

Abstract
This study investigated the effect of strength training on the mechanical properties of the human patella tendon of older individuals. Subjects were assigned to training (n = 9; age 74.3 ± 3.5 years, body mass 69.7 ± 14.8 kg and height 163.4 ± 9.1 cm, mean ±s.d.) and control (n = 9; age 67.1 ± 2 years, body mass 73.5 ± 14.9 kg and height 168.3 ± 11.5 cm) groups. Strength training (two series of 10 repetitions at 80 % of five-repetition maximum) was performed three times per week for 14 weeks using leg extension and leg press exercises. Measurements of tendon elongation during a ramp isometric knee extension were performed before and after training and control periods in vivo using ultrasonography. Training caused a decreased tendon elongation and strain at all levels of force and stress (P < 0.01). Baseline tendon elongation and strain at maximal tendon load were 4.7 ± 1.1 mm and 9.9 ± 2.2 %, respectively (maximum force: 3346 ± 1168 N; maximum stress: 40 ± 11 MPa). After training, these values decreased to 2.9 ± 1.2 mm and 5.9 ± 2.4 % (P < 0.01), respectively (maximum force: 3555 ± 1257 N; maximum stress: 42 ± 11 MPa). Tendon stiffness increased by 65 % (2187 ± 713 to 3609 ± 1220 N mm−1; P < 0.05) and Young's modulus increased by 69 % (1.3 ± 0.3 to 2.2 ± 0.8 GPa; P < 0.01). As a result of these changes, the rate of torque development increased by 27 % (482.8 ± 302.5 to 612.6 ± 401 N m s−1; P < 0.01) following training. No significant changes occurred in any measured variables in the control group (P > 0.05). This study shows for the first time that strength training in old age increases the stiffness and Young's modulus of human tendons. This may reduce the risk of tendon injury in old age and has implications for contractile force production and the rapid execution of motor tasks.
It is relatively well established that ageing is associated with a marked loss of muscle mass and strength (Frontera et al. 2000; Klein et al. 2001), which can limit the performance of activities of daily living (Feder et al. 2000; Khan et al. 2001). A very important, yet scarcely investigated factor in the performance of all functional activities, is the tendon. The tendon is responsible for the transmission of contractile forces from muscle to bone, allowing movement to occur. The tendon however, is not an inextensible tissue, but deforms in response to the applied load in a manner dependent upon its mechanical properties (Elliott, 1965; Butler et al. 1978; Dunn & Silver, 1983). In vitro animal and human tissue studies have shown that ageing is associated with a decrease in the tensile stiffness of tendons and collagenous tissues (Nachemson & Evans, 1968; Tkaczuk, 1968; Noyes & Grood, 1976). In vitro, connective tissues of elderly individuals display a reduced failure stress compared to younger specimens, despite equivalent or even larger tissue cross-sectional areas (Noyes & Grood, 1976). In fact, it has been suggested that there is a progressive deterioration of tendons with ageing, illustrated by a decrease in collagen content and an increase in the more extensible elastin (Verzár, 1957; Vogel, 1991). Recently, human tendon mechanical properties have been studied in vivo using ultrasonography (Ito et al. 1998; Maganaris & Paul, 1999, 2000a,b; Kubo et al. 2001a,b; Magnusson et al. 2001). By applying these methods, it has been shown that in line with previously mentioned in vitro reports, the human gastrocnemius tendons of elderly subjects are more compliant than those of young adults (Maganaris, 2001).
Tendon mechanical properties have a key role in the time course of torque development. In fact, tendon stiffness affects the time required to stretch the series elastic component (SEC) and will therefore affect both the electromechanical delay (EMD) and the rate of torque development (RTD). This is because the EMD depends only partially on the duration of the activation- contraction coupling process, but mostly on the time required for stretching the SEC. Similarly, the RTD depends on the force-velocity properties of the contractile component and on SEC stiffness. Hence, both the EMD and the RTD are factors closely reflecting the stiffness of the tendon and entire SEC (Jewell & Wilkie, 1958). Therefore, the functional implication of reduced tendon stiffness in older individuals is likely to be a slowing in the rate of force development, affecting the time necessary to decelerate the body mass, a factor relating to the prevention of falls. Increased tendon compliance would result in larger strains at any given force, increasing the likelihood of tendon strain injury. The incidence of patella tendon strain injury is in fact higher in older individuals (Kelly et al. 1984; Kannus & Józsa, 1991).
One of the first suggestions that young human tendons may be capable of modifying their material properties in response to increased loading, was the discovery of an increased stiffness of the extensor carpi radialis tendons due to greater loading in the dominant hand (Cronkite, 1936). Subsequently, animal studies have demonstrated that the tendon is capable of adapting to a training stimulus by increasing its stiffness (Woo et al. 1980, 1982; Buchanan & Marsh, 2001). Recently, in vivo ultrasound-based measurements of young adults’ tendon structures have shown an increase in stiffness following a period of isometric (Kubo et al. 2001a) and isotonic resistance training (Kubo et al. 2002). Measurements in both of these studies, however, were determined from the displacement of the aponeurosis. Although conflicting research exists (e.g. Zuurbier et al. 1994), the aponeurosis may be a more compliant structure compared with the free tendon (Lieber et al. 1991; Maganaris & Paul, 2000a,b). Despite these reports on the response of young adult tendons to training, no data are currently available on the adaptation of elderly tendons to regimes of increased loading. Hence, the aim of the present study was to investigate the effects of a 14 week strength training programme on the mechanical properties of the human patella tendon in elderly individuals.
METHODS
Subjects
After receiving ethics committee approval from the Centre for Biophysical and Clinical Research into Human Movement at the Manchester Metropolitan University, 18 subjects (10 females and 8 males) gave written informed consent to participate in this study. All subjects received medical clearance from their general practitioners prior to undertaking the training programme. Subjects did not have any neurological or musculoskeletal disorder preventing their participation. None of the subjects had previously taken part in any type of resistance training. Five females and four males (n = 9, age 74.3 ± 3.5 years, body mass 69.7 ± 14.8 kg and height 163.4 ± 9.1 cm, mean ±s.d.) were assigned randomly to the training group, with the remaining subjects assigned to the control group (n = 9, age 67.1 ± 2 years, body mass 73.5 ± 14.9 kg and height 168.3 ± 11.5 cm). All procedures used in this study conformed with the Declaration of Helsinki.
Strength training programme
The exercise consisted of progressive isotonic resistance training performed three times per week for a period of 14 weeks. All exercises were performed using Technogym (Gambettola, Italy) resistance machines after having completed a 5–10 min warm up on a cycle ergometer at 60–65 % of age-predicted maximum heart rate (Londeree & Moeschberger, 1982). Specific exercises performed were: (1) the bilateral leg extension for the knee extensor muscle group and (2) the bilateral leg press for the hip, knee and ankle extensors, both exercises loading the patella tendon. Other exercises were performed to provide the individuals with an overall increase in muscle strength; these were the bilateral calf raise, chest press, seated row, abdominal crunch and lower back extension. During the first 2 weeks of training, subjects became familiarised with the exercises and gradually increased their workload until they were able to lift the highest possible load for five consecutive times (five-repetition maximum, 5 RM) for each exercise. It was felt that the 5 RM was more appropriate for elderly individuals who were unaccustomed to this type of strenuous exercise, as an alternative to the 1 RM. Thereafter the 5 RM was determined every 2 weeks to assess any increases in strength, so that the same relative training intensity could be maintained. All exercises were performed with the concentric phase of the contraction lasting ∼2 s followed by a ∼3 s eccentric phase. One specific warm-up set of 15 repetitions was performed for each exercise at an intensity of 45 % of the 5 RM followed by 2–3 sets of static stretching for specific muscle groups. For each of the lower limb exercises, two training sets of ∼10 repetitions were performed, initially at an intensity of ∼60–70 % of the 5 RM progressing to 80 % of the 5 RM within 1–3 weeks, dependent upon the subjects’ capabilities. This intensity (80 % of the 5 RM) corresponded to approximately 70–75 % of the predicted 1 RM (Brzycki, 1993). During the sessions, all subjects wore heart rate monitors and if the heart rate rose to within ∼15 beats of the age-predicted maximum heart rate the exercise was stopped. A recovery period of ∼3 min was introduced between sets. Subjects were observed very closely during all exercises to ensure the correct technique was performed and to provide verbal encouragement to maintain motivation. Compliance to training was very high; only ∼7 % of sessions were missed due to holidays, illness, etc.
Study design
All subjects were tested at baseline. Subjects were tested again after 14 weeks of training. The control group continued their usual activities and were then tested after 14 weeks of normal activity.
Measurement of maximal knee extension and flexion torque
Maximal isometric knee extension and flexion torque were measured at a 90 deg knee angle (full extension = 0 deg), with the hip angle at 85 deg (supine position = 0 deg) using an isokinetic dynamometer (Cybex NORM, New York, USA). Straps were positioned at the hip, shoulders and over the right thigh to prevent any extraneous movement. All measurements were performed on the right lower limb. Subjects had previously visited the laboratory on at least one occasion to become familiarised with the procedures involved. Subjects were instructed to perform a maximal isometric knee extension contraction by gradually increasing their effort until they reached maximal torque after ∼4 s. It was expected that a voluntary contractile deficit would exist; therefore the interpolated twitch technique was applied to enable measurement of tendon elongation at maximal force. When the voluntary torque peaked, a superimposed maximal double twitch with 50 μs pulse width and 50 ms inter-stimulus gap was applied (Fig. 1). The doublet was generated by an electrical stimulator (Digitimer stimulator, model DS7, Welwyn Garden City, UK) and was delivered to the quadriceps muscle group through two 7.5 cm × 12.5 cm self-adhesive electrodes (Versa-Stim, CONMED, New York, USA) placed on the distal and proximal regions of the thigh. If the superimposed twitch increased the voluntary torque, the torque and ultrasound data (see section Measurement of patella tendon elongation) produced during the application of the stimulus were then incorporated in the analysis. The voluntary activation capacity of the quadriceps muscle group was calculated as: maximum voluntary torque/(maximum voluntary torque + maximal interpolated torque) as previously reported (Kent-Braun & Ng, 1999; Kent-Braun et al. 2002).
Figure 1. Torque traces from one subject before (A) and after (B) training.
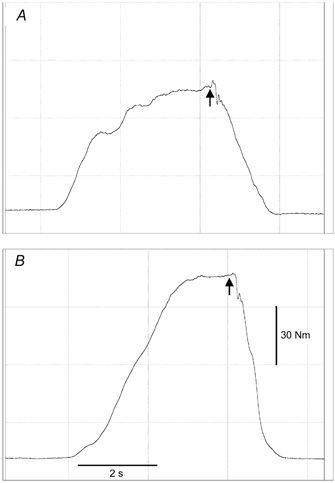
Arrows indicate the application of a superimposed double twitch.
Signals of torque and electromyographic (EMG) activity were displayed on the screen of a computer (Macintosh, G4), interfaced with an acquisition system (Acknowledge, Biopac Systems, Inc., CA, USA) used for analog-to-digital conversion, sampling at a frequency of 2000 Hz.
Measurement of electromyographic activity
Electromyographic activity was assessed from the vastus lateralis (VL) and the long head of the biceps femoris (BF) muscles. Two self-adhesive Ag-AgCl electrodes 10 mm in diameter (Neuroline, Medicotest A/S, Oelstykke, Denmark), were placed in a bi-polar configuration with a constant inter-electrode distance of 20 mm at a site corresponding to the distal one-third of the length of the muscle (Zipp, 1982). To minimise cross-talk from adjacent muscles, electrodes were placed along the mid-sagittal axis of each muscle, guided by axial plane ultrasound scanning. Reference electrodes were placed on the lateral tibial condyle. Electrode placement was always preceded by shaving, skin abrasion and cleansing with an alcohol-based tissue pad. Skin impedance was always reduced below 5000 Ω. The location of all electrodes with respect to anatomical landmarks was recorded and also traced onto an acetate sheet to ensure identical placement on subsequent sessions. The raw EMG signal was pre-amplified and filtered using high- and low-pass filters set at 10 and 500 Hz, respectively. To determine the level of antagonistic coactivation of the knee flexors, the root mean square (RMS) EMG activity of the BF muscle was measured during a ramp isometric knee extension contraction over 50 ms time periods at intervals of 10 % of maximal torque. To determine the maximal activation of the BF muscle when acting as an agonist, a maximal knee flexion isometric contraction was performed at the position studied. The RMS BF muscle EMG activity was measured at the time point of maximal torque over a 50 ms time period, taking into account the electromechanical delay, and was then normalised for a 1 s time period. The antagonistic torque of the knee flexors during a knee extension contraction was calculated assuming a linear EMG-torque relationship (Lippold, 1952), from the EMG-torque relationship of the biceps femoris muscle when acting as an agonist. The addition of this torque to the measured knee extension torque yielded the torque produced by the quadriceps alone.
Measurement of electromechanical delay and rate of torque development
The EMD and RTD were both assessed during an isometric knee extension contraction in which subjects were instructed to reach their maximal contraction torque as rapidly as possible. Subjects were asked to fully relax their thigh muscles prior to initiating this contraction in an attempt to minimise baseline noise from the EMG signal. The EMD was defined as the time lag between the onset of electrical activity and the initiation of torque development. The onset of electrical activity was defined as a rise of 15 μV above baseline in the VL muscle RMS EMG activity. The initiation of torque development was defined as a rise of 2 N m above baseline. The RTD was calculated from the gradient of the torque-time relationship over the first 100 ms after the onset of torque development.
Measurement of patella tendon moment arm length
To allow calculation of tendon forces, the patella tendon moment arm length was assessed using a 0.2 T magnetic resonance imaging (MRI) scanner (Esaote Biomedica, Genoa, Italy). Sagittal-plane scans were taken using a T1-weighted spin-echo sequence with the following scanning parameters: time to echo (TE): 26 ms; time of repetition (TR): 850 ms; field of view (FOV): 180 mm × 180 mm; matrix: 256 × 192; 4 mm slice thickness; and 0.4 mm gap. The patella tendon moment arm length was defined as the perpendicular distance from the midpoint of the patella tendon to the tibio-femoral contact point (Baltzopoulos, 1995; Kellis & Baltzopoulos, 1999). Due to constraints in the size of the coil, it was only possible to image the knee joint at full knee extension. The previously reported ratio of the patella tendon moment arm length between 0 and 90 deg of knee flexion (Baltzopoulos, 1995) was used to estimate the moment arm length at 90 deg of knee flexion for each subject. Measurements of the patella tendon moment arm length have previously demonstrated good reproducibility (r = 0.94) when performed on different days (Smidt, 1973).
Calculation of patella tendon force
Patella tendon force was calculated from the moment equilibrium equation around the knee joint. The sum of the net knee extension torque measured and the estimated antagonistic knee flexor torque was divided by the estimated patella tendon moment arm length at 90 deg to yield patella tendon force.
Measurement of patella tendon elongation
Elongation of the patella tendon was assessed using real-time B-mode ultrasonography (ATL-HDI 3000, Bothwell, USA) during a ramp isometric knee extension performed in ∼4 s with the application of a superimposed double twitch upon maximal contraction (Fig. 1). Measurements were taken after four preconditioning contractions to ensure reproducibility of the measurements (Rigby, 1964; Wood et al. 1988). A 7.5 MHz linear array transducer was positioned in the sagittal-plane over the patella tendon, on an eco-absorptive external marker fixed on the skin with surgical tape (Fig. 2).Any movement of the line cast by the external marker on the ultrasound image indicated movement of the transducer with respect to the scanned structure and this trial would therefore be omitted from any further analysis. Measurements of displacement were analysed at 10 % intervals of maximal torque, using digitising software (NIH image version 1.61, National Institutes of Health, USA). The mean of three measurements for each image was calculated.
Figure 2. Sagittal-plane scans of the patella tendon (PT) at rest, during isometric contraction at 50 % of maximal and at maximal tendon force.
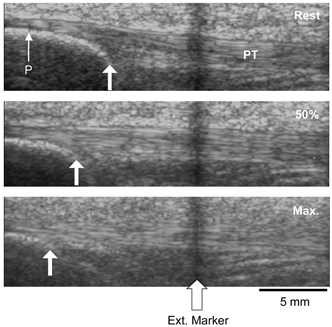
Arrows indicate displacement of the apex of the patella (P) during contraction with respect to an echo-absorptive external marker fixed on the skin.
Measurement of tendon cross-sectional area and calculation of tendon stress
Tendon cross-sectional area (CSA) was measured in the resting state at a knee joint angle of 90 deg after preconditioning, from axial-plane ultrasound images taken at 25, 50 and 75 % of patella tendon length. Three measurements were averaged at each site and the mean of the three sections was used for calculation of tendon stress. Patella tendon stress was calculated by dividing tendon force by tendon CSA.
Measurement of tendon length and calculation of tendon strain
Patella tendon length was assessed at rest after preconditioning, from sagittal-plane ultrasound images and measured from the apex of the patella to the superior aspect of the tibial tuberosity. Tendon strain was calculated as the ratio (%) of tendon displacement to the initial resting tendon length.
Calculation of tendon stiffness and Young's modulus
Patella tendon stiffness (N mm−1) was calculated over each 10 % interval of maximal tendon force from the gradient of the force-elongation relation. Young's modulus (GPa) over the respective stress intervals was calculated by multiplying the stiffness value estimated by the ratio of tendon length to tendon CSA.
Measurement reliability
To assess inter-day reliability, measurements of tendon CSA, length, sub-maximal and maximal tendon elongations, were performed on two separate days in a sub-sample of six elderly subjects. Measurements taken on separate days by the same experimenter showed good agreement: the intra-class correlation coefficients (ICCs) were 0.99 for patella tendon CSA, patella tendon length, sub-maximal and maximal tendon elongations. Typical error was 1.5 mm2 for tendon CSA, 0.6 mm for tendon length, 0.1 mm for sub-maximal tendon elongation and 0.1 mm for maximal tendon elongation.
Statistical analysis
Independent samples Student's t-tests were used to test for differences between the training and control groups at baseline on all of the measured variables. A 2 × 2 factorial analysis of variance (ANOVA) was used to analyse for differences in time (pre; post) and group (training; control) for all of the measured variables. Level of significance was set at P < 0.05. The Scheffé procedure was used for post hoc analysis where necessary. Paired-samples Student's t-tests were used to analyse for differences in 5 RM strength before and after training. Using a one-way random effects model, ICCs were applied to assess the reproducibility of measurements performed on separate days. Typical error was used to assess the random error between reproducibility measurements taken on different days. Typical error was calculated from the equation: s.d.diff/√2 (Hopkins, 2000). Here s.d.diff is the standard deviation of the difference scores between days 1 and 2. Data are reported as means ±s.d.
RESULTS
No significant differences existed between the two groups at baseline for any of the measured variables (P > 0.05). Body mass did not significantly change in either the training (pre: 69.7 ± 14.8 kg; post: 67.2 ± 15.2 kg) or control (pre: 73.5 ± 14.9 kg; post: 75.1 ± 15.4 kg) groups (P > 0.05).
Five-repetition maximum strength
The 5 RM for the leg extension exercise increased by 14 % from 43.5 ± 12 to 49.4 ± 14.1 kg after the 14 weeks of training (P < 0.01). The 5 RM for the bi-lateral leg press exercise increased by 23 % from 178.3 ± 44.7 to 219.1 ± 55.4 kg following training (P < 0.01; Table 1).
Table 1.
Measured and calculated variables pre- and post-training and control periods
| Training group | Control group | |||
|---|---|---|---|---|
| Pre | Post | Pre | Post | |
| Leg extension 5 RM (kg) | 43.5(12) | 49.4(14.1)** | — | — |
| Leg press 5 RM (kg) | 178.3(44.7) | 219.1(55.4)** | — | — |
| Maximal tendon force (N) | 3346(1167.6) | 3555.4(1256.7) | 3254.2(789.6) | 3051(694.5) |
| BF coactivation (μV s−1) | 25.8(9.15) | 24.9(11.3) | 33.2(12.8) | 29.3(13.7) |
| Maximal tendon stress (MPa) | 40(11) | 42.1(10.5) | 39.7(15.6) | 37(12.9) |
| Maximal tendon strain (%) | 9.9(2.2) | 5.9(2.4)** | 10.2(3) | 10.3(1.7) |
| Tendon stiffness (N mm−1) | 2187.4(713.1) | 3609.6(1220)* | 2246.5(726) | 2254.7(684.9) |
| Young's modulus (GPa) | 1.3(0.3) | 2.2(0.8)** | 1.3(0.6) | 1.3(0.4) |
| Tendon CSA (mm2) | 84.2(23.6) | 84.3(21.8) | 87.5(26.6) | 87.2(24.6) |
| Tendon length (mm) | 47.6 (4.7) | 48.3 (4.8) | 48.4 (6.6) | 48.4 (5.9) |
| EMD (ms) | 4.4 (5.4) | 42.9 (5.2) | 50.6 (5.1) | 51.9 (4.4) |
| RTD (N m s−1) | 482.8(302.5) | 612.6(401)** | 501.6(320.3) | 437.2(291.7) |
| Muscle activation capacity | 0.95(0.03) | 0.97(0.03) | 0.91(0.14) | 0.89(0.11) |
5 RM, 5-repetition maximum; BF, biceps femoris; CSA, cross-sectional area; EMD, electromechanical delay; RTD, rate of torque development. Values are means (s.d.); * and ** significantly different from pre (P < 0.05 and P < 0.01, respectively).
Patella tendon force, antagonistic coactivation and voluntary muscle activation capacity
Maximal isometric patella tendon force (taking into account the antagonistic coactivation of the knee flexors) was not significantly different after training (pre: 3346 ± 1167.6 N; post: 3555.4 ± 1256.7 N; P > 0.05) and control (pre: 3254.2 ± 789.6 N; post: 3051 ± 694.5 N; P > 0.05) periods. Antagonistic coactivation from the BF muscle during a maximal isometric knee extension contraction did not change significantly in the training (pre: 25.8 ± 9.15; post: 24.9 ± 11.3 μV s−1; P > 0.05) or control (pre: 33.2 ± 12.8 μV s−1; post: 29.3 ± 13.7 μV s−1; P > 0.05) groups (Table 1). The quadriceps voluntary activation capacity was unchanged following the training and control periods (P > 0.05; Table 1).
Patella tendon force-elongation and stress-strain relations
The apex of the patella moved proximally with increasing force during a ramp isometric knee extension contraction (Fig. 2). Patella tendon elongation and strain at baseline increased in a curvilinear fashion to 4.7 ± 1.1 mm and 9.9 ± 2.2 % respectively, at 3346 ± 1167.6 N and 40 ± 11 MPa (maximal tendon force and stress, respectively) in the training group (Fig. 3 and Fig. 4). The curvilinear force-elongation and stress-strain relations were maintained following training; however, the curves were shifted to the left, indicating a decreased elongation and strain at every 10 % level of maximal tendon force and stress (P < 0.01; Fig. 3 and Fig. 4, respectively). After training, tendon elongation at maximal tendon force decreased by 1.8 mm, from 4.7 ± 1.1 mm at 3346 ± 1167.6 N to 2.9 ± 1.2 mm at 3555.4 ± 1256.7 N (P < 0.01). Tendon strain at maximal tendon stress (42.1 ± 10.5 MPa) decreased by 40 % from 9.9 ± 2.2 to 5.9 ± 2.4 % following training (P < 0.01). In contrast, the tendon force-elongation and stress-strain relations were unchanged in the control group (P > 0.05; Fig. 3 and Fig. 4, respectively). Patella tendon stress and the dimensions of the tendon were not significantly changed after the training and control periods (P > 0.05; Table 1).
Figure 3. PT force-elongation relation for the training and control groups.
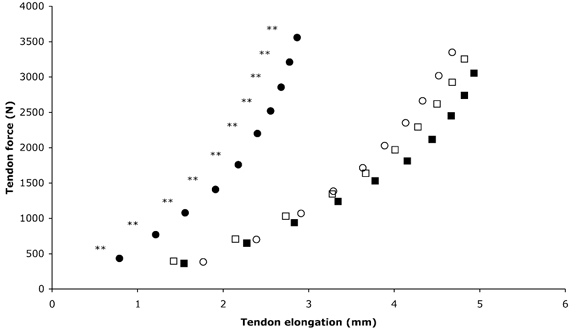
○, Pre, training; •, post, training; □, pre, control; ▪, post, control. ** Significantly (P < 0.01) reduced elongation after training. Values are means; maximal s.d. values are shown in Table 1.
Figure 4. PT stress-strain relation for the training and control groups.
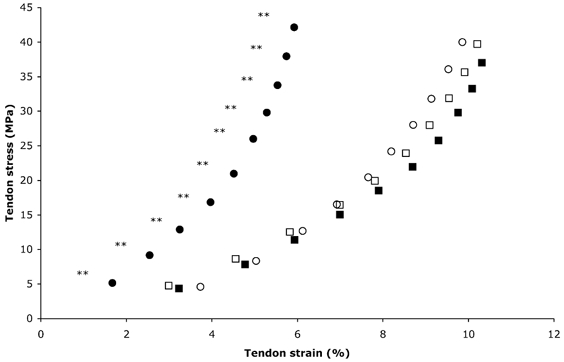
○, Pre, training; •, post, training; □, pre, control; ▪, post, control. ** Significantly (P < 0.01) reduced strain after training. Values are means; maximal s.d. values are shown in Table 1.
Patella tendon stiffness and Young's modulus
Patella tendon stiffness increased with increasing tendon force in both groups at baseline. After training, patella tendon stiffness increased over each 10 % force interval (P < 0.05), whilst this remained unchanged in the control group (P > 0.05; Fig. 5). Tendon stiffness over the 0–10 % force interval increased by 168 % following training (pre: 231.6 ± 105.6; post: 620.4 ± 226.9 N mm−1; P < 0.01), with no change in the control group (pre: 306.7 ± 111.1; post: 248.4 ± 77.8 N mm−1; P > 0.05). Tendon stiffness over the 90–100 % force interval increased by 65 % after training (pre: 2187.4 ± 713.1; post: 3609.6 ± 1220 N mm−1; P < 0.05), but was unchanged after the control period (pre: 2246.5 ± 726; post: 2254.7 ± 684.9 N mm−1; P > 0.05). Young's modulus during maximal tendon stress at baseline was similar in the training (1.3 ± 0.3 GPa) and control (1.3 ± 0.6 GPa) groups. After training Young's modulus increased by 69 % (pre: 1.3 ± 0.3 GPa; post: 2.2 ± 0.8 GPa; P < 0.01), but remained unchanged in the control group (P > 0.05; Fig. 6).
Figure 5. PT stiffness for the training and control groups.
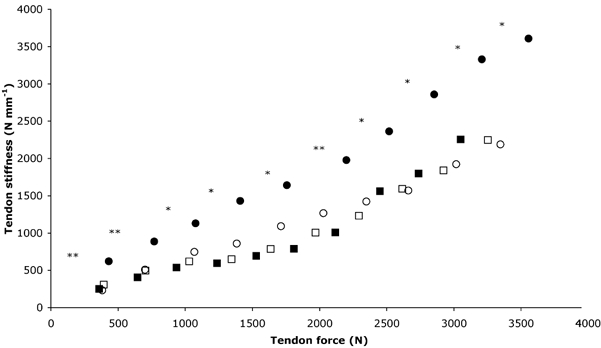
○, Pre, training; •, post, training; □, pre, control; ▪, post, control. Stiffness is significantly (*P < 0.05 and **P < 0.01, respectively) increased after training. Values are means; maximal s.d. values are shown in Table 1.
Figure 6. PT Young's modulus for the training and control groups.
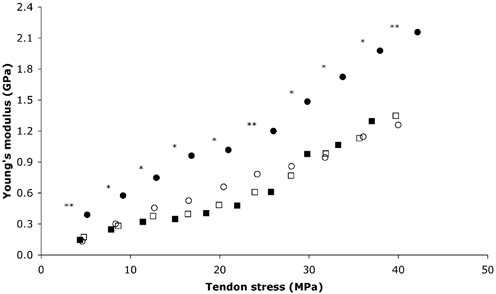
○, Pre, training; •, post, training; □, pre, control; ▪, post, control. Significantly (*P < 0.05 and **P < 0.01, respectively) increased Young's modulus after training. Values are means; maximal s.d. values are shown in Table 1.
Electromechanical delay and rate of torque development
The EMD measured from the VL muscle was unchanged in the training (pre: 43.4 ± 5.4 ms; post: 42.9 ± 5.2 ms; P > 0.05) and control (pre: 50.6 ± 5.1; post: 51.9 ± 4.4 ms; P > 0.05) groups. The RTD increased by 27 % after training (pre: 482.8 ± 302.5; post: 612.6 ± 401 N m s−1; P < 0.01), whilst remaining unaltered (P > 0.05) in the control group (pre: 501.6 ± 320.3 N m s−1; post: 437.2 ± 291.7 N m s−1; Table 1).
DISCUSSION
This study shows for the first time that the mechanical properties of the human patella tendon in older individuals can be altered by a period of strength training. Both the stiffness and Young's modulus of the patella tendon increased following training. The patella tendon force-elongation relation at baseline in both the training and control groups displayed the characteristic curvilinear relationship, with relatively large elongations for a small application of force at the initial concave portion followed by smaller relative elongations as the force increased up to maximum (Fig. 3). As a result of strength training, there was a pronounced shift of the entire force-elongation curve to the left. This demonstrates that for each 10 % level of tendon force there was a reduced tendon elongation after training. This reduction in tendon elongation was greater towards the higher force region, suggesting that the tendon had shown the greatest adaptation in this region. Tendon elongation reduced following training by 1 mm at 10 % of maximal force and by 1.8 mm at maximal force.
The tendon stress-strain relation displayed a very similar pattern to that of the force-elongation relation in both groups at baseline (Fig. 4). Training caused a marked left-shift of the stress-strain curve, indicating reduced tendon strain at each 10 % level of tendon stress. The magnitude of adaptation in the tendon was greater in the higher stress region, illustrated by the greater divergence of the pre- and post-training curves in this region. Tendon strain at maximal stress was reduced from 9.9 % before to 5.9 % after training. Tendon strain values measured in vivo at maximal contraction in humans have previously been reported to be between 3 and 13 % (Maganaris & Paul, 1999, 2000a,b; Magnusson et al. 2001; Muramatsu et al. 2001; Kubo et al. 2001a,b, 2002). This variation is likely to reflect differences in the structures studied. Aponeuroses generally demonstrate larger strains compared to the tendon (Lieber et al. 1991; Maganaris & Paul, 2000a,b). However, differences may exist between tendons performing different physiological functions (Woo et al. 1980). Although the applied loads may vary, in vitro measurements of strain have indicated that tendons function within the elastic range only if strain remains below ∼4 % (Rigby, 1964; Elliott, 1965; Diamant et al. 1972; Ker, 1981) and that tendon fracture may occur at strains of ∼8 % (Ker et al. 1988) or ∼13 % (Wren et al. 2001). Failure strains reported from in vitro testing of human patella tendons are as high as 23–30 % (Haut & Powlison, 1990). Strain values from in vitro tensile tests appear to be generally lower than the majority of values reported from human in vivo testing. However, caution must be stressed when drawing comparisons between these different testing methods as the applied loads may vary. In vitro tests of tendon stiffness may be subject to certain inherent errors, such as specimen slippage due to insufficient clamping and changes in tissue properties due to the extraction and storage processes (Cronkite, 1936; Verzár, 1957; Walker et al. 1964; Bennett et al. 1986; Haut & Powlison, 1990). Furthermore tendons often fail at or near to the site of clamping (Cronkite, 1936; Elliott, 1965; Butler et al. 1978), which is likely to represent premature specimen failure because of an artificially high stress created at the site of the clamps. In the present study, tendon strain at baseline (∼10 %) was higher than most of the values reported from in vivo human testing. However, it should be stressed that all of the measurements to date have been performed on young subjects. It has been demonstrated in vivo in our laboratory that the human gastrocnemius tendon of elderly subjects has a reduced stiffness as compared to that of young adults (Maganaris, 2001). Patella tendon strain observed after training in the present study (∼6 %) approaches the values previously obtained from in vivo testing of young adults. This may indicate training-induced reversibility of the more compliant tendons of the elderly towards stiffer tendons, comparable to those of young adults. A large tendon strain represents a greater predisposition to tendon strain injuries (McMaster, 1933; Bennett et al. 1986; Wren et al. 2001); therefore after strength training in old age, the risk of tendon strain injuries may be reduced. It is clear from the appearance of the force-elongation relation that the patella tendon operated within the elastic range, supported by the observation that when load was removed, the tendon returned to its pre-loading resting length.
Errors in tendon elongation would be incurred if the reference point on the patella did not move solely in the sagittal-plane with respect to the transducer. Studies of patella tracking indicate that a small degree of mediolateral movement of the patella occurs as the lower leg moves through its range of motion (Nagamine et al. 1995; Sheehan et al. 1999). However, the mediolateral movement of the patella caused by the minimal joint rotation during an isometric contraction in the present study is likely to be negligible (Nagamine et al. 1995; Sheehan et al. 1999). Errors in tendon elongation could also be incurred due to unwanted knee joint rotations during the isometric contraction performed. The effect of joint rotation on tendon displacement was investigated in a sub-sample of six elderly subjects. The change in knee joint angle, measured using an electrical goniometer during a maximal isometric knee extension, was found to be 6.7 ± 3 deg in the knee extension direction. Given the negligible tendon displacement observed during passive knee joint rotation over this range (Fig. 7), it was considered unnecessary to correct for such minimal displacements.
Figure 7. PT displacement in the proximal direction during passive knee extension.
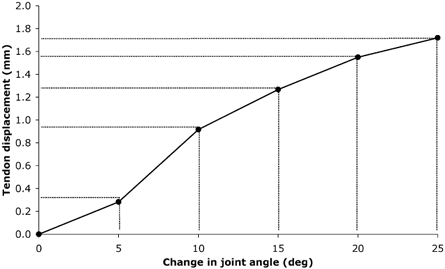
Values are means, (n = 6).
Patella tendon stress at maximal contraction was ∼40 MPa in both groups at baseline. Maximal tendon stress did not change following training because the maximal force was only slightly increased and the tendon cross-sectional area was unaltered. Tendon stress in humans, calculated from in vivo measurements performed on young adults, has been reported to be ∼25 MPa in the tibialis anterior tendon (Maganaris & Paul, 1999) and ∼42 MPa in the triceps surae tendon structures (Magnusson et al. 2001). In vitro tests have demonstrated tendon stress at failure to be between 86 and 100 MPa (Bennett et al. 1986; Wren et al. 2001). These data suggest that the majority of mammalian tendons possess safety factors of ∼4 or even as much as ∼8 (Walker et al. 1964; Ker et al. 1988). Assuming a maximum failure stress of ∼100 MPa (Bennett et al. 1986), the estimated safety factor of the elderly patella tendon from the present study would be ∼2.5, which is lower than that reported from isolated specimens. During rapid eccentric contraction producing forces exceeding those during an isometric contraction (Aagaard et al. 2000), the elderly patella tendon will be closer to the ultimate stress and is therefore likely to be at the greatest risk of injury.
The stiffness of the patella tendon increased over all force levels after training, but the greatest increase in stiffness occurred in the higher force region (Fig. 5). The stiffness values between 90 and 100 % of maximal force of 2187 N mm−1 before and 3610 N mm−1 after training are substantially higher than the values of 161 N mm−1 reported during maximal joint torque in the tibialis anterior tendon (Maganaris & Paul, 1999) and 480 N mm−1 during maximal force in the human triceps surae tendon structures (Magnusson et al. 2001). The relative approach used in the present study to calculate tendon stiffness does not allow comparison over the same absolute force intervals. However, the absolute approach encompasses a number of problems such as missing data points, unequal subject numbers in each condition and omitting data from the higher force regions in order to normalise to the weakest subject. It was the intention of the present study to use all of the generated data, an approach not permitted using absolute force intervals. To incorporate all data points and calculate stiffness over absolute force levels, addition analyses were performed. Missing data points were calculated from polynomial fitting of the tendon force-elongation relation, and stiffness values were calculated over 600 N intervals. This approach showed similar increases in tendon stiffness after training as compared to the relative approach (128 % increase over 0–600 N; 49 % increase over 600–1200 N; 92 % increase over 1200–1800 N; P < 0.01), indicating that the relative approach followed in the present study reflects realistically the training-induced changes in tendon stiffness.
In both groups, antagonistic coactivation of the biceps femoris during maximal isometric knee extension was ∼40 % of the activity when this muscle was acting as an agonist during maximal isometric knee flexion. This indicates that failure to account for the effects of antagonistic coactivation would dramatically underestimate tendon force, tendon stiffness and Young's modulus.
It is noteworthy that training did not cause any measurable change in patella tendon cross-sectional area, indicating that tendon stiffness increased due to a change in the material properties of the tendon and not due to tendon hypertrophy. This is confirmed by the 69 % increase in Young's modulus after training (Fig. 6). The Young's modulus value at baseline was in line with previous in vivo (Maganaris & Paul, 1999) and in vitro (Ker, 1981; Bennett et al. 1986) reports. Increased tendon stiffness after training without any measurable tendon hypertrophy is in agreement with findings from animal studies, showing that tendon stiffness increases in response to increased loading without any change in the size of the tendon in adults (Rollhäuser, 1954), whilst immature tendons adapt to increased loading primarily through tendon hypertrophy (for review see Elliot, 1965). Increased tendon stiffness has been reported from other animal studies using exercise to provide increased levels of tendon loading (Viidik, 1967, 1969; Woo et al. 1980, 1982; Wood et al. 1988). The exact mechanisms responsible for the training-induced increased tendon stiffness are not apparent from the present study. However, based on findings from animal studies it is suggested that both collagen turnover and the packing density of collagen fibrils may have increased (Heikkinen & Vuori, 1972; Woo et al. 1980; Michna & Hartmann, 1989). Also, alterations in the crimp angle of collagen fibrils, affecting tendon stiffness, have been reported to occur following exercise training in rat tendons (Wood et al. 1988). An increased tendon collagen content and decreased crimp angle caused by the increased levels of loading would increase tendon stiffness. If the water content of the tendon had increased in response to training, this may further increase the tensile stiffness of the tendon (Cohen et al. 1976; Hooley et al. 1980; Haut & Haut, 1997).
The EMD is the time lag between the onset of muscle activity and the initiation of torque development. This time period represents the time taken for excitation-contraction coupling of the contractile component and stretching of the tendon and in-series elastic component to occur. The EMD was unchanged after training in the present study. The RTD also encompasses these two main elements, closely reflecting the stiffness of the tendon and series elastic component (Jewell & Wilkie, 1958). The 27 % increase in the RTD measured after training in the present study is consistent with the increased tendon stiffness. This has implications for the rapid execution of motor tasks in old age, such as decelerating the body centre of mass, descending stairs and reacting to a fall. The RTD is also influenced by the force-velocity characteristics of the muscle fibres. This is because muscle contraction is never isometric in vivo and some shortening of muscle fascicles occurs as they stretch the aponeurosis and tendon (Narici et al. 1996; Maganaris et al. 1998; Ito et al. 1998). Therefore, if muscle fascicle length were to increase after training, the addition of in-series sarcomeres may increase shortening velocity and could be another factor contributing to the increased RTD. Such a possibility is supported by recent observations on the effects of strength training in old age on fascicle length (Narici et al. 2000). The increased patella tendon stiffness observed after training may result in a reduced fascicle shortening, which would cause a shift in the optimal angle for force generation, towards resting muscle length. This suggests that training-induced changes in tendon stiffness can actually alter the force-length relation of muscle. Information on the quadriceps force-length relation would provide further insights into the functional relevance of increased patella tendon stiffness following strength training.
In conclusion, 14 weeks of strength training increased patella tendon stiffness and Young's modulus, in older humans. As a consequence, tendon elongation and strain were reduced, decreasing the possibility of tendon strain injury in old age. The rate of torque development was increased after training and it is expected that this would have a positive effect on functional performance by affording a more rapid execution of motor tasks.
Acknowledgments
This study was supported by Italian Space Agency (ASI) funds. The authors would like to express their appreciation to Technogym for providing the resistance machines used in this study. We feel particularly indebted to the participants for their outstanding motivation and adherence to the programme.
REFERENCES
- Aagaard P, Simonsen EB, Anderson JL, Magnusson SP, Halkjaer-Kristensen J, Dyhre-Poulsen P. Neural inhibition during maximal eccentric and concentric contraction: effects of resistance training. J Appl Physiol. 2000;89:2249–2257. doi: 10.1152/jappl.2000.89.6.2249. [DOI] [PubMed] [Google Scholar]
- Baltzopoulos V. A videofluoroscopy method for optical distortion correction and measurement of knee-joint kinematics. Clin Biomech (Bristol, Avon) 1995;2:85–92. doi: 10.1016/0268-0033(95)92044-m. [DOI] [PubMed] [Google Scholar]
- Bennett MB, Ker RF, Dimery NJ, Alexander RM. Mechanical properties of various mammalian tendons. J Zool. 1986;209:537–548. [Google Scholar]
- Brzycki M. Strength testing - predicting a one-rep max from reps to fatigue. J Phys Educ Recreation Dance. 1993;64:88–90. [Google Scholar]
- Buchanan CI, Marsh RL. Effects of long-term exercise on the biomechanical properties of the achilles tendon of guinea fowl. J Appl Physiol. 2001;90:164–171. doi: 10.1152/jappl.2001.90.1.164. [DOI] [PubMed] [Google Scholar]
- Butler DL, Grood ES, Noyes FR, Zernicke RF. Biomechanics of ligaments and tendons. Exerc Sport Sci Rev. 1978;6:125–181. [PubMed] [Google Scholar]
- Cohen RE, Hooley CJ, McCrum NG. Viscoelastic creep of collagenous tissue. J Biomech. 1976;9:175–184. doi: 10.1016/0021-9290(76)90002-6. [DOI] [PubMed] [Google Scholar]
- Cronkite AE. Tensile strength of human tendons. Anat Rec. 1936;64:173–186. [Google Scholar]
- Diamant J, Keller A, Baer E, Litt M, Arridge RGC. Collagen; ultrastructure and its relation to mechanical properties as a function of ageing. Proc R Soc Lond B Biol Sci. 1972;180:293–315. doi: 10.1098/rspb.1972.0019. [DOI] [PubMed] [Google Scholar]
- Dunn MG, Silver FH. Viscoelastic behaviour of human connective tissues: relative contribution of viscous and elastic components. Connect Tissue Res. 1983;12:59–70. doi: 10.3109/03008208309005612. [DOI] [PubMed] [Google Scholar]
- Elliott DH. Structure and function of mammalian tendon. Biol Rev. 1965;40:392–421. doi: 10.1111/j.1469-185x.1965.tb00808.x. [DOI] [PubMed] [Google Scholar]
- Feder G, Cryer C, Donovan S, Carter Y. Guidelines for the prevention of falls in people over 65. BMJ. 2000;321:1007–1011. doi: 10.1136/bmj.321.7267.1007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frontera WR, Hughes VA, Fielding RA, Fiatarone MA, Evans WJ, Roubenoff R. Aging of skeletal muscle: a 12-yr longitudinal study. J Appl Physiol. 2000;88:1321–1326. doi: 10.1152/jappl.2000.88.4.1321. [DOI] [PubMed] [Google Scholar]
- Haut RC, Powlison AC. The effects of test environment and cyclic stretching on the failure properties of human patellar tendons. J Orthop Res. 1990;8:532–540. doi: 10.1002/jor.1100080409. [DOI] [PubMed] [Google Scholar]
- Haut TL, Haut RC. The state of tissue hydration determines the strain-rate-sensitive stiffness of human patellar tendon. J Biomech. 1997;30:79–81. doi: 10.1016/s0021-9290(96)00108-x. [DOI] [PubMed] [Google Scholar]
- Heikkinen E, Vuori I. Effect of physical activity on the metabolism of collagen in aged mice. Acta Physiol Scand. 1972;84:543–549. doi: 10.1111/j.1748-1716.1972.tb05206.x. [DOI] [PubMed] [Google Scholar]
- Hooley CJ, McCrum NG, Cohen RE. The viscoelastic deformation of tendon. J Biomech. 1980;13:521–528. doi: 10.1016/0021-9290(80)90345-0. [DOI] [PubMed] [Google Scholar]
- Hopkins WG. Measures of reliability in sports medicine and science. Sports Med. 2000;30:1–15. doi: 10.2165/00007256-200030010-00001. [DOI] [PubMed] [Google Scholar]
- Ito M, Kawakami Y, Ichinose Y, Fukashiro S, Fukunaga T. Nonisometric behaviour of fascicles during isometric contractions of a human muscle. J Appl Physiol. 1998;85:1230–1235. doi: 10.1152/jappl.1998.85.4.1230. [DOI] [PubMed] [Google Scholar]
- Jewell BR, Wilkie DR. An analysis of the mechanical components in frog striated muscle. J Physiol. 1958;143:515–540. doi: 10.1113/jphysiol.1958.sp006075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kannus P, Józsa L. Histopathological changes preceding spontaneous rupture of tendon. A controlled study of 891 patients. J Bone Joint Surg. 1991;73:1507–1525. [PubMed] [Google Scholar]
- Kellis E, Baltzopoulos V. In vivo determination of the patella tendon and hamstrings moment arms in adult males using videofluoroscopy during submaximal knee extension and flexion. Clin Biomech (Bristol, Avon) 1999;14:118–124. doi: 10.1016/s0268-0033(98)00055-2. [DOI] [PubMed] [Google Scholar]
- Kelly DW, Carter VS, Jobe FV, Kerlan RK. Patellar and quadriceps tendon ruptures - jumper's knee. Am J Sports Med. 1984;12:375–380. doi: 10.1177/036354658401200508. [DOI] [PubMed] [Google Scholar]
- Kent-Braun JA, Ng AV. Specific strength and voluntary muscle activation in young and elderly women and men. J Appl Physiol. 1999;87:22–29. doi: 10.1152/jappl.1999.87.1.22. [DOI] [PubMed] [Google Scholar]
- Kent-Braun JA, Ng AV, Doyle JW, Towse TF. Human skeletal muscle responses vary with age and gender during fatigue due to incremental isometric exercise. J Appl Physiol. 2002;93:1813–1823. doi: 10.1152/japplphysiol.00091.2002. [DOI] [PubMed] [Google Scholar]
- Ker RF. Dynamic tensile properties of the plantaris tendon of sheep (Ovis aries) J Exp Biol. 1981;93:283–302. doi: 10.1242/jeb.93.1.283. [DOI] [PubMed] [Google Scholar]
- Ker RF, Alexander RM, Bennett MB. Why are mammalian tendons so thick? J Zool. 1988;216:309–324. [Google Scholar]
- Klein CS, Rice CL, Marsh GD. Normalised force, activation and coactivation in the arm muscles of young and old men. J Appl Physiol. 2001;91:1341–1349. doi: 10.1152/jappl.2001.91.3.1341. [DOI] [PubMed] [Google Scholar]
- Kubo K, Kanehisa H, Fukunaga T. Effects of resistance and stretching training programmes on the viscoelastic properties of human tendon structures in vivo. J Physiol. 2002;538:219–226. doi: 10.1113/jphysiol.2001.012703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kubo K, Kanehisa H, Ito M, Fukunaga T. Effects of isometric training on the elasticity of human tendon structures in vivo. J Appl Physiol. 2001a;91:26–32. doi: 10.1152/jappl.2001.91.1.26. [DOI] [PubMed] [Google Scholar]
- Kubo K, Kanehisa H, Kawakami Y, Fukunaga T. Influence of static stretching on viscoelastic properties of human tendon structures in vivo. J Appl Physiol. 2001b;90:520–527. doi: 10.1152/jappl.2001.90.2.520. [DOI] [PubMed] [Google Scholar]
- Lieber RL, Leonard ME, Brown CG, Trestik CL. Frog semitendinosis tendon load-strain and stress-strain properties during passive loading. Am J Physiol. 1991;261:C86–92. doi: 10.1152/ajpcell.1991.261.1.C86. [DOI] [PubMed] [Google Scholar]
- Lippold OC. The relationship between integrated action potentials in a human muscle and its isometric tension. J Physiol. 1952;177:492–499. doi: 10.1113/jphysiol.1952.sp004763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Londeree BR, Moeschberger ML. Effect of age and other factors on heart rate. Res Q Exerc Sport. 1982;53:297–304. [Google Scholar]
- McMaster PE. Tendon and muscle ruptures. J Bone Joint Surg. 1933;15:705–722. [Google Scholar]
- Maganaris CN. Proceedings of the Active Life Span Research Symposium, The Plasticity of the Motor System: Adaptations to Increased Use, Disuse and Ageing. UK: Manchester Metropolitan University; 2001. In vivo tendon mechanical properties in young adults and healthy elderly. Available from the website address http://www.mmu.ac.uk/c-a/exspsci/alsymp.html. [Google Scholar]
- Maganaris CN, Baltzopoulos V, Sargeant AJ. In vivo measurements of the triceps surae complex architecture in man: implications for muscle function. J Physiol. 1998;512:603–614. doi: 10.1111/j.1469-7793.1998.603be.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maganaris CN, Paul JP. In vivo human tendon mechanical properties. J Physiol. 1999;521:307–313. doi: 10.1111/j.1469-7793.1999.00307.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maganaris CN, Paul JP. Load-elongation characteristics of in vivo human tendon and aponeurosis. J Exp Biol. 2000a;203:751–756. doi: 10.1242/jeb.203.4.751. [DOI] [PubMed] [Google Scholar]
- Maganaris CN, Paul JP. In vivo human tendinous tissue stretch upon maximum muscle force generation. J Biomech. 2000b;33:1453–1459. doi: 10.1016/s0021-9290(00)00099-3. [DOI] [PubMed] [Google Scholar]
- Magnusson SP, Aagaard P, Rosager S, Dyhre-Poulsen P, Kjaer M. Load-displacement properties of the human triceps surae aponeurosis in vivo. J Physiol. 2001;531:277–288. doi: 10.1111/j.1469-7793.2001.0277j.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michna H, Hartmann G. Adaptation of tendon collagen to exercise. Int Orthop. 1989;13:161–165. doi: 10.1007/BF00268040. [DOI] [PubMed] [Google Scholar]
- Muramatsu T, Muraoka T, Takeshita D, Kawakami Y, Hirano Y, Fukunaga T. Mechanical properties of tendon and aponeurosis of human gastrocnemius muscle in vivo. J Appl Physiol. 2001;90:1671–1678. doi: 10.1152/jappl.2001.90.5.1671. [DOI] [PubMed] [Google Scholar]
- Nachemson AL, Evans JH. Some mechanical properties of the third human lumbar interlaminar ligament (ligamentum flavum) J Biomech. 1968;1:211–220. doi: 10.1016/0021-9290(68)90006-7. [DOI] [PubMed] [Google Scholar]
- Nagamine R, Otani T, White SE, McCarthy DS, Whiteside LA. Patellar tracking measurements in the normal knee. J Orthop Res. 1995;13:115–122. doi: 10.1002/jor.1100130117. [DOI] [PubMed] [Google Scholar]
- Narici MV, Binzoni T, Hiltbrand E, Fasel J, Terrier F, Cerretelli P. In vivo human gastrocnemius architecture with changing joint angle at rest and during graded isometric contraction. J Physiol. 1996;496:287–297. doi: 10.1113/jphysiol.1996.sp021685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Narici MV, Ciuffreda L, Baldi M, Capodaglio P. Unique features of work-induced skeletal muscle hypertrophy in elderly humans. J Physiol. 2000;526.P:35P. [Google Scholar]
- Noyes FR, Grood ES. The strength of the anterior cruciate ligament in humans and rhesus monkeys. Age-related and species-related changes. J Bone Joint Surg. 1976;58:1074–1082. [PubMed] [Google Scholar]
- Rigby BJ. Effect of cyclic extension on the physical properties of tendon collagen and its possible relation to biological ageing of collagen. Nature. 1964;202:1072–1074. doi: 10.1038/2021072a0. [DOI] [PubMed] [Google Scholar]
- Rollhäuser H. Funktionelle anpassung der sehnenfaser im submikroskopischen bereich. Anat Anz Ergänzungsheft. 1954;51:318–322. [Google Scholar]
- Sheehan FT, Zajac FE, Drace JE. In vivo tracking of the human patella using cine phase contrast magnetic resonance imaging. J Biomech Eng. 1999;121:650–656. doi: 10.1115/1.2800868. [DOI] [PubMed] [Google Scholar]
- Smidt GL. Biomechanical analysis of knee flexion and extension. J Biomech. 1973;6:79–92. doi: 10.1016/0021-9290(73)90040-7. [DOI] [PubMed] [Google Scholar]
- Tkaczuk H. Tensile properties of human lumbar longitudinal ligaments. Acta Orthop Scand. 1968;S115:1. doi: 10.3109/ort.1968.39.suppl-115.01. [DOI] [PubMed] [Google Scholar]
- Verzár F. The ageing of connective tissue. Gerontologia. 1957;1:363–378. [PubMed] [Google Scholar]
- Viidik A. The effect of training on the tensile strength of isolated rabbit tendons. Scand J Plast Reconstr Surg. 1967;1:141–147. doi: 10.3109/02844316709022844. [DOI] [PubMed] [Google Scholar]
- Viidik A. Tensile strength properties of achilles tendon system in trained and untrained rabbits. Acta Orthop Scand. 1969;40:261–272. doi: 10.3109/17453676908989506. [DOI] [PubMed] [Google Scholar]
- Vogel HG. Species differences of elastic and collagen tissue - influence of maturation and age. Mech Ageing Dev. 1991;57:15–24. doi: 10.1016/0047-6374(91)90021-q. [DOI] [PubMed] [Google Scholar]
- Walker LB, Harris EH, Benedict JV. Stress-strain relationship in human cadaveric plantaris tendon: a preliminary study. Med Electron Biol Eng. 1964;2:31–38. doi: 10.1007/BF02474358. [DOI] [PubMed] [Google Scholar]
- Woo SL-Y, Gomez MA, Woo Y-K, Akeson WH. Mechanical properties of tendons and ligaments. Biorheology. 1982;19:397–408. doi: 10.3233/bir-1982-19302. [DOI] [PubMed] [Google Scholar]
- Woo SL-Y, Ritter MA, Amiel D, Sanders TM, Gomez MA, Kuei SC, Garfin SR, Akeson WH. The biomechanical and biochemical properties of swine tendons - long term exercise effects of exercise on the digital extensors. Connect Tissue Res. 1980;7:177–183. doi: 10.3109/03008208009152109. [DOI] [PubMed] [Google Scholar]
- Wood TO, Cooke PH, Goodship AE. The effect of exercise and anabolic steroids on the mechanical properties and crimp morphology of the rat tendon. Am J Sports Med. 1988;16:153–158. doi: 10.1177/036354658801600211. [DOI] [PubMed] [Google Scholar]
- Wren TAL, Yerdy SA, Beaupré GS, Carter DR. Mechanical properties of the human achilles tendon. Clin Biomech (Bristol, Avon) 2001;16:245–251. doi: 10.1016/s0268-0033(00)00089-9. [DOI] [PubMed] [Google Scholar]
- Zipp P. Recommendations for the standardization of lead positions in surface electromyography. Eur J Appl Physiol. 1982;50:41–54. [Google Scholar]
- Zuurbier CJ, Everard AJ, Van Der Wees P, Huijing PA. Length-force characteristics of the aponeurosis in the passive and active muscle condition and in the isolated condition. J Biomech. 1994;27:445–453. doi: 10.1016/0021-9290(94)90020-5. [DOI] [PubMed] [Google Scholar]