

Abstract
During fatiguing submaximal contractions a constant force production can be obtained at the cost of an increasing central command intensity. Little is known about the interaction between the underlying central mechanisms driving motor behaviour and cognitive functions. To address this issue, subjects performed four tasks: an auditory choice reaction task (CRT), a CRT simultaneously with a fatiguing or a non-fatiguing submaximal muscle contraction task, and a fatiguing submaximal contraction task alone. Results showed that performance in the single-CRT condition was relatively stable. However, in the fatiguing dual-task condition, performance levels in the cognitive CRT deteriorated drastically with time-on-task. Moreover, in the fatiguing dual-task condition the rise in force variability was significantly larger than during the fatiguing submaximal contraction alone. Thus, our results indicate a mutual interaction between cognitive functions and the central mechanisms driving motor behaviour during fatigue. The precise nature of this interference, and at what level this interaction takes place is still unknown.
Performing a motor task for long periods of time induces motor fatigue, which is generally defined as a decline in a person's ability to exert force (Bigland-Ritchie et al. 1995). Much of this decline is caused by a reversible weakening of the muscles themselves (muscle fatigue). In submaximal contractions associated with muscle fatigue, the central nervous system may still elicit a constant force for some time by activating progressively greater numbers of motoneurones and/or by increasing their discharge rate. Under these conditions, a constant force output is produced at the cost of a progressively increasing central command intensity and subjects are then also often aware of having to exert more ‘effort’ in order to maintain the contraction. Thus, fatiguing muscles place increasing demands on the central mechanisms driving motor behaviour. In addition, part of the progressively increasing motor fatigue, as seen in various types of voluntary behaviour, is caused by mechanisms inside the central nervous system itself (Gandevia, 2001).
Besides the fatigue associated with motor behaviour, there have also been numerous studies of ‘fatigue’ of a more central kind, related to factors such as mental load and time-on-task for cognitive tasks (Hockey, 1993; Meijman, 1997). Such central or ‘mental’ fatigue may be associated with a considerable deterioration of cognitive functions (Lorist et al. 2000), a factor of theoretical as well as practical importance (e.g. in work psychology).
More than 70 years ago, it was already concluded that muscular contraction has an effect on mental processes. The relationship between mental information processing and muscular tension, however, was then, and still is, far from simple (Freeman, 1933). When investigating the interaction between motor functions and cognitive performance, fatigue might be an interesting tool with which to manipulate the contribution of the central mechanisms driving motor behaviour. It is expected that the contribution of central mechanisms increases during fatigue. Moreover, fatigue is a common phenomenon that is related to deterioration in performance, and little is known about the extent to which motor fatigue has an effect on cognitive functions and/or whether and how motor functions are influenced by the simultaneous performance of cognitive tasks. Recently, Schubert and colleagues (1998) addressed this issue using two motor tasks, differing in force requirements, in combination with an auditory classification task. They failed to find effects of ‘fatigue’ on cognitive task performance. Unfortunately, they did not measure the amount of fatigue and it is uncertain whether the intensity of the protocol used was high enough to produce high levels of motor fatigue and related changes in central command intensity. At the end of their experiments subjects were still able to perform the force task.
Our present investigation was designed to explore to what extent interactions take place between simple kinds of fatiguing and non-fatiguing motor behaviour, and a cognitive choice reaction task (CRT). More pronounced effects were expected during fatiguing contractions, which are supposed to induce an increasing involvement of central force-controlling mechanisms. The presence of motor fatigue was determined by measuring the size of the maximal voluntary contraction force.
Methods
Participants
Sixteen healthy adults (10 males, 6 females), ranging in age from 21 to 44 years, were paid for participating in the study. All subjects were right-handed and non-smokers. They all had normal or corrected-to-normal vision and intact hearing. Written informed consent to participate was obtained from each subject prior to the study. All procedures were undertaken with the approval of the local ethics committee and conformed with the standards set out in the Declaration of Helsinki (2000).
Experimental set-up
Subjects sat behind an experimental table with their lower arms on the table and their right hand fixed in the experimental set-up. The elbows were flexed to 135 deg and the right forearm was held in a position halfway between supination and pronation. The abduction force of the first dorsal interosseus muscle of the right hand was measured. For this purpose the right index finger was held slightly abducted within a snugly fitting ring around the proximal interphalangeal joint. The ring was rigidly connected to an isometric force transducer (for details see Zijdewind & Kernell, 1994). The right arm was immobilised with pressure plates and Velcro tape. Electromyographic (EMG) recordings were obtained from the first dorsal interosseus muscle of both hands with a surface electrode (4 mm diameter) placed over the muscle belly and a reference electrode placed at the metacarpophalangeal joint of the index fingers. A band-shaped earth electrode was placed around the right wrist. EMG and force recordings were amplified, filtered, and analysed using a PC equipped with a data-acquisition interface (1401+, Cambridge Electronic Design, Cambridge, UK). The sampling rate was 2000 and 500 Hz for EMG and force recordings, respectively.
Submaximal contraction task with intervening maximal voluntary contractions (MVCs)
Subjects viewed a dual-beam oscilloscope, one beam continuously displaying the isometric force production of the subject and the second displaying the desired level of contraction force. A task started with the presentation of a 2000 Hz tone, indicating that subjects had to perform a MVC, followed 4 s later by a 200 Hz tone, indicating the end of the MVC and 4 s thereafter a second 200 Hz tone, indicating the start of the submaximal contraction. The subjects were instructed to maintain their force steadily at the target level for 1 min periods during the task. After 1 min, subjects performed a 4 s MVC followed by 4 s rest.
Choice reaction task
The stimuli for the CRT were 500 or 900 Hz pure tones, presented binaurally via speakers (duration: 50 ms, intensity: 70 dB(A)). The inter-tone interval varied randomly between 1100 and 1300 ms. Random sequences of ‘frequent’ (70 % occurrence) and ‘infrequent’ stimuli (30 %) were presented in blocks of 50 stimuli over a period of approximately 1 min. The subjects had to respond to the auditory stimuli by pressing one of two response buttons with their left middle or index finger. Half of the subjects were instructed to respond with their middle finger to frequent stimuli and with their index finger to infrequent stimuli. The other half of the subjects received opposite instructions. For half of the subjects in each group, 500 Hz tones were ‘frequent’ stimuli, while for the other half of the subjects these tones served as ‘infrequent’ stimuli. As a result, there were four different versions of the CRT: (1) frequent stimuli 500 Hz, middle finger response; (2) frequent stimuli 900 Hz, middle finger response; (3) frequent stimuli 500 Hz, index finger response; (4) frequent stimuli 900 Hz, index finger response. The presentation of stimuli and the collection of the subjects’ responses were controlled by Micro Experimental Laboratory Professional Software (MEL v2.0; Schneider, 1988), in conjunction with the MEL Serial Response Box.
General procedure (Fig. 1)
Figure 1. Schematic representation of experimental procedures in the two sessions.
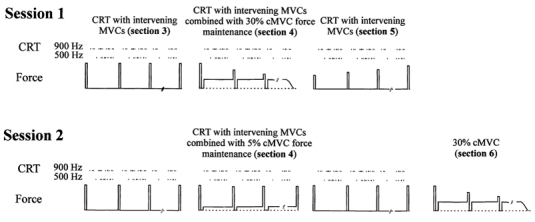
Sections 1 (practice) and 2 (control maximum voluntary contraction, cMVC, determination) have been left out for reasons of clarity. Section 3 consists of a choice reaction task (CRT) with intervening maximal voluntary contractions (MVC). In section 4 the CRT is combined with a submaximal contraction; 30 % cMVC in session 1 and 5 % cMVC in session 2. Section 5 is a repetition of section 3. In section 6 only the submaximal contraction is performed. In this illustration the frequent stimulus is 900 Hz.
Sixteen subjects participated in the first experimental session. A random sample of 10 out of the initial 16 subjects participated in the second session. The two sessions were separated by at least 1 week. Each session started at 13.00 h or 15.00 h and lasted about 1.5 h. Each experimental session consisted of five or six sections with different types of required task performance.
In session 1, five sections were performed, in the following order:
(1) Practice
One group of 150 CRT trials, followed by three blocks of 50 CRT trials with intervening MVCs.
(2) MVC measurements
Three MVCs of index finger abduction at about 1 min intervals. The strongest of these contractions was designated as the ‘control MVC’ (cMVC).
(3) CRT with intervening MVCs
Fourteen blocks of 50 CRT trials, each block being followed by a MVC.
(4) CRT combined with submaximal contraction (dual-task condition)
Blocks of 50 CRT trials performed during a 1 min submaximal contraction at 30 % cMVC followed by a 4 s MVC and 4 s of rest. Subjects continued this sequence (1 min dual task, 4 s MVC, 4 s rest) until they failed to maintain the target force level. On average this happened after 7 min (range 4–18 min).
(5) CRT with intervening MVCs
Same as section 3.
In section 4, subjects were instructed to regard the maintenance of the submaximal force as their primary task while still trying to respond as quickly as possible in the CRT (secondary task), maintaining a high level of accuracy. The procedures of session 2 were similar to those of session 1, except for the following aspects:
section 1 consisted of only three blocks of 50 CRT trials;
the target force in section 4 was reduced from 30 % to 5 % cMVC, each subject performing an equal number of dual-task blocks to those of session 1. To keep visual task aspects similar across sessions the gain of the oscilloscope was increased by a factor of five in session 2;
the number of CRT blocks performed in section 5 was reduced from 14 to 6;
following section 5, the subjects performed an additional submaximal force-maintenance task at 30 % cMVC without any simultaneous CRT (section 6).
Data reduction and statistical procedure
The first two trials of each block of the CRT were regarded as practice trials and were excluded from analysis. Reaction times (RT) were scored as hits when a correct button was pressed within mean RT ± 2 s.d. after stimulus onset; all accepted RT values exceeded 150 ms. All other responses were considered as being incorrect. Mean RTs and error percentages were calculated separately for frequent and infrequent stimuli, for the first block (48 trials) and for the first and second half of each section. In the case of an uneven number of blocks in section 4, the middle block was left out. Mean MVC values were also calculated for the first and second half of each section. Data were subjected to ANOVA (SPSS Inc., Chicago, USA) for repeated measurements, using the ε≈*-adjustment procedure recommended by Quintana & Maxwell (1994). When the main analysis indicated a significant interaction (α = 0.05) between factors, follow-up analyses were performed, adjusting error rates according to Bonferroni. It should be noted that only 10 of the subjects participated in session 2. For comparisons between sessions 1 and 2, only data from these 10 subjects were used for the analysis.
Results
No significant differences were found between CRT performances in the different test versions (frequent stimulus high or low tone; response with index or middle finger). Therefore data were pooled across the four CRT versions. In addition, the results were not dependent upon the absolute size of the cMVC.
General observations
Figure 2 shows the reaction times (A) and the number of incorrect responses (B) for the different sections of the experiments. As can be seen in Fig. 2, in general subjects reacted faster to frequent than to infrequent stimuli (F(1,15) = 175.66, P < 0.001), and more incorrect button presses were made in response to infrequent than to frequent stimuli (F(1,15) = 87.98, P < 0.001).
Figure 2. Mean reaction times and percentage incorrect responses.
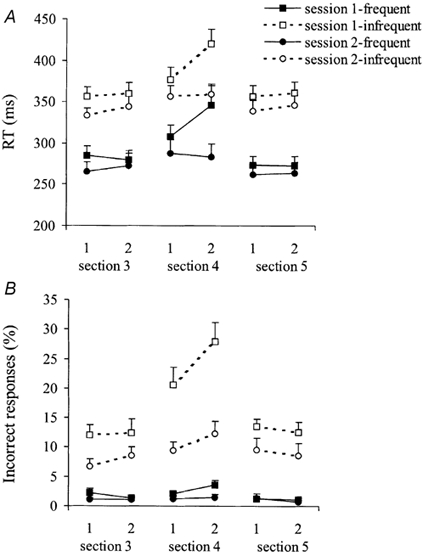
Mean reaction times (± s.e.m.; A) and percentage of incorrect responses (± s.e.m.; B) in the first and second half of sections 3 (single task), 4 (dual task) and 5 (single task) for session 1 (squares; n = 16) and session 2 (circles; n = 10) are shown. Reaction times and percentage of incorrect responses are shown separately for frequent (filled symbols) and infrequent stimuli (open symbols).
Session 1 (30 % cMVC condition)
CRT alone (sections 3 and 5)
No time-on-task effects were seen for CRT performance in the single-task condition. The RTs remained stable during 15 min of single-task performance (F(1,15) = 0.26, n.s.) and the same was true for the number of incorrect responses (F(1,15) = 0.18, n.s.).
Dual task: CRT and 30 % cMVC force maintenance (section 4)
When comparing RTs in the single- and dual-task condition, no significant difference was observed in the 1st minute of task performance (317, 335 and 327 ms for sections 3 (single task), 4 (dual task) and 5 (single task), respectively; F(2,30) = 1.63, n.s.). However, for infrequent stimuli the percentage of incorrect responses in the first block was higher in the dual-task condition than in single-task conditions (20.7 ± 11.7 % and 12.0 ± 7.1 %, respectively; section × stimulus type: F(2,30) = 4.94, P = 0.015). For frequent stimuli, no such differences were observed.
Of major interest for this study were the effects of time-on-task in the dual-task condition. Subjects were asked to maintain a submaximal contraction at 30 % cMVC while simultaneously performing the CRT. In addition, task performance was interrupted every minute by a MVC of the index finger of the dominant (right) hand. During the dual task, the force of these intervening MVCs decreased from 77.3 % cMVC in the first half of the task to 51.9 % cMVC in the second half (Fig. 3; F(1,15) = 141.31, P < 0.001); that is to say, there were clear indications of a progressively increasing degree of motor fatigue. The average levels of submaximal force indicated that subjects were indeed able to produce force levels close to 30 % during the first half of the task. However, the coefficient of variation ((s.d./mean) × 100) of the maintained submaximal force increased from 7.8 % in the first half to 13.6 % in the second half of the test (F(1,15) = 52.09, P < 0.001), thus subject force response became more variable with time-on-task. As is clearly visible in Fig. 2, the increasing degree of motor fatigue was accompanied by a dramatic decline in CRT performance, regarding both reaction times (F(1,15) = 15.85, P < 0.001) and the percentage of correct responses (F(1,15) = 5.96, P < 0.027).
Figure 3. Mean MVC force.
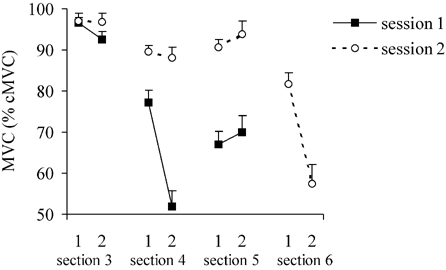
Mean reaction times (± s.e.m.) in the first and second half of sections 3 (single task: CRT), 4 (dual task: CRT and force task), 5 (single task: CRT) and 6 (single task: force task) for session 1 (▪; n = 16) and session 2 (○; n = 10) are shown.
Session 2 (5 % cMVC condition)
The decline of CRT performance in the dual-task condition of session 1, as reflected in the behavioural parameters (RT and the percentage of incorrect responses, Fig. 2), could be the result of the increasing demands placed on central force-controlling mechanisms induced by motor fatigue and/or might be caused by the demands of simultaneously performing a CRT task and a force-control task for a longer time period. To distinguish between these possibilities we added a second test session for 10 of the subjects, now combining the CRT with a maintained contraction task at a force level low enough to cause little or no motor fatigue (5 % cMVC). Furthermore, in session 2 we also evaluated the effect of dual-task performance on motor behaviour. To examine whether the deterioration of motor behaviour during the maintained-contraction task (decline of MVC force: Fig. 3; increase of force variability: Fig. 4) was due to the addition of a secondary cognitive task, we added at the end of session 2 a maintained-contraction task at 30 % cMVC, as performed under single-task conditions (section 6).
Figure 4. Force variation.
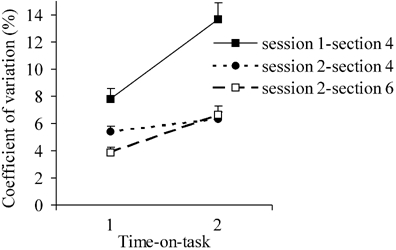
The amount of force variation (± s.e.m.) in the first and second half of the dual-task condition (section 4) in session 1 (▪; n = 16) and session 2 (•: n = 10) and the force-alone condition (section 6; □; n = 10) are shown.
CRT alone (sections 3 and 5)
With regard to RT measurements, the CRT results in session 2 (5 % cMVC condition: sections 3 and 5) were similar to those of session 1 (30 % cMVC condition: section 3, F(1,9) = 1.62, n.s.; section 5, F(1,9) = 2.15, n.s.). In section 3, the percentage of incorrect responses was, unexpectedly, somewhat smaller in session 2 than in session 1 (6.7 % vs 10.8 %; data for first half-section: F(1,9) = 7.36, P = 0.024). For section 3, no significant differences were found in MVC values between the two experimental sessions (F(1,9) = 0.73, n.s.). At the start of section 5, the intermittent MVCs were, as expected, significantly higher following the low-level maintained-contraction task (session 2, 5 % cMVC) than after the fatiguing maintained contractions at 30 % cMVC (session 1; F(1,8) = 48.24, P < 0.001).
Dual task: CRT and 5 % cMVC force maintenance (section 4)
The MVC force showed a considerably smaller (non-significant, F(1,9) = 0.47, n.s.) decline during the low-level force-maintenance of session 2 than that seen in session 1 (F(1,9) = 96.76, P < 0.001); that is to say, there was a significantly lower degree of motor fatigue during the dual task of session 2 than that seen in session 1 (Fig. 3). This smaller degree of fatigue was associated with a significant improvement in the CRT behaviour compared to the 30 % cMVC condition: in the dual-task section of session 2 (5 % cMVC: section 4), no significant deterioration was seen in the RT data with time-on-task (time-on-task: F(1,9) = 0.02, n.s.) and there was a lower percentage of incorrect responses to infrequent stimuli than was found in section 4 of session 1 (Fig. 2: F(1,9) = 15.11, P = 0.004).
As in the 30 % cMVC condition, in the low-level maintained contraction of session 2, an increased variability of force took place during the course of the test. This effect was, however, significantly smaller for the 5 % cMVC contractions of session 2 than for the stronger contractions of session 1 (session × time-on-task: F(1.60,14.42) = 14.01, P = 0.001).
Maintained-contraction task alone (30 % cMVC; section 6)
The reduction in the force of intervening MVCs with time-on-task was similar for the 30 % cMVC single-task condition (session 2) and for the dual-task condition (session 1; F(1,8) = 0.00, n.s.); that is, the addition of a cognitive task did not have an effect on intervening MVCs. Interestingly, there was a difference in force variability between single- and dual-task performance (F(1,9) = 28.33, P < 0.001). In the dual-task condition subjects showed more variability in performance compared to the single-task condition. However, the increase in force variability with time-on-task was less pronounced during the maintained-contraction task performed without the concurrent CRT (session 2, section 6) than for tests in which both of these tasks were combined (session 1, section 4; Fig. 4; session × time-on-task: F(1.85,16.68) = 6.18, P = 0.011). Thus, there was an increase in force variability in both the single- and the dual-task condition, but the observed increase in the force variability was more pronounced in the dual-task condition.
Discussion
During fatiguing submaximal contractions, a constant force production can be obtained at the cost of an increasing central command intensity. However, little is known about the effects of increasing demands on central mechanisms induced by motor fatigue on cognitive task performance. In the present study we evaluated the effects of motor fatigue on cognitive task performance. By using a dual-task protocol we were able to show that cognitive performance is seriously affected by motor fatigue. Subjects performed an auditory CRT either alone or simultaneously with a submaximal contraction task. Performance in the single-task conditions was relatively stable; subjects were able to maintain performance levels at an adequate level throughout 15 min. In the fatiguing dual-task condition (30 % cMVC), however, performance levels in the secondary CRT changed with time-on-task. Subjects reacted more slowly and made more incorrect responses at the end of the task. In the 5 % cMVC dual-task condition, in which the force of interspersed MVCs did not show signs of motor fatigue, no time-related change in the behavioural data was observed. The observed deterioration of task performance with time-on-task in the high force condition may be related to the increased demands placed on central mechanisms driving motor behaviour with increasing motor fatigue. The absence of time-on-task effects in the low force condition, in which no changes in central mechanisms should have occurred, seems to support this conclusion.
In the fatiguing situation, the progressive decrement of the dual-task performance with time-on-task indicates that with increasing motor fatigue, the interference between the motor task and the CRT became more pronounced. In contrast to our findings in the 30 % cMVC conditions, neither a decline in MVC values was observed, nor did the behavioural data show significant changes with time-on-task when subjects performed a submaximal contraction task in which the force level was reduced to 5 % cMVC. The absence of time-on-task effects in the 5 % cMVC force-maintenance task is consistent with the findings reported by Schubert et al. (1998). They combined an auditory CRT with either a force task (about 50 % of maximum force output) or a displacement task (minimal force requirements). In neither condition did their results show differential time-on-task effects. Although the force level in their study was 50 % cMVC, the question is, as already mentioned, whether the intensity of their protocol was high enough to induce motor fatigue, since subjects were still able to perform the force task at the end of their experimental session.
At this point it should be stressed that, for the combination of CRT and force control, time-on-task per se did not influence the CRT responses. Instead, our results indicate that the presence of fatigue and the related involvement of central mechanisms driving motor behaviour are of crucial importance for causing a deterioration of cognitive task performance.
Single- and dual-task performance differed with respect to reaction speed (5 % cMVC) and accuracy (30 % cMVC) at the start of a section when subjects were not fatigued. This decline in performance of the secondary CRT in the dual-task condition showed that simultaneously performing the two tasks was more demanding than single-task performance. Moreover, this decline indicated that the dual-task condition imposed a 100 % workload on the subjects’ limited attentional resources. No residual resources or effort seemed available to compensate for the increasing task demands in the dual-task situation compared to single-task performance. The fatigue-associated deterioration in the secondary task performance might have been related to added resource investment into the primary task, thereby producing a reduction in the resources available for adequate performance in the secondary task. Prolonged investment of attentional resources is in itself fatiguing (Hockey, 1993), therefore it seems reasonable to assume that subjects seek methods to perform a task in a way that conserves resources at acceptable performance levels. This might be done by using heuristics, which are mental shortcuts that provide reasonably good performance without the investment of too much effort (Meijman, 1997). The increase in incorrect responses with time-on-task indicated that subjects used a more risky strategy at the end of the fatiguing dual-task condition. Subjects might have emitted ‘fast guesses’ (Pachella, 1974; Gratton et al. 1988), in which a response is initiated as soon as the stimulus is detected. Little or no evidence for a specific response is then required to exceed the decision criterion and execute a response, and therefore a reduced demand is placed on attentional resources. The nature of this fast guess is that of the most probable response. Indeed, our data showed that most incorrect responses involved pressing the ‘frequent’ button instead of the ‘infrequent’ one, and the mean RTs for these incorrect responses were significantly faster (274 ms) than RTs for correct ‘frequent’ responses (294 ms).
In addition, the subjects showed faster and more accurate responses to frequent, high-probability stimuli as compared to infrequent, low-probability stimuli. We found a response time advantage of 78 ms for stimuli occurring in 70 % compared to stimuli occurring in 30 % of the trials. It has been found that differences between high- and low-probability stimuli are related to the state of preparedness of subjects (Miller, 1998); the distance between motor activation and the threshold for action is relatively small in these prepared conditions, and only a small increment is needed to exceed this threshold, resulting in fast responses.
Furthermore, as argued earlier, the decrement of CRT performance in the dual-task condition might reflect changes in the central force-controlling mechanisms. During motor fatigue, an increase in the amount of activity in other, non-target muscles is often found (Dimitrijevic et al. 1992; Mayston et al. 1994). This associated activity was also observed in the contralateral hand (Zijdewind & Kernell, 2001), which could have resulted in interference with the speed of finger movement in the hand pressing the button in the CRT in the present study. However, it is unlikely that the increase in incorrect responses could be explained by such an increase in associated muscle activity. Moreover, M. M. Lorist & I. Zijdewind (unpublished results) showed that, when using a mathematical task in which no manual response was required instead of the CRT, a similar decline in performance was observed.
What is striking in the present study is that the interference between the motor task and the cognitive task went in both directions. Not only the CRT, but also the performance of the submaximal maintained contraction was negatively affected during the dual-task condition. The subjects’ ability to keep the produced force steady at the target level became impaired. It is well known that fatigue is often associated with an increased variability in force production (Lippold, 1981). In addition to a difference in force variability between single- and dual-task performance at the start of the tasks when subjects were not yet fatigued, we observed that the amount of variation in the maintained submaximal force increased to higher levels in the dual-task condition than that observed in the single 30 % cMVC task. Thus, interference between the two tasks at a central level must have played a role in the increased force variability in the fatiguing dual-task condition. The mechanisms underlying this variability are still unknown.
In conclusion, we observed a mutual interaction during motor fatigue between a motor task and a cognitive task. The interactions resulted in a decline in the performance of the motor task as well as the cognitive task. The question remains as to the nature of this competition, and at which level both tasks interfere. An additional question concerns whether this interaction could also be evoked by non-fatiguing high-effort contractions. Since at the end of a submaximal fatiguing contraction, when the subject is no longer able to produce the desired force, the subject is activating the muscle to near-maximal values. Pilot experiments showed that in high-force conditions, fatigue was induced very rapidly and during short, non-fatiguing contractions the CRT data became unreliable because of the low number of trials.
Information-processing theories assume that there are multiple resources associated with different structural processes (Mulder, 1983; Sanders, 1983; Wickens, 1984; Wickens & Hollands, 2000). In the present study, a submaximal contraction task in which information was presented visually was combined with an auditory CRT. Thus, these tasks relied on distinct perceptual systems. Interference between the two tasks was therefore not expected at the perceptual level and in correspondingly related brain areas. The response modality was similar in both tasks (manual). Possible interference between the two tasks might be related to processes at this stage. The subjects had to translate perceived information into either a button press or the production of maximal or submaximal contraction strength. However, using an experimental set-up in which the secondary cognitive task did not require a manual response, no support for this hypothesis was found (M. M. Lorist & I. Zijdewind, unpublished data).
A possible candidate concerning brain areas playing a role in the observed effects is the dorsolateral prefrontal cortex. Dettmers and colleagues (1995, 1996) showed, using positron emission tomography, that additional areas were activated during a sustained contraction compared to contractions at high force levels. Interestingly, during the sustained task a progressively increasing activity was observed in the dorsolateral prefrontal cortex, an area involved in the organisation and co-ordination of actions in the time domain (Fuster, 1997).
Besides being of interest for understanding (central) processes associated with fatigue and with motor and cognitive behaviour, the present results would also seem relevant as a starting point for further investigations in the applied field of work psychology. A deterioration of cognitive functions, provoked by motor fatigue, might lead to potentially dangerous errors in work situations. Moreover, these results might be of importance in the explanation of cognitive deficits observed in patients suffering from peripheral motor dysfunction. In these patients, tasks requiring low absolute force levels could induce fatigue much faster and therefore they would need to increase the central drive to the motoneurone pool, which could induce a negative effect on cognitive functions.
Acknowledgments
This research was supported by grants from the Netherlands Organization for Scientific Research (NWO; the concerted research action ‘Fatigue at Work’) and the School of Behavioural and Cognitive Neurosciences (BCN; Groningen, The Netherlands).
References
- Bigland-Ritchie B, Rice CL, Garland SJ, Walsh ML. Task-dependent factors in fatigue of human voluntary contractions. In: Gandevia SC, Enoka RM, McComas AJ, Stuart DG, Thomas CK, editors. Fatigue. New York: Plenum; 1995. pp. 361–380. [DOI] [PubMed] [Google Scholar]
- Dettmers C, Fink GR, Lemon RN, Stephan KM, Passingham R, Silbersweig D, Holmes A, Ridding MC, Brooks D, Frackowiak RSJ. The relation between cerebral activity and force in the motor areas of the human brain. Journal of Neurophysiology. 1995;74:802–815. doi: 10.1152/jn.1995.74.2.802. [DOI] [PubMed] [Google Scholar]
- Dettmers C, Lemon RN, Stephan KM, Fink GR, Frackowiak RSJ. Cerebral activation during the exertion of sustained static force in man. NeuroReport. 1996;7:2103–2110. doi: 10.1097/00001756-199609020-00008. [DOI] [PubMed] [Google Scholar]
- Dimitrijevic MR, McKay WB, Sarjanovic I, Sherwood AM, Svirtlih L, VrbovÀ G. Co-activation of ipsi- and contralateral muscle groups during contraction of ankle dorsiflexors. Journal of the Neurological Sciences. 1992;109:49–55. doi: 10.1016/0022-510x(92)90092-y. [DOI] [PubMed] [Google Scholar]
- Freeman GL. The facilitative and inhibitory effects of muscular tension upon performance. American Journal of Psychology. 1933;45:17–52. [Google Scholar]
- Fuster JM. The Prefrontal Cortex. Anatomy, Physiology, and Neuropsychology of the Frontal Lobe. New York: Lippincott-Raven; 1997. [Google Scholar]
- Gandevia SC. Spinal and supraspinal factors in human muscle fatigue. Physiological Reviews. 2001;81:1725–1789. doi: 10.1152/physrev.2001.81.4.1725. [DOI] [PubMed] [Google Scholar]
- Gratton G, Coles MGH, Sirevaag EJ, Eriksen CW, Donchin E. Pre- and poststimulus activation of response channels: a psychophysiological analysis. Journal of Experimental Psychology: Human Perception and Performance. 1988;14:331–344. doi: 10.1037//0096-1523.14.3.331. [DOI] [PubMed] [Google Scholar]
- Hockey GRJ. Cognitive-energetical mechanisms in the management of work demands and psychological health. In: Baddeley A, Weiskrantz L, editors. Attention, Selection, Awareness and Control. Oxford: Oxford University Press; 1993. pp. 328–345. [Google Scholar]
- Lippold O. The tremor in fatigue. In: Porter R, Whelan J, editors. Human Muscle Fatigue. London: Pitman Medical; 1981. pp. 234–248. [Google Scholar]
- Lorist MM, Klein M, Nieuwenhuis S, De Jong R, Mulder G, Meijman TF. Mental fatigue and task control: planning and preparation. Psychophysiology. 2000;37:614–625. [PubMed] [Google Scholar]
- Mayston M, Carter K, Whybrow T, Lockley M, Kelly A, Harrison LM, Stephens JA. Bilateral EMG accompanies unilateral tasks in man. Journal of Physiology. 1994;480. P:44P. [Google Scholar]
- Meijman TF. Mental fatigue and the efficiency of information processing in relation to work times. International Journal of Industrial Ergonomics. 1997;20:31–38. [Google Scholar]
- Miller J. Effects of stimulus-response probability on choice reaction time: Evidence from the lateralized readiness potential. Journal of Experimental Psychology: Human Perception and Performance. 1998;24:1521–1534. [Google Scholar]
- Mulder G. The information processing paradigm: concepts, methods and limitations. Journal of Child Psychology and Psychiatry and Allied Disciplines. 1983;24:19–35. doi: 10.1111/j.1469-7610.1983.tb00100.x. [DOI] [PubMed] [Google Scholar]
- Pachella RG. The interpretation of reaction time in information-processing research. In: Kantowitz B, editor. Human Information Processing: Tutorials in Performance and Cognition. London: Erlbaum; 1974. pp. 41–82. [Google Scholar]
- Quintana SM, Maxwell SE. A Monte Carlo comparison of seven ε-adjustment procedures in repeated measures designs with small sample sizes. Journal of Educational Statistics. 1994;19:57–71. [Google Scholar]
- Sanders AF. Towards a model of stress and human performance. Acta Psychologica. 1983;74:123–167. doi: 10.1016/0001-6918(83)90016-1. [DOI] [PubMed] [Google Scholar]
- Schneider W. Micro experimental laboratory: an integrated system for IBM-PC compatibles. Behavior Research Methods, Instruments, and Computers. 1988;20:206–217. [Google Scholar]
- Schubert M, Johannes S, Kock M, Wieringa BM, Dengler R, MüNTE TF. Differential effects of two motor tasks on ERPs in auditory classification task: Evidence of shared cognitive resources. Neuroscience Research. 1998;30:125–134. doi: 10.1016/s0168-0102(97)00115-6. [DOI] [PubMed] [Google Scholar]
- Wickens CD. Processing resources in attention. In: Parasuraman R, Davies R, editors. Varieties of Attention. New York: Academic Press; 1984. pp. 63–101. [Google Scholar]
- Wickens CD, Hollands JG. Engineering Psychology and Human Performance. Upper Saddle River, NJ, USA: Prentice Hall; 2000. [Google Scholar]
- Zijdewind I, Kernell D. Index finger position and force of the human first dorsal interosseus and its ulnar nerve antagonist. Journal of Applied Physiology. 1994;77:987–997. doi: 10.1152/jappl.1994.77.2.987. [DOI] [PubMed] [Google Scholar]
- Zijdewind I, Kernell D. Bilateral interactions during contractions of intrinsic hand muscles. Journal of Neurophysiology. 2001;85:1907–1913. doi: 10.1152/jn.2001.85.5.1907. [DOI] [PubMed] [Google Scholar]