

Abstract
The amygdala has been implicated in emotional processes, although the precise nature of the emotional deficits following amygdala lesions remains to be fully elucidated. Cognitive disturbances in the perception, recognition or memory of emotional stimuli have been suggested by some, whereas others have proposed changes in emotional arousal. To address this issue, measures of emotional arousal and valence (positivity and negativity) to a graded series of emotional pictures were obtained from patients with lesions of the amygdala and from a clinical contrast group with lesions that spared this structure. Relative to the contrast group, patients with damage to the amygdala evidenced a complete lack of an arousal gradient across negative stimuli, although they displayed a typical arousal gradient to positive stimuli. These results were not attributable to the inability of amygdala patients to process the hostile or hospitable nature of the stimuli, as the amygdala group accurately recognized and categorized both positive and negative features of the stimuli. The relative lack of emotional arousal to negative stimuli may account for many of the clinical features of amygdala lesions.
Keywords: affect, amygdala, arousal, emotion, valence, lesion
INTRODUCTION
Since the classic reports of Kluver and Bucy (1939) on affective blunting following anterior temporal lobe lesions, there has been a rapidly expanding literature on the role of the amygdala in emotion (Bechara et al., 2003). Emotional pictures, contexts or states have been shown to induce amygdala activation as measured by functional brain imaging methods (LaBar et al., 1998; Critchley et al., 2005; Norris et al., 2004; Sabatinelli et al., 2005; Williams et al., 2006). The magnitude of this activation is related to affective intensity, and is generally greater for negative than for positive emotional stimuli methods (Abercrombie et al., 1998; Critchley et al., 2005; Norris et al., 2004; Sabatinelli et al., 2005). Amygdala activation is seen during fear conditioning (LaBar et al., 1998), and lesions of this structure disrupt fear conditioning (Bechara et al., 1995; LeDoux, 2003; LaBar and Cabeza, 2006; Phelps, 2006) and the perception of potential danger (Bauman et al., 2004).
These and other findings clearly implicate the amygdala in negative affective processes (Phelps and LeDoux, 2005; Phelps, 2006), although the precise role of this structure remains to be fully elucidated. While the amygdala appears to have a predominant role in negative emotions, it has also been suggested to be important in appetitive conditioning and positive affect (Hamann et al., 2002; Everitt et al., 2003; Mather et al., 2004) and to code emotional intensity rather than, or in addition to, emotional valence (Adolphs et al., 1999; Anderson et al., 2003; Winston et al., 2005).
There is now a growing literature on human patients with amygdala lesions that provides a rich source of information on cognitive and emotional functioning after damage to this structure. Patients with amygdala damage have been reported to display less intense negative, compared to positive, emotions (Tranel et al., 2006); to be impaired at recognizing facial expressions of negative emotions (Adolphs et al., 2001); to show deficits in episodic or autobiographical emotion-related memories (Phelps and LeDoux, 2005; Buchanan et al., 2006; LaBar and Cabeza, 2006; Phelps; 2006) and to evidence reduced emotional potentiation of memory (Cahill et al., 1995; McGaugh, 2004). An hypothesis has been raised that amygdala lesions may be associated more with a deficiency in the emotional arousal component rather than a cognitive/perceptual deficit in recognition of emotional valence (Glascher and Adolphs, 2003), especially for negative stimuli (Adolphs et al., 1999; Bauman et al., 2004).
To test this hypothesis, and evaluate the generality of the findings, the present study examined the separate valence and arousal dimensions of emotion judgments in the context of a comprehensive evaluative space model (Cacioppo and Berntson, 1994; Cacioppo et al., 2004; Adolphs et al., 2005). Establishing the generality of findings across testing conditions and contexts is especially important in interpretation of lesion effects. The present study is the first to evaluate lesion effects from the perspective of a bivariate rather than bipolar model of affect (Cacioppo and Berntson, 1994; Cacioppo et al., 2004), and to use standardized picture stimuli of the International Affective Picture Series (IAPS, Lang et al., 1999). The present study applied separate bivariate valence (positivity and negativity) ratings and arousal metrics, together with graded emotional picture stimuli (very positive to very negative) to the quantification of evaluative processes in six patients with amygdala lesions, compared to a lesion contrast group and to normative ratings of these picture stimuli (from Lang et al., 1999).
METHODS
Participants
Participants were six patients (four males and two females; age 22–65 years, mean = 39.7) with amygdala lesions and six lesion contrast patients (three males and three females; 33–61 years, mean = 53.0) with damage that spared the amygdala/temporal lobe area and other areas implicated in affective processing. All patients had undergone neuroanatomical characterization according to the standard protocols of the University of Iowa Laboratory of Neuroimaging and Human Neuroanatomy (Frank et al., 1997; Damasio, 2005). The patient selection criteria were: (i) a stable and chronic lesion at least three months after onset; and (ii) involvement of a brain region that either included the amygdala or (for the clinical contrast group) excluded this structure or other areas thought to be critical for processing emotional information (ventromedial prefrontal cortex, orbitofrontal cortex and the insular cortex/SII region). Patient groups did not significantly differ in age, education, IQ (WAIS-III), memory performance (WMS-III), depression (Beck Depression Inventory) or other neuropsychological dimensions tested (Table 1).
Table 1.
Demographics and neuropsychological status
| Group | Amygdala | Contrast |
|---|---|---|
| Means (SEMs) | ||
| Age | 39.7 (18.6) | 53.0 (7.3) |
| Education | 14.0 (1.8) | 14.3 (2.6) |
| Gender | 4M/2F | 3M/3F |
| WAIS-IIIa | ||
| Verbal IQ | 101.8 (20.8) | 96.8 (21.2) |
| Performance IQ | 102.2 (19.6) | 88.2 (10.3) |
| Full Scale IQ | 102.5 (18.7) | 92.5 (16.8) |
| WMS-IIIb | ||
| IMI | 86.7 (7.9) | 88.8 (11.4) |
| GMI | 89.7 (11.4) | 90.7 (11.8) |
| WMI | 108.2 (22.1) | 98.0 (15.9) |
| Wisconsin card sorting task | ||
| categories completed | 5.0 (1.5) | 4.3 (2.1) |
| perseverative errors | 14.3 (10.8) | 21.0 (17.3) |
| Beck depression inventory | 6.0 (6.7) | 5.7 (4.0) |
aWechsler Adult Intelligence Scale (3rd edition).
bWechsler Memory Scale (3rd edition).
Apparatus
Experimental control, stimulus presentation and response recording was by a Pentium-class computer running E-prime (Psychology Software Tools, Inc., Pittsburgh). A mouse served as the response device whereby the subjects indicated valence and arousal ratings to the stimuli. For positivity/negativity ratings, the participants positioned a cursor by means of a mouse onto a bivariate display (5 × 5 unit grid) with one dimension (horizontal) indicating positivity and the other negativity. The arousal rating entailed a similar cursor placement on a single dimension (9 point) scale.
Stimuli
Stimuli were positive, negative, and neutral pictures from the IAPS (Lang et al., 1999). Pictures were matched on normative arousal ratings and on evaluative extremity from the neutral point of the normative scale (12 very positive, 6 moderately positive, 12 neutral, 6 moderately negative and 12 very negative). We limited the number of pictures to 48 in order to minimize potential fatigue and attentional confounds over time, and the stimuli were carefully selected to sample the range of affective ratings, distinct emotions, and social vs nonsocial contexts. A preliminary analysis indicated that results did not differ across social vs nonsocial stimulus contents, nor across threat vs nonthreat contexts.
Procedure
Participants rated the picture stimuli on positivity, negativity and arousal dimensions. Pictures were presented in random order on a computer monitor for 6 s. Participants were instructed to focus on the emotional content of the pictures. After viewing each picture, participants were instructed to rate it on a 5-point bivariate scale of positivity and negativity (see Appendix) and a univalent scale of how aroused it made them feel. Valence ratings were registered by moving a mouse pointer and clicking a location in a 5-point bivariate grid with the horizontal axis indexing positivity (0 = not at all, 4 = extremely) and the vertical axis indicating negativity (0 = not at all, 4 = extremely). The response grid was presented on the screen immediately after termination of the stimulus picture (Cacioppo et al., 2004). After responding, a second screen displayed a single response continuum and the subject was instructed to rate how aroused they felt to the stimulus on a 9-point scale (1 = not at all, 9 = extremely). Three seconds after completing the ratings, the next slide was presented. In addition to the separate ratings of positivity, negativity and arousal for each of the 48 picture stimuli, a net valence rating for each picture was calculated as the positivity rating minus the negativity rating.
Neuropsychological testing
In addition to the experimental ratings, each patient was evaluated in a range of neuropsychological tests. These included the Wechsler Adult Intelligence Scale (3rd edition), the Wechsler Memory Scale (3rd edition), the Wisconsin Card Sorting Task, and the Beck Depression Inventory.
Data analysis
Primary statistical evaluation of ratings data was by between-within (repeated measures on stimulus categories) analysis of variance (ANOVA) with trends, followed up by simple ANOVAs for pairwise contrasts. Derived measures entailing two-sample comparisons were tested by Student's t tests.
RESULTS
The amygdala group showed diminished arousal responses to negative stimuli
As illustrated in Figure 1A, the amygdala group showed markedly reduced arousal ratings to negative emotional stimuli, despite ratings of neutral and positive stimuli that were highly similar to those of the clinical contrast group and to a normative adult sample (Lang et al., 1999). ANOVA with trends revealed a significant effect of picture category (very positive, moderately positive, neutral, moderately negative and very negative) on arousal ratings [F(4,40) = 8.0, P < 0.001], characterized by a significant overall quadratic trend across picture categories [F(1,10) = 20.0, P < 0.001]. The latter reflected the minimal arousal to neutral stimuli and the progressively increasing arousal to either positive or negative stimuli. There also emerged a significant group × picture-category interaction [F(4,40) = 3.7, P = 0.012], characterized by a significant difference between the groups in the linear trend component across picture categories [F(1,10) = 12.7, P = 0.021]. This reflected the reduced arousal ratings, selectively for the negative pictures, of the amygdala group compared to the clinical contrast group and the normative group.
Fig. 1.
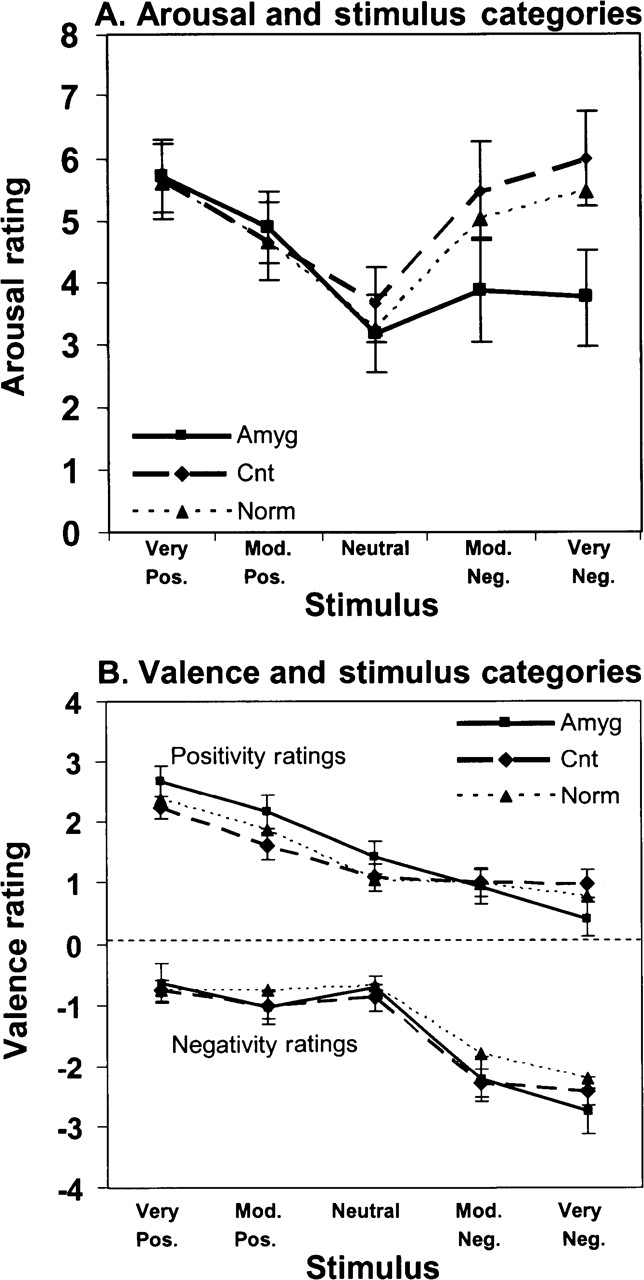
Arousal and valence ratings. Mean (s.e.m.) arousal (A) and valence (B) ratings across stimulus categories, for patients with amygdala lesions (Amyg) compared with the clinical contrast group (Cnt) and normative control data (Norm) from Lang et al. (1999). All groups effectively discriminated the stimulus categories and applied valance ratings accordingly. All groups also displayed comparable arousal functions to positive stimuli, but the amygdala group showed diminished arousal selectively to the negative stimuli.
The amygdala group accurately identified positive and negative picture content
An important question arises as to whether the diminished arousal to negative stimuli in the amygdala group may be attributable to impaired recognition or discriminative processing of the negative features of the pictures. As illustrated in Figure 1B, however, the amygdala group was able to categorize and label the negative picture content accurately, suggesting a fundamental dissociation between the cognitive and affective processing of the stimuli in this group. ANOVA with trends revealed the expected significant effects of picture category on positivity ratings [F(4,40) = 6.1, P < 0.001] and negativity ratings [F(4,40) = 17.86, P < 0.001], each being characterized by a significant linear component [for positivity ratings, [F(1,10) = 23.1, P < 0.001; for negativity, F(1,10) = 20.5, P = 0.001]. There were no significant main effects or interactions of group on either positivity or negativity ratings, although the somewhat higher slope of the positivity-rating function of the amygdala group approached significance [linear trend, F(1,10) = 2.1, P = 0.061]. This could reflect an enhancement of what has been termed the positivity offset (Cacioppo et al., 2004), although this will need to be established in subsequent studies.
Unilateral lesions yield similar effects
Two patients in the amygdala lesion group had bilateral lesions, three had right unilateral lesions, and one had a left unilateral lesion (see ‘Neuroanatomy’ subsequently). Bilateral amygdala lesions have often been found to yield larger effects than unilateral lesions, although unilateral lesions have been reported to have similar although attenuated effects (e.g. Glascher and Adolphs, 2003), or even comparable effects to bilateral lesions (Funayama et al., 2001; Buchanan et al., 2004). Results for patients with bilateral and unilateral lesions are illustrated in Figure 2.
Fig. 2.

Effects of bilateral and unilateral lesions on arousal to positive and negative stimuli. The change in arousal from neutral to the most positive stimuli are illustrated by open symbols and the change from neutral to the most negative stimuli are depicted by solid symbols. Circles depict individual data from the contrast group, squares illustrate the results from patients with bilateral amygdala lesions, and triangles show results from patients with unilateral lesions of the amygdala (right facing triangles = right sided lesions and vice versa). The means are illustrated by the horizontal lines.
Overall, the lesion contrast group evidenced a greater arousal response to negative stimuli than to positive stimuli, whereas the reverse pattern was apparent for the amygdala group. An analysis of variance revealed a significant group × picture-category (positive/negative) interaction on arousal ratings [F(1,10) = 8.46, P = 0.016]. This interaction was attributable to the similar arousal responses of the groups to positive stimuli, and the considerably smaller responses of the amygdala group to negative stimuli. As depicted in Figure 2, both unilateral and bilateral amygdala patients showed an attenuated arousal response to negative stimuli.
Neuroanatomy
Amygdala Group
All subjects had lesions involving the amygdala, although damage often extended beyond the amygdala to the anterior temporal lobe. We use the designation amygdala group for convenience, as the amygdala has been shown to be the most critical structure in the anterior temporal region for affective processing. The etiology of the lesions was either Herpes Simplex Encephalitis or anterior temporal lobectomy to treat intractable seizure disorders. The two cases of bilateral damage were secondary to Herpes Simplex Encephalitis (Figure 3A). In one case, the damage included the entire amygdala, anterior region of the hippocampus and overlaying temporal cortex on the right side, with similar but less extensive damage on the left side. The second subject was similar, with bilateral damage to both the amygdala, anterior parts of the hippocampus and surrounding temporal cortex. Three patients had surgical resections of all or most of the right amygdala, including portions of the hippocampus and anterior pole of the temporal lobe. The final patient had a complete resection of the left amygdala, without visible damage to the hippocampus or adjacent temporal cortex. A representative lesion from the amygdala group is shown in Figure 3A.
Fig. 3.

Illustrative lesions. (A) Bilateral lesion of the amygdala secondary to Herpes Simplex Encephalitis. Although only two of the six patients in the amygdala group had bilateral lesions, the lesions of this patient illustrate the range of completeness of unilateral damage to the amygdala in other patients. (B) Illustration of one of the smaller lesion in the lesion contrast group.
Lesion contrast group
This group included patients with lesions that spared the amygdala, the ventromedial prefrontal cortex, the orbitofrontal cortex and the insular cortex/SII area. In all cases, the etiology was a stroke. One patient had right posterior temporal-occipital damage at the level of the superior temporal gyrus, extending to the posterior horn of the lateral ventricles. There was also damage on the medial side in the posterior ventricular region that involved the retrosplenial cortex and the lower region of the medial aspect of the parietal cortex on the left side, and to a lesser extent on the right. Despite the extensive size of this lesion, the amygdala, hippocampus, ventromedial prefrontal cortex, orbitofrontal cortex, insular cortex and SII were all spared. One patient had a large watershed infarct, which was almost complete (sparing some temporal cortex), but with no involvement of the amygdala, insular cortex or SSII; and another had right temporal cortex damage, mostly of the superior temporal gyrus. A fourth patient had right lateral occipital damage, and another had damage to the left posterior temporal and parietal cortex. The final patient had a left posterior cerebral artery infarct with extensive damage to the hippocampus, but with sparing of the amygdala, the insula and SII. An illustrative lesion from this group is shown in Figure 3B.
DISCUSSION
The present results clarify the impact of amygdala damage on valence and arousal dimensions of affective judgments. Amygdala lesions may not necessarily disrupt the basic perception, categorization and labeling of positive and negative picture content. Rather, they may preferentially attenuate arousal effects to negative, relative to positive stimuli. Although this effect was greater with bilateral damage, unilateral injuries yielded a similar effect.1 These findings are consistent with reports that amygdala lesions may disrupt the development of conditioned autonomic arousal responses, despite the fact that patients may acquire explicit cognitive knowledge about the stimulus and outcome contingencies (Bechara et al., 1995; LaBar et al., 1995; Gazzaniga et al., 2002). Although patients with amygdala lesions have been reported to show deficits in the recognition of emotion in facial displays, these deficits are attributable in part to inadequate visual search and fixation rather than a fundamental inability to discriminate and identify negative facial features (Adolphs et al., 2005). Moreover, in accord with the present findings is that while patients may accurately judge the valence and extremity of both positive and negative facial expressions, they show lower arousal judgments selectively for negative facial displays (Adolphs et al., 1999).
The present findings contrast with the report of Funayama et al. (2001), which reported normal arousal ratings to negative pictures in patients with unilateral temporal lobectomies, although they did observe reduced startle potentiation to the negative pictures in the patients with right hemisphere damage. A potential explanation of this discrepancy may relate to the specific instructions and rating procedures. In the present study, patients were instructed to report on how aroused the pictures made them feel, with an emphasis on their arousal reaction. In the Funayama et al. (2001) study, participants ‘ … rated all pictures, first for arousal, explained as emotional strength or intensity, and then for pleasantness.’ This instruction may have led the subjects to focus on the characteristics or attributes of the pictures, rather than their reaction to the stimuli, and it is clear from the present study that lesion patients can appropriately label emotional content and the intensity or extremity of this content.
Anderson and Phelps (2002) report that patients with lesions of the amygdala show levels of self-reported negative affect on the Positive and Negative Affect Scale (PANAS) that were comparable with control subjects, although they noted that their control subjects showed relatively low base rates of negative affect. These investigators also reported a similarity in the two-factor structure (positive vs negative) in patient and control self-reports. They concluded that the amygdala may be recruited during emotion, but is not necessary for the production of emotional states. This is not necessarily inconsistent with the present results. First, the PANAS is a self-report measure that may be tapping into the patients’ cognitive knowledge of the emotional significance of life contexts. Thus, the reports from amygdala patients may reflect more of a cognitive labeling of emotion, rather than revealing the magnitude of the affective arousal. This is consistent with a recent study of a patient with bilateral amygdala lesions, where self-reports revealed an apparently normal range of affect and emotion, but further evaluation revealed that the patient was highly dispassionate and lacking in a normal sense of danger (Tranel et al., 2006). The contribution of the amygdala may not be so much in the generation of emotion, but in the affective arousal dimension of that emotion, especially for negative emotions (although the amygdala may also play an important role in positive affect as well, e.g. see Hamann et al., 2002).
The present results may explain and organize other findings in the literature. Diminished arousal to negative or threatening stimuli, for example, may underlie the general taming and reduced avoidance behavior after amygdala lesions (Tranel et al., 2006), as originally described by Kluver and Bucy (1939). A diminished arousal effect to the negative outcomes may also account for the poor performance of patients with amygdala lesions in the Iowa Gambling Task (Bechara et al., 1999, 2003). Diminished arousal may also underlie reported deficits in emotional memory and the potentiation of memory by emotional contexts after amygdala damage (Cahill et al., 1995; McGaugh, 2004). There is now a considerable literature documenting memory enhancement by emotional or autonomic/visceral arousal, and an important link in this potentiation appears to be the amygdala (McGaugh, 2004). The arousal deficiency demonstrated in the present study may underlie or contribute to deficits in the emotional potentiation of memory after amygdala damage. Consistent with this interpretation is the finding that memory potentiation is greater for negative and positive stimuli than for neutral stimuli and that amygdala damage is particularly disruptive to memory potentiation in negative contexts (D’Argembeau and Van der Linden, 2005; Buchanan et al., 2006).
In summary, the present results indicate that the amygdala may not be necessary to determine whether and to what extent a stimulus is appetitive or aversive, hostile or hospitable. These results do indicate that the amygdala is important for registering the arousal or emotional impact especially of aversive stimuli. Although the effects of lesions were notable for negative, but not positive stimuli, this does rule out an important contribution of the amygdala to appetitive processes. Indeed, there is considerable evidence for such a contribution (Anderson et al., 2003; Everitt et al., 2003; Mather et al., 2004; Winston et al., 2005). Nevertheless, the present findings raise the question of whether the emotional impact of appetitive stimuli may be governed by a distinct or more broadly distributed or redundant set of neural processes.
Acknowledgments
This research was supported by a program project grant from the National Institute of Neurological Disorders and Stroke to D.T., H.D. and A.B.; a grant from the National Institute on Drug Abuse to A.B., and grants from the National Institute of Mental Health and the Templeton Foundation to J.T.C. We thank Ashley Gruntler and Amanda Crowley for assistance with patient testing.
APPENDIX
Evaluative space grid instructions were based on Russell et al.'s (1989) affect grid instructions. The experimenter shows participants the evaluative space grid on a computer screen and explains:
One way to describe your feelings about a stimulus is in terms of how positive and how negative you feel about it, as shown in the figure. It is in the form of a grid—a kind of map for feelings. The grid asks you two questions: Along the horizontal axis, it asks how positive did the stimulus make you feel from ‘not at all’ at the left to ‘extremely’ at the right. Along the vertical axis, it asks how negative did the stimulus make you feel from ‘not at all’ on the bottom to ‘extremely’ on top.
If you feel positive, but not at all negative, move the mouse into one of these four cells at the bottom edge. The better you feel, the farther to the right you should go. On the other hand, if you feel negative but not at all positive, move the mouse into one of these four cells on the left edge. The worse you feel, the farther up you should go. If you feel neither positive nor negative, move the mouse into the cell in the bottom left. This indicates that you feel not at all positive and not at all negative. Finally, if you feel both positive and negative, move the mouse into one of the cells in the middle. The cell you select will depend on just how positive and just how negative you feel. [Throughout the instructions, the experimenter moved the mouse to highlight the appropriate cell or cells. For example, while saying, ‘The better you feel, the farther to the right you should go,’ they moved the mouse to the right along the bottom edge.]
The grid can be used to describe any pattern of positive and negative feelings. For example, you might feel quite a bit positive and not at all negative or somewhat negative and not at all positive. During the experiment, please place yourself in whichever cell best describes your feelings. After you’ve reached that cell, click to record your response.
Following the instructions, participants were given approximately 30 s to get accustomed to moving around the grid with the mouse.
Footnotes
Conflict of Interest
None declared.
Laterality issues are complex in this literature. Funayama et al. (2001) suggest the right amygdala may modulate fear while directly viewing negative pictures, whereas the left may be more involved in fear associated with cognitive/linguistic representations. Cahill (2003) reports gender differences, with greater left amygdala involvement in emotional memory in females, and greater right amygdala involvement in males. Hamann et al. (2002) suggest that the left amygdala may be more involved in positive emotion, whereas both may be implicated in negative emotion. In view of these complexities, further analysis of the small number of unilateral subjects in the present study is not feasible, although qualitatively, the results are in accord with the Cahill (2003) model of gender differences.
REFERENCES
- Abercrombie HC, Schaefer SM, Larson CL, et al. Metabolic rate in the right amygdala predicts negative affect in depressed patients. NeuroReport. 1998;9:3301–7. doi: 10.1097/00001756-199810050-00028. [DOI] [PubMed] [Google Scholar]
- Adolphs R, Gosselin F, Buchanan TW, Tranel D, Schyns P, Damasio AR. A mechanism for impaired fear recognition after amygdala damage. Nature. 2005;433:68–72. doi: 10.1038/nature03086. [DOI] [PubMed] [Google Scholar]
- Adolphs R, Russell JA, Tranel D. A role for the human amygdala in recognizing emotional arousal from unpleasant stimuli. Psychological Science. 1999;10:167–71. [Google Scholar]
- Adolphs R, Tranel D, Damasio AR. Emotion recognition from faces and prosody following temporal lobectomy. Neuropsychology. 2001;15:396–404. doi: 10.1037//0894-4105.15.3.396. [DOI] [PubMed] [Google Scholar]
- Anderson AK, Phelps EA. Is the human amygdala critical for the subjective experience of emotion? Evidence of intact dispositional affect in patients with amygdala lesions. Journal of Cognitive Neuroscience. 2002;14:709–20. doi: 10.1162/08989290260138618. [DOI] [PubMed] [Google Scholar]
- Anderson AK, Christoff K, Stappen I, et al. Dissociated neural representations of intensity and valence in human olfaction. Nature Neuroscience. 2003;6:196–202. doi: 10.1038/nn1001. [DOI] [PubMed] [Google Scholar]
- Bauman MD, Lavenex P, Mason WA, Capitanio JP, Amaral DG. The development of mother-infant interactions after neonatal amygdala lesions in rhesus monkeys. Journal of Neuroscience. 2004;24:711–21. doi: 10.1523/JNEUROSCI.3263-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bechara A, Damasio H, Damasio AR. Role of the amygdala in decision-making. Annals of the New York Academy of Sciences. 2003;985:356–69. doi: 10.1111/j.1749-6632.2003.tb07094.x. [DOI] [PubMed] [Google Scholar]
- Bechara A, Damasio H, Damasio AR, Lee GP. Different contributions of the human amygdala and ventromedial prefrontal cortex to decision-making. Journal of Neuroscience. 1999;19:5473–81. doi: 10.1523/JNEUROSCI.19-13-05473.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bechara A, Tranel D, Damasio H, Adolphs R, Rockland C, Damasio AR. Double dissociation of conditioning and declarative knowledge relative to the amygdala and hippocampus in humans. Science. 1995;269:1115–8. doi: 10.1126/science.7652558. [DOI] [PubMed] [Google Scholar]
- Buchanan TW, Tranel D, Adolphs R. Anteromedial temporal lobe damage blocks startle modulation by fear and disgust. Behavioral Neuroscience. 2004;118:429–37. doi: 10.1037/0735-7044.118.2.429. [DOI] [PubMed] [Google Scholar]
- Buchanan TW, Tranel D, Adolphs R. Memories for emotional autobiographical events following unilateral damage to medial temporal lobe. Brain. 2006;129:115–27. doi: 10.1093/brain/awh672. [DOI] [PubMed] [Google Scholar]
- Cacioppo JT, Berntson GG. Relationship between attitudes and evaluative space: a critical review with emphasis on the separability of positive and negative substrates. Psychological Bulletin. 1994;115:401–23. [Google Scholar]
- Cacioppo JT, Larsen JT, Smith NK, Berntson GG. In: Feelings and emotions: The Amsterdam conference. In: Manstead ASR, Frijda NH, Fischer AH, editors. New York: Cambridge University Press; 2004. pp. 223–42. [Google Scholar]
- Cahill L, Babinsky R, Markowitsch HJ, McGaugh JL. The amygdala and emotional memory. Nature. 1995;377:295–6. doi: 10.1038/377295a0. [DOI] [PubMed] [Google Scholar]
- Cahill L. Sex- and hemisphere-related influences on the neurobiology of emotionally influenced memory. Progress in Neuropsychopharmacology and Biological Psychiatry. 2003;27:1235–41. doi: 10.1016/j.pnpbp.2003.09.019. [DOI] [PubMed] [Google Scholar]
- Critchley HD, Taggart P, Sutton PM, et al. Activity in the human brain predicting differential heart rate responses to emotional facial expressions. Neuroimage. 2005;24:751–62. doi: 10.1016/j.neuroimage.2004.10.013. [DOI] [PubMed] [Google Scholar]
- D'Argembeau A, Van der Linden M. Influence of emotion on memory for temporal information. Emotion. 2005;5:503–7. doi: 10.1037/1528-3542.5.4.503. [DOI] [PubMed] [Google Scholar]
- Damasio H. Human brain anatomy in computerized images. 2nd edn. Oxford New York: Oxford University Press; 2005. [Google Scholar]
- Everitt BJ, Cardinal RN, Parkinson JA, Robbins TW. Appetitive behavior: impact of amygdala-dependent mechanisms of emotional learning. Annals of the New York Academy of Sciences. 2003;985:233–50. [PubMed] [Google Scholar]
- Frank R, Damasio H, Grabowski TJ. Brainvox: an interactive multimodal visualization and analysis system for neuroanatomical imaging. Neuroimage. 1997;5:13–30. doi: 10.1006/nimg.1996.0250. [DOI] [PubMed] [Google Scholar]
- Funayama ES, Grillon C, Davis M, Phelps EA. A double dissociation in the affective modulation of startle in humans: effects of unilateral temporal lobectomy. Journal of Cognitive Neuroscience. 2001;13:721–9. doi: 10.1162/08989290152541395. [DOI] [PubMed] [Google Scholar]
- Gazzaniga MS, Irvy RB, Mangun GR. Cognitive Neurosciences: The biology of the mind. 2nd edn. New York: Norton; 2002. p. 559. [Google Scholar]
- Glascher J, Adolphs R. Processing of the arousal of subliminal and supraliminal emotional stimuli by the human amygdala. Journal of Neuroscience. 2003;23:10274–82. doi: 10.1523/JNEUROSCI.23-32-10274.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamann SB, Ely TD, Hoffman JM, Kilts CD. Ecstasy and agony: activation of the human amygdala in positive and negative emotion. Psychological Science. 2002;13:135–41. doi: 10.1111/1467-9280.00425. [DOI] [PubMed] [Google Scholar]
- Kluver H, Bucy PC. Preliminary analysis of functions of the temporal lobes in monkeys. Archives of Neurology and Psychiatry. 1939;4:979–1000. doi: 10.1176/jnp.9.4.606. [DOI] [PubMed] [Google Scholar]
- LaBar KS, Cabeza R. Cognitive neuroscience of emotional memory. Nature Reviews Neuroscience. 2006;7:54–64. doi: 10.1038/nrn1825. [DOI] [PubMed] [Google Scholar]
- LaBar KS, Gatenby JC, Gore JC, LeDoux JE, Phelps EA. Human amygdala activation during conditioned fear acquisition and extinction, a mixed-trial fMRI study. Neuron. 1998;20:937–45. doi: 10.1016/s0896-6273(00)80475-4. [DOI] [PubMed] [Google Scholar]
- LaBar KS, LeDoux JE, Spencer DD, Phelps EA. Impaired fear conditioning following unilateral temporal lobectomy in humans. Journal of Neuroscience. 1995;15:6846–55. doi: 10.1523/JNEUROSCI.15-10-06846.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lang PJ, Bradley MM, Cuthbert BN International Affective Picture System (IAPS) Gainesville: FL: Center for Research in Psychophysiology; 1999. Instruction manual and affective ratings (Technical Report No. A-4) [Google Scholar]
- LeDoux J. The emotional brain, fear, and the amygdala. Cellular and Molecular Neurobiology. 2003;23:727–38. doi: 10.1023/A:1025048802629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mather M, Canli T, English T, et al. Amygdala responses to emotionally valenced stimuli in older and younger adults. Psychological Science. 2004;15:259–63. doi: 10.1111/j.0956-7976.2004.00662.x. [DOI] [PubMed] [Google Scholar]
- McGaugh JL. The amygdala modulates the consolidation of memories of emotionally arousing experiences. Annual Review of Neuroscience. 2004;27:1–28. doi: 10.1146/annurev.neuro.27.070203.144157. [DOI] [PubMed] [Google Scholar]
- Norris CJ, Chen EE, Zhu DC, Small SL, Cacioppo JT. The interaction of social and emotional processes in the brain. Journal of Cognitive Neuroscience. 2004;16:1818–29. doi: 10.1162/0898929042947847. [DOI] [PubMed] [Google Scholar]
- Phelps EA. Emotion and cognition: insights from studies of the human amygdala. Annual Review of Psychology. 2006;57:27–53. doi: 10.1146/annurev.psych.56.091103.070234. [DOI] [PubMed] [Google Scholar]
- Phelps EA, Ledoux JE. Contributions of the amygdala to emotion processing: from animal models to human behavior. Neuron. 2005;48:175–87. doi: 10.1016/j.neuron.2005.09.025. [DOI] [PubMed] [Google Scholar]
- Russell JA, Weiss A, Mendelsohn GA. Affect grid: A single-item scale of pleasure and arousal. Journal of Personality and Social Psychology. 1989;57:493–502. [Google Scholar]
- Sabatinelli D, Bradley MM, Fitzsimmons JR, Lang PJ. Parallel amygdala and inferotemporal activation reflect emotional intensity and fear relevance. Neuroimage. 2005;24:1265–70. doi: 10.1016/j.neuroimage.2004.12.015. [DOI] [PubMed] [Google Scholar]
- Tranel D, Gullickson G, Koch M, Adolphs R. Altered experience of emotion following bilateral amygdala damage. Cognitive Neuropsychiatry. 2006;11:219–32. doi: 10.1080/13546800444000281. [DOI] [PubMed] [Google Scholar]
- Winston JS, Gottfried JA, Kilner JM, Dolan RJ. Integrated neural representations of odor intensity and affective valence in human amygdala. Journal of Neuroscience. 2005;25:8903–7. doi: 10.1523/JNEUROSCI.1569-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams LM, Das P, Liddell BJ, Kemp AH, Rennie CJ, Gordon E. Mode of functional connectivity in amygdala pathways dissociates level of awareness for signals of fear. Journal of Neuroscience. 2006;26:9264–71. doi: 10.1523/JNEUROSCI.1016-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]