

Abstract
We have determined the treadmilling rate of brain microtubules (MTs) free of MT-associated proteins (MAPs) at polymer mass steady state in vitro by using [3H]GTP-exchange. We developed buffer conditions that suppressed dynamic instability behavior by ≈10-fold to minimize the contribution of dynamic instability to total tubulin-GTP exchange. The MTs treadmilled rapidly under the suppressed dynamic instability conditions, at a minimum rate of 0.2 μm/min. Thus, rapid treadmilling is an intrinsic property of MAP-free MTs. Further, we show that tau, an axonal stabilizing MAP involved in Alzheimer’s disease, strongly suppresses the treadmilling rate. These results indicate that tau’s function in axons might involve suppression of axonal MT treadmilling. We describe mathematically how treadmilling and dynamic instability are mechanistically distinct MT behaviors. Finally, we present a model that explains how small changes in the critical tubulin subunit concentration at MT minus ends, caused by intrinsic differences in rate constants or regulatory proteins, could produce large changes in the treadmilling rate.
Microtubules (MTs) play important roles in many essential cell functions including mitosis, intracellular transport, positioning of membrane-bounded organelles, and cell shape. They perform many of their cellular tasks by changing their organization and stability in response to the needs of the cell. These tasks seem to depend on the unusual nonequilibrium dynamic behaviors of MTs. One such behavior, called dynamic instability, is characterized by switching at MT ends between episodes of sustained growth and rapid shortening (1–3). Both MT ends can display dynamic instability behavior, with the plus ends displaying more extensive, rapid behavior than the minus ends (2, 4, 5). Another dynamic behavior, called treadmilling, is the growth of a MT at the plus end and shortening at the minus end, which produces a flow of subunits from one end of the MT to the other (6–18). MT dynamics become extremely rapid when mammalian cells progress from interphase to mitosis, when rapid dynamics are required for the construction of the mitotic spindle and for various chromosome movements (19–22). Rapid dynamics consistent with treadmilling, termed flux, occur during metaphase and anaphase of mitosis (23–25). Rapid growth occurs at the plus ends of the MTs tethered to the kinetochores of the chromosomes, and balanced shortening occurs at the minus ends, which are tethered at the centrosomes.
The treadmilling rate of MTs was found to be extremely slow in early studies in vitro with brain MTs composed of a high complement of MT-associated proteins (MAPs; refs. 6 and 12). The method involved assembling the MTs to polymer mass steady state and carrying out pulse and pulse–chase experiments with radiolabeled GTP-tubulin to reveal the sites and rates of tubulin gain and loss. The strategy depended on two properties of the system. The radiolabeled GTP exchanged rapidly with the β-subunits of tubulin dimers when the tubulin was free in solution and became nonexchangeably bound when the tubulin was incorporated into the MTs. Second, the dynamic instability behavior of the MAP-rich MTs was extremely low (2, 26–30), such that exchange of the GTP-tubulin into the MTs by treadmilling was not complicated by significant growing and shortening dynamics caused by dynamic instability.
The treadmilling rate of the MAP-rich MTs was only ≈0.7 μm/h (6). It was argued that such a slow rate could not contribute to MT turnover in cells (31). By using the same approach with radiolabeled GTP-tubulin and MAP-free MTs that displayed dynamic instability, a much more rapid treadmilling rate of ≈50 μm/h was calculated (12). However, in this early work, dynamic instability was estimated from static length distributions determined at different times by electron microscopy at steady state, and the value of 50 μm/h for the treadmilling rate remains uncertain.
Recently, rapid MT treadmilling in the absence of dynamic instability was demonstrated in cells (14, 18). MTs that had become detached from centrosomes treadmilled at rates as high as 10 μm/min. The treadmilling of these MTs and, similarly, the flux of MTs in metaphase and anaphase spindles could be caused by the intrinsic treadmilling capability of MTs. However, regulatory proteins could also create apparent treadmilling behavior (23). Another possibility that may account for the high treadmilling rate in cells is that treadmilling is an intrinsic property of MTs, and regulatory proteins modulate the rate. In support of the idea that accessory proteins might increase the treadmilling rate, the slow treadmilling rate of actin filaments in vitro is increased greatly by cofilin/actin depolymerizing factor (32–34).
Thus, we wanted to determine the intrinsic treadmilling rate of MAP-free MTs. In the present work, we used the rate of radiolabeled GTP-tubulin incorporation into MTs to determine the treadmilling rate under conditions of minimal dynamic instability. The results show that rapid MT treadmilling is an intrinsic property of MAP-free MTs. Further, we show that tau strongly suppresses the treadmilling rate. Finally, we describe the mechanistic distinction between treadmilling and dynamic instability and describe how small changes in the critical tubulin concentration at minus ends, determined by the dissociation rate constant, could produce large changes in the treadmilling rate.
Materials and Methods
Preparation of MAP-Free Tubulin and Tau.
MT protein was prepared by two cycles of assembly/disassembly without glycerol as described (35). Tubulin was purified from the MT protein by elution through a phosphocellulose column (Whatman P-11) equilibrated in 50 mM Pipes/1 mM MgSO4/1 mM EGTA/0.1 mM GTP, drop frozen in liquid nitrogen, and stored at −70°C. The tubulin was greater than 99% pure. No other proteins could be detected by SDS/PAGE on overloaded gels after staining with Coomassie blue (data not shown). Recombinant full-length human tau was expressed in Escherichia coli and purified as described (29).
Video Microscopic Analysis of Dynamic Instability Under Reduced Dynamics Conditions.
A number of solution conditions have been shown to suppress dynamic instability, including the presence of glycerol (31, 36) and the use of high tubulin concentrations (4). In our preliminary experiments, we also found that dynamic instability behavior was suppressed by reducing the temperature from 37°C to 30°C. We settled on 2% (vol/vol) glycerol, 30°C, and a total tubulin concentration of 30 μM as a set of conditions to reduce strongly but not eliminate the dynamic instability behavior of the MTs (see Results). MAP-free tubulin was mixed with sea urchin sperm axonemal seeds (5) in PME buffer (100 mM Pipes/1 mM Mg2+/1 mM EGTA) containing 1 mM GTP and a final concentration of 2% (vol/vol) glycerol. The reaction mixture was incubated at 30°C for 30 min to reach steady state, and the polymerization dynamics of the MTs were recorded by video microscopy and analyzed as described (5). Under the conditions used, MT growth occurred at both ends of the seeds with strongly suppressed dynamics compared with dynamics under normal buffer conditions. The end with the higher growing rates, greater excursion lengths, and larger number of MTs was considered to be the plus end (4, 5, 35). The limit of resolution of the video microscope was ≈0.2 μm (4, 5, 35).
Determination of Treadmilling Rates.
To prepare MT seeds, 30 μM tubulin was polymerized into MTs in PME buffer containing 10% (vol/vol) glycerol at 30°C for 30 min and sheared as described (12). Tubulin (30 μM) was polymerized at the ends of the seeds after diluting the seeds 5-fold in PME buffer containing 100 μM GTP and a GTP-regenerating system (12). The final glycerol concentration was 2% (vol/vol). After 30 min of polymerization at 30°C, the steady-state MTs were pulsed with [3H]GTP at different times, and the kinetics of [3H]GDP incorporation into the MTs were determined after sedimentation (6). Briefly, at the desired times after pulsing, 100-μl portions of MT suspensions were placed gently at the top of 50% (vol/vol) sucrose cushions (4.5 ml; PME buffer; 30°C). Incorporated [3H]GDP was separated from unincorporated [3H]GTP-tubulin by centrifugation at 190,000 × g for 75 min at 30°C (Beckman–Spinco SW 50.1 rotor). MT pellets were dissolved at 0°C; the tubulin concentration was determined; and the amount of [3H]GDP incorporated into the MTs was determined by scintillation counting. The mean length of the MTs and the MT-length distributions were determined by electron microscopy (12). Protein concentration was determined by the Bradford method (37) by using BSA as the standard.
Results
MT Polymerization Under Suppressed Dynamic Instability Conditions.
Our goal was to determine the intrinsic treadmilling rate of steady-state MAP-free brain MTs by using [3H]GTP-tubulin exchange. It was first necessary to develop conditions in which dynamic instability was sufficiently suppressed such that growth and shortening caused by dynamic instability would not complicate the analysis. When purified tubulin (30 μM) was polymerized into MTs in Pipes buffer at 30°C in the presence of 2% (vol/vol) glycerol, the length excursions at the MT ends were strongly suppressed. Polymerization of the MTs under the conditions used is shown in Fig. 1A. Steady state was attained in less than 30 min, and a plateau was maintained for at least 90 min. Two methods were used to determine the extent to which growing and shortening excursions characteristic of dynamic instability were suppressed. If extensive growing and shortening occurs at MT ends in a population, the mean length of the population will increase, and the number concentration will decrease with time because of the loss of MTs that fully depolymerize (1). Thus, we determined the steady-state length distributions of the MT population by electron microscopy at 28 min and 90 min. In the experiment shown in Fig. 1 B and C, the mean length was 20.3 μm ± 0.8 μm (SEM) at 28 min and 21.7 μm ± 0.8 μm (SEM) at 90 min. In 10 separate experiments, the mean length of the MTs did not change significantly (data not shown). The shapes of the distributions, which were nearly Gaussian, also did not change significantly. The absence of significant increases in the mean length or of changes in the length distributions indicates that there was no loss of MTs, including very short MTs. Thus, very little dynamic instability was occurring.
Figure 1.

Length distributions of MAP-free MTs at steady state. (A) Polymerization of tubulin (30 μM) was monitored by light scattering at 350 nm. Length distributions were determined by electron microscopy 28 min (B) and 90 min (C) after initiation of polymerization.
We also quantitated the dynamic instability parameters directly at plus MT ends by video microscopy. Typical life-history traces showing the dynamic instability behavior of nonsuppressed and suppressed MTs are shown in Fig. 2 A and B, respectively. It is clear that, under the stabilizing conditions used, the dynamic instability behavior of the MT plus ends was strongly suppressed. The length grown during a growing event and the length shortened during a shortening event were strongly dampened. The catastrophe frequency was low; the rescue frequency was high; and the overall dynamicity (the total visually detectable growth and shortening per unit of time) was low. Specifically, the growth and shortening rates were reduced 3.9-fold and 10-fold, respectively (Table 1). Similarly, the catastrophe frequency was reduced 3.8-fold, and the overall dynamicity was reduced ≈10-fold. The dynamic instability behavior at the minus ends was also determined and found to be 37% of that at the plus ends, consistent with previous reports (data not shown; refs. 4 and 5).
Figure 2.

Life-history traces of MT plus ends. (A) Growing and shortening length changes under dynamic conditions. Tubulin (13 μM) was polymerized into MTs at the ends of axonemal seeds in 87 mM Pipes/36 mM Mes/1.8 mM Mg2+/1 mM EGTA/1.5 mM GTP, pH 6.8, at 37°C. (B) MT length changes under reduced dynamic instability conditions (see Materials and Methods).
Table 1.
Dynamic instability parameters at steady state
| Parameter | Reduced dynamic condition | Dynamic condition |
|---|---|---|
| Rate μm/min | ||
| Growing | 0.32 ± 0.04 | 1.25 ± 0.5 |
| Shortening | 1.5 ± 0.40 | 16.7 ± 5.5 |
| Mean length, μm per event | ||
| Grown length | 0.90 ± 0.094 | 2.5 ± 0.3 |
| Shortened length | 0.87 ± 0.09 | 6.0 ± 0.5 |
| Frequencies of transitions, min−1 | ||
| Catastrophe | 0.087 ± 0.016 | 0.34 ± 0.06 |
| Rescue | 0.91 ± 0.18 | 0.85 ± 0.24 |
| Dynamicity, μm/min | 0.3 | 3.0 |
Values are shown as means ± SD, except for the mean lengths grown and shortened, which are ± SEM. Microtubules (n = 59) were measured for 352 min for the reduced dynamic condition.
Steady-State MAP-Free MTs Display Rapid Treadmilling Under Suppressed Dynamic Instability Conditions.
Polymerization was carried out by adding glycerol seeds to a prewarmed tubulin solution at 30°C. The final glycerol concentration was 2% (vol/vol). [3H]GTP pulses were added at steady state (30 min), and the quantity of [3H]GDP-tubulin incorporation into the MTs was determined at desired times between 35 min and 80 min. As shown in Fig. 3, net incorporation of [3H]GDP-tubulin into the MTs (mean length = 26.1 μm) occurred in a linear fashion at a rate of 0.38 μm/min. The rate of incorporation per MT was calculated from the concentration of MTs in suspension by using a value of 1,690 tubulin subunits per μm of length (6, 12). The mean rate of incorporation for five independent experiments was 0.32 ± 0.04 μm/min. The mean length of the MTs in all five experiments was 21 μm. Thus, half the length of a typical MT was labeled with the newly incorporated [3H]GDP in ≈30 min.
Figure 3.

Incorporation of [3H]GDP-tubulin into steady-state MTs in the absence (squares) and presence (diamonds) of 2 μM tau. MAP-free tubulin (30 μM) was polymerized into MTs in PME buffer containing 2% (vol/vol) glycerol at 30°C for 30 min (see Materials and Methods). At steady state, the MTs were pulsed with [3H]GTP, and the kinetics of [3H]GDP incorporation into the MTs were determined after centrifugation through 50% (vol/vol) sucrose cushions (see Materials and Methods). The mean lengths in the presence and absence of tau were 29.3 μm ± 1.1 μm and 26.1 μm ± 0.9 μm, respectively.
Steady-state MAP-rich MTs do not display appreciable dynamic instability behavior. Previously, we have shown that these MTs treadmill slowly (at a rate of ≈0.7 μm/h). They display a linear incorporation of [3H]GDP-tubulin into MTs at their net assembly ends (now known to be the plus ends) and a balanced linear loss at the minus ends (6, 11, 12). In a treadmilling system, the addition of tubulin subunits at plus ends is balanced by an equivalent loss of tubulin subunits at the minus ends (6). The rate of tubulin loss from a MT population can be determined by polymerizing the MTs in the presence of [3H]GTP to label the MTs with [3H]GDP fully and then by chasing with excess unlabeled GTP. As shown in Fig. 4, the loss of [3H]GDP-tubulin from the fully labeled MTs occurred in a linear fashion at a rate of 0.26 ± 0.1 μm/min. Thus, the ratio of [3H]GDP incorporation to [3H]GDP loss was 1 to 0.8, which is close to the theoretical ratio of 1:1 required for treadmilling.
Figure 4.

Kinetics of [3H]GDP-tubulin loss from steady-state MTs during a chase with excess unlabeled GTP. MTs were labeled fully with [3H]GDP by polymerizing [3H]GTP-tubulin in PME buffer containing 2% (vol/vol) glycerol at 30°C (see Materials and Methods). [3H]GDP-labeled MTs were chased with a 27-fold excess of GTP, and the loss of [3H]GDP-tubulin from the MTs was determined (see Materials and Methods).
Determination of the Treadmilling Rate.
We used the measured dynamic instability behavior of the axoneme-seeded MTs to estimate the contribution of dynamic instability to total subunit exchange in the steady-state [3H]GTP-tubulin pulse experiments. We calculated the minimum treadmilling rates of the MTs in two ways. For both, we considered that the extent of [3H]GDP-tubulin incorporated into the MTs during a pulse was due to a combination of new growth at plus ends caused by treadmilling and new retained growth caused by dynamic instability. In the first way, we assumed that the extent of new growth that occurred by dynamic instability in a given period of time was equal to the total growth caused by all growing excursions minus the total shortening from all shortening excursions. As determined by video microscopy, MTs spent 65.5% of the total time growing. Thus, a MT could have grown 12.4 μm in 1 h if there were no shortening excursions. Similarly, a MT would have shortened an average of 4.9 μm in 1 h. If all of the growth were caused by dynamic instability, then the average net growth would have been 7.5 μm. Subtracting this value from the total incorporation that would occur in a 1-h [3H]GTP pulse yielded a treadmilling rate of 12 μm/h. Thus, when calculated in this way, a minimum of ≈62.5% of the total incorporation was caused by treadmilling.
In the second way, we reasoned that an average MT would lose a maximum of ≈5 μm of length in 1 h because of dynamic instability if the loss occurred in the absence of any growth excursions. However, during the chase experiment, the MTs lost [3H]GDP-tubulin subunits at a rate of 15.4 μm per h. Thus, individual MTs must have lost a minimum of ≈10.4 μm of length per h because of treadmilling or ≈67.5% of the total loss of radiolabeled GDP-tubulin. Thus, determination of the treadmilling rate with both methods yielded similar results.
Effects of Tau on the Treadmilling Rate.
Tau is an important axonal MAP involved in controlling MT dynamics and stability in axons (38, 39). In neurons, tau colocalizes with axonal MTs, and it is an essential protein for axonal growth. In vitro, low ratios of tau to tubulin strongly suppress dynamic instability (26, 27, 29). In this study, we wanted to determine whether and, if so, how strongly tau could suppress the treadmilling rate.
Tau was added to preassembled MTs at a tau to tubulin ratio of 1:15 (2 μM tau; 30 μM total tubulin), and the reaction mixture was incubated for a additional 25 min at 30°C. At steady state, the MTs were pulsed with [3H]GTP to determine the new tubulin growth rate. As shown in Fig. 3, tau suppressed the rate of [3H]GTP incorporation into the MTs by 68% from a mean of 19.4 μm/h to 6.2 μm/h. Thus, a low ratio of tau to tubulin in a MT suspension strongly suppresses the treadmilling rate.
Discussion
Rapid Treadmilling of MAP-Free Brain MTs Under Suppressed Dynamic Instability Conditions.
We have determined the minimal treadmilling rate of MAP-free bovine brain MTs in vitro in which dynamic instability behavior was strongly suppressed. Under the conditions used, the total detectable dynamic instability behavior, the dynamicity, was reduced 10-fold from 3 μm/min to 0.3 μm/min. The minimum treadmilling rate, determined after subtracting the maximum amount of radiolabeled GTP that could have been incorporated because of dynamic instability, was ≈0.2 μm/min. The intrinsic treadmilling rate of MAP-free MTs in nonstabilizing conditions could be considerably greater. In addition, we show that the neuronal MAP tau, at a ratio of only 1 molecule of tau per 15 molecules of tubulin, suppressed the treadmilling rate by ≈70%. The results show that rapid treadmilling is an intrinsic property of steady-state MAP-free MTs and that tau strongly suppresses the treadmilling rate.
Treadmilling Is Distinct Mechanistically from Dynamic Instability.
Let us compare the mathematical models describing treadmilling and dynamic instability. The theoretical basis for treadmilling, which requires irreversible nucleotide hydrolysis, was described originally by Wegner (40) for actin filaments. We assume here that the association of tubulin-GDP at MT ends is negligible. Experimentally, we have ensured that this assumption is correct by carrying out polymerization in the presence of a high concentration of GTP and a GTP-regenerating system. We also assume that the dissociation of tubulin-GTP from MT ends is negligible, because MTs consist predominantly of tubulin-GDP.
The growth rates at opposite MT ends can be written as
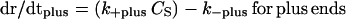 |
1 |
and
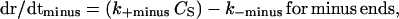 |
2 |
where dr/dt is the growth rate, k+ is the on-rate constant, k− is the off-rate constant, and CS is the solution concentration of tubulin. As mentioned above, because of the hydrolysis of GTP during polymerization, tubulin association at MT ends occurs as GTP-tubulin, and tubulin dissociation occurs as GDP-tubulin. Thus, the association and dissociation reactions are different. The critical concentration for a specific end is the ratio of the dissociation and association rate constants for that end, and the critical concentration for the plus and minus ends can be different because of GTP hydrolysis (40).
Thus, Eqs. 1 and 2 can be rewritten as
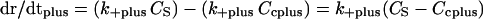 |
3 |
and
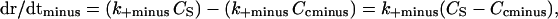 |
4 |
where Ccplus is the critical concentration for the plus ends and Ccminus is the critical concentration for the minus ends. When Cs is greater than Ccplus and lower than Ccminus, the plus end will grow, and the minus end will shorten. It is clear from extensive data that in fact the critical concentration at plus ends is lower than the critical concentration at minus ends (4, 5, 7). Caplow et al. (41) have shown that the free energy released during hydrolysis of GTP to GDP when tubulin is added to MT ends is stored in the MT lattice. This free energy must be used differentially at the opposite MT ends. Thus, the fundamental mechanism responsible for treadmilling is the difference in the critical subunit concentration at the plus and minus ends. At steady state, the concentration of tubulin in solution (Cs) will be maintained between the critical subunit concentrations for each end such that the plus ends will grow and the minus ends will shorten in a balanced fashion.
Treadmilling has been considered to occur through biased dynamic instability at opposite MT ends (see refs. 16 and 17). However, the fundamental mechanisms responsible for treadmilling and dynamic instability are different. Dynamic instability, the switching at MT ends between relatively slow growth and rapid shortening, is postulated to be caused by the gain and loss of a stabilizing “cap” at MT ends consisting of a short stretch of GTP-tubulin, GDP plus Pi-tubulin, or a conformationally stabilizing structure (1, 4, 42, 43). The end grows when it is capped and shortens rapidly when the cap is lost (1, 4, 42). With dynamic instability, MT growth for the plus and minus ends can be written as
 |
5 |
and
 |
6 |
where krs is the shortening rate constant during rapid shortening and Tg and Trs represent the fraction of time growing and rapidly shortening, respectively. Although biased dynamic instability could, in principle, result in net growth at one end and net shortening at the other end, such a situation would be difficult to achieve, because it would require a complex balance of growth and shortening at the opposite ends (4, 16). When the dynamic instability terms are deleted from Eqs. 5 and 6, we have Eqs. 3 and 4. Thus, theoretically, treadmilling can occur independently of dynamic instability. In fact, rapid treadmilling is observed to occur in cells in the absence of detectable dynamic instability (14, 18). To obtain treadmilling, all that is required is a difference in the subunit critical concentration at the opposite MT ends.
The Dissociation Rate Constant at Minus Ends May Control the Treadmilling Rate.
Although regulation of the treadmilling rate could, in principle, be accomplished by modulation of the tubulin dissociation rate at both MT ends, it seems likely that the treadmilling rate is controlled by modulation of the tubulin subunit dissociation at the minus ends. In support of this idea, most MT minus ends in CHO-K1 cells are anchored at the centrosomes and seem to be unavailable for subunit exchange. The predominant dynamic behavior of such MTs is dynamic instability at their plus ends; they do not treadmill (18). In contrast, many of the MTs that are not attached to centrosomes show rapid treadmilling behavior in the absence of detectable dynamic instability (18).
MTs from different tissues or different species could have intrinsically different tubulin subunit dissociation rate constants and, thus, could treadmill at different rates. In addition, regulatory proteins acting at minus ends could greatly modulate the treadmilling rate. Suppose a protein factor or a small ligand increases the critical concentration at MT minus ends by increasing the dissociation rate constant. The resulting depolymerization at the minus ends will tend to increase the soluble tubulin-GTP concentration, thus accelerating plus-end growth. The consequence will be a striking increase in the treadmilling rate as shown for a series of different dissociation rate constants in Table 2. For example, as little as a 20% increase in the minus-end dissociation rate constant would cause a 7.6-fold increase in the treadmilling rate. A 24-fold increase in the treadmilling rate would be achieved with only a 70% increase in the minus-end dissociation rate constant. Extremely rapid MT treadmilling rates (4- 12 μm/min) have been observed in cells (14, 18). In the present work, we determined that the MTs treadmilled at a minimum rate of ≈0.2 μm/min. Only a 55% increase in the minus-end critical concentration would increase the treadmilling rate 24-fold to 4.8 μm/min, approximately that observed in living cells.
Table 2.
Influence of the minus-end critical concentration on the treadmilling rate
| Percentage of increase in kminus | Ccminus | CS | Net growth rate at plus ends, ×k+plus* | Fold increase in treadmilling rate |
|---|---|---|---|---|
| 10 | 11 | 10.5 | 1.3 | 4.3 |
| 20 | 12 | 11.5 | 2.3 | 7.6 |
| 30 | 13 | 12.5 | 3.3 | 11 |
| 50 | 15 | 14.5 | 5.3 | 17.7 |
| 70 | 17 | 16.5 | 7.3 | 24.3 |
k+plus, the association constant at plus ends; CS, the soluble tubulin-GTP concentration; Ccminus, critical subunit concentration at minus ends (μM). We arbitrarily assume that in the absence of a protein factor, CS = 9.5 μM; Ccplus = 9.2 μM; and Ccminus = 10 μM. Under this condition, the net growing rate at the plus ends will be [9.5 (k+plus) − 9.2 (k+plus)] = 0.3 (k+plus). At steady state, the net association must be balanced by the net dissociation. Thus, the treadmilling rate will be 0.3 (k+plus).
*Net growth rate at plus ends is calculated by (k+plus × CS − Ccplus × k+plus).
Interestingly, an analogous situation may hold for actin filaments. Recent evidence indicates that cellular factors such as cofilin interact with the minus ends of actin filaments to increase the treadmilling rate (33, 34). A protein factor recently described in sea urchin egg extracts may be just such a factor for MTs (44). In addition, a stabilizing factor that reduces the catastrophe frequency and shortens the mean length during a shortening excursion or a capping protein that sustains plus-end growth could strongly suppress the contribution of dynamic instability at MT plus ends. Under such conditions, treadmilling could be the predominant mechanism for dynamic tubulin exchange.
Possible Regulation of the MT Treadmilling Rate in Axons by Tau.
The functions of treadmilling in cells are not clear. In mitotic spindles, MTs of fixed length tethered at the kinetochores and poles display rapid poleward flux, which most likely involves treadmilling. Treadmilling in spindles may create a poleward flow of signaling molecules from the kinetochores to the poles. It also may be involved in creating spindle tension (17). Treadmilling may be involved in the transport of materials in neuronal cell processes.
Tau is an axon-specific MAP that stabilizes MTs. A hyperphosphorylated form of tau seems to be the primary protein component of the fibrillary tangles characteristic of Alzheimer’s disease, and it is postulated that tau plays an important role in Alzheimer’s disease (38, 39). We found previously that low ratios of tau to tubulin in MTs powerfully stabilize the dynamic instability of MTs. For example, a ratio of only 1 molecule of tau per 11 molecules of tubulin in a MT suspension is sufficient to reduce the plus-end shortening rate by 69% from 16.6 μm/min to 5.2 μm/min (29). In the present work, we found that a ratio of 1 molecule of tau to 15 molecules of tubulin suppressed the treadmilling rate by ≈70%. Thus, tau also has a powerful ability to suppress treadmilling. Tau most likely binds along MT surfaces, and its ability to suppress treadmilling may be caused by a decrease in the dissociation rate constant at minus ends, thus reducing the difference in the critical concentrations at opposite MT ends. Although axonal MTs display some dynamic behavior, they are relatively stable, and their stability seems important for their function. In axons, the minus ends of MTs are not attached to the centrosomes and are most likely free to exchange tubulin with tubulin in the soluble pool. Thus, they are likely to be able to treadmill, and tau could stabilize axonal MTs by suppressing the treadmilling rate. Because the plus ends of axonal MTs are away from the cell body, suppression of treadmilling by tau could be important in controlling the retrograde transport of molecules from the nerve ending to the cell body.
Acknowledgments
We wish to thank Drs. Mary Ann Jordan, Richard Himes, Robert Margolis, Deborah Feygensen, and Vivian Ngan for valuable discussions and for critically reading the manuscript. This work was supported by U.S. Public Health Service Grant NS13560.
Abbreviations
- MT
microtubule
- MAP
MT-associated protein
References
- 1.Mitchison T, Kirschner M. Nature (London) 1984;312:237–242. doi: 10.1038/312237a0. [DOI] [PubMed] [Google Scholar]
- 2.Horio T, Hotani H. Nature (London) 1986;321:605–607. doi: 10.1038/321605a0. [DOI] [PubMed] [Google Scholar]
- 3.Sammak P J, Borisy G G. Nature (London) 1988;332:724–726. doi: 10.1038/332724a0. [DOI] [PubMed] [Google Scholar]
- 4.Walker R A, O’Brien E T, Pryer N K, Soboeiro M F, Voter W A, Erickson H P, Salmon E D. J Cell Biol. 1988;107:1437–1448. doi: 10.1083/jcb.107.4.1437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Panda D, Jordan M A, Chin K, Wilson L. J Biol Chem. 1996;271:29807–29812. doi: 10.1074/jbc.271.47.29807. [DOI] [PubMed] [Google Scholar]
- 6.Margolis R L, Wilson L. Cell. 1978;13:1–8. doi: 10.1016/0092-8674(78)90132-0. [DOI] [PubMed] [Google Scholar]
- 7.Bergen G L, Borisy G G. J Cell Biol. 1980;84:141–150. doi: 10.1083/jcb.84.1.141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Margolis R L, Wilson L. Nature (London) 1981;293:705–711. doi: 10.1038/293705a0. [DOI] [PubMed] [Google Scholar]
- 9.Cote R H, Borisy G G. J Mol Biol. 1981;150:577–602. doi: 10.1016/0022-2836(81)90382-x. [DOI] [PubMed] [Google Scholar]
- 10.Rothwell S W, Grasser W A, Murphy D B. J Cell Biol. 1985;101:1637–1642. doi: 10.1083/jcb.101.5.1637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Wilson L, Farrell K W. Ann N Y Acad Sci. 1986;466:690–708. doi: 10.1111/j.1749-6632.1986.tb38452.x. [DOI] [PubMed] [Google Scholar]
- 12.Farrell K W, Jordan M A, Miller H P, Wilson L. J Cell Biol. 1987;104:1035–1046. doi: 10.1083/jcb.104.4.1035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hotani H, Horio T. Cell Motil Cytoskeleton. 1988;10:229–236. doi: 10.1002/cm.970100127. [DOI] [PubMed] [Google Scholar]
- 14.Rodionov V I, Borisy G G. Science. 1997;275:215–218. doi: 10.1126/science.275.5297.215. [DOI] [PubMed] [Google Scholar]
- 15.Waterman-Storer C M, Salmon E D. J Cell Biol. 1997;139:417–434. doi: 10.1083/jcb.139.2.417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Waterman-Storer C M, Salmon E D. Curr Biol. 1997;7:R369–R372. doi: 10.1016/s0960-9822(06)00177-1. [DOI] [PubMed] [Google Scholar]
- 17.Margolis R L, Wilson L. BioEssays. 1998;20:830–836. doi: 10.1002/(SICI)1521-1878(199810)20:10<830::AID-BIES8>3.0.CO;2-N. [DOI] [PubMed] [Google Scholar]
- 18.Rodionov V I, Nadezhdina E, Borisy G G. Proc Natl Acad Sci USA. 1999;96:115–120. doi: 10.1073/pnas.96.1.115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hayden J J, Bowser S S, Rieder C. J Cell Biol. 1990;111:1039–1045. doi: 10.1083/jcb.111.3.1039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Gliksman N R, Skibbens R V, Salmon E D. Mol Biol Cell. 1993;4:1035–1050. doi: 10.1091/mbc.4.10.1035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Wordeman L, Mitchison T J. In: MTs. Hyams J, Lloyd C, editors. New York: Wiley–Liss; 1994. pp. 287–301. [Google Scholar]
- 22.Zhai Y, Kronebusch P J, Simon P M, Borisy G G. J Cell Biol. 1996;135:201–214. doi: 10.1083/jcb.135.1.201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Mitchison T J. J Cell Biol. 1989;109:637–652. doi: 10.1083/jcb.109.2.637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mitchison T J, Salmon E D. J Cell Biol. 1992;119:569–582. doi: 10.1083/jcb.119.3.569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Sawin K E, Mitchison T J. Mol Biol Cell. 1994;5:217–226. doi: 10.1091/mbc.5.2.217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Pryer N K, Walker R A, Bourns B D, Soboeiro M F, Salmon E D. J Cell Sci. 1992;103:965–976. doi: 10.1242/jcs.103.4.965. [DOI] [PubMed] [Google Scholar]
- 27.Drechsel D N, Hyman A A, Cobb M H, Kirschner M W. Mol Biol Cell. 1992;3:1141–1154. doi: 10.1091/mbc.3.10.1141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kowalski R J, Williams R C. J Biol Chem. 1993;268:9847–9855. [PubMed] [Google Scholar]
- 29.Panda D, Goode B L, Feinstein S C, Wilson L. Biochemistry. 1995;34:11117–11127. doi: 10.1021/bi00035a017. [DOI] [PubMed] [Google Scholar]
- 30.Dhamodharan R, Wadsworth P. J Cell Sci. 1995;108:1679–1689. doi: 10.1242/jcs.108.4.1679. [DOI] [PubMed] [Google Scholar]
- 31.Kristofferson D, Mitchison T, Kirschner M. J Cell Biol. 1986;102:1007–1019. doi: 10.1083/jcb.102.3.1007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Pollard T D. J Cell Biol. 1986;103:2747–2754. doi: 10.1083/jcb.103.6.2747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Theriot J A. J Cell Biol. 1997;136:1165–1168. doi: 10.1083/jcb.136.6.1165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Carlier M F, Laurent V, Santolini J, Melki R, Didry D, Xia G X, Hong Y, Chua N H, Pantaloni D. J Cell Biol. 1997;136:1307–1322. doi: 10.1083/jcb.136.6.1307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Toso R J, Jordan M A, Farrell K W, Matsumoto B, Wilson L. Biochemistry. 1993;32:1285–1293. doi: 10.1021/bi00056a013. [DOI] [PubMed] [Google Scholar]
- 36.O’Brien E T, Erickson H P. Biochemistry. 1989;28:1413–1422. doi: 10.1021/bi00429a070. [DOI] [PubMed] [Google Scholar]
- 37.Bradford M M. Anal Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- 38.Trojanowski J Q, Lee V M. Am J Pathol. 1994;144:449–453. [PMC free article] [PubMed] [Google Scholar]
- 39.Goedert M, Jakes R, Spillantini M G, Crowther R A. In: MTs. Hyams J, Lloyd C, editors. New York: Wiley–Liss; 1994. pp. 183–200. [Google Scholar]
- 40.Wegner A. J Mol Biol. 1976;108:139–150. doi: 10.1016/s0022-2836(76)80100-3. [DOI] [PubMed] [Google Scholar]
- 41.Caplow M, Ruhlen R L, Shanks J. J Cell Biol. 1994;127:779–788. doi: 10.1083/jcb.127.3.779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Carlier M F. Int Rev Cytol. 1989;115:139–170. doi: 10.1016/s0074-7696(08)60629-4. [DOI] [PubMed] [Google Scholar]
- 43.Chrétien D, Fuller S D, Karsenti E. J Cell Biol. 1995;129:1311–1328. doi: 10.1083/jcb.129.5.1311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Spitte C S, Cassimeris L. Cell Motil Cytoskeleton. 1996;34:324–335. doi: 10.1002/(SICI)1097-0169(1996)34:4<324::AID-CM7>3.0.CO;2-7. [DOI] [PubMed] [Google Scholar]