

Abstract
A Ca2+-requiring catalytic RNA is shown to create 5′ phosphate–phosphate linkages with all nucleotides and coenzymes including CoA, nicotinamide adenine dinucleotide phosphate, thiamine phosphate, thiamine pyrophosphate, and flavin mononucleotide. In addition to these small molecules, macromolecules such as RNAs with 5′-diphosphates, and nonnucleotide molecules like Nɛ-phosphate arginine and 6-phosphate gluconic acid also react. That is, the self-capping RNA isolate 6 is an apparently universal 5′ phosphate-linker, reacting with any nucleophile containing an unblocked phosphate. These RNA reactions demonstrate a unique RNA catalytic capability and imply versatile and specific posttranscriptional RNA modification by RNA catalysis.
RNA molecules are thought to have played prominent roles as both carriers of genetic information and catalysts for chemical transformations in a primordial RNA world (1, 2), from which modern protein-catalyzed metabolism has evolved (3, 4). This RNA world hypothesis requires that RNA molecules be able to catalyze a broad range of chemical reactions that are presently carried out by protein enzymes in modern protein–nucleic acid world. Whereas RNAs have been shown to catalyze a variety of reactions, most of which involve the nucleic acid phosphate–sugar backbone, RNA-catalyzed reactions discovered so far are still quite limited compared with protein-catalyzed reactions. Thus new RNA-catalyzed reactions expand the catalytic RNA repertoire and thereby provide clues to the extent and nature of the RNA world (1, 2).
We selected an RNA (isolate 6) that catalyzed a self-capping reaction with free GDP (5) in the presence of Ca2+, yielding the same 5′-capped structure as is formed on eukaryotic mRNA by protein GTP:RNA guanylyltransferase. The RNA self-capping reaction involves the nucleophilic attack on the 5′ α-phosphate of the pppRNA (RNA with 5′-triphosphate) by the oxygen of β-phosphate of GDP with subsequent release of PPi:
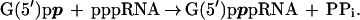 |
1 |
We now report that the self-capping RNA catalyst can react via attack on the α-phosphate of the pppRNA from an oxygen of any nucleophile’s terminal phosphate. The current finding generalizes this catalytic capability of RNA and thereby implies a route to versatile 5′ RNA posttranscriptional modification.
MATERIALS AND METHODS
Reagents.
Mes, 3′-NMPs, 5′-NMP, NDPs, NTPs, adenosine 5′-tetraphosphate (ppppA), guanosine 5′-tetraphosphates (ppppG), thiamine phosphate (thiamine-P), thiamine pyrophosphate (thiamine-PP), reduced and oxidized CoA (90–95% purity), acetyl CoA (90–95% purity), 3′-NADP (oxidized form, 80–90% purity), Nɛ-phosphate arginine (phosphoarginine), and 6-phosphate gluconic acid were from Sigma. FMN (oxidized form) was purchased from Fluka. Calf intestinal alkaline phosphatase (CIAP) was from New England Biolabs. Shrimp alkaline phosphatase (SAP) was from United States Biochemical. G-25 microspin gel filtration columns were from Pharmacia.
RNA Preparation.
Internally 32P-labeled 104-mer self-capping RNA (isolate 6 RNA), 5′-pppggggaguacgggagaggauacuacacgugUCCAUCCACCGUUGUGAUCGUCUGAAGUCAACCACAACAUGCCCUGACUCccauugcauguagcagaagcuuaaa-3′ (lowercase letters were from the constant regions of the original RNA pool), was transcribed by T7 RNA polymerase from its DNA template in the presence of 1 mM NTP and [α-32P]GTP as described (5). The pppRNA was purified by 8% denaturing PAGE (xylene cyanol = 46 cm) to single nucleotide resolution. After elution from the gel, the RNA was EtOH-precipitated and washed with 70:30 EtOH/H2O and redissolved in water. 5′-[γ-32P]-Labeled isolate 6 RNA (*pppRNA, the asterisk indicating the location of isotopic 32P) was prepared by transcription in the presence of 1 μM 5′-[γ-32P]GTP and purified by 8% PAGE (5).
Unlabeled ppRNAs (RNA with 5′-diphosphate), including tRNAAla, 95-mer self-aminoacylating RNA (isolate 29) (6), and 104-mer isolate 6 RNA, were transcribed by T7 RNA polymerase from corresponding DNA templates in the presence of 1 mM NTP and 20 mM GDP. The ppRNAs were purified by 8% denaturing PAGE (xylene cyanol = 15 cm) and recovered as described above. ppRNA concentrations were determined by UV absorbance at 260 nm (1 OD260 unit of RNA = 1,600 pmol tRNAAla, 1,300 pmol 95-mer RNA, and 1,200 pmol 104-mer RNA, respectively).
Reaction of Isolate 6 pppRNA with Phosphate-Containing Nucleophiles.
Internally 32P-labeled isolate 6 pppRNA (5 nM) was incubated with 1 mM of nucleotides, Nɛ-phosphate arginine, and 6-phosphate gluconic acid, respectively, for 60 min at 37°C in 20 mM Mes/2 mM Ca2+, pH 5.5. The reaction products were separated from reactant RNA by 8% denaturing PAGE (xylene cyanol = 46 cm) and visualized by a PhosphorImager (Bio-Rad, model GS525).
Reaction of coenzymes with isolate 6 pppRNA and analysis of the reaction products were conducted as above except for use of 4 mM coenzymes and 20 mM Ca2+.
Reaction of isolate 6 pppRNA with tRNAAla, 95-mer self-aminoacylating RNA (isolate 29) (6), and 104-mer isolate 6 RNA was performed by incubating 5 nM internally 32P-labeled isolate 6 pppRNA with 20 μM unlabeled ppRNA for 30 min at 37°C in 20 mM Mes/50 mM Ca2+, pH 5.5. The reaction products were separated from the reactant isolate 6 RNA by 8% denaturing PAGE (xylene cyanol = 8 cm) and visualized as above.
Reaction Products of Isolate 6 pppRNA.
Reaction products of isolate 6 pppRNA with terminal phosphate-containing nucleophiles were identified by analyzing the released non-RNA-containing product(s) and phosphatase resistance of nucleophile–RNA adducts. Isolate 6 *pppRNA (0.1 μM) was incubated with 1 mM each of ADP, ppppA, thiamine-P and thiamine-PP, Nɛ-phosphate arginine, and 6-phosphate gluconic acid for 60 min at 37°C in 20 mM Mes/2 mM Ca2+, pH 5.5. The reaction mixtures (1 μl) were spotted on a polyethyleneimine cellulose TLC plate and developed in 0.5 M potassium phosphate (pH 6.3) containing 20% methanol. 32P was visualized and quantitated by a PhosphorImager.
Reaction mixtures of internally 32P-labeled pppRNA with ADP and ppppA (under the same reaction conditions as described above) were passed through G-25 microspin gel filtration columns, EtOH-precipitated, recovered, and redissolved in water. Half of the solutions were treated with CIAP (5 units/μl, final concentration) in manufacturer-provided 1× buffer for 30 min at 37°C. As a control, internally 32P-labeled pppRNA was first treated with CIAP under the above conditions. The CIAP-treated RNA was gel-filtered, EtOH-precipitated, and resuspended in water. The RNA solution was then incubated with 1 mM ADP under the reaction conditions. The effect of CIAP on the RNA and the reaction products was analyzed by 8% PAGE (xylene cyanol = 46 cm).
RESULTS
Reaction of Isolate 6 pppRNA with Nucleotides.
Self-capping isolate 6 RNA (5) reacts with all nucleotides tested in a buffered Ca2+ solution (Fig. 1). The reaction products can be separated from the reactant RNA by 8% denaturing PAGE. Depending on the number of phosphates on the nucleophiles, the reaction products have different mobilities. Whereas 3′-NMP (Ap and Gp) products run close to unreacted RNA, the product has progressively slower mobility when the nucleophile contains more phosphates. For example, ppppA and ppppG products move much slower and are totally resolved. This suggests that added 5′-phosphates from the nucleophile are retained in the products. Reaction yields after 1 h at 37°C range from 30% (for NMPs) to 70% (for NDPs), with NTPs, ppppA, and ppppG yielding 50–60%.
Figure 1.

Reaction of nucleotides with isolate 6 pppRNA (5). The RNA was uniformly 32P-labeled by transcription in the presence of [α-32P]GTP and purified to single nucleotide resolution by 8% denaturing PAGE. The reactions were performed with 1 mM nucleotides and 5 nM pppRNA for 1 h at 37°C in 20 mM Mes/2 mM Ca2+, pH 5.5. The reaction products were separated from the reactant RNA by 8% denaturing PAGE (xylene cyanol = 46 cm) and visualized by a PhosphorImager. Np, nucleoside 3′-monophosphate; p(d)N, (2′-deoxyribo)nucleoside 5′-monophosphate; pp(d)N, (2′-deoxyribo)nucleoside 5′-diphosphate; ppp(d)N, (2′-deoxyribo)nucleoside 5′-triphosphate, where N = A, C, G, U, or T.
Reaction of Isolate 6 pppRNA with Coenzymes.
Coenzymes containing a terminal phosphate react with isolate 6 pppRNA and the products are well separated from unreacted RNA (Fig. 2A). As expected, the product of oxidized CoA (CoA dimer connected via a disulfide bond) is more retarded in the 8% gel than the coupling product of reduced CoA (monomer). Thiamine-P and thiamine-PP yield 35% and 65%, respectively, reaction of other coenzymes with isolate 6 pppRNA results in 5% (for 3′-NADP) to 20% (for FMN) products. In reactions with 3′-NADP and CoA and its derivatives, there is one reaction product moving between the top product band (coenzyme-RNA adduct) and bottom unreacted RNA band. This product is probably the result of RNA reaction with the adenosine nucleotide impurity (10–20%) in commercial NADP and CoA and its derivatives, as shown in Fig. 1.
Figure 2.

(A) Reaction of isolate 6 pppRNA with coenzymes. Internally 32P-labeled isolate 6 pppRNA (5 nM) and 4 mM coenzymes were incubated for 1 h at 37°C in 20 mM Mes/20 mM Ca2+, pH 5.5. The reaction products were analyzed as in Fig. 1. (B) Isolate 6 pppRNA reaction with coenzymes before and after phosphatase treatment. Reaction and analysis conditions were the same as above.
To confirm that a terminal phosphate of cofactors is responsible for the coupling reactions, we first treated reactants (GDP, thiamine-PP, CoA, NADP, and FMN) with SAP: 15 mM coenzymes, 10 units/μl SAP, 4 h at 37°C. SAP was deactivated by heating at 65°C for 15 min. The SAP-treated coenzymes were then incubated with isolate 6 pppRNA and analyzed under the same conditions as Fig. 2A. Fig. 2B shows that no reaction of coenzymes with isolate 6 pppRNA is detectable after phosphatase removal of coenzymes’ phosphate. Therefore the terminal phosphate of these molecules is the nucleophilic attacking group.
Reaction of Isolate 6 pppRNA with Nɛ-Phosphate Arginine and 6-Phosphate Gluconic Acid.
Nɛ-Phosphate arginine and 6-phosphate gluconic acid react with isolate 6 pppRNA to yield respectively 30% and 65% products within 60 min at 37°C (Fig. 3), indicating that the nucleotidic character of the above nucleophiles is not required for reaction.
Figure 3.
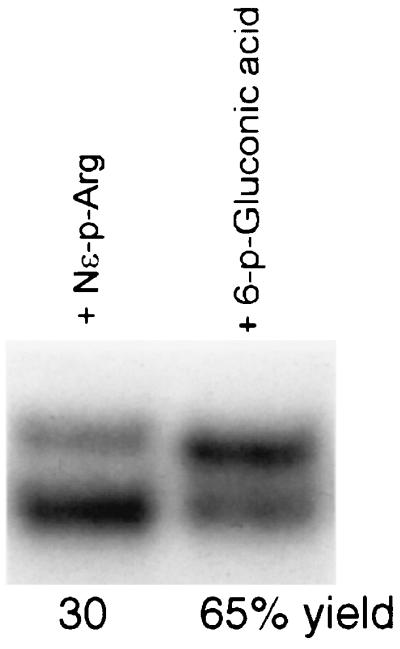
Reaction of isolate 6 pppRNA with Nɛ-phosphate arginine (phosphoarginine) and 6-phosphate gluconic acid. Reaction conditions and product analysis were the same as in Fig. 1.
Reaction Products of Isolate 6 pppRNA.
Reaction of isolate 6 *pppRNA with different nucleophiles uniformly releases the same 32P-labeled single product, *PPi. Fig. 4 shows the release of the 5′-pyrophosphate for near complete reactions with varied nucleophiles including nucleotides, coenzymes, an amino acid, and a sugar. While coupling yields with different nucleophiles vary (Fig. 1), the release of *PPi is more constant. This occurs because isolate 6 pppRNA also has a nucleophile-independent 5′-pyrophosphatase activity (5), and the pyrophosphatase tends to equalize *PPi release. Nevertheless, because only *PPi is released in the presence of different terminal phosphate-containing nucleophiles, the 5′ α-phosphate of isolate 6 pppRNA is always the reaction center, as was true in self-capping reaction with GDP (5). This is consistent with the required selection criteria that were based on 5′-pyrophosphate release by the RNA and subsequent circularization by T4 RNA ligase (5).
Figure 4.

TLC analysis of released 32P-containing product of isolate 6 *pppRNA (5′-[γ-32P]-labeled RNA), reacting with 1 mM ADP, ppppA, thiamine-P and thaimine-PP, Nɛ-phosphate arginine, and 6-phosphate gluconic acid under the same conditions as in Fig. 1. The TLC plate (polyethyleneimine cellulose) was developed in 0.5 M potassium phosphate (pH 6.3) containing 20% methanol. Locations of 32P were visualized by a PhosphorImager.
ADP–pppRNA and ppppA–pppRNA adducts are completely resistant to CIAP, as evidenced by the same product ratio and mobility (5′-hydroxyl, -monophosphate, and -triphosphate RNA have indistinguishable mobility in an 8% denaturing gel) with or without CIAP treatment (Fig. 5, lanes 3–6). Thus phosphates are added in internal linkage, inaccessible to phosphatase digestion. The pretreatment of pppRNA with CIAP completely abolishes reactivity (lane 7), indicating that CIAP was active and removed 5′-phosphates of the pppRNA under these conditions, and that 5′-triphosphate of the RNA is required for activity (5). When phosphate is not present in the nucleophile, there is no reaction (lane 2). The above combined data for varied reactants parallels the mechanism of the self-capping reaction (reaction 1) (5): the 5′ α-phosphate of pppRNA (isolate 6) is attacked by an oxygen of the terminal phosphate of a nucleophile to release *PPi and form a 5′-phosphate–phosphate linkage.
Figure 5.

Phosphatase resistance of reaction products of isolate 6 pppRNA. Conditions are described in Materials and Methods.
Reaction of Isolate 6 pppRNA with ppRNAs.
Different macromolecular RNAs [76-mer tRNAAla, 95-mer self-aminoacylating RNA (6), and 104 mer self-capping RNA isolate 6] with 5′-diphosphates can react with isolate 6 pppRNA (Fig. 6). With 20 μM concentration of ppRNAs acting as nucleophiles, the RNA–RNA coupling reactions yield 1% of product in which the two RNAs are coupled via a 5′–5′-triphosphate linkage (based on the above general reaction mechanism):
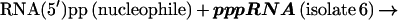 |
2 |
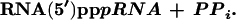 |
While these yields are low, the reactions have not been optimized. Low concentration (20 μM) of RNA nucleophiles may contribute to the low yields. However, it is remarkable that even base paired 5′ termini such as those of tRNA and 95-mer self-aminoacylating RNA (6) give similar detectable yields.
Figure 6.

Reaction of isolate 6 pppRNA with RNAs. Unlabeled 20 μM ppRNAs [tRNAAla, 95-mer self-aminoacylating RNA (6), and self-capping RNA (5)] were incubated with internally 32P-labeled isolate 6 pppRNA (5 nM) for 30 min at 37°C in 20 mM Mes/2 mM Ca2+, pH 5.5. The reaction products were separated from reactant RNA by 8% denaturing PAGE (xylene cyanol = 8 cm) and visualized by a PhosphorImager.
DISCUSSION
We have demonstrated that the isolate 6 pppRNA can react with all tested nucleophiles containing terminal phosphate. We have shown previously that the RNA self-capping reaction involves the nucleophilic attack on the 5′ α-phosphate of the pppRNA by the nonbridging oxygen of β-phosphate of GDP with subsequent release of pyrophosphate (reaction 1) (5). We now conclude that the general RNA mechanism involves the nucleophilic attack on the 5′ α-phosphate of the pppRNA by a nonbridging oxygen of the terminal phosphate with concurrent release of PPi:
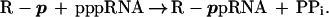 |
3 |
The number of phosphates in the phosphate–phosphate linkage of the reaction product depends on the number of phosphates linked to the terminal phosphate in the nucleophile. Thus, reactions of NMPs, NDPs, NTPs, and nucleoside 5′-tetraphosphates with isolate 6 pppRNA should yield 5′–5′-linked products with pyrophosphate, triphosphate, tetraphosphate, and pentaphosphate linkages, respectively.
Previously, reaction of 5′-phosphate terminated ribooligomers with artificially activated 5′ phosphate-imidazolide oligomers has been shown (7). These present coupling reactions with isolate 6 both use the natural biomolecular pyrophosphate as the leaving group and varied natural biomolecular phosphates in addition to RNA oligomers as the attacking groups. These present reactions are therefore very versatile, perhaps virtually universal: molecules as small as pyrophosphate (5) and as large as 104-mer ppRNA are reactive nucleophiles.
However, the number of phosphates linked to the terminal phosphate affects the coupling yield, with the order, pyrophosphate > triphosphate > tetraphosphate > monophosphate. The rest of the nucleophile does not have significant effect on the reaction (similar yields with different NMPs, similar yields with different NDPs, etc.), although large size tends to reduce the coupling yield (lower yields of CoA and NADP compared with nucleotide AMP). This suggests a large, open binding site for the nucleophile whose affinity may be based, for example, solely on the formation of a phosphate–Ca2+–phosphate complex. Nucleophiles are not restricted to phosphate monoesters (e.g., nucleotides, CoA, NADP, ppRNAs), because phosphoramidates (e.g., Nɛ-phosphate arginine) are also good nucleophiles for the coupling reaction.
A few modified nucleotidelike coenzymes such as nicotinamide guanine dinucleotide and dephosphorylated CoA have been incorporated into a modified Tetrahymena group I ribozyme through the sugar hydroxyl group (8). The ribozyme recognizes guanosine and adenosine with free 2′,3′-hydroxyl groups that must be introduced by the experimenters. On the other hand, incorporation of coenzymes into isolate 6 pppRNA is here extended to virtually all natural coenzymes containing terminal phosphate, such as thiamine-P (thiamine-PP), pyridoxal phosphate, and FMN.
Terminal phosphate-containing molecules are ubiquitous in modern cellular life. Among them are phosphoryl sources (nucleotides), precursors (nucleotides) for nucleic acids, energy sources (ATP, GTP, phosphoarginine, phosphocreatine, phosphoenol pyruvate), biosynthetic intermediates (glucose phosphate, glycerol phosphate, ribose phosphate, 5-phosphoribosyl-1-pyrophosphate, isopentenyl pyrophosphate), enzyme activity regulators (phosphorylated protein enzymes), enzyme coactivators (coenzymes), signaling molecules (guanosine 3′,5′-bispyrophosphate) (9, 10), and so forth. It is clear from present data that the isolate 6 active center will likely link all these terminal phosphate-containing molecules to RNA via specific 5′ phosphate–phosphate linkages.
It has been demonstrated that RNA can catalyze different reactions, including hydrolysis of aminoacyl ester (11), hydrolysis of an amide (12), self-aminoacylation (6), self-alkylation (13), amino acid transfer (14), and self-capping (5). However, the question remains whether RNA has the ability of catalyzing complex chemical reactions in the variety carried out by present protein enzymes. For this purpose, standard RNA is probably at a disadvantage, being composed of 4 monomers as compared with the standard 20 amino acid residues in protein enzymes. Therefore, limitation of functional groups of standard RNA plausibly restricts the range of RNA-catalyzed chemical reactions.
Self-incorporation of terminal phosphate-containing molecules (amino acids, sugars, nucleotides, coenzymes, etc.) into RNA provides a general increase in RNA functional groups beyond the four standard nucleotides. Such a strategy would have been available to RNA in an RNA world, enabling a broad range of chemical reactions. For example, coupled acyl-CoA RNA potentially makes the resulting RNA an acylating agent, allowing the acyl group to be transferred within the RNA molecule or to other molecules, such as ketoses. Incorporation of nicotinamide and flavin into RNA introduces facile oxidation/reduction reactions within the context of RNA catalysis, which makes it conceivable interconversions between alcohol, ketoses/aldehydes, and carboxylic acids. By recruiting thiamine-PP into RNA through self-incorporation, the chemistry of aldehyde transfer could be catalyzed by RNA.
Recruitment of new groups into RNA is not restricted to nucleotides and coenzymes. Incorporation of amino acids into RNA raises the interesting possibility that a selected subset of amino acid residues may have been incorporated into RNA via this route. In particular, phosphoserine, phosphothreonine, phosphotyrosine, and phosphoarginine might have been recruited into RNA (through phosphate–phosphate linkages) to perform limited “nucleoprotein” functions before the appearance of coded peptide synthesis.
Whereas RNA could perform related reactions using noncovalently bound coenzymes (15, 16) and amino acids (17–19), covalently coupling coenzymes and amino acids to RNA through phosphate–phosphate linkage liberates the RNA from the necessity of retaining the information for a strong binding site for the coenzymes and amino acids. Furthermore, covalently coupling coenzymes and amino acids to RNA may render the cofactors more sterically accessible and ensure their availability even under conditions where cofactors are diluted or depleted.
Bidirectional 5′–5′ triphosphate-linked RNA is the product of the coupling of a ppRNA molecule to pppRNA (isolate 6). Such a bidirectional RNA cannot be transcribed from a DNA or RNA template, and may accordingly have special biochemical and/or structural functions in an RNA world. A variety of 5′–5′ phosphate–phosphate-linked ribonucleotide structures are present in modern cells. The 5′-capped structure (a methylated guanosine linked to mRNA through a 5′–5′ triphosphate linkage) stabilizes eukaryotic mRNA and is required for translation (20, 21). Diadenosine 5′–5′ P1,P4-tetraphosphate is a signaling molecule that helps modern organisms to adjust synthesis of RNAs and proteins to environmental changes (22–24). The activity of isolate 6 RNA raises the possibility that these 5′–5′ phosphate–phosphate-linked RNA-like structures may be remnants of ancestral bidirectional 5′–5′ phosphate–phosphate-linked RNAs or 5′-capped RNA molecules.
Acknowledgments
We thank Mali Illangasekare for the DNA template for 95-mer self-aminoacylating RNA (6) and the members of our lab for comments on a draft manuscript. F.H. is the recipient of a National Institutes of Health postdoctoral fellowship (National Research Service Award). This work was also supported by National Institutes of Health Grants GM30881 and GM48080 to M.Y.
ABBREVIATIONS
- CIAP
calf intestinal alkaline phosphatase
- ppRNA
RNA with 5′-diphosphate
- pppRNA
RNA with 5′-triphosphate
- ppppA
adenosine 5′-tetraphosphate
- ppppG
guanosine 5′-tetraphosphate
- SAP
shrimp alkaline phosphatase
- thiamine-P
thiamine phosphate
- thiamine-PP
thiamine pyrophosphate
References
- 1.Gilbert W. Nature (London) 1986;319:618. [Google Scholar]
- 2.Joyce G F. Nature (London) 1989;338:217–224. doi: 10.1038/338217a0. [DOI] [PubMed] [Google Scholar]
- 3.Benner S A, Ellington A D, Tauer A. Proc Natl Acad Sci USA. 1989;86:7054–7058. doi: 10.1073/pnas.86.18.7054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.White H B. J Mol Biol. 1976;7:101–104. doi: 10.1007/BF01732468. [DOI] [PubMed] [Google Scholar]
- 5.Huang F, Yarus M. Biochemistry. 1997;36:6557–6563. doi: 10.1021/bi970475b. [DOI] [PubMed] [Google Scholar]
- 6.Illangasekare M, Sanchez G, Nickles T, Yarus M. Science. 1995;267:643–647. doi: 10.1126/science.7530860. [DOI] [PubMed] [Google Scholar]
- 7.Chapman K A, Szostak J W. Chem Biol. 1995;2:325–333. doi: 10.1016/1074-5521(95)90051-9. [DOI] [PubMed] [Google Scholar]
- 8.Breaker R R, Joyce G F. J Mol Evol. 1995;40:551–558. doi: 10.1007/BF00160500. [DOI] [PubMed] [Google Scholar]
- 9.Bremer H, Ehrenberg M. Biochim Biophys Acta. 1995;1262:15–36. doi: 10.1016/0167-4781(95)00042-f. [DOI] [PubMed] [Google Scholar]
- 10.Svitil A L, Cashel M, Zyskind J W. J Biol Chem. 1993;268:2307–11. [PubMed] [Google Scholar]
- 11.Piccirilli J A, McConnell T S, Zaug A J, Noller H F, Cech T R. Science. 1992;256:1420–1424. doi: 10.1126/science.1604316. [DOI] [PubMed] [Google Scholar]
- 12.Dai X, De Mesmaeker A, Joyce G F. Science. 1995;267:237–240. doi: 10.1126/science.7809628. [DOI] [PubMed] [Google Scholar]
- 13.Wilson C, Szostak J W. Nature (London) 1995;374:777–782. doi: 10.1038/374777a0. [DOI] [PubMed] [Google Scholar]
- 14.Lohse P A, Szostak J W. Nature (London) 1995;381:442–444. doi: 10.1038/381442a0. [DOI] [PubMed] [Google Scholar]
- 15.Burgstaller P, Famulok M. Angew Chem. 1994;33:1084–1087. [Google Scholar]
- 16.Lorsch J R, Szostak J W. Biochemistry. 1994;33:973–982. doi: 10.1021/bi00170a016. [DOI] [PubMed] [Google Scholar]
- 17.Yarus M. Science. 1988;240:1751–1758. doi: 10.1126/science.3381099. [DOI] [PubMed] [Google Scholar]
- 18.Connell G J, Illangasekare M, Yarus M. Biochemistry. 1993;32:5497–5502. doi: 10.1021/bi00072a002. [DOI] [PubMed] [Google Scholar]
- 19.Majerfeld I, Yarus M. Nat Struct Biol. 1994;1:287–292. doi: 10.1038/nsb0594-287. [DOI] [PubMed] [Google Scholar]
- 20.Adams J M, Cory S. Nature (London) 1975;255:28–33. doi: 10.1038/255028a0. [DOI] [PubMed] [Google Scholar]
- 21.Muthukrishnan S, Both G W, Furruichi Y, Shatkin A J. Nature (London) 1975;255:33–40. doi: 10.1038/255033a0. [DOI] [PubMed] [Google Scholar]
- 22.Varshavsky A. Cell. 1983;34:711–712. doi: 10.1016/0092-8674(83)90526-3. [DOI] [PubMed] [Google Scholar]
- 23.Bocher B R, Lee P C, Wilson S W, Cutler C W, Ames B N. Cell. 1984;37:225–232. doi: 10.1016/0092-8674(84)90318-0. [DOI] [PubMed] [Google Scholar]
- 24.Johnstone D B, Farr S B. EMBO J. 1991;10:3897–3904. doi: 10.1002/j.1460-2075.1991.tb04959.x. [DOI] [PMC free article] [PubMed] [Google Scholar]