top panel had a higher discharge rate during vibration than the holding phase. Another subject exhibited a lower motor unit discharge rate during the vibration than the holding phase (bottom panel).
Motor unit discharge rate during sustained contraction without/with vibration
In the second experiment, we analysed action potentials from seven motor units during a sustained isometric 20 % MVC in which the application of vibration started after 2 min of contraction. The initial firing rate of these motor units was 10.2 ± 1.6 Hz. This firing rate was similar to that of the motor units that demonstrated an increase or no change in firing rate in the main experiment (10.9 ± 2.4 Hz). As can be seen in Fig. 5, the motor units initially demonstrated a decrease in firing rate that increased again after the vibration was applied. The initial decline in discharge rate follows the same pattern as for motor units recorded in a previous experiment in which subjects performed sustained isometric 20 % MVC fatiguing contractions (Garland et al. 1997).
Figure 5. Mean discharge rate of motor units with and without vibration.

The lines represent data from three different experimental sets: 1, main experiment (n = 22); 2, second experiment (n = 7); 3, previous experiment (20 % MVC sustained isometric fatiguing contraction with no vibration applied (n = 20) (Garland et al. 1997). The mean firing rate for every 10 s from the start of the fatiguing contraction without vibration (8 s for the experiments with vibration) was calculated for each motor unit. The individual motor unit data were plotted against the subject's endurance time and the curves of best fit were determined. These curves were averaged for each of the three experiments in order to summarise the behaviour of motor units with and without vibration.
Comparison of endurance limits
The muscular wisdom hypothesis suggested that a slowing of motor unit firing rate during a sustained MVC would minimize fatigue. Considering that few motor units demonstrated a decrease in discharge rate during the main experiment, a separate set of experiments was conducted to investigate the impact of this maintenance of motor unit firing rate on the development of fatigue under submaximal conditions. The endurance time and the MVC torque were compared between two fatigue protocols, one with and one without vibration.
The endurance times did not differ between the two tasks. The mean endurance time for the sustained 20 % MVC with and without vibration was 5.4 ± 1.3 and 5.2 ± 1.5 min, respectively. The decline in MVC torque following fatigue was also similar in the two cases (45.8 ± 12.7 % without vibration and 45.6 ± 13.7 % with vibration). Thus the application of vibration, and the subsequent maintenance of motor unit discharge rate, did not have a negative effect on endurance time or the extent of fatigue.
DISCUSSION
The main findings from this study were that (1) motor unit firing rate is maintained in submaximal isometric contractions with the periodic activation of muscle spindles and (2) manipulation of the motor unit discharge rate by the application of vibration did not affect the functional outcome of the fatiguing protocol. These findings shed new light on the muscle wisdom hypothesis and the mechanisms underlying the modulation of motor unit firing rate during fatigue.
Motor unit discharge rate during vibration
We expected that the vibration-induced excitatory input from the spindles onto the motoneurone pool would increase the discharge rate of previously active motor units or recruit additional motor units during the vibration. Although we were able to analyse only seven motor units during the vibration phase, motor unit recruitment was evident during the vibration in one experiment (see Fig. 2). Interestingly, the motor unit discharge rate during vibration did not always increase. In Fig. 4, two motor units are depicted during the 2 s vibrations and during the 8 s holding phases. In the top panel, the vibration was associated with an increase in discharge rate over that in the holding phase and also a large torque augmentation. In the bottom panel, the discharge rate of the motor unit was less than that found in the holding phase and the torque augmentation was modest. Because subjects were provided with visual feedback of the force, one subject may have lowered the central drive to the motor units during the vibration (knowing that the vibration will assist the contraction). This would have resulted in a smaller torque augmentation and a lower motor unit discharge rate during the vibration. In other subjects, the vibration may have added excitation to an unchanged central drive resulting in larger force augmentation and an increase in motor unit firing rate.
In the present study, the added increment of force imposed by vibration lessened as fatigue progressed. Cafarelli & Layton-Wood (1986) found that the increased sensation of force that occurred during vibration in non-fatigued muscle did not occur if the muscle was fatigued. Perhaps the subjects participating in that study did not sense any additional force after fatigue because there had not been any additional force evoked. The development of muscle fatigue could have offset any incremental force that otherwise would have resulted from the vibration. However, it is hard to ascertain in the present experiment whether the vibration evoked less force during fatigue or whether the subjects learned to relax the muscle during the vibration period by the end of the task.
The most important aspect of the motor unit discharge rate during vibration for the present study is that the changes in motor unit discharge rate over time were similar whether the motor unit was recorded during vibration or during the holding phase. This verifies that the data recorded during the holding phases are a valid representation of the effects of spindle activation on motor unit discharge rate (see below).
Motor unit discharge rate during the holding phase
Discharge rate in the majority of motor units did not decline during a sustained isometric 20 % MVC that was interrupted by brief periodic tendon vibration. However, the firing rate of the majority of motor units declined from the onset of contraction during a sustained 20 % MVC of the triceps brachii muscle without vibration (Garland et al. 1997). In addition, few motor units demonstrated a decline in firing rate when the contraction was interrupted by repeated arm movements (Miller et al. 1996; Griffin et al. 1998). When an isometric force pulse protocol was used to recreate the motor unit activity patterns that occur during a dynamic contraction (but without the movement), the firing rate of the majority of motor units also decreased (Griffin et al. 2000). Together these findings suggest that muscle spindle activation can prevent the reduction in motor unit firing rate that typically occurs during submaximal isometric fatigue tasks. The reduction in motor unit firing rate demonstrated in isometric contractions may represent a unique situation and may not be relevant for most activities of daily living that incorporate movement (and consequently, spindle assistance to the motoneurone pool).
Only 6 of the 22 motor units that were active from the beginning of the task displayed a temporary reduction in discharge rate with fatigue that later increased back toward baseline (Fig. 3). An increase in motor unit discharge rate following an initial decline has been observed previously during sustained fatiguing submaximal isometric MVCs of the triceps brachii (Garland et al. 1997) and biceps brachii (Garland et al. 1994) muscles and during intermittent 20 % MVCs of the biceps brachii muscle (Dorfman et al. 1990). The reversal of discharge rate may stem from an increase in central drive to maintain the target force as the active motor units fatigue or possibly from an increase in fusimotor activity. For instance, during 40 Hz stimulation of the triceps surae muscles of the decerebrated cat, Ljubisavljevic et al. (1994) found that the initial reduction in Ia afferent discharge rate gradually returned to above baseline values by a few minutes of contraction. The late increase was proposed to stem from activation of chemoreceptive group III and IV afferents (see Ellaway et al. 1982; Appelberg et al. 1983; Jovanovic et al. 1990).
The question of the role of muscle spindles during fatigue has been of interest since Marsden et al. (1976) proposed that an increased gain of the stretch reflex occurs following fatigue. Several investigations confirmed that the amplitude of the EMG response to muscle stretch is greater following fatiguing exercise in humans (Darling & Hayes 1983; Hakkinen & Komi 1983; Kirsch & Rymer 1987). However, Balestra et al. (1992) demonstrated a decrease in the stretch reflex following fatigue evoked by repetitive stretch-shortening cycles. Even in non-fatiguing contractions, Suzuki et al. (1990) found that repeated isometric contractions resulted in a decrease of motor unit recruitment threshold force that could be ‘reset’ by passive stretch of the muscle. The thixotropic nature of intrafusal actin-myosin bonds can also increase magnitude of the stretch reflex (see Proske et al. 1993, for review). It is possible that the thixotropic nature of the spindle influences motor unit discharge behaviour during fatigue. Although the mechanisms whereby the spindle activity is integrated with motoneurone firing are not addressed, the present study affirms the role of muscle spindle input in maintaining motor unit discharge rate during fatiguing contractions.
Muscular wisdom
The muscular wisdom hypothesis proposed that a reduction in motor unit firing rate occurs to minimize fatigue. This hypothesis was developed originally using sustained MVCs as a model (Marsden et al. 1983; Bigland-Ritchie et al. 1983a, b). The applicability of the muscle wisdom hypothesis to submaximal fatiguing contractions has been questioned. Force-frequency curves have been found to shift to the right instead of the left during various fatigue protocols (e.g. Edwards et al. 1977; Binder-Macleod & McDermond 1992; Fuglevand 1996). That is, higher frequencies of stimulation (not lower as predicted by the muscular wisdom hypothesis) were required to produce a 50 % MVC after fatigue than before fatigue. For single motor units in the human thenar muscle, the frequency required to generate 50 % MVC either increased or decreased depending on whether the muscle was potentiating or fatiguing, respectively (Thomas et al. 1991). In addition, the majority of motor units demonstrated a decline in firing rate during a sustained 20 % MVCs of the triceps brachii muscle with no change in whole muscle half-relaxation time (Garland et al. 1997).
The present investigation demonstrated that the prevention of a reduction in motor unit discharge rate did not influence the endurance time nor did it create a precipitous decline in MVC torque. Interestingly, the mean endurance time for the third experiment was slightly longer and the decline in the MVC torque was greater as compared to the main study in which motor units were recorded (5.4 vs. 4.7 min and 46 vs. 31 % MVC torque decline in the third and main experiments, respectively). The differences in the prefatigue MVC torque of subjects may affect the endurance time and the MVC decline following fatigue. The subjects had a mean MVC of 44.7 ± 20.2 N m in the third experiment, whereas those in the main study had a mean MVC of 55.7 ± 19.0 N m. Thus, stronger subjects have shorter endurance times and smaller declines in MVC following fatigue.
If the lowering of motor unit firing rate during sustained submaximal contractions was serving to optimize force by decreasing the rate of fatigue, then maintaining the motor unit firing rate in the present investigation ought to have shortened the endurance time or resulted in a greater decline in MVC torque. This did not occur. Thus, the functional relevance of the modulation of motor unit firing rate during fatiguing submaximal isometric contractions is unknown.
Acknowledgments
This work was supported by a grant awarded to S. J. Garland from Natural Sciences and Engineering Research Council, Canada and NSERC PGS-B scholarships awarded to L. Griffin and R. Gossen.
References
- Appelberg B, Hulliger M, Johansson H, Sojka P. Actions on gamma-motoneurones elicited by electrical stimulation of group III muscle afferent fibres in the hind limb of the cat. Journal of Physiology. 1983;335:275–292. doi: 10.1113/jphysiol.1983.sp014533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balestra C, Duchateau J, Hainaut K. Effects of fatigue on the stretch reflex in a human muscle. Electroencephalography and Clinical Neurophysiology. 1992;85:46–52. doi: 10.1016/0168-5597(92)90101-g. [DOI] [PubMed] [Google Scholar]
- Bigland-Ritchie B, Cafarelli E, Vollestad NK. Fatigue of submaximal static contractions. Acta Physiologica Scandinavica. 1986;128(suppl. 556):137–148. [PubMed] [Google Scholar]
- Bigland-Ritchie B, Johansson R, Lippold OCJ, Smith S, Woods JJ. Changes in motoneurone firing rates during sustained maximal voluntary contractions. Journal of Physiology. 1983a;340:335–346. doi: 10.1113/jphysiol.1983.sp014765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bigland-Ritchie B, Johansson R, Lippold OCJ, Woods JJ. Contractile speed and EMG changes during fatigue of sustained maximal voluntary contractions. Journal of Neurophysiology. 1983b;50:313–324. doi: 10.1152/jn.1983.50.1.313. [DOI] [PubMed] [Google Scholar]
- Binder-Macleod SA, McDermond LR. Changes in the force-frequency relationship of the human quadriceps femoris muscle following electrically and voluntarily induced fatigue. Physical Therapy. 1992;72:95–104. doi: 10.1093/ptj/72.2.95. [DOI] [PubMed] [Google Scholar]
- Bongiovanni LG, Hagbarth KE. Tonic vibration reflexes elicited during fatigue from maximal voluntary contractions in man. Journal of Physiology. 1990;423:1–14. doi: 10.1113/jphysiol.1990.sp018007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bongiovanni LG, Hagbarth KE, Stjernberg L. Prolonged muscle vibration reducing motor output in maximal voluntary contractions in man. Journal of Physiology. 1990;423:15–26. doi: 10.1113/jphysiol.1990.sp018008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cafarelli E, Layton-Wood J. Effect of vibration on force sensation in fatigued muscle. Medicine and Science in Sports and Exercise. 1986;18:516–521. [PubMed] [Google Scholar]
- Darling WG, Hayes KC. Human servo responses to load disturbances in fatigued muscle. Brain Research. 1983;267:345–351. doi: 10.1016/0006-8993(83)90887-9. [DOI] [PubMed] [Google Scholar]
- Dorfman LJ, Howard JE, McGill KC. Triphasic behavioral response of motor units to submaximal fatiguing exercise. Muscle and Nerve. 1990;13:621–628. doi: 10.1002/mus.880130711. [DOI] [PubMed] [Google Scholar]
- Edin BB, Vallbo AB. Dynamic response of human muscle spindle afferents to stretch. Journal of Neurophysiology. 1990;63:1297–1306. doi: 10.1152/jn.1990.63.6.1297. [DOI] [PubMed] [Google Scholar]
- Edwards RHT, Hill DK, Jones DA, Merton PA. Fatigue of long duration in human skeletal muscle after exercise. Journal of Physiology. 1977;272:769–778. doi: 10.1113/jphysiol.1977.sp012072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellaway PH, Murphy PR, Tripathi A. Closely coupled excitation of gamma motoneurones by group III muscle afferents with low mechanical threshold in the cat. Journal of Physiology. 1982;331:481–498. doi: 10.1113/jphysiol.1982.sp014385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Enoka RM, Robinson GA, Kossev AR. A stable, selective electrode for recording single motor-unit potentials in humans. Experimental Neurology. 1988;99:761–764. doi: 10.1016/0014-4886(88)90189-6. [DOI] [PubMed] [Google Scholar]
- Fuglevand AJ. Re-evaluation of muscle wisdom using physiological stimulation rates. 26th Annual Meeting, Washington, Society for Neuroscience Abstracts. 1996;22(1098):436.4. [Google Scholar]
- Garland SJ, Enoka RM, Serrano LP, Robinson GA. Behavior of motor units in human biceps brachii during a submaximal fatiguing contraction. Journal of Applied Physiology. 1994;76:2411–2419. doi: 10.1152/jappl.1994.76.6.2411. [DOI] [PubMed] [Google Scholar]
- Garland SJ, Griffin L, Ivanova T. Motor unit discharge rate is not associated with muscle relaxation time in sustained submaximal contractions in humans. Neuroscience Letters. 1997;239:25–28. doi: 10.1016/s0304-3940(97)00885-9. [DOI] [PubMed] [Google Scholar]
- Griffin L, Garland SJ, Ivanova T. Discharge patterns in human motor units during fatiguing arm movements. Journal of Applied Physiology. 1998;85:1684–1692. doi: 10.1152/jappl.1998.85.5.1684. [DOI] [PubMed] [Google Scholar]
- Griffin L, Garland SJ, Ivanova T. Muscle spindle activation can sustain motor unit firing rates during submaximal fatigue. 29th Annual Meeting, Miami Beach, Society for Neuroscience Abstracts. 1999;25(part 1):1146. [Google Scholar]
- Griffin L, Ivanova T, Garland SJ. Role of limb movement in the modulation of motor unit discharge rate during fatiguing contractions. Experimental Brain Research. 2000;130:392–400. doi: 10.1007/s002219900253. [DOI] [PubMed] [Google Scholar]
- Hakkinen K, Komi PV. Electromyographic and mechanical characteristics of human skeletal muscle during fatigue under voluntary and reflex conditions. Electroencephalography and Clinical Neurophysiology. 1983;55:436–444. doi: 10.1016/0013-4694(83)90132-3. [DOI] [PubMed] [Google Scholar]
- Jovanovic K, Anastasijevic R, Vuco J. Reflex effects on gamma fusimotor neurones of chemically induced discharges in small-diameter muscle afferents in decerebrate cats. Brain Research. 1990;521:89–94. doi: 10.1016/0006-8993(90)91528-o. [DOI] [PubMed] [Google Scholar]
- Kirsch RF, Rymer WZ. Neural compensation for muscular fatigue: evidence for significant force regulation in man. Journal of Neurophysiology. 1987;57:1893–1910. doi: 10.1152/jn.1987.57.6.1893. [DOI] [PubMed] [Google Scholar]
- Ljubisavljevic M, Jovanovic K, Anastasijevic R. Fusimotor responses to fatiguing muscle contractions in non-denervated hindlimb of decerebrate cats. Neuroscience. 1994;61:683–689. doi: 10.1016/0306-4522(94)90444-8. [DOI] [PubMed] [Google Scholar]
- Macefield G, Hagbarth K-E, Gorman R, Gandevia SC, Burke D. Decline in spindle support to α-motoneurones during sustained voluntary contractions. Journal of Physiology. 1991;440:497–512. doi: 10.1113/jphysiol.1991.sp018721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macefield VG, Fuglevand AJ, Howell JN, Bigland-Ritchie B. Discharge behaviour of single motor units during maximal voluntary contractions of a human toe extensor. Journal of Physiology. 2000;528:227–234. doi: 10.1111/j.1469-7793.2000.00227.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marsden CD, Meadows JC, Merton PA. Muscular wisdom’ that minimizes fatigue during prolonged effort in man: Peak rates of motoneuron discharge and slowing of discharge during fatigue. In: Desmedt JE, editor. Motor Control Mechanisms in Health and Disease. New York: Raven Press; 1983. pp. 169–211. [PubMed] [Google Scholar]
- Marsden CD, Merton PA, Morton HB. Servo action in the human thumb. Journal of Physiology. 1976;257:1–44. doi: 10.1113/jphysiol.1976.sp011354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller KJ, Garland SJ, Ivanova T, Ohtsuki T. Motor-unit behavior in humans during fatiguing arm movements. Journal of Neurophysiology. 1996;75:1629–1636. doi: 10.1152/jn.1996.75.4.1629. [DOI] [PubMed] [Google Scholar]
- Person RS, Kudina LP. Discharge frequency and discharge pattern of human motor units during voluntary contraction of muscle. Electroencephalography and Clinical Neurophysiology. 1972;32:471–483. doi: 10.1016/0013-4694(72)90058-2. [DOI] [PubMed] [Google Scholar]
- Proske U, Morgan DL, Gregory JE. Thixotropy in skeletal muscle and in muscle spindles: a review. Progress in Neurobiology. 1993;41:705–721. doi: 10.1016/0301-0082(93)90032-n. [DOI] [PubMed] [Google Scholar]
- Suzuki S, Hayami A, Suzuki M, Watanabe S, Hutton RS. Reductions in recruitment force thresholds in human single motor units by successive voluntary contractions. Experimental Brain Research. 1990;82:227–230. doi: 10.1007/BF00230858. [DOI] [PubMed] [Google Scholar]
- Thomas CK, Johansson RS, Bigland-Ritchie B. Attempts to physiologically classify human thenar motor units. Journal of Neurophysiology. 1991;65:1501–1508. doi: 10.1152/jn.1991.65.6.1501. [DOI] [PubMed] [Google Scholar]