

Abstract
Embryological and genetic evidence indicates that the vertebrate head is induced by a different set of signals from those that organize trunk–tail development1–6. The gene cerberus encodes a secreted protein that is expressed in anterior endoderm and has the unique property of inducing ectopic heads in the absence of trunk structures1. Here we show that the cerberus protein functions as a multivalent growth-factor antagonist in the extracellular space: it binds to Nodal, BMP and Wnt proteins via independent sites. The expression of cerberus during gastrulation is activated by earlier nodal-related signals in endoderm and by Spemann-organizer factors that repress signalling by BMP and Wnt. In order for the head territory to form, we propose that signals involved in trunk development, such as those involving BMP, Wnt and Nodal proteins, must be inhibited in rostral regions.
The principal activities of injected cerberus (cer) messenger RNA in Xenopus embryos are the inhibition of trunk mesoderm and neuralization of the ectoderm1. Xenopus cerberus protein (Cer) is structurally related to a family of cystine-knot secreted proteins that behave as antagonists of members of the transforming growth factor-β (TGF-β) family7,8. To determine the molecular mechanism of Cer function, we prepared soluble Xenopus Cer protein tagged with the Flag epitope. In animal cap cells expressing cer mRNA, Cer–Flag was secreted as a soluble protein of relative molecular mass (Mr) 46,000, designated Cer-long (Cer-L); a minor form of Mr 33,000, designated Cer-short (Cer-S), was also produced (Fig. 1a, lane 1). Transfected human 293T cells (Fig. 1a, lane 2) produced almost exclusively the proteolytically processed Cer-S form (Fig. 1b). To test whether Cerberus binds to any of the three main mesoderm-inducing candidate factors, we produced biologically active preparations of activin, Vg1 and Xnr-1 (Xenopus nodal-related-1, ref. 9), tagged with haemagglutinin (HA) at the amino terminus (Fig. 1c). In immunoprecipitation experiments Cer–Flag proteins bound Xnr-1 but not activin or Vg1 proteins, displaying a remarkable degree of specificity (Fig. 1d). In the presence of either Cer-L or Cer-S protein, the Xbra-inducing activity of Xnr-1 was blocked, but that of activin or Vg1 proteins was not (Fig. 1e). In animal cap assays, we found that at 10−9 M, Cer-L and Cer-S proteins titrated Xnr-1 signalling stoichiometrically, indicating that the affinity of the interaction must be in the subnanomolar range (Fig. 1f).
Figure 1.
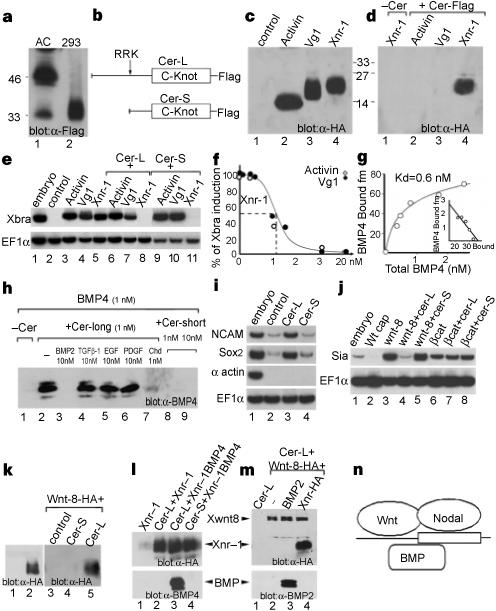
Cerberus protein binds to Xnr-1, BMP-4 and Xwnt-8. a, Cer–Flag secreted by Xenopus animal cap (AC) and cultured 293T cells. b, The two Cer protein products. c, HA-tagged activin, Vg1 and Xnr-1 secreted by Xenopus oocytes. d, Xnr-1 is bound specifically by Cer-S–Flag (lanes 2−4) or Cer-L–Flag (not shown). e, Cer binding inactivates Xnr-1 signalling. Animal cap explants were treated with oocyte control medium, activin (15 pM), Vg1 (0.3 nM) or Xnr-1 (1 nM) proteins, either alone (lanes 2−5) or together with 5 nM Cer-S (lanes 6−8) or Cer-L (lanes 9−11). f, Cer inhibits Xnr-1 with high affinity; 2 nM Xnr-1 was incubated with increasing concentrations of Cer-S (closed circles) or Cer-L (open circles). g, Cer-L–Flag binds BMP-4 with a KD of 0.6 nM. h, Binding of Cer-L–Flag to BMP-4 is competed for by BMP-2 but not by TGF-β-1, EGF or PDGF; Cer-S does not bind BMP-4. i, Cer-L (10 nM), but not Cer-S (20 nM), is a direct neural inducer of animal cap cells sensitized by brief dissociation–re-aggregation (but not of intact caps). j, Cer inhibits Xwnt8 but not β-catenin mRNA; induction of Siamois, a target of Wnt signalling, was assayed at stage 10+. k, Lanes 1, 2, soluble Xwnt-8–HA protein is secreted by mRNA-injected oocytes. Lanes 3−5, binding of Xwnt-8-HA (5 nM) to Cer–Flag proteins (10 nM). l, m, The Xwnt-8–HA, Xnr-1–HA and BMP binding sites in Cer-L are independent. Cer-L–Flag (2 nM) was bound to Xnr-1 (1 nM) or Xwnt-8 (2 nM) and competed with BMP-4 (10 nM), BMP-2 (27 nM) or Xnr-1 (8 nM). n, Multiple ligand-binding sites on Cerberus.
The neuralizing activity of cer mRNA can be antagonized by BMP-4 (refs 1,7). We performed co-immunoprecipitation experiments to test whether Xenopus Cerberus proteins bind BMPs directly. Cer-L–Flag protein bound BMP-4, with a dissociation constant (KD) of 0.6 nM (Fig. 1g). This interaction was highly specific, as it could be displaced by a 10-fold excess of the related molecule BMP-2, but not by TGF-β-1, epidermal growth factor (EGF) or platelet-derived growth factor (PDGF; Fig. 1h, lanes 3−6). In biological assays, Cer-L protein had neutralizing activity (Fig. 1i, lane 3), whereas the proteolytically processed Cer-S protein did not bind BMP-4 (even in 10-fold excess, Fig. 1h, lane 9) and did not induce neural markers (Fig. 1i, lane 4).
An additional property of cer mRNA is that it blocks the induction of secondary axes by Xwnt-8 mRNA4. We found that in animal caps, full-length cer mRNA inhibited Xwnt-8 but not β-catenin signalling (Fig. 1j), suggesting that Cer-L might antagonize Xwnt-8 extracellularly. To test this directly, we incubated Xwnt-8–HA protein produced in oocytes with Cer-L–Flag or Cer-S–Flag and immunoprecipitated with anti-Flag antibody. We found that Xwnt-8 bound to Cer-L, but not to the Cer-S protein (Fig. 1k, lanes 3−5). Thus, the Cer-L protein is a trifunctional antagonist that can bind Xnr-1, BMP and Xwnt-8 in solution.
The binding of Xnr-1 to Cer-L–Flag could not be displaced by excess BMP (Fig. 1l) and Xwnt-8 binding was not displaced by excess BMP or Xnr-1 (Fig. 1m). This suggests that Cer-L binds each ligand at independent binding sites (Fig. 1n). In addition, the biochemical studies show that Cer-L is proteolytically processed into the Cer-S protein containing the cystine-knot (Fig. 1b), which retains only the anti-Xnr-1 activity. This proteolytic cleavage could be important in regulating Cerberus function in vivo; a regulatory proteolytic step has been described in the case of chordin (Chd), in which active BMP ligands are released from inactive Chd/BMP complexes10.
Xenopus Cerberus has three distinct inhibitory activities and, as shown below, each one is required for the induction of ectopic head-like structures. To study the phenotypic effects of specifically inhibiting nodal-related signals in the context of the Xenopus embryo, we generated a construct encoding a secreted form of Cer-S, designated cer-S. Injection of cer-S mRNA into single blastomeres did not induce ectopic heads but instead gave rise to anterior defects in the endogenous head, including cyclopia (not shown). Embryos in which each blastomere was injected at the four-cell stage with cer-S mRNA lacked axial structures and the expression of the trunk mesoderm markers Xbra and Xwnt-8 was blocked (Fig. 2a–d). In addition to maintaining a mesodermal fate, nodal-related signals are required for the formation of anterior endoderm: expression of endogenous cerberus was blocked by cer-S mRNA (Fig. 2e, lane 2) and induced by Xnr-1 mRNA (Fig. 2f). Interestingly, endodermal patterning by Xnr-1 mRNA appears to function early in development, as injection of an Xnr-1 DNA construct expressed after mid-blastula was unable to induce cerberus (Fig. 2f, lanes 6−8). Taken together, these gain- and loss-of-function studies support the view that nodal-related signals have a central role in the patterning of anterior endoderm and in trunk mesoderm formation in Xenopus, in agreement with genetic studies in other vertebrates11–17.
Figure 2.
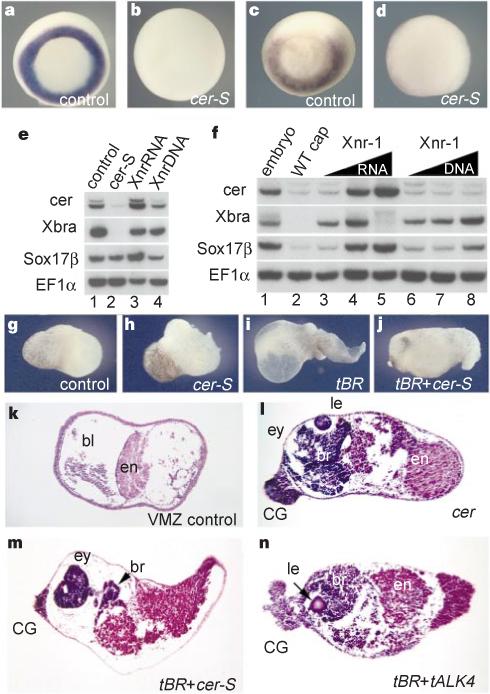
Phenotypic effects of cer-S mRNA, an inhibitor of nodal-related signals. Embryos were injected radially at the 4-cell stage. a–d, In situ hybridization for Xbra (a,b) and Xwnt-8 (c,d) in wild type and in embryos injected with 120 pg of cer-S mRNA per blastomere. e, Anterior endoderm formation requires early nodal-related signals in injected embryos: endogenous cerberus transcripts are inhibited by cer-S mRNA (200 pg total) and induced by Xnr-1 mRNA (50 pg). f, Dual function for Xnr-1; Xnr-1 mRNA (6,12, 24 pg), but not pCS2–Xnr-1 DNA (20, 40, 80 pg), can induce cer expression in animal caps. g–n, Head induction by simultaneous repression of BMP and nodal signals in explants of ventral marginal zone (VMZ). g, k, Wild-type VMZs consisting histologically of endoderm (en) and blood (bl). h, cer-S mRNA (150 pg); i, tBR mRNA (600 pg); j, m, co-injection of tBR mRNA (600 pg) and cer-S mRNA (150 pg) generates head-like structures (76%, n = 17), similar to those produced by cerberus mRNA (l, 50%, n = 10), consisting only of anterior structures with cement gland (CG), brain (br) and eye (ey) containing a lens (le). n, Blocking BMP and Nodal signalling by tBR (600 pg) and tALK4 (1.5 ng) RNAs is sufficient to induce head-like structures.
Since the anti-nodal activity of cer-S was not sufficient to induce ectopic head-like structures, we next tested it in combination with anti-BMP reagents in explants of ventral marginal zone (VMZ), which normally develops into postero-ventral mesoderm containing blood (Fig. 2g, k). Microinjection of cer-S or of tBR, a dominant-negative BMP receptor mRNA6, did not cause head formation (Fig. 2h, i). However, injection of combined cer-S and tBR produced a striking change in VMZ fate into head-like structures (Fig. 2j, m). The explants contained a large cyclopic eye (often with crystalline lens), brain, cement gland and endoderm, but no notochord or somites, indicating the generation of head organizer activity. Indistinguishable head-like structures were formed by injection of full-length cer mRNA (Fig. 2l). Co-injection of a dominant-negative putative nodal receptor18, tALK4, with tBR also induced head-like structures (Fig. 2n), showing that the cer constructs do not have an instructive role. Although these experiments in VMZs show that simultaneous inhibition of nodal-related and BMP signalling is sufficient for head specification, they do not indicate whether the anti-Wnt activity of cerberus is required for head formation. Indeed, in VMZs the main Wnt, Xwnt-8, is inhibited at the gene-expression level in embryos injected with cer-S (Fig. 2d) or tBR mRNA19.
To test whether triple inhibition of Wnt, nodal-related and BMP signalling is required for ectopic head formation, we carried out epistatic combinatorial experiments in the context of the whole embryo. Co-injection of chd (anti-BMP) and cer-S (anti-nodal) into a ventral blastomere resulted in ectopic heads containing a cyclopic eye (58%, n = 115); this induction was blocked by co-injection of Xwnt-8 DNA (Fig. 3a, b). Similarly, selective inactivation of the anti-nodal activity of full-length cer mRNA by co-injection of a constitutively active ALK-4 receptor18, even at doses that only weakly promote trunk mesoderm formation, abolished ectopic head induction by cer (Fig. 3c, d). Mimicking BMP signalling by co-injecting a constitutively active BMP receptor construct also antagonized ectopic head induction (Fig. 3e). These epistatic experiments indicate that all three inhibitory activities of cerberus are required for the formation of ectopic head structures.
Figure 3.
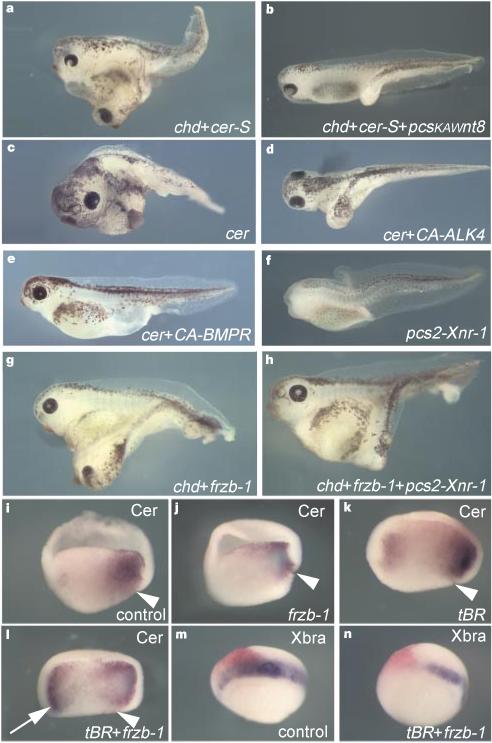
Head induction requires the triple inhibition of nodal-related, Wnt and BMP signals. Embryos were injected into one ventral blastomere with the indicated mRNAs or DNAs at the 4-cell stage. a, Ectopic head induction by a combination of chd (50 pg) and cer-S (200 pg) mRNA (58%, n = 115). b, pCSKA–Xwnt-8 DNA (50 pg) antagonizes head induction by chd/cer-S (1% ectopic heads, n = 111). c, Ectopic heads formed by injection of 150 pg cer mRNA (57%, n = 66) are abolished in d and e by co-injection of 1 ng CA–ALK4 mRNA (0%, n = 36) or 400 pg of CA–BMPR mRNA (0%, n = 21). f, Dorsal injection of pCS2-Xnr-1 (32 pg) causes head reduction (53%, n = 60). g, Co-injection of chd (25 pg) and Frzb-1 (200 pg) mRNAs mediates formation of a complete secondary axis with cyclopic head (49%, n = 175). h, Co-injection of 16 pg of pCS2–Xnr-1 DNA inhibits head induction by chd/Frzb-1 (5%, n = 133). i–n, Simultaneous repression of BMP and Wnt signalling leads to ectopic activation of cer in endoderm and concomitant downregulation of Xbra in mesoderm. Embryos were cut sagittally in two halves after fixation and before whole-mount hybridization. Arrowhead indicates the dorsal blastopore lip. Frzb-1 mRNA (300 pg) synergizes with tBR mRNA (800 pg) in upregulating cer in the ventral endoderm (arrow). Note in n the inhibition of Xbra in the injected area, correlating with the activation of cer (an anti-Nodal agent) observed in l.
Niehrs and co-workers have shown that simultaneous inhibition of BMP and Wnt signalling is sufficient to induce secondary axes containing heads with cyclopic eyes4. This is puzzling, for this head induction would occur without apparent inhibition of nodal-related signals. We confirmed these results using co-injection of chd10 and Frzb-1 (ref. 20) mRNAs, which encode secreted inhibitors specific for BMP and Wnt proteins, respectively (Fig. 3g). Head formation by chd and Frzb-1 was suppressed by co-injection of Xnr-1 DNA (Fig. 3h and 3f), suggesting that inhibition of nodal signalling could also be involved in this experimental situation, perhaps by activating transcription of cerberus. As shown in Fig. 3i–l, co-injection of Frzb-1 and tBR mRNA led to strong ectopic expression of cerberus in ventral endoderm at levels comparable to those seen with endogenous cerberus mRNA (Fig. 3l, arrow), with concomitant inhibition of the trunk mesodermal marker Xbra near the site of injection (Fig. 3m, n).
In mice, the anterior visceral endoderm (AVE) has a fundamental role in head formation2,14,16,21 and is the topological equivalent of the anterior endoderm that expresses cerberus in Xenopus2,22. Robertson and colleagues have shown that nodal activity in the AVE is required for head formation in chimaeric embryos14, whereas we report here that inhibition of nodal-related signalling by cerberus is required for head formation. These paradoxical observations can be reconciled by the proposal that, in mice, nodal signalling14–17 may be required in the AVE to express secreted factors that antagonize the formation of mesoderm in the overlying epiblast16,17, in which the head forms. In Xenopus, we have found evidence that cerberus expression requires an early nodal-related function mimicked by injection of Xnr-1 mRNA, but not DNA (Fig. 2e, f). This early signal, which correlates with the expression of Xnr-1 in endoderm at late blastula9, leads to the expression of a multivalent antagonist, Cerberus, which inhibits the function of nodal-related signals in rostral regions at later stages, preventing trunk mesoderm from forming in the head field. As shown in Fig. 4, the expression of cerberus in anterior endoderm at the mid-gastrula stage1 results from earlier signals provided by TGF-β proteins expressed in the endoderm9,23 as well as by organizer-specific secreted inhibitors of the Wnt5,20,21 and BMP6 pathways. In this view, the secretion of Cerberus into the extracellular space would be an important downstream event that would lock the head-organizer programme in place by simultaneously antagonizing three signalling pathways involved in trunk formation, thereby restricting the trunk territory to the posterior part of the body.
Figure 4.
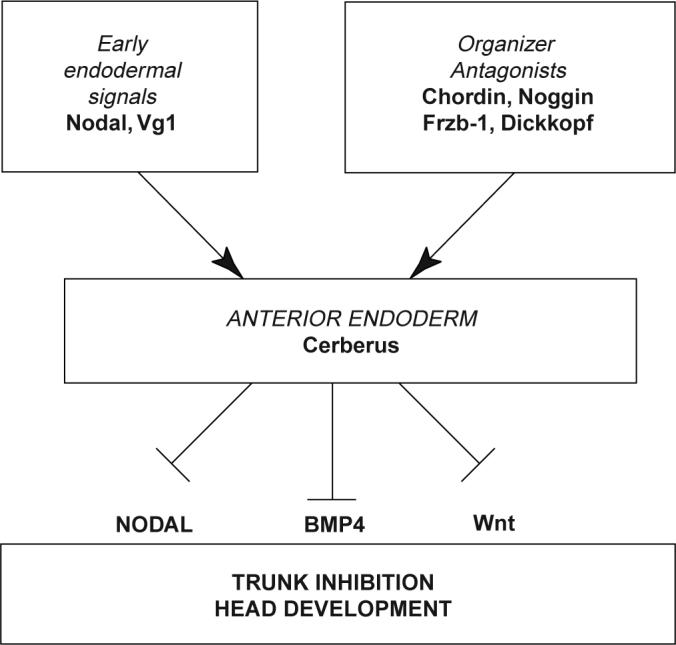
Model of the formation and function of anterior endoderm in Xenopus head development.
Methods
DNA constructs
To generate HA-tagged soluble Xnr-1, Vg1, activin and Xwnt-8, we designed a secreted protein-expression cassette vector that contains the pre-pro region and proteolytic cleavage site of activin-βB24 followed by the sequence RGL(YPYDVPDYA)LE (pCS2–proAct–HA). The two amino acids (LE) after the HA tag were inserted to provide an XhoI cloning site. The mature regions of signalling proteins were generated by the PCR reaction using primers flanked by XhoI and XbaI sequences and subcloned into the HA expression cassette. Mature signalling sequences started at residue C259 of activin-βB, S247 of Vg1, N286 of Xnr-1 and V26 of Xwnt-8. pCS2–cer-S was generated by PCR, deleting residues R26 to A116 of cer.
Protein binding
Soluble Xwnt-8, Xnr-1, Vg1 and activin were secreted by manually defolliculated Xenopus oocytes injected with 50 ng of mRNA and incubated in 5−10 μl per oocyte of OR2 medium for 2 days24. Cer-S was prepared by transfection of 293T cells10; Cer-L was prepared from animal caps of embryos injected with 1 ng cer–Flag mRNA. Typically, 10 caps were dissociated at stage 9 and incubated in 100 μl of Ca–Mg-free medium for 3 h. For immunoprecipitations, protein mixtures were incubated on ice (2 h) in 20 mM Tris–HCl pH 7.5, 150 mM NaCl, 1 mM CaCl, 1 mM MgCl, 2% glycerol, 0.1% BSA and 0.1% each of the detergents Triton X-100, CHAPS and octylglucoside (Pierce). Protein-A beads pre-bound with anti-Flag polyclonal antibody (Santa Cruz Inc.) were then added and samples incubated with end-over-end rotation for 1 h at 4 °C. After three 5-min washings in the same buffer, protein complexes were analysed by electrophoresis.
Acknowledgements
We thank L. S. Mathews, N. Ueno, R. Moon, D. Melton and K. Cho for gifts of plasmids, K. Masuhara for anti-BMP4 mAb, Genetics Institute for BMPs, and members of the De Robertis laboratory for comments. S.P., E.A., S.B. and H.G were supported by Telethon, ARC, MSTP and DFG, respectively. This work was supported by a grant from the NIH. E.M.D.R. is a Howard Hughes Medical Institute investigator.
References
- 1.Bouwmeester T, Kim SH, Sasai Y, Lu B, De Robertis EM. Cerberus is a head-inducing secreted factor expressed in the anterior endoderm of Spemann's organizer. Nature. 1996;382:595–601. doi: 10.1038/382595a0. [DOI] [PubMed] [Google Scholar]
- 2.Thomas P, Beddington R. Anterior primitive endoderm may be responsible for patterning the anterior neural plate in the mouse embryo. Curr. Biol. 1996;6:1487–1496. doi: 10.1016/s0960-9822(96)00753-1. [DOI] [PubMed] [Google Scholar]
- 3.Shawlot W, Behringer RR. Requirement for Lim1 in head-organizer function. Nature. 1995;374:425–430. doi: 10.1038/374425a0. [DOI] [PubMed] [Google Scholar]
- 4.Glinka A, Wu W, Onichtchouk D, Blumenstock C, Niehrs C. Head induction by simultaneous repression of Bmp and Wnt signalling in Xenopus. Nature. 1997;389:517–519. doi: 10.1038/39092. [DOI] [PubMed] [Google Scholar]
- 5.Glinka A, et al. Dickkopf-1 is a member of a new family of secreted proteins and functions in head induction. Nature. 1998;391:357–362. doi: 10.1038/34848. [DOI] [PubMed] [Google Scholar]
- 6.Harland R, Gerhart J. Formation and function of Spemann's organizer. Annu. Rev. Cell Dev. Biol. 1997;13:611–667. doi: 10.1146/annurev.cellbio.13.1.611. [DOI] [PubMed] [Google Scholar]
- 7.Hsu DR, Economides AN, Wang X, Eimon PM, Harland RM. The Xenopus dorsalizing factor gremlin identifies a novel family of secreted proteins that antagonize BMP activities. Mol. Cell. 1998;1:673–683. doi: 10.1016/s1097-2765(00)80067-2. [DOI] [PubMed] [Google Scholar]
- 8.Belo JA, et al. Cerberus-like is a secreted factor with neuralizing activity expressed in the anterior primitive endoderm of the mouse gastrula. Mech. Dev. 1997;68:45–57. doi: 10.1016/s0925-4773(97)00125-1. [DOI] [PubMed] [Google Scholar]
- 9.Jones CM, Kuehn MR, Hogan BLM, Smith JC, Wright CVE. Nodal-related signals induce axial mesoderm and dorsalize mesoderm during gastrulation. Development. 1995;121:3651–3662. doi: 10.1242/dev.121.11.3651. [DOI] [PubMed] [Google Scholar]
- 10.Piccolo S, et al. Cleavage of Chordin by Xolloid metalloprotease suggests a role for proteolytic processing in the regulation of Spemann organizer activity. Cell. 1997;91:407–416. doi: 10.1016/s0092-8674(00)80424-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Feldman B, et al. Zebrafish organizer development and germ-layer formation require nodal-related signals. Nature. 1998;395:181–185. doi: 10.1038/26013. [DOI] [PubMed] [Google Scholar]
- 12.Sampath K, et al. Induction of the zebrafish ventral brain and floor plate requires cyclops/nodal signalling. Nature. 1998;395:185–189. doi: 10.1038/26020. [DOI] [PubMed] [Google Scholar]
- 13.Rebagliati MR, Toyama R, Haffter P, Dawid IB. cyclops encodes a nodal-related factor involved in midline signaling. Proc. Natl Acad. Sci. USA. 1998;95:9932–9937. doi: 10.1073/pnas.95.17.9932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Varlet I, Collignon J, Robertson EJ. Nodal expression in the primitive endoderm is required for the specification of the anterior axis during mouse gastrulation. Development. 1997;124:1033–1044. doi: 10.1242/dev.124.5.1033. [DOI] [PubMed] [Google Scholar]
- 15.Conlon FL, et al. A primary requirement for nodal in the formation and maintenance of the primitive streak in the mouse. Development. 1994;120:1919–1928. doi: 10.1242/dev.120.7.1919. [DOI] [PubMed] [Google Scholar]
- 16.Waldrip WR, Bikoff EK, Hoodless PA, Wrana JL, Robertson EJ. Smad2 signaling in extraembryonic tissues determines anterior-posterior polarity of the early mouse embryo. Cell. 1998;92:797–808. doi: 10.1016/s0092-8674(00)81407-5. [DOI] [PubMed] [Google Scholar]
- 17.Nomura M, Li E. Smad2 role in mesoderm formation, left-right patterning and craniofacial development. Nature. 1998;393:786–790. doi: 10.1038/31693. [DOI] [PubMed] [Google Scholar]
- 18.Chang C, Wilson PA, Mathews LS, Hemmati-Brivanlou A. A Xenopus type I activin receptor mediates mesodermal but not neural specification during embryogenesis. Development. 1997;124:827–837. doi: 10.1242/dev.124.4.827. [DOI] [PubMed] [Google Scholar]
- 19.Hoppler S, Moon RT. BMP-2/4 and Wnt-8 cooperatively pattern the Xenopus mesoderm. Mech Dev. 1998;71:119–129. doi: 10.1016/s0925-4773(98)00004-5. [DOI] [PubMed] [Google Scholar]
- 20.Leyns L, Bouwmeester T, Kim SH, Piccolo S, De Robertis EM. Frzb-1 is a secreted antagonist of Wnt signaling expressed in the Spemann Organizer. Cell. 1997;88:747–756. doi: 10.1016/s0092-8674(00)81921-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Rhinn M, et al. Sequential roles for Otx2 in visceral endoderm and neuroectoderm for forebrain and midbrain induction and specification. Development. 1998;125:845–856. doi: 10.1242/dev.125.5.845. [DOI] [PubMed] [Google Scholar]
- 22.Bouwmeester T, Leyns L. Vertebrate head induction by anterior primitive endoderm. BioEssays. 1997;19:855–863. doi: 10.1002/bies.950191005. [DOI] [PubMed] [Google Scholar]
- 23.Henry GL, Melton DA. Mixer, a homeobox gene required for endoderm development. Science. 1998;281:91–96. doi: 10.1126/science.281.5373.91. [DOI] [PubMed] [Google Scholar]
- 24.Kessler DS, Melton DA. Induction of dorsal mesoderm by soluble, mature Vg1 protein. Development. 1995;121:2155–2164. doi: 10.1242/dev.121.7.2155. [DOI] [PubMed] [Google Scholar]