

Abstract
Interpreting receiver responses to on-territory playback of aggressive signals is problematic. One solution is to combine such receiver-perspective experiments with a sender-perspective experiment that allows subjects to demonstrate how their choice of singing strategies is associated with their approach behavior. Here we report the results of a sender-perspective study on the banded wren (Thryothorus pleurostictus), and combine information on context and results of previous receiver-perspective experiments to clarify function. Territorial males were presented with a 5-min playback consisting of song types present in their repertoire. We assessed the degree to which the subjects’ song matching rate, overlapping rate, and song-type versatility were correlated with their approach latency, closeness of approach, latency to first retreat, and time spent close to the speaker. Male age, breeding stage, and features of the playback stimuli were also considered. Song matching was associated with rapid and close approach, consistent with the receiver-perspective interpretation of type matching as a conventional signal of aggressive motivation. Overlapping was associated with earlier retreat, and together with the aversive receiver response to our previous overlapping playback experiment suggests that overlapping is a defensive withdrawal signal. High versatility was associated with slower first retreat from the speaker and high levels of reciprocal matching between subject and playback. Males with fledglings sang with particularly low versatility and approached the speaker aggressively, whereas males with nestlings overlapped more and retreated quickly. Finally, older males matched more but overlapped less.
Keywords: Aggressive motivation, playback design, path analysis, signal function, singing versatility, song-type repertoires
Identifying the precise function of signals and the information they encode is a major focus of animal communication research (Otte, 1974; Bradbury and Vehrencamp, 1998; Searcy and Nowicki 2005). Researchers have three types of data from which they can hope to infer signal function: 1) the ambient contexts that a sender might encode in its signals, including sender characteristics such as sex, age, condition, and motivation, and external characteristics such as nearby conspecifics, predators, and food; 2) the responses of the receiver to the signals, including approach and retreat behaviors; and 3) the sender’s associated behavior, which may or may not be dependent on the receiver’s response. The function of some signals may be fairly straightforward to resolve, for example, begging signals that increase with food deprivation and cause appropriate receivers to feed the sender (Kilner, 2002), and alarm signals associated with the appearance of a predator that cause receivers to flee (Seyfarth et al., 1980). Determining the precise function of signals given in agonistic contexts is more difficult because a) there are usually several to many of these signals in the repertoire of a given species, b) signals are given in chains with the two parties alternating sender and receiver roles, and c) the range of possible functions at each step in a chain is large, e.g., challenge, directed rival pointing, intention to attack or retreat, motivation to escalate or de-escalate, dominance, submission, fear, ambivalence, fighting ability, defensive action, and victory declaration (Andersson, 1980; Archer, 1988; Bradbury and Vehrencamp, 1998).
The calls and songs of birds that defend territories acoustically have long been popular signals for functional studies because of the tractability of recording the signals, observing behavioral interactions, conducting sound playback experiments, and reliably locating the subjects (Collins, 2004). The most common protocol is to present territorial owners with alternative signal-variant treatments (simultaneous or sequential), and then to quantify their approach to the speaker. Such experiments are designed to investigate the receiver perspective of the communication dyad (data strategy 2 above). However, Searcy and Nowicki (2000) and others have argued that the differential responses of territory owners to putatively aggressive signal variants broadcast on a subject’s territory cannot be used to infer relative threat function. On the one hand, a sufficiently threatening signal ought to elicit retreat or a weak approach response. On the other, a highly motivated territorial owner might elect to escalate the contest and exhibit a stronger approach response to the more threatening signal. Receiver responses confound threat level with relative receiver condition and motivation to defend the territory. The evidence does indeed indicate that playback of the signal variant believed to be the more aggressive threat sometimes keeps territory owners at a greater distance, as demonstrated for song overlapping (Todt and Naguib, 2000; Osiejuk et al. 2004), dominant frequency (Hardouin et al., 2007), frequency matching (Mennill and Ratcliffe, 2004), and certain note types (Järvi et al., 1980)), and sometimes elicits a stronger approach, as demonstrated for song-type matching (Krebs et al., 1981; Burt et al., 2001; Nielsen and Vehrencamp, 1995), trill length (Leitao and Riebel, 2003), and frequency modulation (Slabbekoorn and tenCate, 1997).
One alternative protocol for testing the repulsive effect of territorial signal variants is a speaker-replacement experiment, in which a territory owner is removed and replaced by a speaker broadcasting the signal treatments. Stronger threat signals are expected to keep intruders out of the territory for a longer period (Searcy and Nowicki 2000). This approach still has many problems, since receivers are a mixture of neighbors and floaters who may vary in their motivation to take over a new area, and who may not rank the relative salience of threat signals in the same way as territory defenders. Moreover, testing the keep-out function of matching and overlapping signal variants is logistically challenging because intruders often do not sing and their repertoires are unknown. Another alternative protocol is to use a lure to bring the subject very close to the speaker before playing the treatment. A stronger or faster movement away from the speaker implies retreat from a more threatening stimulus. However, subjects who receive an extremely threatening treatment first may remember the lure when the second, less threatening, stimulus is presented, so careful design and analysis of order effects are required (Beecher and Campbell, 2005; Hall et al., 2006).
Investigating the sender’s perspective (data strategies 1 and 3 above) provides a third alternative or supplementary strategy for resolving signal function. Careful documentation of the precise context of signal-variant use and associated sender acts during natural aggressive encounters may identify sender motivation or intentions (Kramer et al., 1985; Simpson, 1985; Weary et al., 1988; Smith and Smith, 1996). However, whether attack follows the presumed more aggressive signal depends on whether the opponent retreats in response to the threat, an effect which must be examined and controlled for with proper conditional statistical analyses (Waas, 1991). A better method is to present territorial subjects with an experimental opponent, allow them to signal in ways of their own choosing, and monitor their subsequent actions (Vehrencamp, 2001; Searcy et al., 2006). The context and opponent acts are more controlled than in natural interactions, and potential cause and effect relationships between vocal signals and prior or subsequent movements can be evaluated.
We used the latter strategy in a study of male singing interactions in the banded wren Thryothorus pleurostictus. Males in this neotropical, sedentary, territorial songbird possess song-type repertories of about 25 discrete types, share a large fraction of these types with neighbors, and generally sing with immediate variety (Molles and Vehrencamp, 1999). Vigorous singing occurs throughout the 5-month breeding season, both during a dawn chorus and at intervals throughout the day. Counter-singing increases during boundary disputes, indicating that song plays an important role in territory defense (Trillo and Vehrencamp, 2005). We focused on three singing patterns with known receiver responses from prior alternative-treatment playback studies: song-type matching, song overlapping, and variation in song-type switching rate and diversity. These singing patterns all involve song-type choice or timing that can be performed in principle by all individuals and changed based on momentary decisions or sender motivation. We subjected territorial males to a long playback (5 min) consisting of six song types similar to ones in their own repertoires. We then examined the associations between the males’ rate of matching, overlapping, and switching during the playback and their approach behavior, along with known contexts such as male age, breeding stage, presence of the female, and time of day. Path analysis was used to assess possible cause and effect relationships between singing and approach behaviors. In the discussion we compare these sender associations with the results of previous receiver-response experiments.
METHODS
Study site and subjects
We conducted the playback experiment from 2–29 July 2005 in Santa Rosa National Park in the Guanacaste Conservation Area of northwestern Costa Rica (10° 51′ N, 85° 38′ W). The habitat consists of tropical dry deciduous forest, mixed with occasional patches of regenerating secondary growth. For details on habitat, breeding season, and study population see Molles and Vehrencamp (1999). The 34 subjects were individually color-banded males known to be resident on their territories for 1 to 5 years (absolute age was known for 12 males). Mated status, breeding stage, and territorial boundaries were monitored from May through August in the 2005 breeding season.
Playback protocol
Stimulus songs with high signal-to-noise ratios were selected from recordings of 33 males from the same neighborhood, but not adjacent to subject males. Source males were recorded from approximately 0500 to 0700, which included the dawn chorus, using a Sennheiser ME67 directional microphone and Marantz PMD 690 digital solid-state recorder. Exemplars of six song types in the repertoire of the subject were selected from the same source male, noise was filtered out using the cursor-delimited band-pass filter in Syrinx (J. Burt, http://syrinxpc.com), and songs were amplified to the same peak amplitude. Each song type was pasted four times at intervals of about 12.5 sec before switching to the next song type to create a 5-min stimulus sound file with 24 songs. Stimulus sound files were played from a Creative Nomad Jukebox 3 through an Anchor Audio Mini-Vox PB-25 speaker at levels approximating natural song (90 dB at 1m).
The playback trials were conducted between 0700 and 0930 hr from a central position on the territory. The speaker was placed 1.5 to 2 m high and a 15 m radius circle was marked with flagging around the speaker. Trials lasted 15 min, with a 5-min pre-playback period, a 5-min playback period, and a 5-min post playback period. This simulation represents a highly aggressive intrusion by a stranger, which would only be observed during a territory take-over. The entire trial was recorded with the same recording equipment mentioned above, and two additional observers scored the timing of movements by the focal male in and out of the 15 m circle using time-synchronized handheld computers (Palm 100). All three observers communicated behavioral observations and estimated distances of the male from the speaker with walkie-talkies, which were recorded on the second channel of the stereo recorder.
From the audio and computer records of each trial, we scored the following spatiotemporal responses: latency from start of playback to first approach toward the loudspeaker (flight of at least 2 m); time of entry into the 15 m circle; latency of first retreat out of the circle; closest approach of the subject to the loudspeaker at any time during the trial; and amount of time spent within the circle (computed separately for during- and post-playback periods and with multiple entrances into the circle summed). For two birds that did not enter the 15 m circle, we made an equivalent measure of their latency to first retreat after their closest approach. The following vocal events were quantified for each phase of the experiment (pre-, during, and post-playback): the number of songs sung by the focal male, the song type and length of each song, and the occurrence of temporal overlapping of the subject’s and playback’s songs.
With this information we computed the three key singing variables of interest. 1) Matching rate - the proportion of the male’s songs that matched a playback song type. Although we distinguished between immediate matches (male matched most recent playback song type) and delayed matches (male matched any other earlier playback song type), both types of matches were summed for the primary analysis of during-playback matching rate to obtain a normally distributed variable. Delayed matches up to a maximum lag of 5 min were therefore included with this scoring method, which seems to be biologically meaningful (Molles, 2006; Burt and Vehrencamp, unpubl. microphone array recordings). 2) Overlapping rate - the proportion of the male’s songs that overlapped the playback (subject started to sing more than 0.2 sec before playback song had ended). 3) Song-type versatility – computed as switching rate × diversity, where switching rate = number of switches to a different song type per songs sung, and diversity = number of song types per songs sung. Switching rate and diversity were highly correlated with each other (r = 0.569, P = 0.001, n = 30, excludes 4 males that sang fewer than 5 songs). The derived versatility variable was more normally distributed and produced better residual plots in the multivariate analyses than did switching rate or song-type diversity alone. This versatility index ranges from 0 for completely repetitive delivery of one song type to 1.0 for switching to a new type with each subsequent song. In addition to these singing patterns, we scored song rate (songs/min) during each phase of playback, and the mean length of male songs delivered during and after playback. We also noted whether the subject’s immediate matches and overlaps of the playback occurred before, during, or after his closest approach to the speaker.
From the audio record we scored three characteristics of the playback stimulus: the number of playback songs that matched the subject’s songs (sum of immediate and delayed matches by playback, including playback delayed matches of subject songs sung in the 5-min pre-playback period), the number of playback songs that overlapped the subject’s songs (playback song started at least 0.2 sec before male’s song ended), and the mean length of playback songs. Several additional context variables were included in the final dataset: whether or not the female mate responded by vocalizing or approaching the speaker; start time of the playback; breeding stage of the subject, with categories of nest-building, incubating, feeding nestlings, feeding fledglings, or no known nest (including cases of recent predation and apparently unpaired males); and minimum male age (years since first banded).
Statistical analyses
We transformed continuous variables with skewed distributions using natural log, square-root, or arcsine as appropriate to normalize residual distributions and achieve linear pairwise relationships. To identify variables associated with each singing pattern, we first performed separate forward stepwise general linear model (GLM) analyses on each of the three key singing measures during the 5-min playback period (matching rate, overlapping rate, or versatility) as dependent variables. Independent variables included latency to first approach, latency from approach to first retreat, proportion of time within the circle during plus post playback, closest approach, male song rate in the pre- and during-playback periods, male song length, the other two target singing variables, matches by the playback, overlaps by the playback, time of day, presence of female, male age, and breeding stage. For illustration purposes, we also generated a combined approach score using principal components analysis on the four approach variables. The first principal component had an eigenvalue of 2.08 and explained 52.0% of the variation. Time in the circle and latency to retreat loaded positively (0.542 and 0.580, respectively), while latency to approach and closest approach loaded negatively (−0.275 and −0.542, respectively), so positive values of this approach score indicate rapid, close, and long-duration approach to the loudspeaker. JMP 5.0 statistical software (SAS Institute, Cary NC) was used for the multivariate analyses and residual plots. JMP generates a series of dummy variables for categorical variables with 3 or more levels (e.g., breeding stage) and considers these variables for entry into the stepwise model along with the continuous variables. JMP also produces residual leverage scatter plots that illustrate the effect size of each independent variable in the final multivariate model while controlling for other terms in the model.
To assess the possible cause and effect relationship between the singing and approach variables identified in the stepwise analysis, we employed path analysis using the graphically driven software package AMOS 7.0 (SPSS, Chicago IL; Arbuckle, 2006). The Specification Search feature allows the user to set up an initial model with causal arrows pointing in both directions between the target singing and approach variable, as well as from all independent variables to these two variables, and then systematically removes links to find the best models. In separate analyses, we declared each singing variable and its associated approach response variable as endogenous variables with arrows connecting them in both directions. All of the context variables (breeding stage, male age, time of day, pre-playback song rate, playback matches and overlaps, female presence) were included as exogenous variables, with pair-wise correlations among them and causal arrows from each one pointing to both of the endogenous variables. Breeding stage was converted into four separate variables (building, incubating, nestlings, and fledglings) coded with +1 or −1 for inclusion in the exogenous variable set. The search process reports the ten best models. As recommended, we chose the one with the lowest BIC (Bayesian Information Criterion), which penalizes model complexity and therefore tends to pick the most parsimonious models. We report the final path models graphically, showing the retained arrows and their standardized regression coefficients (β) and significance.
RESULTS
All 34 subjects sang and approached the loudspeaker in response to playback. Two males did not enter the 15 m circle but approached to 16 and 18 m, respectively. Sixteen males entered, retreated, and then re-entered the 15 m circle, usually in the post-playback period. Table 1 shows the pair-wise correlation matrix among the four approach response variables, the three key singing variables, and the subset of other continuous variables that entered one or more of the stepwise models. Table 2 summarizes the final stepwise models.
Table 1.
Correlation matrix (Pearson r values) for approach, male singing, and playback variables.
| Prop time <15m |
Closest appr |
Latency to first retreat |
Matching rate |
Overlap rate |
Versatility | Pre-pbk song rate |
Dur-pbk song rate |
Male song length |
Overlaps by playback |
Matches by playback |
Male age |
|
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Latency first appr | −0.210 | 0.149 | −0.177 | −0.260 | 0.062 | 0.064 | 0.248 | 0.197 | −0.031 | 0.040 | 0.037 | 0.209 |
| Prop time <15m | −0.418* | 0.520* | 0.323 | −0.206 | −0.011 | −0.134 | 0.023 | 0.266 | 0.065 | 0.179 | −0.240 | |
| Closest approach | −0.548** | −0.257 | 0.024 | −0.134 | 0.355* | 0.129 | 0.070 | 0.109 | −0.195 | 0.266 | ||
| Latency to retreat | 0.395* | −0.338* | 0.228 | −0.232 | −0.146 | 0.333 | −0.221 | 0.291 | −0.074 | |||
| Matching rate | 0.013 | 0.115 | −0.075 | −0.128 | 0.431* | 0.031 | 0.177 | 0.072 | ||||
| Overlapping rate | 0.096 | 0.061 | 0.177 | −0.266 | 0.140 | 0.241 | −0.279 | |||||
| Versatility | 0.467* | −0.265 | 0.156 | −0.360* | 0.634** | −0.007 | ||||||
| Pre-pbk song rate | 0.164 | 0.239 | −0.061 | 0.400* | 0.270 | |||||||
| Dur-pbk song rate | −0.181 | 0.791** | 0.088 | 0.206 | ||||||||
| Male song length | −0.116 | 0.201 | 0.134 | |||||||||
| Overlaps by playback | −0.067 | 0.104 | ||||||||||
| Matches by playback | 0.051 |
P < 0.05,
P < 0.001.
Table 2.
Final stepwise GLM models. F statistic for whole model and independent effects of each variable in the model, P-values and direction of effect.
| Dependent variable | Effects | F (df) | P | Direction |
|---|---|---|---|---|
| Matching rate | Whole model | 6.29 (6,27) | 0.0003 | |
| Latency first approach | 6.04 (1,27) | 0.043 | − | |
| Closest approach | 5.73 (1,27) | 0.024 | − | |
| Male song length | 10.04 (1,27) | 0.004 | + | |
| Male overlapping | 4.02 (1,27) | 0.055 | + | |
| Male minimum age | 4.50 (1,27) | 0.043 | + | |
| Breeding stage (B+I+F vs N+X)* | 10.13 (1,27) | 0.004 | + | |
|
| ||||
| Overlapping rate | Whole model | 5.40 (3,30) | 0.004 | |
| Latency first retreat | 9.59 (1,30) | 0.004 | − | |
| Matches by playback | 6.62 (1,30) | 0.015 | + | |
| Male minimum age | 5.11 (1,30) | 0.031 | − | |
|
| ||||
| Song-type versatility | Whole model | 13.70 (4,27) | <0.0001 | |
| Matches by playback | 7.57 (1,27) | 0.011 | + | |
| Pre-playback song rate | 7.75 (1,27) | 0.010 | + | |
| During playback song rate | 9.85 (1,27) | 0.004 | − | |
| Latency first retreat | 5.99 (1,27) | 0.021 | + | |
| Time of day | 3.92 (1,27) | 0.058 | + | |
| Breeding stage (F vs X+B+I+N)* | 16.37 (1,27) | 0.0004 | − | |
|
| ||||
| Post-pbk delayed matching rate | Whole model | 2.85 (3,27) | 0.056 | |
| Post time < 15 m | 7.15 (1,27) | 0.013 | + | |
| Male minimum age | 4.68 (1,27) | 0.040 | + | |
| Time of day | 2.17 (1,27) | 0.153 | − | |
Breeding stages: X = none, B = building, I = incubating, N = nestlings, F = fledglings
Males that matched the playback at higher rates with immediate plus delayed song-type matches approached the speaker more quickly and more closely (Fig. 1). Moreover, males that matched the playback also sang longer songs during the playback period. This song length effect occurred in part because matching males more often delivered double (compound) songs, one of which matched a playback song. Although there was no simple correlation between matching rate and overlapping rate (Table 1), after correcting for other variables in the model, males with a high matching rate also showed a tendency to overlap the playback more. To determine whether immediate and delayed matching constitute a graded signal, we separated males into three classes – those that immediately matched one or more times, those that only delay-matched, and non-matchers – and compared their approach responses. We found a significant linear increase in proportion of time spent in the 15 m circle for non-matchers, delayed matchers, and immediate matchers (linear regression with matching strategy treated as an ordinal variable: F2,32 = 3.51, P = 0.042, Fig. 2).
Figure 1.

Significant effects associated with matching rate during playback; all graphs show leverage residuals for x and y axes. (A) Matching rate as a function of latency to first approach, showing higher matching associated with more rapid approach. (B) Matching rate as a function of closest approach to the speaker, showing higher matching associated with closer approach. (C) Matching rate as a function of song length. (D) Matching rate as a function of male age, showing increased matching for older males.
Figure 2.
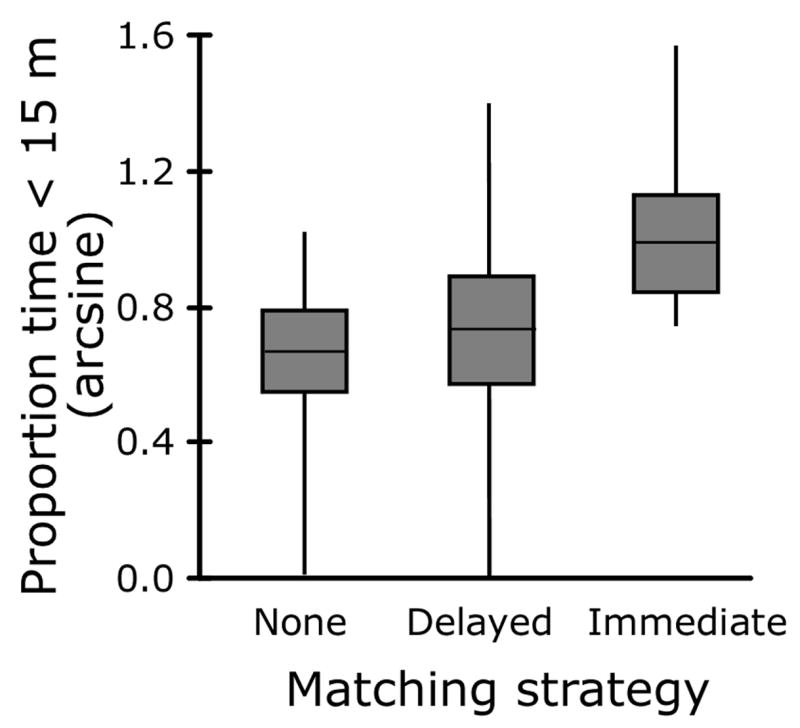
Proportion of time spent within the 15 m circle during plus post playback for males that did not match (n=10), delay-matched only (n=11) and immediately type matched (n=13). Box and whiskers plots show mean, ±SE, and range.
The path analysis using closest approach yielded no significant causal links between matching and closest approach. The equivalent analysis with approach latency retained both arrows between matching rate and latency to first approach, but only the link from matching to latency of first approach was significant (Fig. 3a). We examined the temporal relationship between matching and approach for thirteen males that immediately matched a playback song (range: 1 to 4 times). Seven of these males matched shortly before or at the time of their major approach towards the speaker, and the remaining six males matched while closest to the speaker; no males immediately matched while retreating.
Figure 3.
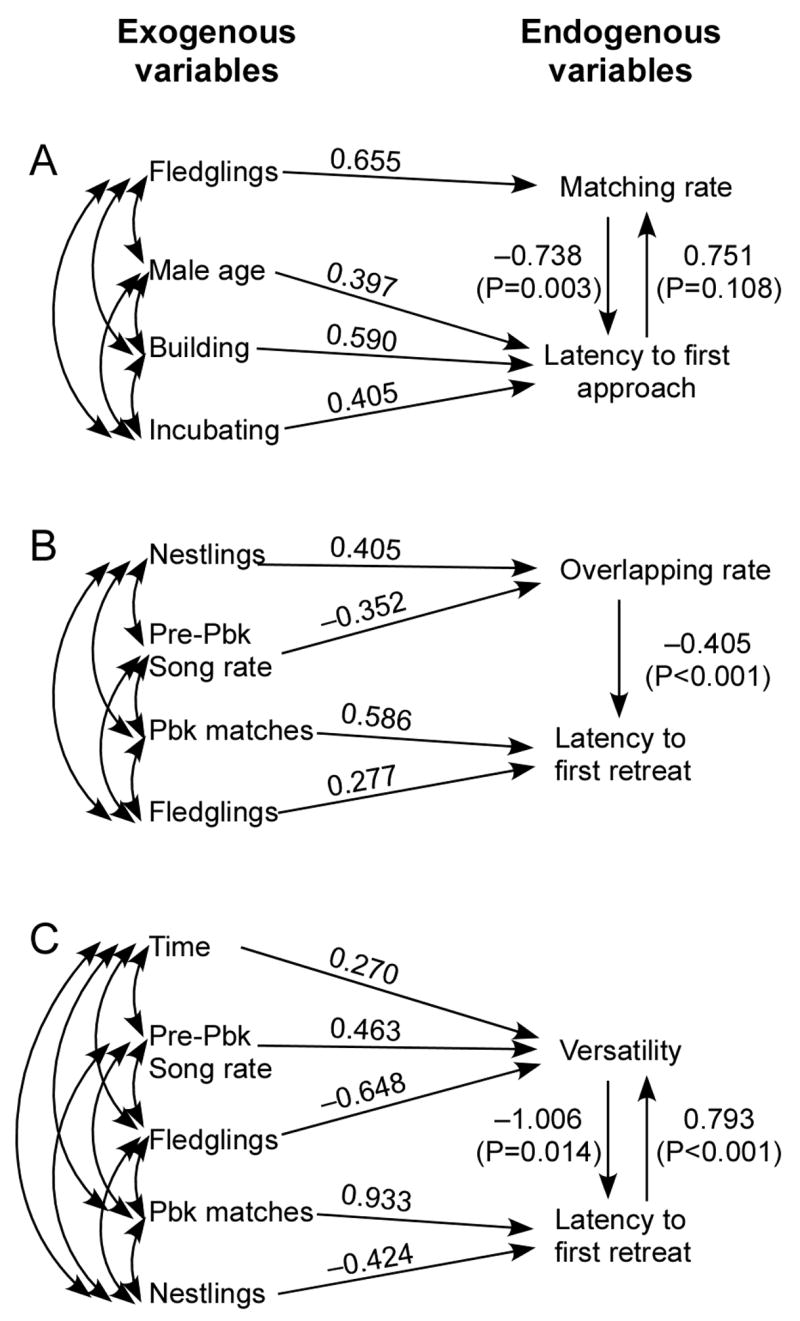
Final path models testing cause and effect relationships between singing and approach variables; initial set of exogenous variables included all context variables except other during-playback singing and response variables. (A) Matching rate versus latency to first approach. (B) Overlapping rate versus latency to first retreat. (C) Versatility versus latency to first retreat. Values on arrows are standardized regression coefficients (β).
To examine the possible role of delayed matches in the post-playback period, we performed a separate stepwise regression analysis with post-playback delayed matching rate as the dependent variable, and all of the post-playback singing measures (song rate, versatility, song length), post time spent in the circle, post closest approach, and the context variables breeding stage, time of day, and male age as initial independent variables (n = 31 because three males did not sing in the post-playback period). We found that post-playback delayed matching rate was positively associated with post time spent inside the circle (Table 2). Post-playback delayed matching rate was also strongly correlated with during-playback matching rate by the male (r = 0.518, n = 31, P = 0.003).
Males that overlapped more playback songs retreated significantly sooner from the 15 m circle (Table 2 and Fig. 4). In addition, subjects that were more frequently matched by the playback stimulus were more likely to overlap the playback. Nineteen of the 34 subjects overlapped the playback 1 to 4 times. Seven of these males overlapped before approaching, 7 overlapped while close to the speaker, and 5 overlapped while retreating. The path analyses provided a clear and consistent resolution of the causal relationship regardless of the initial set of exogenous variables, with overlap rate significantly predicting latency to first retreat but not the reverse (Fig. 3b). The primary exogenous variable causing a high rate of overlapping was the nestling stage of breeding.
Figure 4.
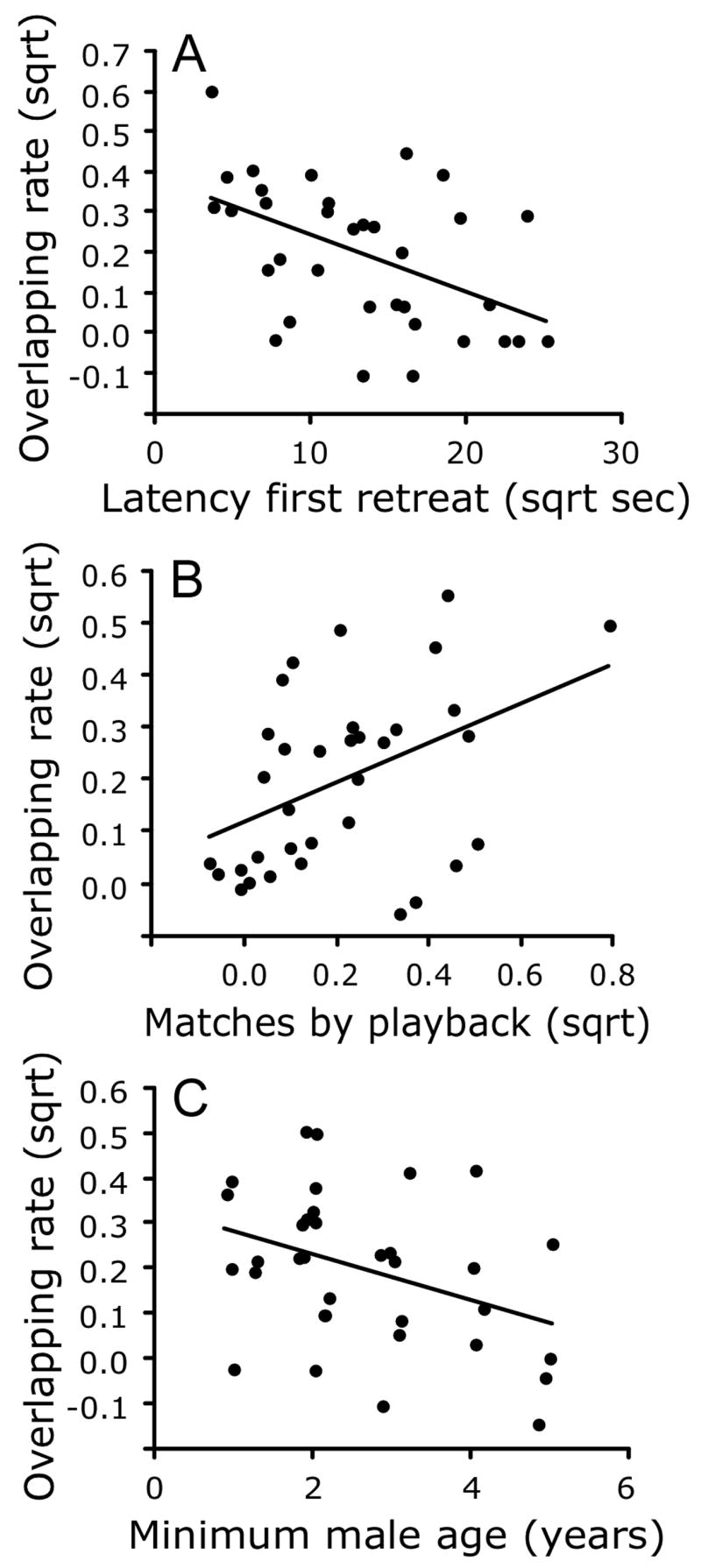
Significant effects associated with overlapping rate during playback; all graphs show leverage residuals for x and y axes. (A) Overlapping rate as a function of latency to first retreat, showing higher overlapping associated with more rapid retreat. (B) Overlapping rate as a function of delayed and immediate song-type matches of the subject by the playback, with more frequent overlapping by the subject associated with greater playback matching. (C) Overlapping rate as a function of male age, showing reduced overlapping by older males.
Males that sang with higher song-type versatility took longer to retreat out of the circle after approaching the speaker (Table 2 and Fig. 5). Males also sang with higher versatility the more frequently they were matched by the playback. High-versatility singing was associated with a higher pre-playback song rate, but a lower during-playback song rate. Singing versatility during the playback period tended to be higher for subjects tested later in the morning; a similar trend has been observed during natural singing in the 0700 to 0930 period (Trillo and Vehrencamp, unpublished data).
Figure 5.
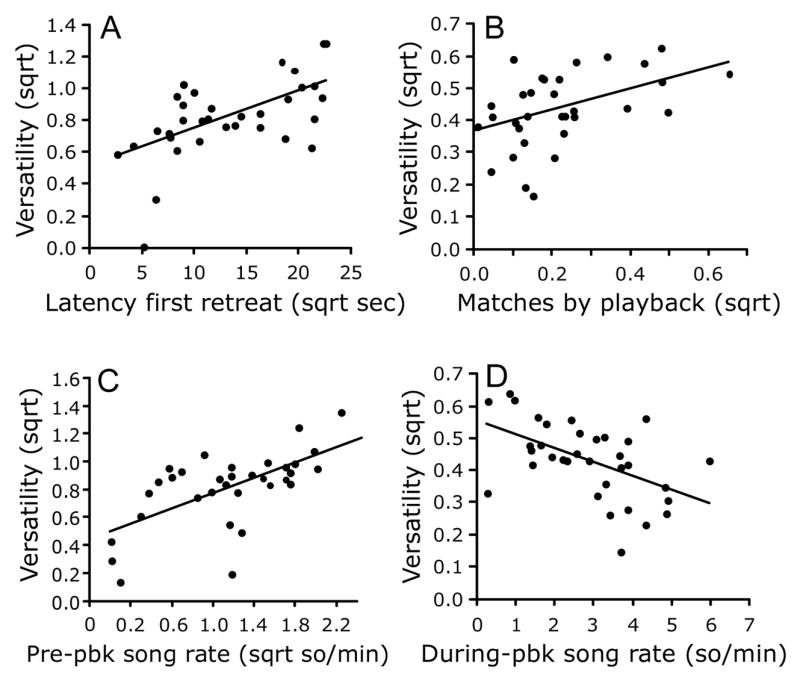
Significant effects associated with song-type versatility during playback; all graphs show leverage residuals for x and y axes. (A) Versatility versus latency to first retreat, showing higher versatility associated with slower retreat. (B) Versatility versus number of matches by playback, with higher versatility associated with greater playback matching. (C) Versatility as a function of pre-playback song rate, showing higher versatility associated with higher rate of pre-playback singing. (D) Versatility as a function of during-playback song rate, with more repetitive singing associated with higher song rate during playback.
The path analysis model for versatility retained both arrows between the singing variable and latency to first retreat, and both were significant (Fig. 3c). This result indicates that versatility both determines and is affected by the birds’ retention time within the 15 m circle. Models with different initial sets of exogenous variables retained most of the same variables, but connected them differently to versatility and latency to retreat. Thus we cannot make a definitive conclusion about causation by the exogenous variables. However, there were three strong association patterns. 1) Males with fledglings sang with low versatility and remained close to the speaker for a long time. 2) Males that were frequently matched by the playback sang with high versatility and remained close for a long time. 3) Males that were singing at a high rate before the playback started sang with high versatility during playback but retreated quickly.
The breeding stage of focal males affected their approach and singing responses in several ways. Figure 6 illustrates the simple mean values of the key singing and approach variables for different breeding stages, and Table 2 and Figure 3 include the significance of the breeding stage variables that entered the stepwise and path models, respectively. Males with fledglings sang repetitively, matched the playback at high rates, and exhibited a significantly stronger composite aggressive response score. In contrast, males with nestlings overlapped the playback most and tended to exhibit shorter retreat latency and less time spent within the circle. Males in the nest-building stage were intermediate in terms of singing variables and tended to approach the speaker more slowly and less closely. Presence or absence of the female herself during the playback experiment did not enter any stepwise or path analysis.
Figure 6.
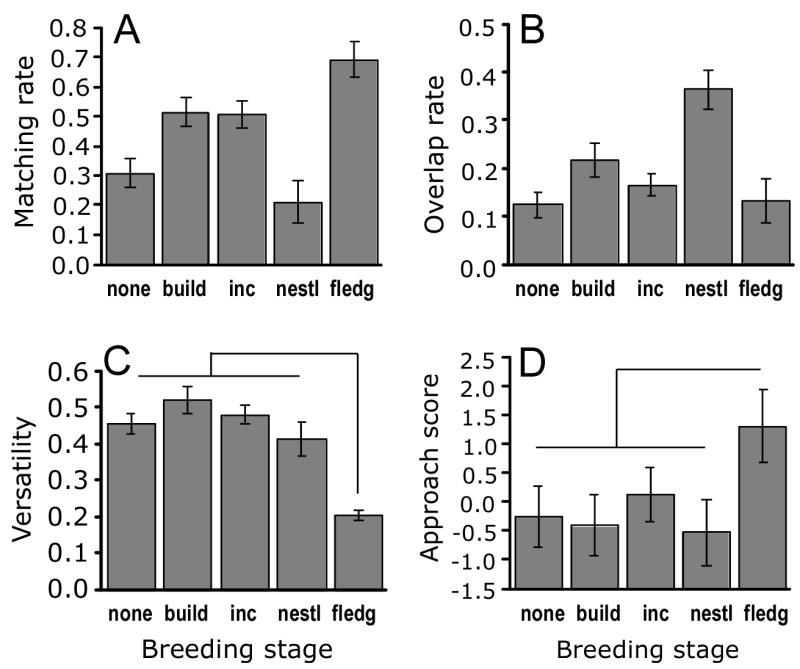
Effects of breeding stage on singing variables and approach; bars show means ±SE. Breeding stages and sample sizes are: none (no nest) n = 7; build (nest building) n = 7, inc (incubating eggs) n = 9; nestl (feeding nestlings) n = 6; fledg (feeding fledglings) n = 5. (A) Matching rate as a function of breeding stage, contrast between build-incubate-fledgling stages versus none-nestling stages significant at P = 0.004 (Table 2). (B) Overlapping rate as a function of breeding stage. (C) Versatility as a function of breeding stage, contrast between fledgling versus all other stages significant at P = 0.0004 (Table 2). (D) Approach score as a function of breeding stage, contrast between fledging versus all other stages significant at F1,29 = 5.41, P = 0.027.
Male age affected several of the singing variables. Older males were significantly more likely to match both during and after playback, and they were less likely to overlap the playback (Table 2). However, older males did not display more aggressive behavior, but tended to approach more slowly and less closely and retreat faster (Table 1).
DISCUSSION
Our study provides a sender’s perspective on the meaning of different singing patterns. Territory owners were exposed to a relatively lengthy playback of song types in their repertoires, allowing them to demonstrate how their choice and timing of songs was related to different measures of their speaker-approaching behavior. Banded wrens are particularly good subjects for such a subject-choice experiment because most individuals sing in response to songs broadcast on their territories, approach and investigate the stimulus, and vary in their tendency to remain close or leave the vicinity of the speaker. Combining this controlled singing and response information with results of previous work on natural counter-singing interactions and receiver responses to playback of alternative singing pattern treatments provides a deeper understanding of these acoustic signals. In the discussion, we show how information about sender context, receiver response, and associated sender behavior has led to our conclusions about the signal functions of matching, overlapping, and song-type versatility, respectively.
Song-type matching
Context
Song-type matching in the banded wren necessarily occurs in the context of counter-singing interactions between neighboring males and during close boundary encounters. Two illustrations of such interactions can be found in Molles and Vehrencamp (2001) and Molles (2006). Switching rapidly between song types shared by both individuals occurs at the initial stages of vocal engagement, resulting in frequent delayed matches and occasional single immediate matches. The birds do not sequentially switch and match each other; bouts of sequential matching always entail both birds singing the same song type in repeat mode. Such bouts of matched-type counter-singing occur partway into a prolonged interaction when the birds are close to each other and seem to represent an escalation. These intensive interactions are more likely to occur when neighboring males are renegotiating their territory boundary (Trillo and Vehrencamp, 2005). This study also found that males with fledglings matched more than males in other stages of breeding, and that older males matched more often than younger males.
Receiver response
Receivers exposed to an intensive interactive playback that continuously matched them approached the speaker more rapidly, relative to playback that delivered non-matching shared or unshared song types (Molles and Vehrencamp, 2001). Close approach increases the risk of being attacked, and was proposed as evidence for the required receiver retaliation cost of a conventional threat signal (Hurd, 1997; Hurd and Enquist, 1998; Vehrencamp, 2000).
Sender behavior
Males that matched the playback more often with immediate plus delayed matches approached the speaker faster and closer, compared to males that matched less often. This study found that immediate matching in particular occurred only prior to or during closest approach to a simulated intruder, never during retreat. Matching males that retreated before the playback ended overlapped the playback at that time. Immediate matchers spent more total time during and after playback close to the speaker than delay matchers, and non-matchers spent even less time close. Males that continued to delay-match the playback song types in the post-playback period also remained close to the speaker for longer than males that did not delay-match.
Function
Matching is a graded signal of short-term aggressive motivation. The strong association between immediate matching and aggressive approach in this sender-perspective study, together with the rapid approach to a matching stimulus found in the receiver-perspective study and the observations of higher levels of matching during periods of boundary change, by males defending fledglings, and by older males, are consistent with the interpretation of immediate matching as a strong offensive threat signal in the banded wren. Although the path analysis suggested that matching “predicts” rapid approach, the timing of immediate matches with respect to the birds’ movements does not support such a strong causative relationship, and we prefer to conclude only that matching is associated with a rapid and close approach response and strong aggressive motivation. Sender-perspective studies of type-matching in song sparrows and other species have sometimes found an association with aggressive approach (Vehrencamp 2001, Burt et al. 2001), although Searcy et al. (2006) found only a trend for type-matching and a stronger association between soft song and actual attack behavior.
Delay-matching rate in the post-playback period was correlated with time spent close to the speaker and to during-playback matching rate, suggesting that similar motivational factors may drive both during- and post-playback matching behavior. Delayed matching seems to be a moderately agonistic signal that indicates willingness to engage at close range and continue in a vocal counter-singing interaction. A similar argument was made for delayed matching in the nightingale (Luscinia megarhynchos), although here the delay ranged from a fraction to a few seconds between song starts of two rivals and resulted in graded levels of overlapping and song alternation (Todt and Naguib, 2000). Our scoring method for delayed matches in the banded wren counted delays up to 5 min. The fact that the percentage of delayed matches was correlated with measures of approach indicated that these relatively long intervals with other intervening song types are nevertheless meaningful. In this study, some males failed to sing any of the six playback song types even though they possessed all of these types in their repertoires (approximately 25% random expectation of matching). These birds did not approach the speaker as quickly or closely or remain close for as long as the matchers. Avoidance of matching therefore signals less aggressive or de-escalating motivation. During interactions among song-type sharing neighbors in both banded wrens and song sparrows, singing a non-shared song type both prevents the neighbor from matching and is a clear non-matching signal associated with the lowest likelihood of aggression (Beecher and Campbell, 2005; Molles and Vehrencamp, 2001).
Song overlapping
Context
Some overlapping clearly occurs by chance. Overlapping rates are higher when song rates are higher as in the dawn chorus (Burt and Vehrencamp, 2005). Similarly, in the current study males that sang at higher rates were more often overlapped by the playback by chance (Table 1). Preliminary analyses of microphone array recordings indicate that overlapping is more prevalent when singing males are either close to each other or far away, compared to intermediate distances (Vehrencamp and Burt, unpublished data). Overlapping type matches sometimes occur during counter-singing interactions, and on occasion one male will start a song and the other will finish it or overlap the terminal trill. Other relevant aspects of context found in this study include higher overlapping by younger males, by males provisioning nestlings, and by males that were more often matched by playback.
Receiver response
Hall et al. (2006) found that receivers responded in an aversive manner to being repeatedly overlapped, shortening their songs and tending to stay further from the speaker. There was a significant effect of treatment order in that study (which employed a preliminary lure design), with birds that received the overlapping treatment first singing shorter songs and approaching less closely in the alternating treatment, compared to birds that received the alternating treatment first. In receiver-perspective studies on other species, overlapped birds have been described as “uncomfortable”, “disturbed”, “agitated”, or “aversive” (Todt and Naguib 2000; Mennill and Radcliffe 2004; Hall et al. 2006).
Sender behavior
In the Hall et al. (2006) overlapping experiment, 15% of the subjects overlapped the playback, and those birds showed a tendency to spend less time close to the speaker compared to males that did not overlap. The current study strongly corroborated this result because males that overlapped playback at high rates left the vicinity of the speaker sooner. The path analysis showed that overlapping predicted retreat. This conclusion is consistent with our observation that it occurred before and during retreat away from the speaker. Overlapping rate was not in any way associated with latency to approach or closeness of approach.
Function
Most research on song overlapping has focused on the perspective of the receiver (the bird being overlapped and eavesdroppers), and little is known of the context and sender perspective. Although previous receiver-response experiments with overlapping playback have argued that it is a highly effective threat signal that repels rivals (Todt and Naguib, 2000), or a rival-directed signal of elevated sender arousal (Dabelsteen et al., 1996, 1997), the current sender-perspective study suggests a very different interpretation. Overlapping masks or jams the song of the opponent, in particular the terminal trill, which in banded wrens is the loudest song component and may carry information about male quality (Illes et al., 2006). By masking this information, the overlapper may be signaling that he is “not interested” in listening to the rival’s full song or in interacting further (Todt and Naguib, 2000). Because overlapping both holds receivers at a distance and predicts sender withdrawal in the banded wren, it appears to function as an intention-to-retreat signal or as a “defensive” (i.e., opposite of “offensive”) threat. Our results are consistent with those of Langemann et al. (2000), who found an association between overlapping and singing further from the speaker in their study of the great tit (Parus major).
Song-type versatility
Context
In contrast to the prior two vocal signals, singing with different song-type versatility patterns can occur in the absence of a counter-singing rival. Banded wrens can vary song-type switching rate and type diversity somewhat independently, but the two strategies are highly correlated: a very low switching rate, i.e., completely repetitive singing, is necessarily associated with very low diversity. Molles (2006) thought the two strategies might send different messages, but could not specify the separate functions. We found it more useful to combine them into a single index of song-type versatility, since the birds vary switching rate and diversity along a continuous scale.
The extremes of completely repetitive singing and high-switch-high-diversity singing certainly occur in different contexts. The contexts for repetitive singing include: 1) when solo singing, such as the first male to start the dawn chorus, and mid-morning solo bouts from either the center or edge of the territory; 2) during the escalation stage of a boundary dispute when neighboring males are close to each other; and 3) after a close encounter, chase, or fight by the first bird to start singing (Molles and Vehrencamp, 1999; Trillo and Vehrencamp, 2005; Molles, 2006). The current study revealed a very strong effect of breeding stage context on the subjects’ use of song-type versatility. Males with fledglings sang repetitively compared to males in all other stages, and they also responded aggressively to the playback. Males with recently fledged young react vigorously and defensively to our presence on the territory at this time as well. The strong response to a simulated invader may occur because food resources on the territory are most limited at this stage, or because juveniles are directly threatened by enemies of all types. In another Thryothorus wren, Logue and Gammon (2004) also found that males with juveniles approached playback more closely than males without juveniles.
Contexts for high-versatility singing include: 1) the dawn chorus, 2) unmated males in the hour after dawn chorus, and 3) short courtship bouts of soft singing to receptive mates (Molles and Vehrencamp 1999; Burt and Vehrencamp 2005; M. Hall, A. Illes, S. Vehrencamp, personal observations). An intermediate level of versatility, with high rates of switching among a subset of 2–4 song types, occurs when males are counter-singing with a neighbor from a distance (Molles, 2006). During these interactions, a male preferentially uses song types shared with the neighbor, and the opponent may either switch among the same subset to produce frequent delayed matching, or may switch among a different subset of shared song types (Trillo and Vehrencamp, 2005; Molles, 2006). Most of the time counter-singing rivals alternate songs rather than overlap, suggesting that they are paying attention to the other’s song type choice.
Receiver response
Molles (2006) presented territorial males with three alternative versatility treatments, using songs recorded from a neighbor and broadcast from the territorial boundary with that neighbor. Receivers spent significantly more time close to the speaker during the repetitive treatment than during the two rapid-switching treatments (that differed in number of types delivered). Moreover, their singing response tended to parallel the song versatility of the playback, with higher playback versatility causing higher subject versatility and more delayed matching. One interpretation of this result is that repetitive singing is a stronger aggressive threat signal than versatile singing.
Sender behavior
The current study was consistent with this interpretation in some respects, but not in others, and the dichotomy was reflected in the path analysis. Males with fledglings sang remarkably repetitively and approached the speaker aggressively, including remaining close for a long time, compared to males in other stages of breeding. Males that had been singing at a high rate prior to the playback sang with high versatility and retreated quickly. These two patterns are consistent with the receiver-response experiment. On the other hand, males that were more frequently matched by the playback sang with very high versatility but also remained close to the speaker for a long time. This result seems to contradict the receiver experiment. In the current study, matches by playback were more frequent when males first matched the playback and stayed on the same type so that the repetitive playback matched them back. The very high positive correlation between versatility and matches by playback suggests that some males became strongly engaged in a reciprocal matching and switching vocal interaction with the playback, and remained close to the speaker for a long time.
Function
Low-versatility (repetitive) singing appears to be the default non-interactive mode of solo singing. Switching among several to many song types commences immediately when another bird starts to sing nearby, and initiates a bout of vocal interaction with variable levels of switching and matching. High-versatility singing seems to indicate that a male will not immediately approach very closely or attack and that the interaction can continue for some period of time. We speculate that these extended vocal interactions allow rivals to negotiate or learn something about each other based on rates of immediate and delayed matching, tendencies to follow versus lead, and/or performance quality of different song types. When a counter-singing male then shifts into repetitive singing, he may be signaling that he is no longer “interested” in learning or negotiating, while continuing to sing and not backing down. Repetitive singing is probably not an aggressive threat signal per se, but rather a non-interactive mode of counter-singing.
This interpretation of the function of repetitive singing could reconcile the apparent contradiction between the two playback experiments. Neighbors already know each other well, so repetitive singing from the boundary may be interpreted as an unfriendly challenge that elicits a strong approach response from the receiver-owner. The simulated territorial invasion by a “local stranger” in the current study caused many of the subjects to remain moderately close and sing interactively, possibly to learn more about the unknown bird. The five subjects with fledglings did not attempt to negotiate with the stranger, but instead sang vigorously and repetitively for a long period of time. Further progress on understanding the switching and matching rules of negotiation can only be obtained from analysis of natural counter-singing bouts recorded with microphone arrays, where distance between males and their movements can be accurately documented and the fine details of song structure can be examined.
In conclusion, an on-territory playback experiment designed to allow the sender to choose a variety of singing patterns and then demonstrate its associated spatial response, coupled with careful measurements of both approach and retreat behaviors, reveals the sender’s perspective of signal message. In conjunction with information on receiver responses and social contexts, we can achieve a more complete understanding of the function of signals. We argue that on-territory playback experiments are a valid technique for inferring signal function, so long as both receiver and sender perspectives are obtained.
Acknowledgments
We thank the staff of the Area Conservación de Guanacaste, especially Roger Blanco and Maria Marta Chavarria, for logistical support of our field research in Parque Nacional Santa Rosa. This research was funded by the National Institute of Mental Health (grant R01-MH60461 to S.L.V.).
References
- Andersson M. Why are there so many threat displays? J Theor Bio. 1980;186:773–781. doi: 10.1016/0022-5193(80)90310-0. [DOI] [PubMed] [Google Scholar]
- Arbuckle JL. Amos 7.0 User’s Guide. Spring House, PA: Amos Development Corp; 2006. [Google Scholar]
- Archer J. The Behavioral Biology of Aggression. Cambridge: Cambridge University Press; 1988. [Google Scholar]
- Beecher MD, Campbell SE. The role of unshared songs in singing interactions between neighbouring song sparrows. Anim Behav. 2005;70:1297–1304. [Google Scholar]
- Bradbury JW, Vehrencamp SL. Principles of Animal Communication. Sunderland MA: Sinauer Associates; 1998. [Google Scholar]
- Burt JM, Vehrencamp SL. Dawn chorus as an interactive communication network. In: McGregor PK, editor. Animal Communication Networks. Cambridge: Cambridge University Press; 2005. pp. 320–343. [Google Scholar]
- Burt JM, Campbell SE, Beecher MD. Song type matching as threat: a test using interactive playback. Anim Behav. 2001;62:1163–1170. [Google Scholar]
- Collins S. Vocal fighting and flirting: the functions of birdsong. In: Marler P, Slabbekoorn H, editors. Nature’s Music: the Science of Birdsong. London: Elsevier Academic Press; 2004. pp. 39–79. [Google Scholar]
- Dabelsteen T, McGregor PK, Holland J, Tobias JA, Pedersen SB. The signal function of overlapping singing in male robins. Anim Behav. 1997;53:249–256. [Google Scholar]
- Dabelsteen T, McGregor PK, Shepherd M, Whittaker X, Pedersen SB. Is the signal value of overlapping different from that of alternating during matched singing in great tits. J Avian Biol. 1996;27:189–194. [Google Scholar]
- Hall MH, Illes AE, Vehrencamp SL. Overlapping signals in banded wrens: long-term effects of prior experience on males and females. Behav Ecol. 2006;17:260–269. doi: 10.1093/beheco/arj022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hardouin LA, Reby D, Bavoux C, Burneleau G, Bretagnolle V. Communication of male quality in owl hoots. Am Nat. 2007;169:552–562. doi: 10.1086/512136. [DOI] [PubMed] [Google Scholar]
- Hurd PL. Is signalling of fighting ability costlier for weaker individuals? J Theor Biol. 1997;184:83–88. [Google Scholar]
- Hurd PL, Enquist M. Conventional signalling in aggressive interactions: the importance of temporal structure. J Theor Biol. 1998;192:197–211. [Google Scholar]
- Illes AE, Hall ML, Vehrencamp SL. Vocal performance influences male receiver response in the banded wren. Proc R Soc Lond B. 2006;273:1907–1912. doi: 10.1098/rspb.2006.3535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Järvi T, Radesäter T, Jakobsson S. The song of the willow warbler Phylloscopus trochilus with special reference to singing behavior in agonistic situations. Ornis Scand. 1980;11:236–242. [Google Scholar]
- Kilner RM. The evolution of complex begging displays. In: Wright J, Leonard ML, editors. The Evolution of Begging: Competition, Cooperation and Communication. Dordrecht: Kluwer Academic Publishers; 2002. pp. 87–106. [Google Scholar]
- Kramer HG, Lemon RE, Morris MJ. Song switching and agonistic stimulation in the song sparrow (Melospiza melodia): five tests. Anim Behav. 1985;33:135–149. [Google Scholar]
- Krebs JR, Ashcroft R, van Orsdol K. Song matching in the great tit, Parus major L. Anim Behav. 1981;29:918–923. [Google Scholar]
- Langemann U, Tavares JP, Peake TM, McGregor PK. Response of great tits to escalating patterns of playback. Behaviour. 2000;137:451–471. [Google Scholar]
- Leitao A, Riebel K. Are good ornaments bad armaments? Male chaffinch perception of songs with varying flourish length. Anim Behav. 2003;66:161–167. [Google Scholar]
- Logue DM, Gammon DE. Duet song and sex roles during territory defence in a tropical bird, the black-bellied wren, Thryothorus fasciatoventris. Anim Behav. 2004;68:721–731. [Google Scholar]
- Mennill DJ, Ratcliffe LM. Overlapping and matching in the song contests of black-capped chickadees. Anim Behav. 2004;67:441–450. [Google Scholar]
- Molles LE. Singing complexity of the banded wren Thryothorus pleurostictus: Do switching and song-type diversity send different messages? Auk. 2006;123:991–1003. [Google Scholar]
- Molles LE, Vehrencamp SL. Repertoire size, repertoire overlap, and singing modes in the banded wren (Thryothorus pleurostictus) Auk. 1999;116:677–689. [Google Scholar]
- Molles LE, Vehrencamp SL. Songbird cheaters pay a retaliation cost: evidence for auditory conventional signals. Proc R Soc Lond B. 2001;268:2013–2019. doi: 10.1098/rspb.2001.1757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nielsen BMB, Vehrencamp SL. Responses of song sparrows to song-type matching via interactive playback. Behav Ecol Sociobiol. 1995;37:109–117. [Google Scholar]
- Osiejuk TS, Ratynska K, Cygan JP. Signal value of alternating and overlapping singing in the Yellowhammer Emberiza citrinella. J Ethol. 2004;22:55–61. [Google Scholar]
- Otte D. Effects and functions in the evolution of signaling systems. Ann Rev Ecol Syst. 1974;5:385–417. [Google Scholar]
- Searcy WA, Nowicki S. Male-male competition and female choice in the evolution of vocal signaling. In: Espmark Y, Amundsen T, Rosenqvist G, editors. Animal Signals: Signalling and Signal Design in Animal Communication. Trondheim: Tapir Academic Press; 2000. pp. 301–315. [Google Scholar]
- Searcy WA, Nowicki S. The evolution of animal communication: reliability and deception in signaling systems. Princeton: Princeton University Press; 2005. [Google Scholar]
- Searcy WA, Anderson RC, Nowicki S. Bird song as a signal of aggressive intent. Behav Ecol Sociobiol. 2006;60:234–241. [Google Scholar]
- Seyfarth RM, Cheney DL, Marler P. Monkey responses to three different alarm calls: Evidence of predator classification and semantic communication. Science. 1980;210:801–803. doi: 10.1126/science.7433999. [DOI] [PubMed] [Google Scholar]
- Simpson BS. Effects of location in territory and distance from neighbours on the use of song repertoires by Carolina wrens. Anim Behav. 1985;33:793–804. [Google Scholar]
- Slabbekoorn H, tenCate C. Stronger territorial responses to frequency modulated coos in collared doves. Anim Behav. 1997;54:955–965. doi: 10.1006/anbe.1997.0499. [DOI] [PubMed] [Google Scholar]
- Smith WJ, Smith AM. Information about behaviour provided by Louisiana waterthrush Serius motacilla (Parulinae) songs. Anim Behav. 1996;51:785–799. [Google Scholar]
- Todt D, Naguib M. Vocal interactions in birds: The use of song as a model in communication. Adv Study Behav. 2000;29:247–296. [Google Scholar]
- Trillo PA, Vehrencamp SL. Song types and their structural features are associated with specific contexts in the banded wren. Anim Behav. 2005;70:921–935. doi: 10.1016/j.anbehav.2005.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vehrencamp SL. Handicap, index, and conventional signal elements of bird song. In: Espmark Y, Amundsen T, Rosenqvist G, editors. Animal signals: signalling and signal design in animal communication. Trondheim: Tapir Academic Press; 2000. pp. 277–300. [Google Scholar]
- Vehrencamp SL. Is song-type matching a conventional signal of aggressive intentions? Proc R Soc Lond B. 2001;268:1637–1642. doi: 10.1098/rspb.2001.1714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Waas JR. Do little blue penguins signal their intentions during aggressive interactions with strangers? Anim Behav. 1991;41:375–382. [Google Scholar]
- Weary DM, Krebs JR, Eddyshaw R, McGregor PK, Horn A. Decline in song output by great tits: Exhaustion or motivation? Anim Behav. 1988;36:1241–1244. [Google Scholar]