

Abstract
We investigated the role of the hyperpolarization-activated mixed cation current, IH, in the control of spontaneous action potential firing of rat cerebellar Purkinje neurons in brain slices. Extracellular recordings revealed that the continual action potential firing of Purkinje neurons was disrupted by the pharmacological blockade of IH. Blockade of IH revealed spontaneous transitions between periods of tonic action potential firing and quiescence, without effects on the frequency or variance of action potential generation. Whole-cell recordings revealed that blockade of IH unmasked a form of membrane potential bistability, where transitions between tonic firing and quiescent states (separated by ∼20 mV) were evoked by excitatory and inhibitory postsynaptic potentials, or by the delivery of brief (20 ms) somatic or dendritic positive and negative current pulses. The stable upper state of tonic action potential firing was maintained by the recruitment of axo-somatic voltage-activated sodium, but not calcium, channels. Negative modulation of IH by serotonin unmasked bistability, indicating that bistability of Purkinje neurons is likely to occur under physiological conditions. These data indicate that IH acts as a ‘safety net’, maintaining the membrane potential of Purkinje neurons within the range necessary for the generation of tonic action potential firing. Following the downregulation of IH, synaptic inhibition can generate long periods (seconds) of quiescence, the duration of which can be controlled by climbing fibre activation and by the underlying ‘tone’ of parallel fibre activity.
Purkinje neurons are the sole output neurons of the cerebellum. In vivo they fire action potentials tonically at high rates (Granit & Phillips, 1956; Armstrong & Rawson, 1979; Stratton et al. 1988), despite the presence of high levels of spontaneous inhibitory synaptic activity (Eccles et al. 1966b; Häusser & Clark, 1997). Superimposed upon this tonic firing are complex bursts of action potentials generated by the periodic activation of the climbing fibres originating from the inferior olive (Eccles et al. 1966a; Lang et al. 1999), as well as increases in firing rate caused by activity in granule cells, the origin of the parallel fibres (Eccles et al. 1966a; Armstrong & Rawson, 1979). The energetically costly nature of tonic action potential firing in Purkinje neurons suggests an important role in the function of the cerebellar system.
The frequency of tonic action potential firing in Purkinje neurons in vivo has been shown to be maintained in brain slice preparations in vitro where synaptic inputs have been blocked (Häusser & Clark, 1997) and following the acute disassociation of Purkinje neuron somata (Nam & Hockberger, 1997; Raman & Bean, 1999). This demonstrates that tonic action potential firing in cerebellar Purkinje neurons is generated by intrinsic membrane conductances, with a recent study implicating voltage-activated sodium channels as the main conductance involved in generating the inward current necessary to maintain tonic action potential firing (Raman & Bean, 1999).
Observations from other spontaneously active central neurons indicate that the hyperpolarization-activated mixed cationic current IH plays a crucial role in the control of the periodicity of both the tonic and burst firing of action potentials (McCormick & Pape, 1990; Maccaferri & McBain, 1996; Williams et al. 1997; Ghamari-Langroudi & Bourque, 2000). As Purkinje neurons express IH (Crepel & Penit-Soria, 1986; Roth & Häusser, 2001), it may be expected to influence tonic action potential firing in these neurons. In acutely dissociated Purkinje neurons, however, previous findings have indicated that IH contributes little inward current at membrane potentials near the action potential threshold, and that the pharmacological blockade of IH does not alter the frequency of tonic action potential firing (Raman & Bean, 1999). This suggests that IH is not directly involved in tonic action potential firing in acutely dissociated Purkinje neurons. The situation in more intact preparations may be different, however, due to the influence of ongoing inhibitory synaptic activity (Eccles et al. 1966b; Häusser & Clark, 1997) and dendritically located ion channels (Llinas & Sugimori, 1980a, b; Stuart & Häusser, 1994).
Here, we explore the role of IH in the control of tonic action potential firing in Purkinje neurons maintained in brain slice preparations. Our observations reveal that IH acts as a ‘safety net’, holding the membrane potential within the range necessary for activation of the inward current required for tonic action potential firing. Removal of this ‘safety net’ following pharmacological blockade of IH or downregulation by serotonin leads to the appearance of membrane potential bistability. As a consequence, excitatory and inhibitory synaptic inputs can switch the Purkinje neurons between quiescent and tonically active states.
METHODS
Experiments were performed according to methods approved by the Animal Experimentation Ethics Committee of the Australian National University and the UK Home Office. Wistar rats (postnatal days 18–25) were anaesthetized by inhalation of halothane, decapitated, and 300-μm-thick sagittal brain slices of the cerebellar vermis or somatosensory cortex were prepared using standard procedures. Slices were perfused with oxygenated Ringer solution of composition (mm): NaCl 125; NaHCO3 25; KCl 3; NaH2PO4 1.25; CaCl2 2; MgCl2 1 and glucose 25. Somatic cell-attached and whole-cell voltage- and current-clamp recordings (pipette resistance 3–6 MΩ), or simultaneous somatic and dendritic (pipette resistance 8–15 MΩ) whole-cell current-clamp recordings, were made from visually identified cerebellar Purkinje or neocortical layer 5 pyramidal neurons, as described previously (Stuart et al. 1993). For somatic cell-attached voltage-clamp recordings, the holding potential was set to 0 mV. During whole-cell voltage-clamp experiments, a series of depolarizing and hyperpolarizing voltage steps (5 mV increments, 1.5 s duration, 0.1 Hz) were delivered from a holding potential of −45 mV using ≥ 95 % series resistance compensation in the presence of lag values ≤ 10 μs. Cell-attached and current-clamp experiments were performed at 32–33 °C, while voltage-clamp experiments were made at room temperature (20–24 °C). Patch electrodes were filled with (mm): potassium gluconate 135; NaCl 7; Hepes 10; EGTA 0.5; Na2-ATP 2; Na2-GTP 0.3; disodium phosphocreatine 10; MgCl2 2 (pH 7.2 adjusted with KOH; osmolarity 270 mosmol l−1). In some experiments, 0.5 mm EGTA was replaced by 20 mm BAPTA, and potassium gluconate lowered to maintain osmolarity. No correction was made for liquid junction potentials. For current-clamp recordings, voltage signals were filtered at 10–30 kHz, while for cell-attached and voltage-clamp recordings current signals were filtered at 2–5 kHz. Voltage and current signals were acquired at 50–100 kHz, using an ITC-16 or ITC-18 interface (Instrutech, USA) controlled by an Apple Power Macintosh G3 running Axograph (Axon Instruments, USA) acquisition and analysis software. ‘Steady-state’ activation curves determined from tail-current analysis were fitted with a single Boltzmann equation of the form: y = 1/(1 + e(V1/2-V)/k) where V1/2 is the voltage of half-maximal activation and k is a constant.
Synaptic potentials were evoked electrically using an isolated stimulator (200 μs pulse, ≤ 100 μA) connected to a patch pipette (4–10 MΩ) filled with extracellular solution. Climbing fibres were activated by stimulating in the granule cell layer at low frequency (0.1–0.2 Hz) in the presence of the GABAA receptor antagonist bicuculline methiodide (20 μm). Inhibitory postsynaptic potentials (IPSPs) were evoked selectively by stimulation in the molecular layer, either with ‘off-beam’ stimulation (≥ 250 μm from the Purkinje cell soma) or in the presence of the excitatory amino-acid receptor antagonists 4-hydroxyquinoline-2-carboxylic acid (Kynurenic acid; 3 mm) and 6-cyano-7-nitroquinoxaline-2,3-dione (CNQX; 10 μm). Tetrodotoxin (TTX; 0.1 μm) dissolved in extracellular solution was applied, where indicated, locally by pressure application. The application pipette (similar to that used for somatic whole-cell recording) was placed either close (∼10 μm) to the axon, 10–40 μm from the soma in the granule cell layer, or within the dendritic arbor (120–140 μm from the soma) under visual guidance and positive pressure applied (150–200 mmHg), resulting in a spatially localized ejection area (Williams & Stuart, 1999). Numerical values are given in the text as mean ± s.e.m. Statistical analysis was performed with Student's t test, and statistical significance was determined with α = 0.05.
RESULTS
Action potential firing is independent of dI/dt in Purkinje neurons
Purkinje neurons fire action potentials spontaneously at high rates in the absence of synaptic inputs. To explore the mechanisms controlling this spontaneous firing, we made whole-cell current-clamp recordings from the somata of Purkinje neurons and evoked action potential firing with slow ramps of positive current (0.5–2 nA s−1, 2 s) delivered from a hyperpolarized membrane potential (−70.9 ± 1.5 mV; n = 7). These ramps generated a gradual membrane potential depolarization that abruptly triggered high-frequency action potential firing, typically in the range of 50–70 Hz (Fig. 1A, B, top). Importantly, the initial action potential firing rate (first inter-spike interval) was only very weakly dependent upon the slope of the current ramp (dI/dt; correlation coefficient 0.32 ± 0.10; n = 7; Fig. 1C, top), indicating that Purkinje neurons adopt an initial firing rate that is, broadly, independent of the rate of membrane potential depolarization. In contrast, action potential firing in layer 5 neocortical pyramidal neurons evoked with similar ramps did not exhibit an abrupt transition to high-frequency action potential firing (Fig. 1A, B, bottom), with the initial firing frequency increasing as a smooth function of the slope of current ramps (n = 4; Fig. 1C, bottom). These results reveal that under control conditions Purkinje neurons display hallmarks of a bistable system, where the virtual independence of the initial action potential firing rate from dI/dt indicates that action potential firing is a quantitatively stable state with a high minimum value.
Figure 1. Action potential firing is independent of dI/dt in Purkinje neurons.
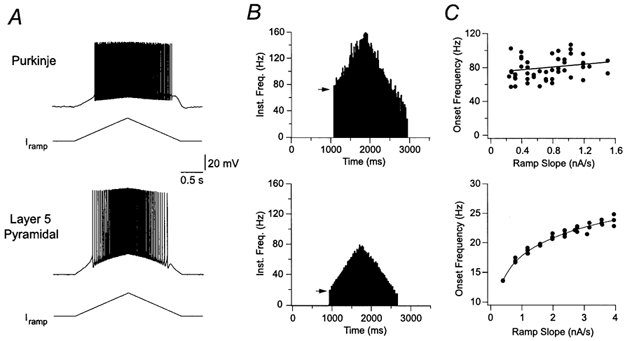
A, somatic whole-cell recording of action potential firing elicited by slow (2.5 s) ramp currents in a Purkinje neuron (top panel; ramp peak 0.9 nA), and a layer 5 pyramidal neuron (bottom panel; ramp peak 2.0 nA). Prior to the ramp the Purkinje neuron was hyperpolarized to −65 mV by the injection of a negative holding current, whereas the pyramidal neuron was resting at −65 mV. B, plot of the instantaneous firing frequency during the ramp currents shown in A. Note that the onset frequency (indicated by an arrow) is very different for the Purkinje and pyramidal neurons. C, relationship between the slope of injected current and the onset frequency for different ramps. Lines represent linear (top) or polynomial (bottom) fits.
Blockade of IH reveals state transitions of spontaneous firing
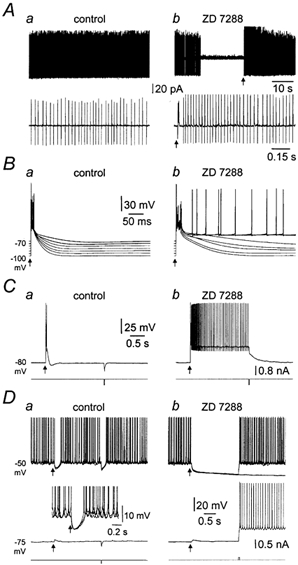
During somatic cell-attached patch-clamp recordings, Purkinje neurons fired action potentials spontaneously with a mean frequency of 41.8 ± 7.4 Hz and coefficient of variation (CV) of 0.36 ± 0.12 (n = 15; Fig. 2A, B), as described previously (Häusser & Clark, 1997). Bath application of ZD 7288 (10 μm), a selective and irreversible pharmacological blocker of IH (BoSmith et al. 1993; Harris & Constanti, 1995; Roth & Häusser, 2001), led to the appearance of long periods of quiescence (≥ 5 s) where no spontaneous action potential firing occurred (Fig. 2A). These quiescent periods were terminated by the abrupt reappearance of sustained tonic action potential firing, with a mean frequency and CV that were not significantly different from control values (Fig. 2B).
Figure 2. Blockade of IH transforms tonic action potential firing in Purkinje neurons.
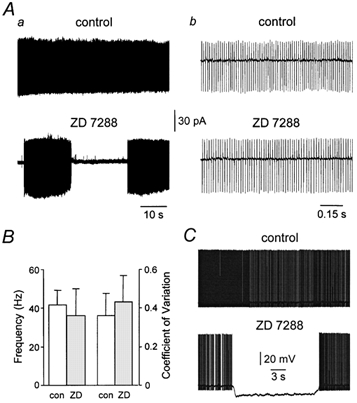
Aa, somatic cell-attached (Vpipette= 0 mV) recording of spontaneous action potential firing under control condtions and in the presence of ZD 7288 (10 μm). The periodicity of action potential firing is shown more clearly on an exapanded time base (b). B, summary data of the mean frequency and coefficient of variation of spontaneous action potential firing under control (□) and in the presence of ZD 7288 ( ). C, somatic whole-cell current-clamp recording of spontaneous action potential firing under control conditions and in the presence of ZD 7288 (10 μm). Note the sudden spontaneous hyperpolarization and cessation of action potential firing.
). C, somatic whole-cell current-clamp recording of spontaneous action potential firing under control conditions and in the presence of ZD 7288 (10 μm). Note the sudden spontaneous hyperpolarization and cessation of action potential firing.
We further investigated this phenomenon using somatic whole-cell current-clamp recordings. During whole-cell recording under control conditions, Purkinje neurons fired action potentials spontaneously and tonically in a manner identical to that observed during extracellular recording (Fig. 2C). Application of ZD 7288 led to the appearance of long quiescent periods, which were associated with spontaneous membrane potential hyperpolarizations of up to −25 mV (Fig. 2C; −16.6 ± 1.1 mV; n = 15). Quiescence was terminated by spontaneous membrane depolarization and the restoration of tonic action potential firing (Fig. 2C). These data indicate that following pharmacological blockade of IH, Purkinje neurons fluctuate spontaneously between two qualitatively different states, tonic action potential firing and quiescence at a hyperpolarized membrane potential.
Properties of IH in Purkinje neurons
To better understand the role of IH in the control of tonic action potential firing we characterized its electrophysiological properties in Purkinje neurons using somatic whole-cell current- and voltage-clamp recording techniques. As shown previously (Crepel & Penit-Soria, 1986; Roth & Häusser, 2001), Purkinje neurons responded to hyperpolarizing current pulses (from a holding potential of −64 ± 0.5 mV; n = 22) with a slow depolarizing sag back to the holding potential (Fig. 3Aa). The amplitude of this sag (calculated by subtracting the steady-state potential measured just before the offset of negative current steps from the peak potential) increased in a linear manner with the magnitude of peak voltage deflections (Fig. 3Ab; n = 22). Depolarizing current steps evoked voltage deviations that were characterized by the appearance of a slow transient depolarizing potential at their onset (Fig. 3Aa). Transient hyperpolarizing and depolarizing potentials were generated at the offset of negative and positive current steps, respectively (Fig. 3Aa). The application of ZD 7288 abolished the depolarizing sag generated during negative current steps and led to a substantial membrane potential hyperpolarization (−16.0 ± 1 mV; n = 20) and an increase in apparent input resistance (Fig. 3B). This increase in apparent input resistance was found to be significantly greater following the restoration of the membrane potential to control levels by the injection of tonic current through the recording pipette (Fig. 3B). Furthermore, transient hyperpolarizing and depolarizing potentials evoked at the offset of positive and negative current steps, respectively, were abolished by ZD 7288, indicating that they are mediated exclusively by IH.
Figure 3. Physiological and pharmacological properties of IH in Purkinje neurons.
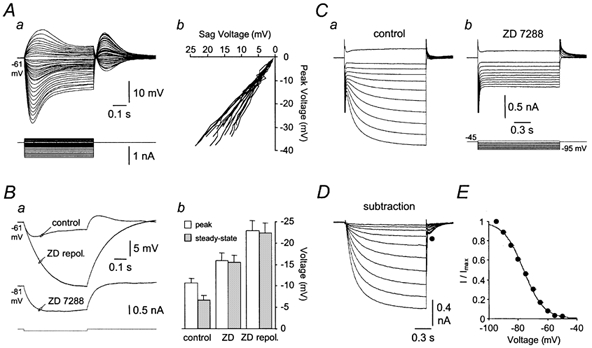
Aa, voltage responses (upper traces) to positive and negative current steps (lower traces) injected via the recording pipette during whole-cell current-clamp recording. Note the appearance of a slow depolarizing sag during the course of the hyperpolarizing voltage responses. The holding potential was set to −61 mV. Ab, the amplitude of the sag during negative current steps is linearly related to the peak amplitude of hyperpolarizing voltage responses, like those shown in Aa. This relationship is shown for 22 neurons. Ba, hyperpolarizing voltage responses evoked by −100 pA current steps (bottom) recorded under control conditions and in the presence of ZD 7288 (10 μm). Note that ZD 7288 hyperpolarized the membrane potential by 20 mV. Restoration of the membrane potential to control levels by positive current injection revealed a dramatic increase in apparent input resistance and blockade of the depolarizing sag. Bb, summary data of the peak and steady-state amplitude of hyperpolarizing voltage responses generated by −100 pA steps recorded at −64 ± 0.5 mV under control conditions and in the presence of ZD 7288. C, a family of current responses recorded under whole-cell voltage-clamp evoked by a series of depolarizing and hyperpolarizing voltage steps (lower traces in Cb) in control conditions (Ca) and following the application of ZD 7288 (10 μm) (Cb). D, the result of digital subtraction of traces recorded in ZD 7288 from those recorded under control conditions. Note the presence of a slow inward current with voltage-dependent kinetics. E, ‘steady-state’ activation curve obtained by tail-current analysis of digitally subtracted traces like those shown in D (mean ± s.e.m.; n = 6). The continuous line is the fit with a single Boltzmann function.
To further explore the properties of IH we made somatic whole-cell voltage-clamp recordings in the presence of the sodium channel blocker tetrodotoxin (TTX; 0.5 μm). Under these conditions, negative voltage steps delivered from a holding potential of −45 mV activated slow inward currents (Fig. 3Ca), which were abolished by the application of ZD 7288 (Fig. 3Cb; n = 6). Digital subtraction of families of currents recorded in control conditions and in the presence of ZD 7288 revealed the presence of a slow ZD 7288-sensitive inward current that increased in magnitude and possessed kinetics that accelerated with membrane negativity (Fig. 3D). Tail-current analysis of digitally subtracted currents was used to construct the ‘steady-state’ activation curve for the ZD 7288-sensitive component of the whole-cell current. The voltage dependence of activation was well described by a single Boltzmann function, with the current first activating at approximately −55 mV and a half-maximal point of −76.4 mV (Fig. 3E; n = 6). These data demonstrate that IH is active at membrane potentials near action potential threshold in Purkinje neurons, and that its pharmacological blockade leads to a dramatic increase in apparent input resistance and a substantial hyperpolarization of the membrane potential.
Modulation of intrinsic bistability by IH
The spontaneous transitions between periods of tonic firing and quiescence observed in the presence of ZD 7288 suggest that these states represent stable states within a bistable system. To examine this possibility we injected small (0.46 ± 0.03 nA), short (20 ms) pulses of depolarizing current from hyperpolarized membrane potentials under control conditions and in the presence of ZD 7288, in the continual presence of a GABAA receptor antagonist to block spontaneous inhibitory postsynaptic activity. Under control conditions, brief depolarizing current steps generated electrotonic potentials crowned by the firing of one to two action potentials that repolarized rapidly to the pre-pulse membrane potential (Fig. 4Aa). In the presence of ZD 7288, however, the same depolarizing current steps delivered from the same membrane potential evoked tonic action potential firing (Fig. 4Ab; n = 33). Tonic firing under these conditions could be stopped by the delivery of a small, brief negative current step, following which the membrane potential was reset to the pre-pulse membrane potential (Fig. 4Ab). If a negative current step was not applied, spontaneous firing could proceed for long periods (Fig. 4B). These data demonstrate that pharmacological blockade of IH reveals two clearly distinguishable stable membrane potential states in Purkinje neurons; the upper state corresponding to tonic action potential firing and the lower state corresponding to quiescence at a hyperpolarized membrane potential (−70.5 ± 0.7 mV; n = 33). The membrane potential in the upper state was measured during inter-spike intervals at times following the action potential after-hyperpolarization, yielding a voltage separation between upper and lower stable states averaging 18.4 ± 0.9 mV (n = 33).
Figure 4. Properties of intrinsic bistability.
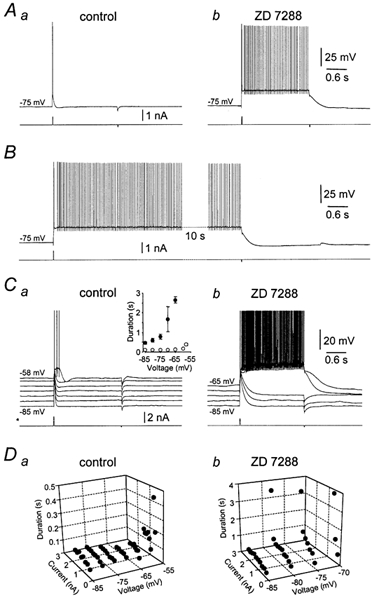
A, voltage responses (upper traces) evoked by brief positive current pulses (20 ms, 0.7 nA) under control conditions (a) and in the presence of ZD 7288 (10 μm) (b) during whole-cell current-clamp recording. Note in the presence of ZD 7288 that the positive-current pulse triggers a long period of action potential firing that could be terminated by the delivery of a brief negative-current pulse (20 ms, 0.2 nA). The holding potential was set to −75 mV. B, in the presence of ZD 7288, tonic action potential firing evoked by a brief positive-current pulse (20 ms, 0.8 nA) could continue for a long period until stopped by a brief negative-current pulse (20 ms, 0.2 nA). Note the 10 s gap in the record. C, voltage dependence of the pattern of voltage responses evoked by just-threshold positive-current pulses, recorded in control conditions (a) and in the presence of ZD 7288 (10 μm) (b). The graph (inset) shows summary data of the duration of voltage responses evoked by this protocol for six neurons recorded under control conditions (○) and in ZD 7288 (•), see Results for details. D, graphs demonstrating the voltage dependence of the relationship between the duration of voltage responses evoked by positive-current steps of different magnitude, recorded under control conditions (a) and in the presence of ZD 7288 (b; 10 μm).
To examine further the voltage dependence of this form of intrinsic bistability, we injected pulses of positive current of sufficient magnitude to evoke action potential firing from different membrane potential levels under control conditions and in the presence of ZD 7288 (Fig. 4C). Under control conditions we observed that just-threshold positive current pulses generated voltage deflections that were crowned by action potential firing that rapidly repolarized to pre-pulse values across a wide membrane potential range (Fig. 4Ca; n = 6). At the most depolarized levels examined (∼-60 mV), just-threshold positive current pulses evoked short-duration plateau potentials that led to the firing of 2–6 action potentials (Fig. 4Ca). These plateau potentials were terminated by a sudden and spontaneous repolarization and undershoot of the membrane potential, as described previously (Llinas & Sugimori, 1980a, b). In the presence of ZD 7288, positive current pulses of the same amplitude delivered over the same range of membrane potentials produced a strikingly different pattern of activity. From the most hyperpolarized membrane potentials examined, just-threshold positive current pulses evoked significantly longer voltage perturbations than in the control condition (Fig. 4Cb). As the membrane potential was made more depolarized, the duration of voltage perturbations increased slightly, until at membrane potentials between −70 and −65 mV, positive current pulses were capable of switching the neurons to the upper stable state where tonic action potential firing occurred (Fig. 4Cb).
This behaviour was summarized over a group of neurons (n = 6) by measuring the duration of voltage responses evoked by brief positive current pulses under control conditions and in ZD 7288 (Fig. 4Ca; inset). The duration of voltage responses was calculated as the time from positive current pulse onset until the membrane potential spontaneously repolarized to the pre-pulse value, or was terminated by the delivery of a negative current step. The voltage dependence of these state transitions was explored further by analysing the duration of voltage responses evoked by positive current pulses of different magnitude delivered over a wide range of membrane potentials. Under control conditions the duration of voltage perturbations did not show a strong dependence on the magnitude of current pulses over a broad range of membrane potentials, only increasing near action potential firing threshold, where small plateau potentials were observed (Fig. 4Da). In ZD 7288, however, we observed that transition from lower to upper stable states could occur over a wide range of membrane potentials, with the magnitude of positive current pulses required to produce state transition being greater at more hyperpolarized membrane potentials (Fig. 4Db). Importantly, we observed that from a membrane potential depolarized to ∼−70 mV neurons did not settle at the lower stable state, but spontaneously transitioned to the upper tonic action potential firing state. These data indicate that a region of membrane potential instability exists over a potential range between −70 mV and action potential firing threshold, as would be predicted in a bistable system.
Synaptic control of bistability
The properties of intrinsic bistability revealed by brief depolarizing and hyperpolarizing current steps suggests that similar state transitions may be triggered by excitatory and inhibitory synaptic inputs. To determine if this is the case, we first examined whether climbing fibre activation could restart tonic action potential firing during quiescent periods observed during somatic cell-attached recordings in the presence of ZD 7288. As predicted from the effect of depolarizing current pulses, during periods of quiescence, activation of the climbing fibre input, which triggered a burst of action potentials (Eccles et al. 1966a), was followed by sustained periods of tonic action potential firing (Fig. 5A; n = 5). Next, we examined the voltage dependence of this effect of climbing fibre activation under control conditions and following the pharmacological blockade of IH. Under control conditions, climbing fibre activation generated a discrete and decremental burst of action potentials when evoked over a wide range of membrane potentials (Fig. 5Ba). The application of ZD 7288 increased dramatically the duration of the climbing fibre response over the entire membrane potential range examined, and when activated from membrane potentials depolarized to −70 mV evoked a burst of action potentials that was followed by tonic action potential firing (Fig. 5Bb). This effect is more apparent in Fig. 5C, where the discrete nature of the climbing fibre burst under control conditions was transformed following the blockade of IH, such that climbing fibre activation led to a transition from the lower state to the stable upper action potential firing state (Fig. 5Cb; n = 10). Similar transitions from quiescence to tonic action potential firing could be achieved by parallel fibre stimulation (n = 4), or by the injection of somatic current waveforms designed to mimic the time course of excitatory postsynaptic potentials (EPSPs; τrise 0.3 ms; τdecay 3 ms; amplitude 1.1 ± 0.1 nA; n = 5; data not shown).
Figure 5. Synaptic control of intrinsic bistability.
A, somatic cell-attached recording of spontaneous action potential firing under control conditions (a) and in the presence of ZD 7288 (10 μm) (b). In the presence of ZD 7288 activation of a single climbing fibre (arrow) was capable of restarting tonic action potential firing (b). The pattern of action potential firing is shown on an expanded time base in the lower traces. Note the burst discharge of action potentials evoked by climbing fibre activation (Ab lower traces, arrow). B, voltage dependence of climbing fibre EPSPs recorded under control conditions (a) and in the presence of ZD 7288 (b). Note EPSPs generated from −75 and −70 mV (thick lines) evoke a long train of action potential firing in the presence of ZD 7288. C, climbing fibre EPSPs evoked under control conditions (a) and in the presence of ZD 7288 (b) are shown on a longer time base. Note that in the presence of ZD 7288 the climbing fibre EPSP leads to a long period of action potential firing that could be terminated by the delivery of a negative current pulse. D, IPSP inhibition of action potential firing under control conditions (a) and in the presence of ZD 7288 (b; three superposed traces in each panel). Note that under control conditions single IPSPs pause action potential firing (inset) and are reversed in polarity at hyperpolarized membrane potentials (holding potential shown by the side of traces). In the presence of ZD 7288, IPSP amplitude and duration are massively amplified and are capable of the complete annihilation of spontaneous firing, which could be restarted by the delivery of a small pulse of positive current.
We next examined whether IPSPs were able to generate transitions from the upper to the lower stable state in ZD 7288. Under control conditions, IPSPs were capable of pausing spontaneous action potential firing (Fig. 5Da). Following blockade of IH, however, IPSPs led to a transition from the upper tonic action potential firing state to the lower quiescent state (Fig. 5Db; n = 5), indicating a massive amplification of both the amplitude (control: −4.6 ± 1.1 mV; ZD 7288: −13.8 ± 2.4 mV) and duration of IPSPs. These data indicate that following blockade of IH, single EPSPs and IPSPs are capable of switching the membrane potential of Purkinje neurons between the upper and lower stable states.
Spread of bistable states in the dendritic tree
Simultaneous somatic and dendritic whole-cell recordings (42–115 μm from the soma; Fig. 6) were used to examine the spread of bistability in Purkinje neurons. Transitions between lower and upper states could be achieved by brief (20 ms) pulses of positive current delivered at either dendritic (Fig. 6Aa) or somatic (Fig. 6Ab) locations. The magnitude of current pulses required to generate transitions to the upper stable state in ZD 7288 was similar during dendritic and somatic current injections (soma: 0.32 ± 0.043 nA; dendrite 42–115 μm from the soma: 0.36 ± 0.063 nA; n = 9). We also found that the voltage difference between lower and upper stability points was slightly smaller at more distal dendritic locations (Fig. 6B), as a consequence of the more depolarized upper stability point at the soma (−50.5 ± 1.6 mV compared to −52.4 ± 1.6 mV; n = 9). Although these differences were not statistically significant, they suggest that the inward currents responsible for the generation of the upper stable state are located close to the soma.
Figure 6. Bistable states are efficiently spread throughout the dendritic tree.
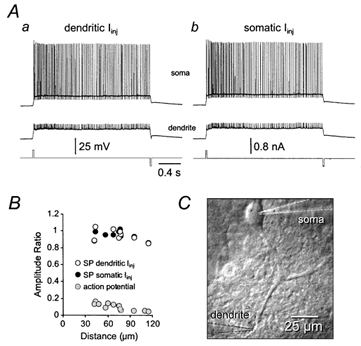
A, bistable voltage responses in the presence of ZD 7288 (10 μm) initiated and terminated by 20 ms pulses of positive and negative current, respectively (lower traces), delivered through the dendritic (a) or somatic (b) recording pipette during simultaneous whole-cell recording. Note the large decrement of action potential amplitude from soma to dendrites, but the lack of attenuation of stable bistability points. Dendritic recording was made 68 μm from the soma. B, summary data demonstrating the spatial profile of attenuation of action potentials ( ) and the voltage difference between stability points (SP) following initiation of bistability at dendritic (○) or somatic (•) sites. C, the image shows the morphology of a Purkinje neuron and placement of the recording pipettes.
) and the voltage difference between stability points (SP) following initiation of bistability at dendritic (○) or somatic (•) sites. C, the image shows the morphology of a Purkinje neuron and placement of the recording pipettes.
Ionic basis of bistability
Previous studies have suggested that voltage-activated sodium and calcium channels play a role in the formation of plateau potentials in Purkinje neurons (Llinas & Sugimori, 1980a, b; Callaway & Ross, 1997). To investigate the role of sodium channels in bistability we used the sodium channel blocker TTX. Bath application of TTX (0.2–0.5 μm) blocked intrinsic bistability in a time-dependent manner (Fig. 7A; n = 6). Following TTX application, the duration and integral of voltage responses evoked by positive current steps in ZD 7288 decreased progressively until both action potentials and bistability were blocked (Fig. 7A). Analysis of the time course of blockade of bistability and action potentials by TTX revealed that bistability was fully blocked at times preceding any statistically significant reduction in action potential amplitude (Fig. 7B, C). To investigate the cellular location of the sodium channels responsible for the formation of bistability we applied TTX locally (0.1 μm in a pipette) to axonal and dendritic sites (Fig. 7D, E). Prolonged (10–90 s) pressure application of TTX to dendritic sites did not alter bistability (voltage separation between stability points, control: 22 ± 2 mV, TTX 22 ± 2 mV; integral, control: 46 ± 3 mV s, TTX: 47 ± 4 mV s; n = 5). In the same experiments, however, brief (3–6 s) axonal application of TTX reversibly abolished bistable behaviour (voltage separation between stability points, control: 22 ± 2 mV, TTX 0.9 ± 0.2 mV, wash: 25 ± 3 mV; integral, control: 46 ± 3 mV s, TTX: 5 ± 1 mV s, wash: 48 ± 3 mV s; n = 5). These data indicate that bistability is a consequence of the recruitment of axo-somatic sodium channels.
Figure 7. Bistable states are generated by the recruitment of voltage-activated sodium channels.
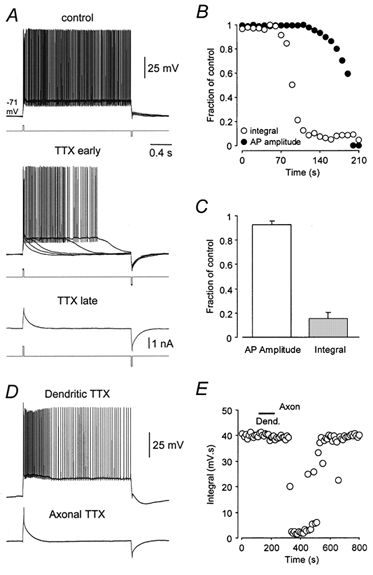
A, bistable voltage responses recorded in the presence of ZD 7288 (10 μm) evoked by brief positive-current pulses and terminated by brief negative-current pulses (upper five overlaid traces). The bath application of TTX (0.2 μm) resulted in a gradual reduction of bistable behaviour (middle traces, five overlaid traces), until both bistability and action potentials were blocked (lower trace). The holding potential was −71 mV. B, time course of blockade of bistable behaviour by TTX measured as the integral of voltage responses over a 2 s period following initiation by positive-current pulses (○), and action potential (AP) amplitude (•). Values expressed as a fraction of the control. C, summary data (n = 6) showing that bistable responses () are greatly reduced at time points before any significant effect of TTX on the amplitude of action potentials is noted (□). D, bistable voltage responses are insensitive to local dendritic TTX (0.1 μm; 90 s; 120 μm from the soma), but are blocked reversibly by local axonal TTX application (3 s, 20 μm from the soma). E, time series showing the integral of voltage responses during dendritic and axonal TTX application, same neurons as in D.
To examine the involvement of calcium channels, we bath-applied the broad-spectrum calcium channel antagonist cadmium (200 μm; n = 5). Rather than decreasing bistability, application of cadmium enhanced bistability, statistically significantly increasing the integral of voltage responses and the voltage separation between the lower and upper stability points (Fig. 8A, B). This led to the progressive inactivation of somatic action potentials following transitions to the upper stable state (Fig. 8A). In the same experiments the efficacy of cadmium to block calcium channels was verified by the total blockade of climbing fibre responses, and thus the bistable behaviour they generated (Fig. 8C). To explore the possible role of calcium channel subtypes, we applied the specific P/Q-type calcium channel antagonist ω-agatoxin GVIA (0.5–1.0 μm; Dove et al. 1998). In common with the effects of cadmium, blockade of P/Q-type calcium channels did not reduce, but rather facilitated bistable behaviour, increasing the voltage separation between stability points and the integral of voltage responses (Fig. 8B; n = 5). These data indicate that the upper stable state of tonic action potential firing is generated as a consequence of the recruitment of voltage-activated sodium, but not calcium channels.
Figure 8. Bistability is not dependent upon the activation of calcium channels.
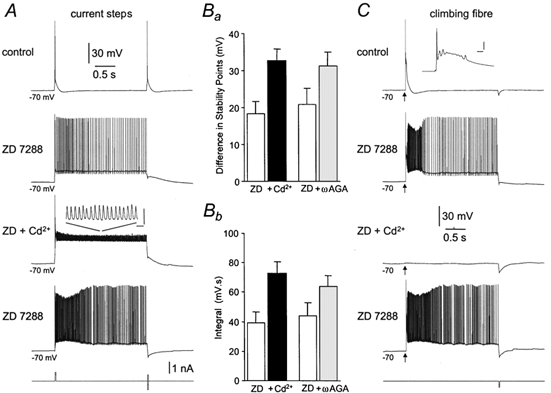
A, bistability recorded in the presence of ZD 7288 (10 μm) evoked by brief pulses of positive current and terminated by brief pulses of negative current are augmented, in a reversible manner, by the application of cadmium (200 μm). Note that in the presence of cadmium, the amplitude of somatic action potentials are smaller, presumably due to inactivation (inset calibration: 10 ms; 10 mV). B, summary of the effects of calcium channel antagonists on the voltage separation between upper and lower bistability points (a) and the integral of bistable responses (b): □, control; ▪, cadmium (200 μm; n = 5); , ω-agatoxin GVIA (0.5–1 μm; n = 5). C, in the same experiment, climbing fibre evoked activity is completely and reversibly blocked by cadmium.
To determine the role of calcium-activated potassium channels in the control of bistability, we dialysed Purkinje neurons with 20 mm of the rapid calcium chelator BAPTA by including it in the pipette internal solution. Intracellular dialysis of a Purkinje neurons with BAPTA-containing solution rapidly increased the spontaneous firing frequency (Fig. 9A). Within 15 s of establishing the whole-cell configuration, the spontaneous firing rate rose from 48.7 ± 12.3 Hz (measured in the cell-attached configuration just before break in) to 100.0 ± 14.2 Hz (n = 5; Fig. 9A). Continued dialysis of the neurons with BAPTA in the pipette internal for several minutes converted the spontaneous tonic firing pattern into a bistable pattern where prolonged periods of silence alternated with high-frequency spikes that rapidly inactivated (Fig. 9B). The membrane potential during the silent period during dialysis with BAPTA averaged −61.3 ± 1.4 mV (n = 10), a value considerably depolarized compared to that of the lower state observed during application of ZD 7288. Furthermore, the initial firing frequency in the upper state was substantially higher than seen in control conditions or during ZD 7288 application, averaging 344.1 ± 45.8 Hz (n = 10). Inactivation of these spikes led to a depolarized plateau potential with an average membrane potential of −29.8 ± 0.95 mV. When dialysis with BAPTA internal solution was combined with bath application of ZD 7288 (40 μm), spontaneous bistability was also observed (Fig. 9B), with the onset frequency being 118.7 ± 18.6 Hz (n = 4), the depolarized plateau potential following sodium spike inactivation averaging −29.6 ± 0.95 mV, and the membrane potential during the silent period averaging −93.8 ± 1.38 mV. These results indicate that calcium-activated potassium channels help to regulate the transition between tonic firing and bistability, and influence the voltage level of the upper and lower states.
Figure 9. Spontaneous bistability can be enhanced by blockade of calcium-activated potassium channels.
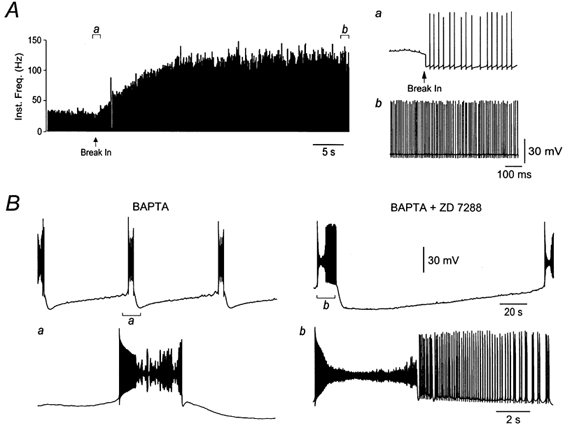
A, internal BAPTA rapidly increased the spontaneous firing frequency of Purkinje neurons. A continuous histogram of action potential firing rate demonstrates the increase in frequency following establishment of a whole-cell recording using an intra-pipette solution containing 20 mm BAPA (see Methods). Voltage traces (right) show action potentials in the cell-attached and whole-cell mode (a; firing rate 55 Hz) and 15 s following break-in (b, firing rate 140 Hz). B, in another Purkinje neuron, continued dialysis with BAPTA led to a spontaneous bistable activity pattern (left). The inset (a) shows state transitions at a faster time base. Addition of ZD 7288 (40 μm) enhanced the spontaneous bistable activity pattern (right); stereotyped bursts of firing were interspersed with periods of quiescence lasting several minutes. The inset (b) shows state transitions on a faster time base.
Neuromodulation of IH reveals bistability
IH is a common site of neuromodulation and is known to be regulated by the neurotransmitters of the ascending reticular activating system (Pape, 1996). In Purkinje neurons, activation of adenergic and sertonergic receptors have been demonstrated to negatively modulate IH (Crepel et al. 1987; Li et al. 1993). To explore whether intrinsic bistability emerges as a consequence of neuromodulation of IH we investigated the effects of serotonin (100 μm, n = 6) on bistability. Bath application of serotonin generated a small membrane potential hyperpolarization (−3 ± 0.5 mV) and decreased the sag in the response to long negative current steps by 18.2 ± 3.8 % (Fig. 10A) across the entire negative range of the voltage-current relationship (Fig. 10B). In the presence of serotonin, the bistability triggered by brief current pulses was enhanced (Fig. 10C), such that the integral of voltage responses compared to control conditions was dramatically increased (Fig. 10D), similar to observations following the blockade of IH with ZD 7288 (compare with Fig. 4C). The voltage separation between the lower and upper stable states in the presence of serotonin was significantly smaller (10.2 ± 0.9 mV) than observed in the presence of ZD 7288, consistent with the incomplete block of IH by serotonin.
Figure 10. Serotonin unmasks intrinsic bistability through a negative modulation of IH.
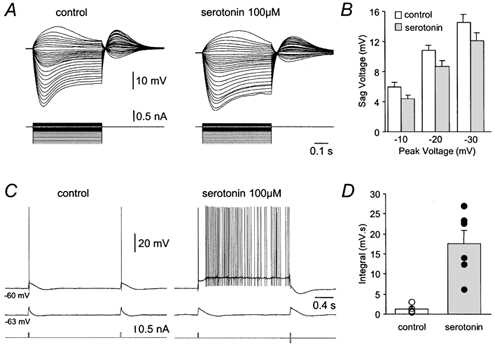
A, families of voltage responses (upper traces) evoked by a series of positive and negative current steps (lower traces) recorded under control conditions and in the presence of serotonin (100 μm). Note the serotonin-induced reduction of depolarizing sag potentials generated during the course of hyperpolarizing voltage responses. B, summary of the amplitude of the sag generated during hyperpolarizing voltage responses to the indicated peak amplitudes under control conditions (□) and in the presence of serotonin () (n = 6). C, voltage responses (upper traces) evoked by brief pulses of positive current (lower traces) under control conditions and in the presence of serotonin (100 μm) at two different holding potentials (shown to the left of voltage traces). Note that in the presence of serotonin, voltage responses evoked from −60 mV exhibit bistable behaviour. D, summary of the integral of voltage responses recorded under control conditions (□ and ○) and in the presence of serotonin (and •), evoked by the protocol shown in C (n = 6).
We next explored if state transitions could also be unmasked by serotonin in response to synaptic inhibition (‘off-beam’ inhibition, 10 stimuli at 100 Hz repeated every second). Under control conditions, bursts of IPSPs transiently paused spontaneous firing (Fig. 11A). In the presence of serotonin, however, bursts of inhibitory synaptic inputs triggered prolonged pauses in Purkinje cell spontaneous firing, occasionally leading to the complete cessation of firing during the period between IPSP bursts (Fig. 11B; n = 5). As with ZD 7288, the action potential firing rate was not statistically significantly altered by serotonin, indicating that its primary effect was to enhance bistability rather than affecting the action potential firing rate per se. Taken together, these data indicate that negative neuromodulation of IH by serotonin can act to enhance membrane potential bistability in Purkinje neurons.
Figure 11. Serotonin enhances intrinsic bistability triggered by inhibitory synaptic input.
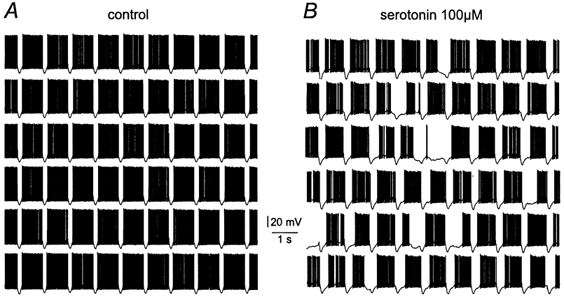
A, spontaneous action potential firing under control conditions. Brief gaps in spontaneous firing were generated by rhythmic (1 Hz) activation of off-beam inhibitory synaptic fibres in a burst pattern (10 stimuli at 100 Hz). B, in the presence of 100 μm serotonin, the same rhythmic (1 Hz) activation of off-beam inhibitory synaptic fibres generated long pauses in spontaneous action potential firing, consistent with the enhancement of bistability by partial blockade of IH.
DISCUSSION
The pioneering studies by Llinás and colleagues demonstrated the existence of dendritic electrogenesis in Purkinje neurons and the formation of long-lasting plateau potentials mediated, in part, by the recruitment of sodium currents (Llinas & Sugimori, 1980a, b). More recently it has been shown that Purkinje neurons are not quiescent, but intrinsically generate action potentials at high rates in the absence of synaptic inputs (Häusser & Clark, 1997; Nam & Hockberger, 1997; Raman & Bean, 1999), with voltage-activated sodium channels implicated as the main inward current necessary for the maintenance of tonic action potential firing (Raman & Bean, 1999). Our observations are consistent with these findings, but extend them by demonstrating that IH plays a critical role in the maintenance of excitability in cerebellar Purkinje neurons.
Our results indicate that blockade of IH leads to spontaneous periods of quiescence that can last for tens of seconds, the duration of which can be controlled by synaptic input. In contrast to previous findings made from spontaneously active central neurons (McCormick & Pape, 1990; Maccaferri & McBain, 1996; Williams et al. 1997; Ghamari-Langroudi & Bourque, 2000), we find that IH does not control the frequency of action potential firing itself (Raman & Bean, 1999). Rather, IH maintains the membrane potential within a range where the inward sodium current responsible for the generation of spontaneous action potential firing in Purkinje neurons operates. In this way IH acts as a ‘safety net’ for tonic action potential firing, ensuring that tonic firing is maintained even when challenged by a high level of spontaneous inhibitory postsynaptic inputs (Eccles et al. 1966b; Häusser & Clark, 1997). In support of this hypothesis, we have demonstrated that IPSPs could annihilate tonic action potential firing for long periods following pharmacological blockade of IH. This behaviour was shown to result from the enhancement of membrane potential bistability, which is characterized by the existence of a hyperpolarized lower stable state (more negative than −70 mV) and an upper stable state associated with tonic action potential firing. These observations demonstrate both a novel biophysical property of Purkinje neurons and the critical role played by IH in the control of their excitability.
Ionic mechanisms underlying bistability
The properties of sodium currents in Purkinje neurons are unique, possessing both a resurgent component enabling partial recovery from inactivation at depolarized membrane potentials, and a persistent component (Gahwiler & Llano, 1989; Raman & Bean, 1997, 1999; Kay et al. 1998). Both these components of the sodium current have been proposed to play a critical role in the generation of spontaneous action potential firing by supplying inward current during inter-spike intervals (Raman & Bean, 1999). The persistent component of the sodium current (INap) has been shown to be available for activation from membrane potentials more positive than ∼70 mV, a level that is approximately 20 mV more negative than the action potential threshold in Purkinje neurons (Llinas & Sugimori, 1980b; Gahwiler & Llano, 1989; Nam & Hockberger, 1997; Kay et al. 1998; Raman & Bean, 1999). In support of the role of INap in tonic action potential firing, we find that bistability is abolished by the blockade of sodium currents by TTX prior to any effect on action potentials. Presumably the difference in the TTX sensitivity of INap and action potentials (Azouz et al. 1996; Stuart, 1999) occurs as a consequence of the large reserve of sodium channels available for action potential generation (Mainen et al. 1995; Rapp et al. 1996; Madeja, 2000). Furthermore, we show that axo-somatic sodium channels are responsible for the formation of bistability, a finding that supports the suggestion of a high density of axonal sodium channels (Mainen et al. 1995; Rapp et al. 1996).
Our data indicate that other ionic currents also play a role in the control of bistability in Purkinje neurons. Blockade of calcium currents enhanced bistability by depolarizing the upper stability state. Non-selective blockade of calcium-dependent mechanisms with internal BAPTA also enhanced bistability. This indicates, consistent with previous work (Raman & Bean, 1999), that the net effect of calcium currents on spontaneous firing is not to provide inward current, but rather to activate calcium-activated mechanisms, presumably potassium currents, which play a key role in regulating the excitability of the Purkinje neuron. As a consequence, the blockade of calcium-activated currents with BAPTA depolarized the upper stability point and could trigger spontaneous bistable firing. In the presence of BAPTA, unlike other results, the upper stable state was marked by action potential inactivation and exhibited hallmarks of calcium electrogenesis.
We therefore suggest that the upper stable state is maintained primarily by INap, which is available for activation from membrane potentials positive to ∼70 mV. Depolarizing voltage perturbations that enter this membrane potential region activate this persistent inward current and so give rise to a tonic membrane potential depolarization that initiates spontaneous action potential firing as an upper stability point. The rate of tonic action potential firing is, however, controlled by the activation of calcium- and voltage-dependent potassium currents that operate to prevent runaway depolarization and subsequent inactivation of sodium action potentials (see Fig. 9). Under control conditions, IH acts to maintain the membrane potential within this range, thereby helping to sustain tonic action potential firing. Following the blockade or downregulation of IH, however, membrane potential hyperpolarizations caused by IPSPs, or the after-hyperpolarizations that follow climbing fibre evoked complex spikes or intense activity, can switch Purkinje neurons into a lower stable state, generating periods of quiescence that can last for many seconds.
Our findings are in contrast with previous modelling studies that have demonstrated a critical role of P/Q-type calcium channels in the formation of long-lasting plateau potentials of Purkinje neurons (De Schutter & Bower, 1994; Jaeger et al. 1997). Previous observations have indicated that membrane potential bistability will arise in any neuronal system that possesses a non-inactivating inward current, if, and only if, the derivative of the inward current is greater than the sum of all other transmembrane currents (Williams et al. 1997; Hughes et al. 1999). Theoretical evidence suggests that in Purkinje neurons, the activation of dendritic P/Q-type calcium currents fulfills this requirement (Yuen et al. 1995). We find, however, that selective blockade of P/Q-type calcium channels fails to block bistability. Furthermore, dendritic recordings revealed the absence of dendritic calcium electrogenesis accompanying the upper state (Fig. 6; see, in contrast, Jaeger et al. 1997), although dendritic electrogenesis could be evoked at these sites in response to large-amplitude positive-current steps (data not shown). We suggest, therefore, that under our experimental conditions, the upper tonic action potential firing state fails to reach the threshold for the recruitment of a significant fraction of dendritic P/Q-type calcium channels. The emergence of bistability following blockade of IH, however, indicates that the magnitude of INap and other transmembrane currents are balanced in such a way as to favour the stable generation of membrane potential bistability. The existence of intrinsic bistability in central neurons has been demonstrated previously in a small fraction of thalamocortical projection neurons, where bistability mediated by the activation of a persistent window component of an inactivating calcium current was shown to emerge following the pharmacological blockade of IH (Williams et al. 1997; Hughes et al. 1999). IH control of membrane potential bistability mediated by the persistent activation of inward currents may, therefore, form a common mechanism in many central neurons.
Physiological role of IH in the control of Purkinje neuron output
Our observations indicate that Purkinje neurons behave in a bistable manner, with IH acting to maintain the membrane potential near the upper stability point. Based on the ability of serotonin to promote bistability in the presence of an inhibitory input, we predict that the negative neuromodulation of IH is likely to disrupt tonic action potential firing in Purkinje neurons in vivo. Consistent with this idea, electrical stimulation of the Raphé complex has previously been demonstrated to inhibit spontaneous action potential firing in Purkinje neurons for long periods (Strahlendorf et al. 1979; Weiss & Pellet, 1982). Harmaline, an agent that increases central and peripheral serotonergic activity, has also been shown to result in the appearance of long alternating periods of tonic action potential firing and quiescence in Purkinje neurons without influences on the inferior olivary system (Stratton et al. 1988). Moreover, cerebellar serotonin levels are known to be increased during locomotor activity (Mendlin et al. 1996). Our data suggest that the ability of synaptic inputs to influence the action potential output of Purkinje neurons will depend upon the state- and activity-dependent modulation of IH. In the absence of neuromodulation of IH, stable tonic action potential firing will be favoured, with only brief interruptions by inhibition. Conversely, following downregulation of IH, synaptic inhibition can interrupt tonic action potential firing for long periods (see Fig. 11), the duration of which will be controlled in vivo by climbing fibre activation and the underlying ‘tone’ of parallel fibre activity. These mechanisms will therefore extend the dynamic range of Purkinje neuron firing during locomotor activity.
Acknowledgments
This work was supported by the Wellcome Trust.
REFERENCES
- Armstrong DM, Rawson JA. Activity patterns of cerebellar cortical neurones and climbing fibre afferents in the awake cat. Journal of Physiology. 1979;289:425–448. doi: 10.1113/jphysiol.1979.sp012745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Azouz R, Jensen MS, Yaari Y. Ionic basis of spike after-depolarization and burst generation in adult rat hippocampal CA1 pyramidal cells. Journal of Physiology. 1996;492:211–223. doi: 10.1113/jphysiol.1996.sp021302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bosmith RE, Briggs I, Sturgess NC. Inhibitory actions of ZENECA ZD 7288 on whole-cell hyperpolarization activated inward current (If) in guinea-pig dissociated sinoatrial node cells. British Journal of Pharmacology. 1993;110:343–349. doi: 10.1111/j.1476-5381.1993.tb13815.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Callaway JC, Ross WN. Spatial distribution of synaptically activated sodium concentration changes in cerebellar Purkinje neurones. Journal of Neurophysiology. 1997;77:145–152. doi: 10.1152/jn.1997.77.1.145. [DOI] [PubMed] [Google Scholar]
- Crepel F, Debono M, Flores R. Alpha-adrenergic inhibition of rat cerebellar Purkinje cells in vitro: a voltage-clamp study. Journal of Physiology. 1987;383:487–498. doi: 10.1113/jphysiol.1987.sp016423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crepel F, Penit-Soria J. Inward rectification and low threshold calcium conductance in rat cerebellar Purkinje cells. An in vitro study. Journal of Physiology. 1986;372:1–23. doi: 10.1113/jphysiol.1986.sp015993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Schutter E, Bower JM. An active membrane model of the cerebellar Purkinje cell. I. Simulation of current clamps in slice. Journal of Neurophysiology. 1994;71:375–400. doi: 10.1152/jn.1994.71.1.375. [DOI] [PubMed] [Google Scholar]
- Dove LS, Abbott LC, Griffith WH. Whole-cell and single-channel analysis of P-type calcium currents in cerebellar purkinje cells of leaner mutant mice. Journal of Neuroscience. 1998;18:7687–7699. doi: 10.1523/JNEUROSCI.18-19-07687.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eccles JC, Llinas R, Sasaki K. The excitatory synaptic action of climbing fibres on the purkinje cells of the cerebellum. Journal of Physiology. 1966a;182:268–296. doi: 10.1113/jphysiol.1966.sp007824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eccles JC, Llinas R, Sasaki K. Intracellularly recorded responses of the cerebellar Purkinje cells. Experimental Brain Research. 1966b;1:161–183. doi: 10.1007/BF00236869. [DOI] [PubMed] [Google Scholar]
- Gahwiler BH, Llano I. Sodium and potassium conductances in somatic membranes of rat Purkinje cells from organotypic cerebellar cultures. Journal of Physiology. 1989;417:105–122. doi: 10.1113/jphysiol.1989.sp017793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghamari-Langroudi M, Bourque CW. Excitatory role of the hyperpolarization-activated inward current in phasic and tonic firing of rat supraoptic neurones. Journal of Neuroscience. 2000;20:4855–4863. doi: 10.1523/JNEUROSCI.20-13-04855.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Granit R, Phillips CG. Excitatory and inhibitory processes acting upon individual Purkinje cells of the cerebellum in cats. Journal of Physiology. 1956;133:520–547. doi: 10.1113/jphysiol.1956.sp005606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harris NC, Constanti A. Mechanism of block by ZD 7288 of the hyperpolarization-activated inward rectifying current in guinea pig substantia nigra neurones in vitro. Journal of Neurophysiology. 1995;74:2366–2378. doi: 10.1152/jn.1995.74.6.2366. [DOI] [PubMed] [Google Scholar]
- Häusser M, Clark BA. Tonic synaptic inhibition modulates neuronal output pattern and spatiotemporal synaptic integration. Neuron. 1997;19:665–678. doi: 10.1016/s0896-6273(00)80379-7. [DOI] [PubMed] [Google Scholar]
- Hughes SW, Cope DW, Toth TI, Williams SR, Crunelli V. All thalamocortical neurones possess a T-type Ca2+‘window’ current that enables the expression of bistability-mediated activities. Journal of Physiology. 1999;517:805–815. doi: 10.1111/j.1469-7793.1999.0805s.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jaeger D, De Schutter E, Bower JM. The role of synaptic and voltage-gated currents in the control of Purkinje cell spiking: a modeling study. Journal of Neuroscience. 1997;17:91–106. doi: 10.1523/JNEUROSCI.17-01-00091.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kay AR, Sugimori M, Llinas R. Kinetic and stochastic properties of a persistent sodium current in mature guinea pig cerebellar Purkinje cells. Journal of Neurophysiology. 1998;80:1167–1179. doi: 10.1152/jn.1998.80.3.1167. [DOI] [PubMed] [Google Scholar]
- Lang EJ, Sugihara I, Welsh JP, Llinas R. Patterns of spontaneous Purkinje cell complex spike activity in the awake rat. Journal of Neuroscience. 1999;19:2728–2739. doi: 10.1523/JNEUROSCI.19-07-02728.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li SJ, Wang Y, Strahlendorf HK, Strahlendorf JC. Serotonin alters an inwardly rectifying current (Ih) in rat cerebellar Purkinje cells under voltage clamp. Brain Research. 1993;617:87–95. doi: 10.1016/0006-8993(93)90617-v. [DOI] [PubMed] [Google Scholar]
- Llinas R, Sugimori M. Electrophysiological properties of in vitro Purkinje cell dendrites in mammalian cerebellar slices. Journal of Physiology. 1980a;305:197–213. doi: 10.1113/jphysiol.1980.sp013358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Llinas R, Sugimori M. Electrophysiological properties of in vitro Purkinje cell somata in mammalian cerebellar slices. Journal of Physiology. 1980b;305:171–195. doi: 10.1113/jphysiol.1980.sp013357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maccaferri G, McBain CJ. The hyperpolarization-activated current (Ih) and its contribution to pacemaker activity in rat CA1 hippocampal stratum oriens-alveus interneurones. Journal of Physiology. 1996;497:119–130. doi: 10.1113/jphysiol.1996.sp021754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCormick DA, Pape HC. Properties of a hyperpolarization-activated cation current and its role in rhythmic oscillation in thalamic relay neurones. Journal of Physiology. 1990;431:291–318. doi: 10.1113/jphysiol.1990.sp018331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Madeja M. Do neurones have a reserve of sodium channels for the generation of action potentials? A study on acutely isolated CA1 neurones from the guinea-pig hippocampus. European Journal of Neuroscience. 2000;12:1–7. doi: 10.1046/j.1460-9568.2000.00871.x. [DOI] [PubMed] [Google Scholar]
- Mainen ZF, Joerges J, Huguenard JR, Sejnowski TJ. A model of spike initiation in neocortical pyramidal neurones. Neuron. 1995;15:1427–1439. doi: 10.1016/0896-6273(95)90020-9. [DOI] [PubMed] [Google Scholar]
- Mendlin A, Martin FJ, Rueter LE, Jacobs BL. Neuronal release of serotonin in the cerebellum of behaving rats: an in vivo microdialysis study. Journal of Neurochemistry. 1996;67:617–622. doi: 10.1046/j.1471-4159.1996.67020617.x. [DOI] [PubMed] [Google Scholar]
- Nam SC, Hockberger PE. Analysis of spontaneous electrical activity in cerebellar Purkinje cells acutely isolated from postnatal rats. Journal of Neurobiology. 1997;33:18–32. doi: 10.1002/(sici)1097-4695(199707)33:1<18::aid-neu3>3.0.co;2-g. [DOI] [PubMed] [Google Scholar]
- Pape HC. Queer current and pacemaker: the hyperpolarization-activated cation current in neurones. Annual Review of Physiology. 1996;58:299–327. doi: 10.1146/annurev.ph.58.030196.001503. [DOI] [PubMed] [Google Scholar]
- Raman IM, Bean BP. Resurgent sodium current and action potential formation in dissociated cerebellar Purkinje neurones. Journal of Neuroscience. 1997;17:4517–4526. doi: 10.1523/JNEUROSCI.17-12-04517.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raman IM, Bean BP. Ionic currents underlying spontaneous action potentials in isolated cerebellar Purkinje neurones. Journal of Neuroscience. 1999;19:1663–1674. doi: 10.1523/JNEUROSCI.19-05-01663.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rapp M, Yarom Y, Segev I. Modeling back propagating action potential in weakly excitable dendrites of neocortical pyramidal cells. Proceedings of the National Academy of Sciences of the USA. 1996;93:11985–11990. doi: 10.1073/pnas.93.21.11985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roth A, Häusser M. Compartmental models of rat cerebellar Purkinje cells based on simultaneous somatic and dendritic patch-clamp recordings. Journal of Physiology. 2001;535:445–472. doi: 10.1111/j.1469-7793.2001.00445.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strahlendorf JC, Strahlendorf HK, Barnes CD. Modulation of cerebellar neuronal activity by raphe stimulation. Brain Research. 1979;169:565–569. doi: 10.1016/0006-8993(79)90405-0. [DOI] [PubMed] [Google Scholar]
- Stratton SE, Lorden JF, Mays LE, Oltmans GA. Spontaneous and harmaline-stimulated Purkinje cell activity in rats with a genetic movement disorder. Journal of Neuroscience. 1988;8:3327–3336. doi: 10.1523/JNEUROSCI.08-09-03327.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stuart G. Voltage-activated sodium channels amplify inhibition in neocortical pyramidal neurones. Nature Neuroscience. 1999;2:144–150. doi: 10.1038/5698. [DOI] [PubMed] [Google Scholar]
- Stuart GJ, Dodt HU, Sakmann B. Patch-clamp recordings from the soma and dendrites of neurones in brain slices using infrared video microscopy. Pflügers Archiv. 1993;423:511–518. doi: 10.1007/BF00374949. [DOI] [PubMed] [Google Scholar]
- Stuart G, Häusser M. Initiation and spread of sodium action potentials in cerebellar Purkinje cells. Neuron. 1994;13:703–712. doi: 10.1016/0896-6273(94)90037-x. [DOI] [PubMed] [Google Scholar]
- Weiss M, Pellet J. Raphe-cerebellum interactions. II. Effects of midbrain raphe stimulation and harmaline administration on single unit activity of cerebellar cortical cells in the rat. Experimental Brain Research. 1982;48:171–176. doi: 10.1007/BF00237212. [DOI] [PubMed] [Google Scholar]
- Williams SR, Stuart GJ. Mechanisms and consequences of action potential burst firing in rat neocortical pyramidal neurones. Journal of Physiology. 1999;521:467–482. doi: 10.1111/j.1469-7793.1999.00467.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams SR, Toth TI, Turner JP, Hughes SW, Crunelli V. The ‘window’ component of the low threshold Ca2+ current produces input signal amplification and bistability in cat and rat thalamocortical neurones. Journal of Physiology. 1997;505:689–705. doi: 10.1111/j.1469-7793.1997.689ba.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuen GL, Hockberger PE, Houk HC. Bistability in cerebellar Purkinje cell dendrites modelled with high-threshold calcium and delayed-rectifier potassium channels. Biological Cybernetics. 1995;73:375–388. doi: 10.1007/BF00199473. [DOI] [PubMed] [Google Scholar]