

Abstract
It has been postulated that alcoholism is associated with abnormalities in glutamatergic neurotransmission. This study examined the density of glutamate NMDA receptor subunits and its associated proteins in the noradrenergic locus coeruleus (LC) in deceased alcoholic subjects. Our previous research indicated that the NMDA receptor in the human LC is composed of obligatory NR1 and regulatory NR2C subunits. At synapses, NMDA receptors are stabilized through interactions with postsynaptic density protein (PSD-95). PSD-95 provides structural and functional coupling of the NMDA receptor with neuronal nitric oxide synthase (nNOS), an intracellular mediator of NMDA receptor activation. LC tissue was obtained from 10 alcohol-dependent subjects and 8 psychiatrically healthy controls. Concentrations of NR1 and NR2C subunits, as well as PSD-95 and nNOS, were measured using Western blotting. In addition we have examined tyrosine hydroxylase (TH), the rate-limiting enzyme in the synthesis of norepinephrine. The amount of NR1 was lower in the rostral (−30%) and middle (−41%)portions of the LC of alcoholics as compared to control subjects. No differences in the amounts of NR2C, PSD-95, nNOS and TH were detected comparing alcoholic to control subjects. Lower levels of NR1 subunit of the NMDA receptor in the LC implicates altered glutamate-norepinephrine interactions in alcoholism.
Keywords: glutamate, NMDA receptor, postsynaptic density protein 95, neuronal nitric oxide synthase, tyrosine hydroxylase, locus coeruleus
1. Introduction
Accumulating evidence suggests that neurophysiological and pathological effects of ethanol are mediated, at least in part, through the glutamatergic system. NMDA receptors are among the highest affinity ethanol targets in the brain Grant and Lovinger, 1995. Ethanol inhibits NMDA receptor function and this effect does not appear to be mediated by any of the well-characterized binding sites on the receptor (e.g. glutamate, glycine, phencyclidine, polyamine sites). Instead, recombinant receptor studies have shown that ethanol may have a selective effect on specific NMDA receptor subunits, with different effects on certain subunit combinations (Jin and Woodward, 2006; Kumari and Ticku, 2000).
NMDA receptors are heteromeric proteins assembled from NR1 and NR2 subunits and, less commonly, the NR3 subunit. The NR2 family contains 4 subunits (A-D) that arise from separate genes. The NR1 subunit has 8 splice variants that are generated by alternative splicing of exon 5, 21, and 22 (Zukin and Bennett, 1995). Both NR1 and NR2 subunits are required to form functional NMDA receptors. The subunit composition of the NMDA receptor affects a number of physiological and pharmacological properties of the NMDA receptor including sensitivity to inhibition by ethanol (Chu et al., 1995; Jin and Woodward 2006).
The locus coeruleus (LC), the major source of norepinephrine in the brain, is regulated by excitatory glutamatergic input from the nucleus paragiganocellularis and medial prefrontal cortex (Aston-Jones et al., 1991; Ennis and Aston-Jones, 1986; Jodo et al., 1998). LC activity is inhibited by ethanol (Aston-Jones et al., 1982; Shefner and Tabakoff, 1985) possibly in part through inhibition of glutamatergic activation of the LC. Our previous study revealed that the NMDA receptor in the human LC is predominantly composed of NR1/NR2C subunits (Karolewicz et al., 2005). Based on recombinant receptor studies, NMDA receptors composed of NR1/NR2C subunits have a lower current amplitude, shorter open time, smaller single conductance, and reduced sensitivity to Mg2+ (Ishii et al., 1993; Kutsuwada et al. 1992; Laurie and Seeburg, 1994; Monyer et al., 1994). The relevance of these recombinant studies to the function of NMDA receptors in the LC is unknown. A recent study demonstrated an elevated concentration of the NR2C subunit in the LC from major depressive subjects as compared to normal control subjects (Karolewicz et al., 2005). These findings imply that altered subunit composition of the NMDA receptor in the LC may contribute to disruption of noradrenergic transmission in psychiatric illnesses.
There have been no previous studies of NMDA receptor subunits and its associated proteins in postmortem tissue of alcohol dependent subjects. Rather, investigators have investigated the effects of chronic exposure of rodents to ethanol on the expression of NMDA receptor polypeptides (for review see Kumari and Ticku, 2000). Therefore, the present study was designed to determine whether alcoholism is associated with abnormalities of NMDA receptor subunits in the LC. In addition, signaling molecules linked to NMDA receptor activation were also evaluated. At glutamatergic synapses, NMDA receptors are stabilized through interactions with postsynaptic density protein-95 (PSD-95). PSD-95 immobilizes the NMDA receptor and serves as a scaffold for the assembly of macromolecular complexes that modulate the receptor’s transduction pathways, including it’s coupling to neuronal nitric oxide synthase, nNOS (Brenman et al., 1996; Kornau et al., 1995). Concentrations of NR1 and NR2C subunits as well as PSD-95 and nNOS were measured in LC tissue from alcohol dependent and healthy control subjects using Western blotting. In addition, tyrosine hydroxylase (TH), the rate limiting enzyme in the synthesis of norepinephrine, was measured in the same study subjects.
2. Methods
2.1. Human subjects
Brain tissue was obtained at autopsy at the Coroner's Office of Cuyahoga County, Cleveland, OH, USA. An ethical protocol approved by the Institutional Review Boards at Case Western Reserve University and the University of Mississippi Medical Center was used and informed written consent was obtained from the next of kin for all subjects. Retrospective, informant-based psychiatric assessments were performed for all alcohol dependent subjects and healthy controls. A trained interviewer administered the Schedule for Affective Disorders and Schizophrenia: lifetime version (SADS-L) to knowledgeable next of kin of 4 alcoholics. The Structured Clinical Interview for DSM-IV Psychiatric Disorders (SCID-IV) was administered to next of kin of the 6 remaining alcohol dependent subjects. Blood and urine samples from all subjects were examined by coroner’s office for psychotropic medications and substances of abuse, including ethanol. Control subjects (n=8) did not meet criteria for any Axis I disorder at the time of death, except for nicotine dependence as noted (Table 1). Based on the next of kin assessment, all alcohol dependent subjects (n=10) were described as long-term “heavy” drinkers. The onset age of drinking was reported to occur between the teen years and the 20s for almost all alcohol dependent subjects. Nine of the 10 alcohol dependent subjects were consuming large quantities of alcohol daily until death. Available data indicate that only one alcohol dependent subject was abstaining from ethanol for 3 weeks before death. Only 2 out of the ten subjects had historical information indicating signs of withdrawal (sweating, tremors) when unsuccessfully attempting to discontinue their habitual drinking. Subjects were matched across groups as closely as possible for age, gender, postmortem interval, smoking and tissue pH (Table 1). The paired design was maintained throughout all experimental procedures. Additionally, body mass index (BMI) was computed as the ratio between body weight (kg) and height (m2).
Table 1.
Demographics for subjects used in the study.
| Parameter | Controls (n=8) | Alcoholics (n=10) |
|---|---|---|
| Age range | 27 to 58 y | 42 to 64 y |
| Age (mean±SEM) | 44 ± 4 y | 50 ± 3 y |
| PMIa range | 9 to 32 h | 14 to 30 h |
| PMI (mean±SEM) | 23 ± 2 | 22 ± 2 |
| pH range | 5.95 to 6.87 | 5.82 to 6.95 |
| pH (mean±SEM) | 6.60 ± 0.11 | 6.65 ± 0.11 |
| Genderb (F:M) | 4:4 | 2:8 |
| Toxicology | Cocaine metabolites, n=1; NDDc, n=7 | Lidocaine, n=2; Codeine, n=1; Ethanol, n=5; NDD, n=2 |
| Natural (heart disease, n=7) | Natural (heart disease, n=6; pneumonia, n=2) | |
| Cause of death | Accidental (lightning, n=1) | Accidental (carbon monoxide poisoning, n=1) |
| Suicide (carbon monoxide poisoning, n=1) | ||
| Smokers | n=7 | n=9 |
PMI, postmortem interval
F, female; M, male
NDD, no drugs detected
2.2. Dissection and anatomical positioning of measurements
LC tissue blocks were sectioned according to a method published previously (Karolewicz et al., 2004; 2005). Frozen tissue blocks were cut along the entire length of the LC, with histological sections taken at 1 mm intervals to evaluate anatomical position along the LC axis. The exact location of the rostral and caudal end of the LC was individualized for each subject based on cresyl violet (Nissl) staining and subsequent cell counting. The LC had its rostral border defined as a point where at least 25 ± 5 neuromelanin-containing cells were identified. The caudal border was defined near the caudal end at a point where 25 ± 5 or less neuromelanin-containing cells were present. The whole LC was punched into 50 µm thick sections. Tissue punches were collected in centrifuge tubes and stored at −80° C until assayed.
For each anatomical level of the LC, tissue was collected from 2 mm of sections that were each centered at points that represented 25%, 50%, and 75% of the total rostro-caudal length of the LC. Semi-quantitative estimation of neuromelanin-containing cell profiles was performed using cresyl violet stained sections (20 µm) cut at 1 mm intervals along the LC. Counts are reported as the average of the left and right LC (Figure 4).
Figure 4.
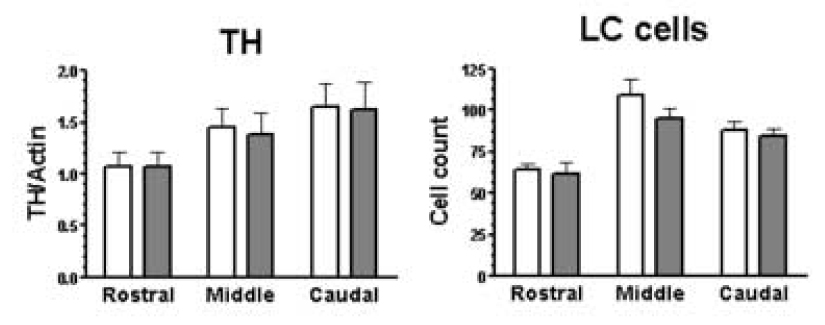
Amount of TH immunoreactivity and neuromelanin containing cell numbers from control subjects (open bars; n=8) and alcohol dependent subjects (filled bars; n=10). Immunoreactivities and LC cell numbers were estimated at the rostral, middle and caudal portions of the LC.
2.3. Immunoblotting
LC tissue samples were prepared according to a method published previously (Karolewicz et al., 2005). Samples were homogenized in ice-cold TE buffer (10 mM Tris-HCl and 1 mM ethylene-diaminetetraacetate, EDTA) containing protease inhibitors (Protease Inhibitor Cocktail Tablets - Complete™, Boehringer Mannheim GmbH). Total protein concentrations for all samples were determined using the bicinchoninic acid (BCA) method (#23227; Pierce). Samples were mixed with sample buffer (125 mM Tris base, 20% glycerol, 4% SDS, 10% mercaptoethanol, 0.05% bromophenol blue, pH 6.8) and heated at 95°C for 8 min. Solubilized protein (20 µg per lane) was subjected to 10% Criterion Precast Tris-HCl gel electrophoresis (#345-0009; Bio-Rad Laboratories) and transferred to nitrocellulose membranes (Hybond ECL; #RPN303D; Amersham Biosciences). After transfer, blots were blocked in 5% non-fat milk/TBS (20 mM Tris base and 0.5 M NaCl, pH 7.5) or 5% non-fat milk/PBS (137 mM NaCl, 2.7 mM KCl, 8 mM Na2HPO4 and KH2PO4, pH 7.4) for 2 h, then incubated (overnight at 4° C) with mouse anti-NR1 monoclonal antibodies diluted 1:1000 (# 556308; Pharmingen/BD Biosciences). NR2C subunit was labeled using rabbit polyclonal antibodies diluted 1:500 (# OPA1-04020; Affinity BioReagents). PSD-95 was labeled using a mouse monoclonal antibody diluted 1:1000 (# 05-494; Upstate Cell Signaling). nNOS was labeled using mouse monoclonal antibodies diluted 1:1000 (# 610309; BD Transduction Laboratories/BD Biosciences). TH was labeled using a mouse monoclonal antibody diluted 1:1000 (# MAB318; Chemicon International). Membranes were washed three times for 10 min in TBS (or PBS) buffer and incubated with secondary anti-mouse antibody for NR1, PSD-95, nNOS, TH (diluted 1:2000; # NA931; Amersham Biosciences) and anti-rabbit secondary antibody for NR2C (diluted 1:3000; # NA934; Amersham Biosciences). After incubation, blots were washed 3 times for 15 min and developed using enhanced chemiluminescence detection (ECL; #NEL104; PerkinElmer) and immediately exposed to film (Hyperfilm-ECL; #RPN3103K; Amersham Biosciences). As a control for transfer and loading, actin was detected on each blot using an anti-actin mouse monoclonal antibody diluted 1:5000 (# MAB1501; Chemicon International). Immunoreactivity of NR1, NR2C, PSD-95, nNOS and TH was investigated in pairs of the same alcohol dependent and matched control subjects. Pairs of subjects were immunoblotted on the same gel with duplicates on the separate gels. All results were generated from the rostral, middle and caudal portions of the LC. Each gel was loaded with 3 concentrations of LC standard (dissected from a healthy control subject) consisting of 10, 20, and 40 µg of total protein.
2.4. Relationship between the optical density and the total protein concentration
In order to determine the relationship between optical density values and the concentrations of protein immunoreactivities, 10, 20, and 40 µg of total LC protein (LC standard) was immunoblotted with anti-NR1, anti-NR2C, anti-PSD-95, anti-nNOS, anti-TH, and anti-actin antibodies (Figure 2). Optical density values of immunoreactive bands were measured and are presented as a function of total LC protein concentration (expressed in µg). Analyses of blots revealed an approximately 1:1 relationship between changes in optical density values and changes in protein concentrations. That is, a 100% increase in total protein loading resulted in an approximately 100% increase in the optical density of all protein immunoreactive bands (Figure 1).
Figure 2.
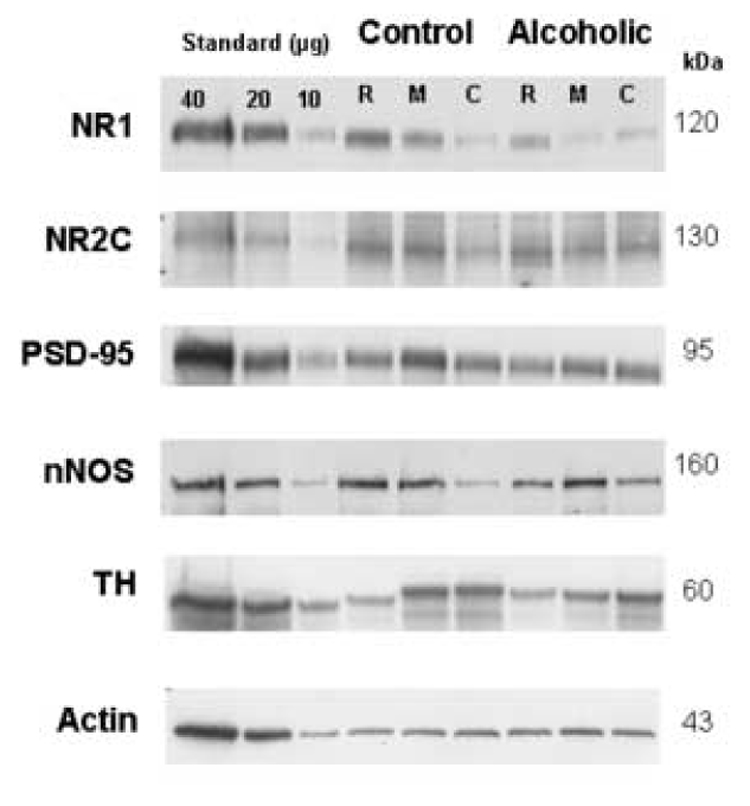
Immunoblots of NR1, NR2C, PSD-95, nNOS, TH and actin from a representative pair of control/alcoholic subjects used in the analysis. Shown are blots from three separate anatomical levels of the LC: rostral (R), middle (M), and caudal (C). Each well was loaded with 20 µg of total protein. Additionally, each gel was loaded with three concentrations of LC standard (dissected from a healthy control subject) consisting of 10–40 µg of total protein.
Figure 1.
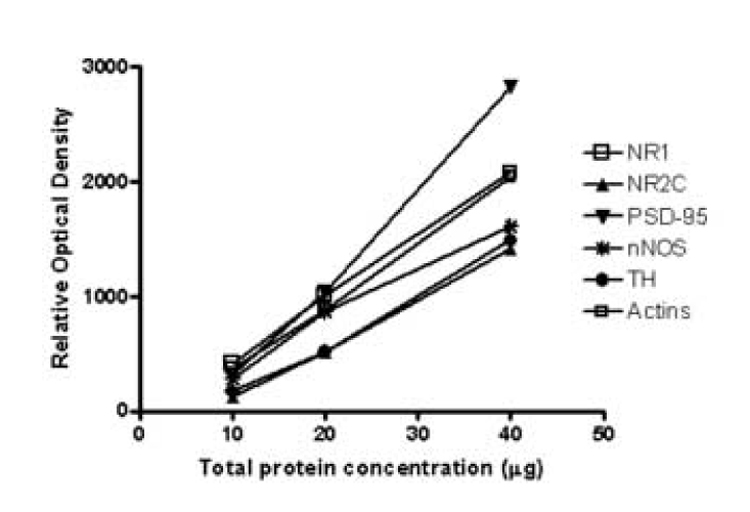
Relationship between the optical density values of NR1, NR2C, PSD-95, nNOS, TH, actin and total protein concentration in the human LC. Wells were loaded with 10, 20, and 40 µg of total human LC protein.
2.5. Statistical analysis
Band densities were analyzed using MCID Elite 7.0 (Imaging Research, St. Catherines, Ontario, Canada). Relative optical density values were converted to arbitrary protein units using the standards, normalizing with actin immunoreactivity on the same gel. Data from each set of experiments, i.e. for each protein measured, were analyzed using one-way ANOVA followed by Bonferroni post tests for regional differences (GraphPad Prism 4.0, GraphPad Software Incorporated, San Diego, CA). Summary statistics are reported as the mean ± S.E.M. Linear regression analyses were used to compute potential correlations between immunoreactivities and potentially confounding factors. A p value < 0.05 was considered significant.
3. Results
For NMDA receptor subunits, immunoreactive bands corresponding to molecular masses of ~120 and ~130 kDa for NR1 and NR2C, respectively, were observed (Figure 2). Amounts of NR1-immunoreactivity from alcohol dependent subjects were significantly lower than those of control subjects (F5,54 = 9.28, p<0.0001; Figure 3). Post hoc comparisons revealed that amounts of NR1 immunoreactivity were significantly lower at the rostral and middle LC (p<0.01) from alcoholics relative to control subjects. In contrast, amounts of NR2C immunoreactivity from alcohol dependent subjects were not significantly different from that of controls (Figure 3). PSD-95 and nNOS immunoreactivities appeared as bands with molecular masses of ~95 and ~160 kDa, respectively, and amounts of these proteins in the LC of alcoholic subjects were not significantly different from those of control subjects (Figure 2 and 3). TH immunoreactivity appeared as a band at ~60 kDa and amounts of this protein were normal in alcoholics as compared to controls (Figure 2 and 4). The number of neuromelanin-containing cell profiles counted at the 3 anatomical levels of the LC was not significantly different comparing alcoholic to control subjects (Figure 4).
Figure 3.

Amounts of NR1, NR2C, PSD-95, and nNOS immunoreactivities in the LC from control subjects (open bars; n=8) and alcohol dependent subjects (filled bars; n=10). Immunoreactivities were measured at the rostral, middle and caudal portions of the LC. Asterisks indicate statistical significance (p<0.01) Comparing amounts of immunoreactivity in alcoholic subjects to control subjects at the same anatomical level.
Ages, postmortem intervals, and brain pH values were not significantly correlated with amounts of NR1 in control or alcohol dependent subjects. Similarly, effects of age, postmortem interval, or pH on NR2C, PSD-95, nNOS and TH were not detected. It is worth noting that one alcohol dependent subject had committed suicide. This subject had an amount of NR1 immunoreactivity near average of immunoreactivity of all alcohol dependent subjects dying of natural causes. The average body mass index (BMI) values of alcoholics (28.12 ± 2) and controls (30.6 ± 2) were not significantly different and ranged from 15 to 40 in controls and 19 to 38 in alcoholics (Figure 5).
Figure 5.

Body mass index (BMI) in psychiatrically normal control subjects (open bars, n=8) and in subjects diagnosed with alcohol dependence (filled bars; n=10). Each bar represents the mean ± SEM. There was no significant difference in BMI comparing alcoholics to control subjects.
4. Discussion
To the best of our knowledge, the present study is the first to analyze quantities of glutamate signaling proteins in postmortem tissue from alcohol dependent subjects. The present study provides evidence that the amount of the NR1 subunit immunoreactivity is significantly lower in the LC of alcoholics as compared to healthy controls. In contrast, overall amounts of NR2C, PSD-95, nNOS and TH were unchanged in the LC of the same alcohol dependent subjects. Thus, the low level of NR1 subunit was unique among the 5 proteins studied. Based on the retrospective interviews with next of kin (see Methods) only one of the 10 alcohol dependent subjects was abstaining from ethanol for 3 weeks before death. Nine of the ten alcoholics were consuming large quantities of ethanol daily until death. Moreover, toxicology screening demonstrated the presence of blood and urine ethanol in half of the investigated alcoholics (Table 1). Therefore, abnormalities in NR1 subunit are likely related to ethanol exposure, rather than to ethanol withdrawal.
To date, few studies have investigated NMDA receptors in postmortem brain from alcohol dependent subjects. Radioligand binding to NMDA receptors has been shown to be unchanged (Dodd et al., 1992; Freund and Anderson, 1999), modestly increased (Freund and Anderson, 1996), or reduced (Cummins et al., 1990; Michaelis et al., 1990) in brains from alcoholics. However, there has been no previous study of NMDA receptor subunits in postmortem tissue from alcohol dependent subjects. Rather, investigators have investigated the effects of chronic exposure of rodents to ethanol on the expression of subunits of the NMDA receptor. Trevisan et al., (1994) observed up-regulation of NR1 protein in rats exposed to ethanol in the hippocampus but not in the cortical region, nucleus accumbens or striatum. Snell et al., (1996) found increased NR1 protein, but not mRNA, in hippocampus and cerebellum after chronic ethanol administration to mice. In contrast, unchanged levels of NR1 subunit mRNA in cortex, hippocampus and cerebellum has been reported in rats chronically treated with ethanol (Follesa and Ticku, 1995). Overall, these rodent findings are not consistent with the present observations in postmortem human tissue. This discrepancy between animal and human studies may be a result of differences in the duration of ethanol exposure. The average age of alcohol dependent subjects used in the present study is 50 ± 3 y (Table 1). The onset age of drinking was reported to occur between the teen years and the 20s for alcohol dependent subjects (see 2.1. Human subjects). Therefore it is estimated that the average duration of drinking was 20–30 years. Interestingly, a minimum of 6 days of chronic ethanol treatment in adult rats appeared to be sufficient to observe an up-regulation of NR1 and NR2A-B polypeptide levels in hippocampus and cerebral cortex (Kalluri et al., 1998; Snell et al., 1996). Another obvious difference between rodent and human studies is the lack of dietary control in human studies. Poor diet in alcoholics may contribute to alcohol-induced neuropathological changes. However, body mass index (BMI), which provides evidence of the nutritional status, was not statistically different in alcohol dependent subjects as compared to controls (Figure 5). It is worth noting that both groups of subjects had the BMI values above 25 (ideal BMI). This weight data, albeit limited to the time of death, suggests that malnutrition is not a factor likely to be contributing to altered NMDA receptor subunit composition.
Recent studies demonstrate that ethanol has complex regulatory effects on splice variants of the NR1 subunit. The NR1 subunit has 8 splice variants that are generated by alternative splicing of single gene exons: 5, 21, and 22 (Zukin and Bennet, 1995). Darstein et al., (2000) and Hardy et al., (1999) observed reductions in specific splice variants of NR1 mRNA in ethanol-treated rats during ethanol exposure and after withdrawal, while total amounts of NR1 mRNA were unchanged. In addition, chronic exposure of fetal mouse cortical neurons to ethanol increases expression of splice variants that lack exon 5 and exon 22, but decreased expression of splice variants that contain exon 5 (e.g. NR1-3b and NR1-4b). Hence, the effect of ethanol on NR1 subunits is splice variant-specific. In the present study of alcohol dependent subjects, the NR1 subunit was measured using an antibody raised against a recombinant fusion protein containing amino acids 660 to 811, and this antibody does not discriminate the 8 splice variants of NR1 (Siegel et al., 1994). Hence, it is possible that even more robust effects of ethanol on NR1 could be found if splice variant-specific antibodies were used.
Based on in vivo and in vitro studies, acute administration of ethanol inhibits the function of glutamatergic NMDA receptors in regionally specific manner. In contrast, sustained ethanol administration is associated with 1) increased NMDA receptor function, 2) increased ligand binding, and 3) elevated levels of protein and/or mRNA for some NMDA receptor subunits in hippocampus and cerebral cortex (for review see Hoffman, 2003; Krystal et al., 2003; Kumari and Ticku, 2000). However, no previous study has examined the effect of ethanol administration on NMDA receptor expression or function in the LC.
Laboratory animal studies have shown that NR2A and NR2B, as opposed to NR2C or NR2D, are more likely to be regulated by chronic ethanol exposure. NMDA receptors containing NR2C or NR2D subunits are less sensitive to ethanol-induced inhibition than are those containing NR2A or NR2B subunits (Chu et al., 1995) suggesting that functional sensitivity to ethanol may translate to sensitivity to regulation by chronic exposure to ethanol. Interestingly, among all 32 combinations of NR1/NR2 subunits, the lowest degree of inhibition by ethanol was observed for NR2C and NR2D receptors containing NR1-3b or NR1-4b subunits, on the other hand highest degree of inhibition was observed for the NR1-2b/NR2C receptors (Jin and Woodward, 2006). The functional and pharmacological properties of NR1/NR2C receptors may depend on NR1 splice variant present and may show marked region-specific differences. Given that various NR1 splice variants convey differential sensitivity to ethanol, further study of alcoholism using specific antibodies for protein products of splice variants of NR1 mRNA in the human LC is warranted.
At excitatory synapses, NMDA receptors are organized into multiprotein signaling complexes. A prominent scaffolding and anchoring protein is PSD-95 which couples the NMDA to intracellular proteins and signaling enzymes (Kornau et al. 1995). PSD-95 interacts with the C terminus of NR2 subunits, but not with NR1 subunits (Lau et al. 1996). As a result, PSD-95 enhances NMDAR clustering at synapses and inhibits NR2-mediated internalization. In the present study, no differences in the amount of PSD-95 immunoreactivity were observed in alcoholic compared to control subjects, just as there were no differences in NR2C subunit immunoreactivity. Presently, there are no previous experimental studies on the effect of chronic ethanol exposure on PSD-95 levels in the LC. Previously, evidence has been provided that ethanol withdrawal leads to bi-directional and sex-selective effects on PSD-95 (Alele and Devaud, 2005). PSD-95 levels were significantly increased in the female rat cerebral cortex and decreased in the hippocampus by ethanol. This is in contrast to a study reporting that chronic ethanol administration to neuronal cultures did not alter PSD-95 levels (Chandler et al., 1999).
The synaptic expression of NR1 subunits, through their C termini, is stabilized by interactions with calmodulin, alpha-actinin-2, Yotiao or neurofilamin-light (Ehlers et al., 1996; Lin et al., 1998; Ratnam and Teichberg, 2005; Wyszynski et al., 1997). Examination of these latter proteins may provide further insight into the pathology of NR1 signaling machinery in the LC of alcoholics.
In the CNS, nitric oxide (NO) is predominantly produced by neuronal nitric oxide synthase (nNOS) and nNOS activity is strongly linked to the NMDA receptor pathway. Studies in rodents have demonstrated that NO and the nNOS gene are critically involved in the regulation of neurobehavioral effects of ethanol (Rezvani et al., 1995; Spanagel at al., 2002). However, in the present study nNOS protein was found unchanged in the LC from alcoholics. A previous postmortem study has reported that nNOS protein content was significantly increased in the limbic brain regions such as frontal cortex and nucleus accumbens, but not in the LC in alcohol dependent subjects (Gerlach et al., 2001). Presently, there are no previous experimental studies on the effect of chronic ethanol exposure on nNOS protein levels in the LC. nNOS activity has been measured in rat frontal cortex, hippocampus, striatum and cerebellum following chronic alcohol exposure (Naassila et al., 1997). The Km and Vmax values were significantly increased in the frontal cortex and in the striatum, and were not affected in the cerebellum and hippocampus. In contrast, nNOS activity in the various brain regions of mice remained normal after acute and chronic administration of ethanol (Ikeda et al., 1999).
Based on reports of reductions in neuron numbers in the LC of alcoholics (Arango et al., 1994) and in rats exposed to ethanol (Lu et al., 1997), it seemed possible that TH levels and LC neuron numbers would be low in alcoholics. Instead, we found normal levels of TH and neuromelanin-containing neurons in all investigated levels of the LC (Figure 2). These findings are consistent with other postmortem studies reporting no loss of pigmented neurons in alcoholics (Halliday et al., 1992), and unchanged TH immunoreactivity or TH activity in the LC of alcoholics (Baker et al., 1994; Tabakoff et al., 1985).
One of the shortcomings of this study is that NR1 protein immunoreactivity was measured in postmortem tissue homogenates as opposed to measuring functional NR1 subunit protein that is expressed on the surface of the plasma membrane. Whether the decrease in the total cellular NR1 protein immunoreactivity is positively correlated with a decrease in the number of functional subunits expressed on the cell surface cannot be answered based on the present experiments. Considering that NR1 is an essential subunit of NMDA receptors, it seems possible that a loss of NR1 subunit may result in a lower number of functional NMDA receptors. NR1 subunits are synthesized in a large excess (~10-fold) when compared with NR2 subunits (Huh and Wenthold 1999). The excess production of NR1 optimizes the chance that NR2 subunits will find NR1 partners. Unassembled NR1 subunits with a half-life of approximately 2 h are retained in the endoplasmic reticulum (ER) and degraded. Therefore, there is a possibility that lower total NR1 protein would indicate increased rate of NR1 protein degradation in the ER.
Another unresolved issue is the cellular localization of the change of NR1 subunits within the region of the LC. The LC was dissected with a 5 mm bore punch centered over the cellular region of the LC. Hence, noradrenergic cells within the LC were collected, as were glia and terminals of neurons that project to the LC, including serotonergic and GABAergic neurons (Aston-Jones et al., 1991).
The present findings indicate that amounts of NR1 protein are low in the LC of alcoholic subjects. A likely causal factor for this neuropathological finding is decades of ethanol exposure, although improper diet, metabolic abnormalities secondary to liver dysfunction, and other issues that associate with alcoholism may also contribute. The possibility that reduced NR1 could be due to alcohol-related structural pathology in the LC seems unlikely based on findings of unchanged LC cell number as well as normal levels of NR2C, PSD-95, nNOS and TH in the same alcohol dependent subjects. Finally, it cannot be ruled out that low NR1 subunit protein in the LC is a predisposing factor to the development of alcoholism, rather than an effect of ethanol exposure.
Acknowledgements
The authors thank Dr. Maria Soledad Romero for technical assistance in the preparation of proteins for assays. We gratefully acknowledge the work of Drs James C. Overholser, Herbert Y. Meltzer, Bryan L. Roth, George Jurjus, Ginny Dilley, Lisa Konick, and Lesa Dieter in the retrospective psychiatric diagnoses. The excellent assistance of the Cuyahoga County Coroner’s Office, Cleveland, OH is greatly appreciated. This publication was made possible by Grant Number RR17701 from National Center for Research Resources (NCRR), a component of the National Institutes of Health (NIH) and MH63187, MH46692, MH02031.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Alele PE, Devaud LL. Differential adaptations in GABAergic and glutamatergic systems during ethanol withdrawal in male and female rats. Alcoholism: Clinical and Experimental Research. 2005;29:1027–1034. doi: 10.1097/01.alc.0000167743.96121.40. [DOI] [PubMed] [Google Scholar]
- Arango V, Underwood MD, Mann JJ. Fewer pigmented neurons in the locus coeruleus of uncomplicated alcoholics. Brain Research. 1994;650:1–8. doi: 10.1016/0006-8993(94)90199-6. [DOI] [PubMed] [Google Scholar]
- Aston-Jones G, Foote SL, Bloom FE. Low doses of ethanol disrupt sensory responses of brain noradrenergic neurones. Nature. 1982;296:857–860. doi: 10.1038/296857a0. [DOI] [PubMed] [Google Scholar]
- Aston-Jones G, Shipley MT, Chouvet G, Ennis M, van Bockstaele E, Pieribone V, Shiekhattar R, Akaoka H, Drolet G, Astier B, et al. Afferent regulation of locus coeruleus neurons: anatomy, physiology and pharmacology. Progress in Brain Research. 1991;88:47–75. doi: 10.1016/s0079-6123(08)63799-1. [DOI] [PubMed] [Google Scholar]
- Baker KG, Halliday GM, Harper CG. Effect of chronic alcohol consumption on the human locus coeruleus. Alcoholism: Clinical and Experimental Research. 1994;18:1491–1496. doi: 10.1111/j.1530-0277.1994.tb01455.x. [DOI] [PubMed] [Google Scholar]
- Brenman JE, Chao DS, Gee SH, McGee AW, Craven SE, Santillano DR, Wu Z, Huang F, Xia H, Peters MF, Froehner SC, Bredt DS. Interaction of nitric oxide synthase with the postsynaptic density protein PSD-95 and alpha1-syntrophin mediated by PDZ domains. Cell. 1996;84:757–767. doi: 10.1016/s0092-8674(00)81053-3. [DOI] [PubMed] [Google Scholar]
- Chandler LJ, Norwood D, Sutton G. Chronic ethanol upregulates NMDA and AMPA, but not kainate receptor subunit proteins in rat primary cortical cultures. Alcoholism: Clinical and Experimental Research. 1999;23:363–370. [PubMed] [Google Scholar]
- Chu B, Anantharam V, Treistman SN. Ethanol inhibition of recombinant heteromeric NMDA channels in the presence and absence of modulators. Journal of Neurochemistry. 1995;65:140–148. doi: 10.1046/j.1471-4159.1995.65010140.x. [DOI] [PubMed] [Google Scholar]
- Cummins JT, Sack M, von Hungen K. The effect of chronic ethanol on glutamate binding in human and rat brain. Life Science. 1990;47:877–882. doi: 10.1016/0024-3205(90)90601-m. [DOI] [PubMed] [Google Scholar]
- Darstein MB, Landwehrmeyer GB, Feuerstein TJ. Changes in NMDA receptor subunit gene expression in the rat brain following withdrawal from forced long-term ethanol intake. Naunyn Schmiedeberg’s Archives of Pharmacology. 2000;361:206–213. doi: 10.1007/s002109900180. [DOI] [PubMed] [Google Scholar]
- Dodd PR, Thomas GJ, Harper CG, Kril JJ. Amino acid neurotransmitter receptor changes in cerebral cortex in alcoholism: effect of cirrhosis of the liver. Journal of Neurochemistry. 1992;59:1506–1515. doi: 10.1111/j.1471-4159.1992.tb08467.x. [DOI] [PubMed] [Google Scholar]
- Ehlers MD, Zhang S, Bernhadt JP, Huganir RL. Inactivation of NMDA receptors by direct interaction of calmodulin with the NR1 subunit. Cell. 1996;84:745–755. doi: 10.1016/s0092-8674(00)81052-1. [DOI] [PubMed] [Google Scholar]
- Ennis M, Aston-Jones G. A potent excitatory input to the nucleus locus coeruleus from the ventrolateral medulla. Neuroscience Letters. 1986;71:299–305. doi: 10.1016/0304-3940(86)90637-3. [DOI] [PubMed] [Google Scholar]
- Follesa P, Ticku MK. Chronic ethanol treatment differentially regulates NMDA receptor subunit mRNA expression in rat brain. Brain Research Molecular Brain Research. 1995;29:99–106. doi: 10.1016/0169-328x(94)00235-7. [DOI] [PubMed] [Google Scholar]
- Freund G, Anderson KJ. Glutamate receptors in the frontal cortex of alcoholics. Alcoholism: Clinical and Experimental Research. 1996;20:1165–1172. doi: 10.1111/j.1530-0277.1996.tb01106.x. [DOI] [PubMed] [Google Scholar]
- Freund G, Anderson KJ. Glutamate receptors in the cingulate cortex, hippocampus, and cerebellar vermis of alcoholics. Alcoholism: Clinical and Experimental Research. 1999;23:1–6. [PubMed] [Google Scholar]
- Gerlach M, Blum-Degen D, Ransmayr G, Leblhuber F, Pedersen V, Riederer P. Expression, but not activity, of neuronal nitric oxide synthase is regionally increased in the alcoholic brain. Alcohol and Alcoholism. 2001;36:65–69. doi: 10.1093/alcalc/36.1.65. [DOI] [PubMed] [Google Scholar]
- Grant KA, Lovinger DM. Cellular and behavioral neurobiology of alcohol: receptor-mediated neuronal processes. Clinical Neuroscience. 1995;3:155–164. [PubMed] [Google Scholar]
- Halliday G, Ellis J, Harper C. The locus coeruleus and memory: a study of chronic alcoholics with and without the memory impairment of Korsakoff's psychosis. Brain Research. 1992;598:33–37. doi: 10.1016/0006-8993(92)90164-5. [DOI] [PubMed] [Google Scholar]
- Hardy PA, Chen W, Wilce PA. Chronic ethanol exposure and withdrawal influence NMDA receptor subunit and splice variant mRNA expression in the rat cerebral cortex. Brain Research. 1999;819:33–39. doi: 10.1016/s0006-8993(98)01340-7. [DOI] [PubMed] [Google Scholar]
- Hoffman PL. NMDA receptors in alcoholism. International Review of Neurobiology. 2003;56:35–82. doi: 10.1016/s0074-7742(03)56002-0. [DOI] [PubMed] [Google Scholar]
- Huh KH, Wenthold RJ. Turnover analysis of glutamate receptors identifies a rapidly degraded pool of the N-methyl-D-aspartate receptor subunit, NR1, in cultured cerebellar granule cells. Journal of Biological Chemistry. 1999;274:151–157. doi: 10.1074/jbc.274.1.151. [DOI] [PubMed] [Google Scholar]
- Ikeda M, Komiyama T, Sato I, Himi T, Murota S. Neuronal nitric oxide synthase is resistant to ethanol. Life Science. 1999;64:1623–1630. doi: 10.1016/s0024-3205(99)00099-5. [DOI] [PubMed] [Google Scholar]
- Ishii T, Moriyoshi K, Sugihara H, Sakurada K, Kadotani H, Yokoi M, Akazawa C, Shigemoto R, Mizuno N, Masu M, et al. Molecular characterization of the family of the N-methyl-D-aspartate receptor subunits. Journal of Biological Chemistry. 1993;268:2836–2843. [PubMed] [Google Scholar]
- Jin C, Woodward JJ. Effects of 8 different NR1 splice variants on the ethanol inhibition of recombinant NMDA receptors. Alcoholism: Clinical and Experimental Research. 2006;30:673–679. doi: 10.1111/j.1530-0277.2006.00079.x. [DOI] [PubMed] [Google Scholar]
- Chiang C, Aston-Jones G. Potent excitatory influence of prefrontal cortex activity on noradrenergic locus coeruleus neurons. Neuroscience. 1998;83:63–79. doi: 10.1016/s0306-4522(97)00372-2. [DOI] [PubMed] [Google Scholar]
- Kalluri HS, Mehta AK, Ticku MK. Up-regulation of NMDA receptor subunits in rat brain following chronic ethanol treatment. Brain Research Molecular Brain Research. 1998;58:221–224. doi: 10.1016/s0169-328x(98)00112-0. [DOI] [PubMed] [Google Scholar]
- Karolewicz B, Stockmeier CA, Ordway GA. Elevated Levels of the NR2C Subunit of the NMDA Receptor in the Locus Coeruleus in Depression. Neuropsychopharmacology. 2005;30:1557–1567. doi: 10.1038/sj.npp.1300781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karolewicz B, Szebeni K, Stockmeier CA, Konick L, Overholser JC, Jurjus G, Roth BL, Ordway GA. Low nNOS protein in the locus coeruleus in major depression. Journal of Neurochemistry. 2004;91:1057–1066. doi: 10.1111/j.1471-4159.2004.02792.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kornau HC, Schenker LT, Kennedy MB, Seeburg PH. Domain interaction between NMDA receptor subunits and the postsynaptic density protein PSD-95. Science. 1995;269:1737–1740. doi: 10.1126/science.7569905. [DOI] [PubMed] [Google Scholar]
- Krystal JH, Petrakis IL, Mason G, Trevisan L, D'Souza DC. N-methyl-D-aspartate glutamate receptors and alcoholism: reward, dependence, treatment, and vulnerability. Pharmacology &. Therapeutics. 2003;99:79–94. doi: 10.1016/s0163-7258(03)00054-8. [DOI] [PubMed] [Google Scholar]
- Kumari M, Ticku MK. Regulation of NMDA receptors by ethanol. Progress in Drug research. 2000;54:152–189. [PubMed] [Google Scholar]
- Kutsuwada T, Kashiwabuchi N, Mori H, Sakimura K, Kushiya E, Araki K, Meguro H, Masaki H, Kumanishi T, Arakawa M, et al. Molecular diversity of the NMDA receptor channel. Nature. 1992;358:36–41. doi: 10.1038/358036a0. [DOI] [PubMed] [Google Scholar]
- Lau LF, Mammen A, Ehlers MD, Kindler S, Chung WJ, Garner CC, Huganir RL. Interaction of the N-methyl-D-aspartate receptor complex with a novel synapse-associated protein, SAP102. Journal of Biological Chemistry. 1996;271:21622–21628. doi: 10.1074/jbc.271.35.21622. [DOI] [PubMed] [Google Scholar]
- Laurie DJ, Seeburg PH. Ligand affinities at recombinant N-methyl-D-aspartate receptors depend on subunit composition. European Journal of Pharmacology. 1994;268:335–345. doi: 10.1016/0922-4106(94)90058-2. [DOI] [PubMed] [Google Scholar]
- Lin JW, Wyszynski M, Madhavan R, Sealock R, Kim JU, Sheng M. Yotiao a novel protein of neuromuscular junction and brain that interacts with specific splice variants of NMDA receptor subunit NR1. Journal of Neuroscience. 1998;18:2017–2027. doi: 10.1523/JNEUROSCI.18-06-02017.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu W, Jaatinen P, Rintala J, Sarviharju M, Kiianmaa K, Hervonen A. Effects of lifelong ethanol consumption on rat locus coeruleus. Alcohol and Alcoholism. 1997;32:463–470. doi: 10.1093/oxfordjournals.alcalc.a008281. [DOI] [PubMed] [Google Scholar]
- Michaelis EK, Freed WJ, Galton N, Foye J, Michaelis ML, Phillips I, Kleinman JE. Glutamate receptor changes in brain synaptic membranes from human alcoholics. Neurochemistry Research. 1990;15:1055–1063. doi: 10.1007/BF01101704. [DOI] [PubMed] [Google Scholar]
- Monyer H, Burnashev N, Laurie DJ, Sakmann B, Seeburg PH. Developmental and regional expression in the rat brain and functional properties of four NMDA receptors. Neuron. 1994;12:529–540. doi: 10.1016/0896-6273(94)90210-0. [DOI] [PubMed] [Google Scholar]
- Naassila M, Beauge F, Daoust M. Regulation of rat neuronal nitric oxide synthase activity by chronic alcoholization. Alcohol & Alcoholism. 1997;32:13–17. doi: 10.1093/oxfordjournals.alcalc.a008228. [DOI] [PubMed] [Google Scholar]
- Ratnam J, Teichberg VI. Neurofilament-light increases the cell surface expression of the N-methyl-D-aspartate receptor and prevents its ubiquitination. Journal of Neurochemistry. 2005;92:878–885. doi: 10.1111/j.1471-4159.2004.02936.x. [DOI] [PubMed] [Google Scholar]
- Rezvani AH, Grady DR, Peek AE, Pucilowski O. Inhibition of nitric oxide synthesis attenuates alcohol consumption in two strains of alcohol-preferring rats. Pharmacology Biochemistry & Behavior. 1995;50:265–270. doi: 10.1016/0091-3057(94)00310-f. [DOI] [PubMed] [Google Scholar]
- Shefner SA, Tabakoff B. Basal firing rate of rat locus coeruleus neurons affects sensitivity to ethanol. Alcohol. 1985;2:239–243. doi: 10.1016/0741-8329(85)90052-7. [DOI] [PubMed] [Google Scholar]
- Siegel SJ, Brose N, Janssen WG, Gasic GP, Jahn R, Heinemann SF, Morrison JH. Regional, cellular, and ultrastructural distribution of N-methyl-D-aspartate Receptor subunit 1 in monkey hippocampus. Proceedings of the National Academy of Sciences of the United States of America. 1994;91:564–568. doi: 10.1073/pnas.91.2.564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Snell LD, Nunley KR, Lickteig RL, Browning MD, Tabakoff B, Hoffman PL. Regional and subunit specific changes in NMDA receptor mRNA and immunoreactivity in mouse brain following chronic ethanol ingestion. Brain Research Molecular Brain Research. 1996;140:71–78. doi: 10.1016/0169-328x(96)00038-1. [DOI] [PubMed] [Google Scholar]
- Spanagel R, Siegmund S, Cowen M, Schroff KC, Schumann G, Fiserova M, Sillaber I, Wellek S, Singer M, Putzke J. The neuronal nitric oxide synthase gene is critically involved in neurobehavioral effects of alcohol. Journal of Neuroscience. 2002;22:8676–8683. doi: 10.1523/JNEUROSCI.22-19-08676.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tabakoff B, Hoffman PL, Valverius P, Borg S, Lee JM, Jaffe R, U'Prichard D, DeLeon-Jones F. Characteristics of receptors and enzymes in brains of human alcoholics. Alcohol. 1985;2:419–423. doi: 10.1016/0741-8329(85)90107-7. [DOI] [PubMed] [Google Scholar]
- Trevisan L, Fitzgerald LW, Brose N, Gasic GP, Heinemann SF, Duman RS, Nestler EJ. Chronic ingestion of ethanol up-regulates NMDAR1 receptor subunit immunoreactivity in rat hippocampus. Journal of Neurochemistry. 1994;62:1635–1638. doi: 10.1046/j.1471-4159.1994.62041635.x. [DOI] [PubMed] [Google Scholar]
- Wyszynski M, Lin J, Rao A, Nigh E, Beggs AH, Craig AM, Sheng M. Competitive binding of alpha-actinin and calmodulin to the NMDA receptor. Nature. 1997;385:439–442. doi: 10.1038/385439a0. [DOI] [PubMed] [Google Scholar]
- Zukin RS, Bennett MV. Alternatively spliced isoforms of the NMDARI receptor subunit. Trends in Neuroscience. 1995;18:306–313. doi: 10.1016/0166-2236(95)93920-s. [DOI] [PubMed] [Google Scholar]