

Abstract
Angiogenesis is a prerequisite for tumour progression and is highly regulated by growth factors and cytokines a number of which also stimulate the production of nitric oxide. Asymmetric dimethylarginine is an endogenous inhibitor of nitric oxide synthesis. Asymmetric dimethylarginine is metabolised by dimethylarginine dimethylaminohydrolase. To study the effect of dimethylarginine dimethylaminohydrolase on tumour growth and vascular development, the rat C6 glioma cell line was manipulated to overexpress the rat gene for dimethylarginine dimethylaminohydrolase I. Enhanced expression of dimethylarginine dimethylaminohydrolase I increased nitric oxide synthesis (as indicated by a two-fold increase in the production of cGMP), expression and secretion of vascular endothelial cell growth factor, and induced angiogenesis in vitro. Tumours derived from these cells grew more rapidly in vivo than cells with normal dimethylarginine dimethylaminohydrolase I expression. Immunohistochemical and magnetic resonance imaging measurements were consistent with increased tumour vascular development. Furthermore, dimethylarginine dimethylaminohydrolase activity was detected in a series of human tumours. This data demonstrates that dimethylarginine dimethylaminohydrolase plays a pivotal role in tumour growth and the development of the tumour vasculature by regulating the concentration of nitric oxide and altering vascular endothelial cell growth factor production.
British Journal of Cancer (2002) 87, 673–680. doi:10.1038/sj.bjc.6600518 www.bjcancer.com
© 2002 Cancer Research UK
Keywords: dimethylarginine dimethylaminohydrolase, nitric oxide, nitric oxide synthase, vascular endothelial growth factor, asymmetric dimethylarginine
Angiogenesis is the formation of new capillaries from existing blood vessels. It is a multistep process that involves dissociation of endothelial cells (EC) from adjacent pericytes, remodelling of the extracellular matrix, proliferation and migration of EC and capillary differentiation. Insight into the regulation of this process is central to our understanding of a number of biological processes whether physiological, as occurs during development, the reproductive cycle and wound healing, or pathological, as occurs during atherogenesis, diabetic retinopathy and tumour progression (Carmeliet and Jain, 2000; Folkman, 1995).
There is strong evidence to suggest that nitric oxide (NO), produced from arginine by the nitric oxide synthases (NOS), is a crucial signalling molecule and regulator of angiogenesis (Chinje and Stratford, 1997). NO enhances vascular permeability, induces extracellular matrix degradation, endothelial cell proliferation and migration (Ziche et al, 1994; Trachtman et al, 1996; Fukumura et al, 1997) and stimulates the expression of vascular endothelial growth factor (VEGF) while also mediating many of its angiogenic effects (Chin et al, 1997; Papapetropoulos et al, 1997). The role of NO in tumour progression is still unclear, since NO has been shown to inhibit angiogenesis (Pipili-Synetos et al, 1995) and NO produced by activated macrophages can be cytotoxic to tumour cells (Stuehr and Nathan, 1989). However, NO has been positively correlated with tumour grade in human gynaecological, breast, neuronal, and head and neck tumours (Thomsen et al, 1994; Cobbs et al, 1995; Gallo et al, 1998; Reveneau et al, 1999).
Regulation of NO production may therefore play an important role in the regulation of angiogenesis and consequently tumour progression. Methylated analogues of arginine such as asymmetric dimethylarginine (ADMA) and N-mono methylarginine (L-NMMA) are competitive inhibitors of NO synthesis. Free ADMA is found in plasma and urine of healthy individuals and is synthesised by the post-translational methylation of protein arginine residues and liberated upon their hydrolysis (Kakimoto and Akazawa, 1970; Fickling et al, 1993). The intracellular concentration of ADMA reaches levels sufficient to inhibit NO synthesis (Macallister et al, 1994) and can be modulated by changes in the activity of the enzyme dimethylarginine dimethylaminohydrolase (DDAH) (Ogawa et al, 1989; Macallister et al, 1996).
Two isoforms of DDAH have been reported (Leiper et al, 1999). DDAH I predominates in tissues that express the neuronal isoform of NOS, while DDAH II distribution mainly follows endothelial NOS (Leiper et al, 1999). Dysfunction of DDAH and elevated levels of its substrate ADMA have been implicated in pathological conditions including hypertension, pre-eclampsia, renal failure and atherosclerosis (Matsuoka et al, 1997; Holden et al, 1998; Miyazaki et al, 1999). Recently, high levels of ADMA have been correlated with impaired angiogenesis in hypercholesterolemic mice (Jang et al, 2000). The aim of our study was to test the hypothesis that tumour-derived ADMA plays a significant role in the regulation of tumour growth and vascular development. We demonstrate that increased DDAH expression results in decreased tumour ADMA, increased NO production, tumour growth and angiogenesis, and that these effects are mediated through changes in the expression of VEGF. We also show preliminary evidence of DDAH activity in human tumours.
METHODS
Cell culture and transfections
Rat C6 glioma cells (European Collection of Cell Cultures, Centre for Applied Microbiology and Research, UK) were maintained in Ham's F-10 medium containing 5 mM L-glutamine, 100 U ml−1 penicillin, 0.1 mg ml−1 streptomycin and 10% (v v−1) foetal calf serum. The full length rat DDAH I cDNA was generated by RT–PCR using mRNA isolated from C6 cells, forward primer: GGAGCAAGCTTCGCCACCATGGCCGGCCTCAGCCA and reverse primer: GCCTGCTCGAGTCAAGAGTCTGTCTTCTTGTTAAT and subsequently cloned into HindIII/XhoI sites in pcDNA 3.1/hygro (InVitrogen). C6 cells were transfected using the poly L-ornithine method (5×106 cells per 9 cm plate; 10 μg DNA plasmid; 10 μg ml−1 poly L-ornithine) and selected in medium supplemented with 500 μg ml−1 hygromycin B (Dong et al, 1994).
Animals and tumours
Female (7–8 weeks old) MF1/nu mice were injected subcutaneously in the flanks with 2×106 cells in 0.1 ml PBS. Tumour growth was measured at daily intervals after anaesthetising with fluothane in 2% N2O, 5% O2. The tumour volume was calculated using the formula: tumour volume=π/6·(length×width×depth). All procedures were performed in accordance to the Home Office Scientific Procedures Act 1986.
Western blot analysis
The protein concentration of cell lysates and tumour homogenates were determined by Bradford assay and equal amounts analysed by SDS–PAGE. DDAH expression was detected using a monoclonal antibody against rat DDAH I (Kimoto et al, 1995).
DDAH activity assay
Cells were seeded on 24-well plates at a density of 1.5×105 cells per well and incubated in HEPES buffered Krebs solution containing [14C] L-NMMA (0.8 μCi ml−1, specific activity 56 mCi mMol−1) for 1 h at 37°C. Cells were washed twice in ice-cold Krebs solution and lysed in 0.1% (w v−1) SDS. Human tumour biopsy specimens taken during surgical removal of the tumour, or mouse tumour tissues were homogenised in 10 mM sodium phosphate buffer pH 6.6 using a motor homogeniser. Aliquots of 500 μl were incubated with [14C] L-NMMA (0.4 μCi ml−1) for 1 h at 37°C. The formation of [14C] citrulline was determined as previously described (Macallister et al, 1996). The enzymatic activity was corrected for the protein concentration.
HPLC analysis
Assessment of the concentration of ADMA in tumour homogenates and in culture medium was performed as previously described (Macallister et al, 1994).
cGMP assay
Estimation of the production of cGMP in tumour cells was performed as previously described (Whitley, 1994).
In vitro assay of endothelial cell migration
A human endothelial cell line SGHEC-7 derived from SV40 transfected human umbilical vein endothelial cells (Fickling et al, 1992) was cultivated on microcarrier (MC) beads for 2 days and embedded in fibrin gels (Nehls and Drenckhahn, 1995). Conditioned medium from confluent cultures mixed 1 : 1 with fresh medium was added. After 2 days incubation, digital images were acquired and the extent of migration was determined using Image Pro-Plus software (Media Cybernetics). Invasion was determined as the maximum length of invasive processes minus the radius of the bead.
Northern blot analysis
Total RNA was prepared from cells using TRIZOL (Gibco BRL). The probe for VEGF was a 540 bp BamHI-HindIII fragment of the human VEGF121 cDNA (nt 565–1012). The high identity in the cDNA sequences of human and rat or mouse VEGF ensures the crossreactivity of the probe (Shima et al, 1996). The probe for the 18S subunit corresponds to nucleotides 552–1429 nt of the human 18S gene.
VEGF ELISA
Culture medium was collected from confluent 6-well plates and analysed for VEGF expression using a murine VEGF ELISA kit (R&D Systems, detection limit 3 pg ml−1). Tumours were excised at an early stage of development (day 15), weighed, cut in small pieces and maintained in culture medium for 4 days. VEGF secretion was measured in the culture medium. Values were correlated with the protein concentration of cells or the weight of tumours, respectively.
Magnetic resonance imaging (MRI)
A separate cohort of size-matched D27 and C6 wild type tumours were imaged at 18 and 23 days post-passage respectively. 1H MR imaging was performed on a Varian spectrometer at 4.7 T using a two-turn surface coil (1 cm diameter). Mice bearing tumours derived from D27 (mean volume 1.23±0.2 cm3, n=8) or C6 (mean volume 1.25±0.2 cm3, n=8) wild type cells were anaesthetised with a single intraperitonal dose of 10 ml kg−1 Hypnorm/Hypnovel and placed so the tumour hung into the centre of the surface coil. The mouse core temperature was maintained at 37°C by a warm water blanket. Multi-gradient echo (MGRE) images were acquired from three transverse 1 mm thick slices through the centre of the tumour. The readout gradient was oscillated to generate a series of eight echoes, utilising a repetition time of 100 ms, initial echo of 5 ms and an echo spacing of 5 ms, resulting in sets of images with decreased R2* weighting. Tumour R2* maps for each slice were generated using all eight gradient-echo image sets by fitting an exponential model on a pixel-by-pixel basis (Robinson et al, 2001; Griffiths et al, 2002). For each slice, R2* was determined from a region-of-interest (ROI) encompassing the whole tumour but excluding the surrounding skin/muscle.
Fluorescence microscopy
Mice from the same cohort as used for the MRI and bearing similarly size-matched tumours were injected intravenously with 15 mg kg−1 of the perfusion marker Hoechst 33342 (Sigma) 1 min prior to cervical dislocation. Tumours were rapidly excised, frozen and sections cut (10 μm). Acetone fixed sections were visualised by fluorescence microscopy. Fluorescence signals from entire tumour sections were recorded with digital images acquired under identical conditions. The area of tumour stained by Hoechst 33342 as a percentage of the total area of tumour section was calculated using anaLYSIS (Soft Image Analysis).
Ethics
Local Ethical Committee approval was obtained for the collection of human tissues.
RESULTS
Overexpression DDAH in C6 glioma cells
To study the role of DDAH in tumour biology, the rat C6 glioma cell line was transfected with the pcDNA vector, containing the coding region of the rat DDAH I gene. Three stable clones, (D20, D26, D27) that constitutively express DDAH I and mock transfected clones (M lines, transfected with the empty vector as control) were analysed. Western blot analysis of D20, D26 and D27 cells revealed similarly high levels of DDAH I protein expression (Figure 1A). Consistent with the high protein expression these cell lines displayed, on average, a four-fold increase in enzymatic activity compared to wild type C6 and control transfected M4 cells, as determined by the conversion of 14C radiolabelled L-NMMA to L-citrulline (Figure 1B). Since the function of DDAH is to catabolise the endogenous inhibitors of NO, ADMA and L-NMMA, the levels of ADMA in the culture medium of the overexpressing cell lines was examined. Overexpression of DDAH I was shown to reduce the levels of ADMA by 70–75% (Figure 1C). Consistent with the role of ADMA as an inhibitor of NO, the decreased levels of ADMA led to an increase in NO production as indicated by a two-fold increase in the production of cGMP (C6: 12.33±1.4, D27: 26.67±1.5, mean±s.e.mean). In vitro studies showed no apparent correlation between transfection with DDAH (compared to empty vector) and growth (Figure 1D). However transfected cells generally showed slower growth rates than C6 wild type cells. All three DDAH I overexpressing cell lines grew similarly to M4 and M6 (Figure 1D) and other mock transfected lines (data not shown).
Figure 1.
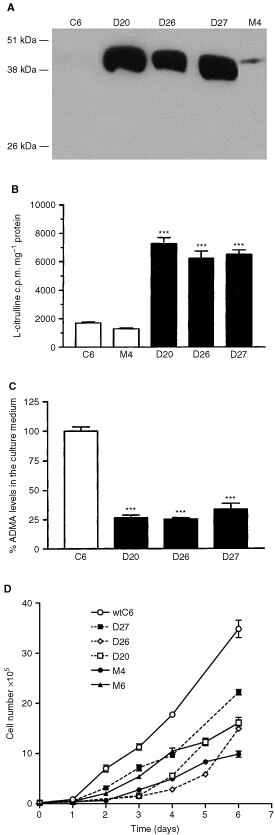
Characterisation of the stable DDAH I transfected cell lines in vitro. (A) Western blot analysis of DDAH I overexpressing cell lines (20 μg protein per lane). An increase in a band at approximately 38 kDa, indicating the presence of DDAH I was observed. (B) Assessment of DDAH activity in cell lysates by the conversion of [14C] L-NMMA to L-citrulline revealed that the D20, D26 and D27 cell lines had a four-fold increase in DDAH enzymatic activity. Results are means+s.e.m. of triplicates of three separate experiments (***P<0.001 compared to control, Student's t-test). (C) Overexpression of DDAH I reduced the concentration of ADMA by 70–75% in the culture medium as determined by HPLC analysis. Results are mean %+s.e.m. of triplicate determinations from three separate experiments (***P<0.001, Student's t-test). (D) The in vitro growth rate of DDAH transfected cells (dashed lines) D20, D26, D27, mock transfected cells (solid lines) M4, M6 and wild type C6 cells is shown. Results are means±s.e.m. of three experiments performed in at least duplicate.
Effect of DDAH I overexpression on tumour growth in vivo
In contrast, when D26 and D27 cells were implanted into the flanks of nude mice their growth rate increased (doubling time: ∼2 days) compared to the wild type or the control M4 cells (doubling time: ∼4 days, Figure 2A). In a limited number of experiments the growth of C6 tumours was allowed to continue until day 26 when they reached a similar size to that of D27 at 20 days (data not shown). Western blot analysis of tumours demonstrated higher DDAH I protein expression in D27 compared to C6 tumours (Figure 2B). Homogenates from D26 and D27 tumours displayed a 2.5-fold increase in DDAH activity (Figure 2C). The concentration of ADMA also decreased from 2.6 μg mg−1 protein in C6 tumours to undetectable levels in D27 tumours.
Figure 2.
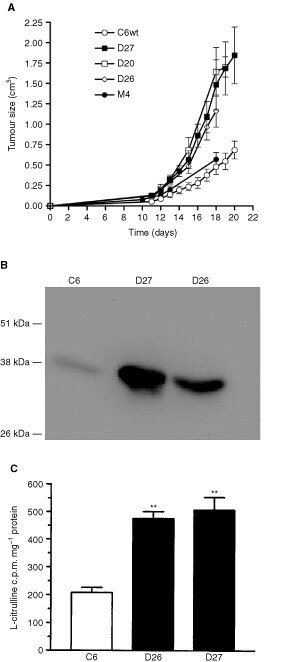
Overexpression of DDAH I increases the in vivo tumour growth rate. (A) In vivo, overexpression of DDAH I caused an increase in the growth rate of D20, D26 and D27 tumours compared to M4 and C6 tumours. Results are means±s.e.m. of at least two different inoculations of six mice. (B) Western blot analysis of tumours (100 μg protein per lane) confirmed that the overexpressing DDAH tumours had higher amounts of DDAHI protein in vivo. (C) Assessment of DDAH activity in tumour homogenates by the conversion of [14C] L-NMMA to L-citrulline. Tumour homogenates derived from D26 and D27 revealed a two- and a half-fold increase in their DDAH activity compared to C6 tumours. Results are means+s.e.m. of duplicates from two separate experiments (**P<0.02 compared to control, Student's t-test).
Effect of DDAH I on tumour vascular development
Non-invasive Magnetic Resonance Imaging (MRI) was employed to assess the blood vessel development in vivo. Deoxyhaemoglobin is paramagnetic and creates magnetic susceptibility perturbations around blood vessels, increasing the MRI transverse relaxation time R2*. Multi-gradient recalled echo (MGRE) MR images allow the relaxation rate R2* to be quantified, which is directly proportional to the tissue content of deoxyhaemoglobin and hence a sensitive index of tissue vascularity (Robinson et al, 2001; Griffiths et al, 2002). The average R2* of D27 and C6 tumours was 65.6±5 s−1 and 42.5±4 s−1 respectively. The significantly faster R2* displayed by the D27 tumours compared to the wild type (Student's t-test, P<0.001) indicates a higher concentration of deoxyhaemoglobin and increased tumour vascularisation (Figure 3A). In addition to the higher growth rate, tumours derived from the D27 cells were shown upon excision to bleed profusely and to be more vascularised compared to wild type tumours (Figure 3B).
Figure 3.
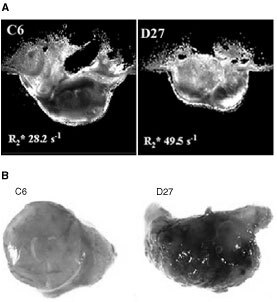
Overexpression of DDAH I affects the tumour vasculature. (A) Representative R2* maps obtained from size-matched C6 and D27 tumours, and their associated average R2* relaxation rates, imaged 23 days and 18 days post-passage respectively. The more intense R2* map (brighter image) of the D27 tumour is consistent with a high concentration of deoxyhaemoglobin and hence better vessel development. (B) Representative photographs of size-matched tumours derived from C6 and D27 cells. C6 and D27 tumours were excised 26 and 20 days post-implantation respectively. DDAH I overexpressing tumours are better vascularised compared to wild type or control tumours.
DDAH I expression results in increased tumour perfusion
The effect of DDAH I on tumour perfusion was examined using the fluorescent marker Hoechst 33342 (Smith et al, 1988). This is a nuclear dye that, upon injection, stains endothelial cells and cells immediately adjacent to tumour blood vessels. Since the time of experiment was limited to 1 min to avoid diffusion of the dye, Hoechst 33342 marked only the functionally perfused vessels. Fluorescence microscopy showed that the extent of the staining is greater in D27 tumours compared to wild type tumours indicating better tumour perfusion and increased number of functional vessels (C6: 5.1%±0.5, D27: 9%±0.5, mean±s.e.m, n=4).
DDAH I induces in vitro vascular endothelial cell migration
The role of DDAH I on vessel development was examined further using an in vitro assay of endothelial migration. Human endothelial cells, SGHEC-7, grown on microcarrier (MC) beads and embedded in fibrin gels were incubated with conditioned medium from confluent cultures of C6, D27 or SGHEC-7 cells and the formation of invasive processes was analysed over a period of 2 days. Invasion of endothelial cells into the fibrin gel was used as an indicator of the angiogenic potential of factors produced by the D27 cells. The average of the maximum length of the process minus the radius of the bead from 40 MC beads was determined. Statistical analysis showed that conditioned medium from D27 cells significantly stimulated the migration of SGHEC-7 cells into the fibrin gel compared to medium from C6 or SGHEC-7 cells (Figure 4).
Figure 4.
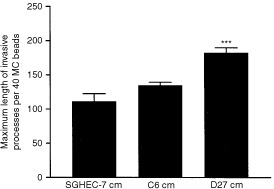
Conditioned medium from DDAH I overexpressing cells induces endothelial cell migration in vitro. A human endothelial cell line SGHEC-7 was grown on beads in three dimensional fibrin gels and incubated with conditioned medium (cm) from confluent SGHEC-7, D27 or C6 cells. Quantitative analysis of invasive process formation was expressed as the average of the maximum length of process minus the radius of the bead per 40 MC beads. Results are means+s.e.m. of 40 individual measurements in three separate experiments (***P<0.001 compared to control, Student's t-test).
DDAH I expression increases the production of VEGF
Since it has previously been suggested that NO can mediate some of the angiogenic effects of VEGF, we examined whether the effects of DDAH and ADMA on tumour angiogenesis were mediated through changes in VEGF. In vitro analysis of VEGF mRNA displayed a two-fold increase in D20, D26, D27 compared to control M4 and wild type C6 cells as determined by Northern blot analysis (Figure 5A). This correlated with an average two-fold increase in VEGF secretion in the culture medium as determined by ELISA (Figure 5B). Ex vivo analysis of VEGF secretion displayed a two-fold increase in D27 compared to C6 tumour culture supernatant (Figure 5C).
Figure 5.
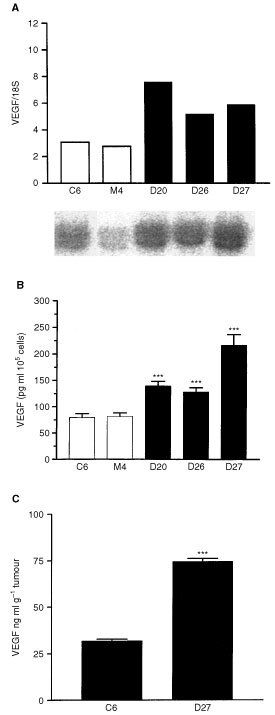
Overexpression of DDAH I results in upregulation of VEGF. (A) VEGF mRNA expression in the D27, D26, D20, M4 and parent C6 cell lines assessed by Northern blot analysis. RNA loading was corrected according to the levels of the 18S ribosomal subunit. The signal intensity was quantified by densitometry and expressed as the ratio of VEGF/18S. The lower panel shows a representative Northern blot analysis of VEGF expression in cell lysates. (B) VEGF expression in the culture medium of D27, D26, D20, M4 and parent C6 cell lines assessed by ELISA. Results are means+s.e.m. of duplicates of three individual experiments (***P<0.001 compared to control, Student's t-test). (C) VEGF secretion in the culture medium of D27 and C6 derived tumours were assessed by ELISA. Results are means+s.e.m. of triplicates of three tumours respectively (***P<0.001, Student's t-test).
DDAH I activity in human tumours
DDAH activity was detected in eight human tumours, including those from the brain (Figure 6).
Figure 6.
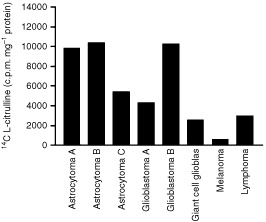
Presence of DDAH activity in human tumours. DDAH activity in tumour homogenates derived from eight individual human tumour biopsy specimens taken during surgical removal of the tumour. DDAH activity was assessed by the conversion of [14C] L-NMMA to L-citrulline. DDAH activity varied in the different tumour types.
DISCUSSION
Tumour angiogenesis is driven by the requirement of the tumour for nutrients and oxygen. Angiogenesis involves the coordination of a number of independent events including the production of proteases and angiogenic factors that have both a chemotactic and mitogenic effect on endothelial cells. We demonstrate a novel mechanism by which a reduction in tumour ADMA, as a result of increased DDAH activity, stimulates tumour growth and angiogenesis through increased NO and VEGF expression.
Regulation of the production of NO through DDAH activity is important since NO can have pleiotropic effects on diverse aspects of tumour biology. There is positive correlation between NOS activity and tumour grade in human breast, gynaecological, head, neck and neuronal tumours (Thomsen et al, 1994; Cobbs et al, 1995; Gallo et al, 1998; Reveneau et al, 1999) and NOS inhibitors have been shown to decrease tumour growth (Thomsen et al, 1997). However, the role of tumour-derived NO is still unclear since overexpression of iNOS in tumour cells can promote or inhibit tumour growth and metastasis (Dong et al, 1994; Jenkins et al, 1995; Xie et al, 1995).
Increased malignancy of human primary brain tumours and the formation of metastasis is associated with increased neovascularisation. Astrocytomas represent one of the most vascularised human neoplasms and the progression from low to high grade is characterised by an increase in neovascularisation, focal necrosis and cellular proliferation. Tumours derived from the rat C6 glioma cell line are histologically classified as astrocytomas and represent a well-established model of human glioblastomas (Barth, 1998). C6 glioma cells express both eNOS and iNOS (Simmons and Murphy, 1993; Barna et al, 1996).
The result of increased DDAH I activity in C6 cells and the concomitant fall in the concentration of ADMA led to a two-fold increase in NO synthesis as observed by increased cGMP production. However this did not alter the in vitro growth properties of the glioma cell lines. The in vivo growth rate of D20, D26 and D27 cells was significantly increased compared to the wild type or the control M4 cells. Our data suggest that DDAH/ADMA pathway does not have a direct effect on the cell cycle or cell proliferation but indirectly affects the tumour growth, possibly by interfering with the tumour vessel development.
To establish a role for ADMA and DDAH in tumour angiogenesis in vivo, MGRE-MRI was used as a non-invasive assay of blood vessel development of D27 and wild type C6 tumours, followed by gross histology and fluorescence microscopy using the nuclear dye Hoechst 33342. It is clear from these experiments that DDAH I expression leads to higher blood volume, better tumour perfusion and increased number of functional vessels. We can therefore speculate that the increased growth observed in D27 tumours is dependent on increased blood vessel development.
Sprouting of endothelial cells from pre-existing vessels, the invasion into the surrounding tissue and the reorganisation into new capillary structures are important steps in vessel development. DDAH I expression and ADMA concentration does not affect the proliferation rate of endothelial cells (V Kostourou, unpublished observations). Therefore, a possible role of DDAH I in vessel development could be the facilitation of the endothelial cell invasion. In support of this, conditioned medium from D27 cells significantly stimulated the migration of SGHEC-7 cells into the fibrin gel compared to medium from C6 or SGHEC-7 cells. These results demonstrate that DDAH activity and the intracellular concentration of ADMA regulate the release of an angiogenic factor or factors in vitro that affect the invasive properties of the endothelial cells.
Vessel growth and development is under the control of a number of factors the most potent being VEGF. VEGF is a largely endothelial cell-specific mitogen that is induced by hypoxia. It promotes angiogenesis, blood vessel permeability and maintains tumour vessel integrity (Shweiki et al, 1992; Yancopoulos et al, 2000). Increased VEGF expression is related to neovascularisation and tumour progression (Takahashi et al, 1995; Zhang et al, 1995) while inhibition of VEGF or its receptors inhibits tumour growth in vivo (Kim et al, 1993; Saleh et al, 1996). Since NO may function as an upstream regulator of expression as well as a downstream effector of the action of VEGF, we examined whether the effects of DDAH and ADMA on tumour angiogenesis were mediated through changes in VEGF. In vitro analysis of VEGF mRNA production as well as VEGF protein secretion in the culture medium demonstrated that DDAH I expression stimulates VEGF production. The increase in VEGF production was similar to that induced by hypoxia in the same cells (V Kostourou, unpublished observations).
Our findings indicate that human tumours exhibit DDAH activity. Although this study is limited in number it does suggest that human brain tumours express particularly high levels of DDAH activity. Increased NO production has also been reported in human CNS tumours (Cobbs et al, 1995). Our experimental data demonstrates that DDAH I can promote tumour growth and increase tumour perfusion. Since tumours with a high grade of malignancy are better perfused and vascularised it is interesting to speculate that DDAH I could facilitate tumour progression and correlate with tumour malignancy. The relationship between DDAH activity and tumour grade is currently under investigation.
In summary, this study shows that increased DDAH expression results in decreased tumour ADMA, increased NO and VEGF production, vascularisation and tumour growth. Our results provide the first demonstration of the importance of the ADMA/DDAH pathway in the regulation of vessel development. It also has important implications for the regulation of angiogenesis in other pathological and physiological circumstances in which VEGF expression or NO production is involved. The fact that tumour development is so dramatically altered by regulating this pathway adds a novel level of complexity to a system where the precise balance between pro- and anti-angiogenic factors is of key importance. Therapeutic interventions in oncology using the DDAH/ADMA pathway are therefore plausible.
Acknowledgments
This work was supported by a St. George's Hospital Medical School (SGHMS) studentship to V Kostourou, the Cancer Research Campaign, UK, (CRC) grant SP1971/0404 and the British Heart Foundation (BHF). SP Robinson is the recipient of a Royal Society University Research Fellowship. The authors would like to thank Drs P Wood and J Leiper for helpful early discussions, Dr M Murphy for human tumour biopsies, Professor A Johnstone and JR Griffiths at SGHMS and Professor P. Vallance at UCL for their useful comments on the manuscript, and Mr C Brown and his staff for care of the animals.
References
- BarnaMKomatsuTReissCS1996Activation of type III nitric oxide synthase in astrocytes following a neurotropic viral infection Virology 223331343 [DOI] [PubMed] [Google Scholar]
- BarthRF1998Rat brain tumor models in experimental neuro-oncology: the 9L, C6, T9, F98, RG2 (D74), RT-2 and CNS-1 gliomas J Neurooncol 3691102 [DOI] [PubMed] [Google Scholar]
- CarmelietPJainRK2000Angiogenesis in cancer and other diseases Nature 407249257 [DOI] [PubMed] [Google Scholar]
- ChinKKurashimaYOguraTTajiriHYoshidaSEsumiH1997Induction of vascular endothelial growth factor by nitric oxide in human glioblastoma and hepatocellular carcinoma cells Oncogene 15437442 [DOI] [PubMed] [Google Scholar]
- ChinjeECStratfordIJ1997Role of nitric oxide in growth of solid tumours: a balancing act Essays Biochem 326172 [PubMed] [Google Scholar]
- CobbsCSBrenmanJEAldapeKDBredtDSIsraelMA1995Expression of nitric oxide synthase in human central nervous system tumors Cancer Res 55727730 [PubMed] [Google Scholar]
- DongZYStaroselskyAHQiXXXieKPFidlerIJ1994Inverse correlation between expression of inducible nitric-oxide synthase activity and production of metastasis in K-1735 murine melanoma-cells Cancer Res 54789793 [PubMed] [Google Scholar]
- FicklingSAToozeJAWhitleyGS1992Characterization of human umbilical vein endothelial-cell lines produced by transfection with the early region of Sv40 Exp Cell Res 201517521 [DOI] [PubMed] [Google Scholar]
- FicklingSAWilliamsDVallancePNusseySSWhitleyGS1993Plasma concentrations of endogenous inhibitor of nitric oxide synthesis in normal pregnancy and pre-eclampsia Lancet 342242243 [DOI] [PubMed] [Google Scholar]
- FolkmanJ1995Angiogenesis in cancer, vascular, rheumatoid and other disease Nature Medicine 12731 [DOI] [PubMed] [Google Scholar]
- FukumuraDYuanFEndoMJainRK1997Role of nitric oxide in tumor microcirculation. Blood flow, vascular permeability, and leukocyte-endothelial interactions Am J Pathol 150713725 [PMC free article] [PubMed] [Google Scholar]
- GalloOMasiniEMorbidelliLFranchiAFini-StorchiIVergariWAZicheM1998Role of nitric oxide in angiogenesis and tumor progression in head and neck cancer J Natl Cancer Inst 90587596 [DOI] [PubMed] [Google Scholar]
- GriffithsJRMcSheehyPMJRobinsonSPTroyHChungY-LLeekRDWilliamsKJStratfordIJHarrisALStubbsM2002Metabolic changes detected by in vivo magnetic resonance studies of Hepa-1 wild-type tumours and tumours deficient in hypoxia-inducible-factor-1b: evidence of an anabolic role for the HIF-1 pathway Cancer Res 62688695 [PubMed] [Google Scholar]
- HoldenDPFicklingSAWhitleyGSNusseySS1998Plasma concentrations of asymmetric dimethylarginine, a natural inhibitor of nitric oxide synthase, in normal pregnancy and preeclampsia Am J Obstet Gynecol 178551556 [DOI] [PubMed] [Google Scholar]
- JangJJHoHKKwanHHFajardoLFCookeJP2000Angiogenesis is impaired by hypercholesterolemia: role of asymmetric dimethylarginine Circulation 10214141419 [DOI] [PubMed] [Google Scholar]
- JenkinsDCCharlesIGThomsenLLMossDWHolmesLSBaylisSARhodesPWestmoreKEmsonPCMoncadaS1995Roles of nitric oxide in tumor growth Proc Natl Acad Sci USA 9243924396 [DOI] [PMC free article] [PubMed] [Google Scholar]
- KakimotoYAkazawaS1970Isolation and identification of N-G,N-G- and N-G,N′-G-dimethyl-arginine, N-epsilon-mono-, di-, and trimethyllysine, and glucosylgalactosyl- and galactosyl-delta-hydroxylysine from human urine J Biol Chem 24557515758 [PubMed] [Google Scholar]
- KimKJLiBWinerJArmaniniMGillettNPhillipsHSFerraraN1993Inhibition of vascular endothelial growth factor-induced angiogenesis suppresses tumor-growth in vivo Nature 362841844 [DOI] [PubMed] [Google Scholar]
- KimotoMWhitleyGSTsujiHOgawaT1995Detection of N-G,N-G-Dimethylarginine dimethylaminohydrolase in human tissues using a monoclonal-antibody J Biochem 117237238 [DOI] [PubMed] [Google Scholar]
- LeiperJMSanta MariaJChubbAMacAllisterRJCharlesIGWhitleyGSVallanceP1999Identification of two human dimethylarginine dimethylaminohydrolases with distinct tissue distributions and homology with microbial arginine deiminases Biochem J 343209214 [PMC free article] [PubMed] [Google Scholar]
- MacAllisterRJFicklingSAWhitleyGSVallanceP1994Metabolism of methylarginines by human vasculature - implications for the regulation of nitric-oxide synthesis Br J Pharmacol 1124348 [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacAllisterRJWhitleyGSParryHKimotoMOgawaTHodsonHMoncadaSVallanceP1996Regulation of NO synthesis by dimethylarginine dimethylaminohydrolase Br J Pharmacol 117P76P76 [DOI] [PMC free article] [PubMed] [Google Scholar]
- MatsuokaHItohSKimotoMKohnoKTamaiOWadaYYasukawaHIwamiGOkudaSImaizumiT1997Asymmetrical dimethylarginine, an endogenous nitric oxide synthase inhibitor, in experimental hypertension Hypertension 29242247 [DOI] [PubMed] [Google Scholar]
- MiyazakiHMatsuokaHCookeJPUsuiMUedaSOkudaSImaizumiT1999Endogenous nitric oxide synthase inhibitor – A novel marker of atherosclerosis Circulation 9911411146 [DOI] [PubMed] [Google Scholar]
- NehlsVDrenckhahnD1995A novel, microcarrier-based in vitro assay for rapid and reliable quantification of three-dimensional cell migration and angiogenesis Microvasc Res 50311322 [DOI] [PubMed] [Google Scholar]
- OgawaTKimotoMSasaokaK1989Purification and properties of a new enzyme, Ng,Ng-dimethylarginine dimethylaminohydrolase, from rat-kidney J Biol Chem 2641020510209 [PubMed] [Google Scholar]
- PapapetropoulosAGarcia-CardenaGMadriJASessaWC1997Nitric oxide production contributes to the angiogenic properties of vascular endothelial growth factor in human endothelial cells J Clin Invest 10031313139 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pipili-SynetosEPapageorgiouASakkoulaESotiropoulouGFotsisTKarakiulakisGMaragoudakisME1995Inhibition of angiogenesis, tumour growth and metastasis by the NO-releasing vasodilators, isosorbide mononitrate and dinitrate Br J Pharmacol 11618291834 [DOI] [PMC free article] [PubMed] [Google Scholar]
- ReveneauSArnouldLJolimoyGHilpertSLejeunePSaint-GiorgioVBelichardCJeanninJF1999Nitric oxide synthase in human breast cancer is associated with tumor grade, proliferation rate, and expression of progesterone receptors Lab Invest 7912151225 [PubMed] [Google Scholar]
- RobinsonSPRodriguesLMHoweFAStubbsMGriffithsJR2001Effects of different levels of hypercapnic hyperoxia on tumour R2* and arterial blood gases Magnetic Resonance Imaging 19161166 [DOI] [PubMed] [Google Scholar]
- SalehMStackerSAWilksAF1996Inhibition of growth of C6 glioma cells in vivo by expression of antisense vascular endothelial growth factor sequence Cancer Res 56393401 [PubMed] [Google Scholar]
- ShimaDTKurokiMDeutschUNgYSAdamisAPD'AmorePA1996The mouse gene for vascular endothelial growth factor. Genomic structure, definition of the transcriptional unit, and characterization of transcriptional and post-transcriptional regulatory sequences J Biol Chem 27138773883 [DOI] [PubMed] [Google Scholar]
- ShweikiDItinASofferDKeshetE1992Vascular endothelial growth-factor induced by hypoxia may mediate hypoxia-initiated angiogenesis Nature 359843845 [DOI] [PubMed] [Google Scholar]
- SimmonsMLMurphyS1993Cytokines regulate L-arginine-dependent cyclic-GMP production in rat glial-cells Eur J Neurosci 5825831 [DOI] [PubMed] [Google Scholar]
- SmithKAHillSABeggACDenekampJ1988Validation of the fluorescent dye Hoechst 33342 as a vascular space marker in tumours Br J Cancer 57247253 [DOI] [PMC free article] [PubMed] [Google Scholar]
- StuehrDJNathanCF1989Nitric oxide. A macrophage product responsible for cytostasis and respiratory inhibition in tumor target cells J Exp Med 16915431555 [DOI] [PMC free article] [PubMed] [Google Scholar]
- TakahashiYKitadaiYBucanaCDClearyKREllisLM1995Expression of vascular endothelial growth-factor and its receptor, Kdr, correlates with vascularity, metastasis, and proliferation of human colon-cancer Cancer Res 5539643968 [PubMed] [Google Scholar]
- ThomsenLLLawtonFGKnowlesRGBeesleyJERiverosmorenoVMoncadaS1994Nitric-oxide synthase activity in human gynecological cancer Cancer Res 5413521354 [PubMed] [Google Scholar]
- ThomsenLLScottJMJTopleyPKnowlesRGKeerieAJFrendAJ1997Selective inhibition of inducible nitric oxide synthase inhibits tumor growth in vivo: Studies with 1400W, a novel inhibitor Cancer Res 5733003304 [PubMed] [Google Scholar]
- TrachtmanHFutterweitSGargPReddyKSinghalPC1996Nitric oxide stimulates the activity of a 72-kDa neutral matrix metalloproteinase in cultured rat mesangial cells Biochem Biophys Res Commun 218704708 [DOI] [PubMed] [Google Scholar]
- WhitleyGS1994Extraction and assay of cyclic nucleotides Methods Mol Biol 27189197 [DOI] [PubMed] [Google Scholar]
- XieKHuangSDongZJuangSHGutmanMXieQWNathanCFidlerIJ1995Transfection with the inducible nitric oxide synthase gene suppresses tumorigenicity and abrogates metastasis by K-1735 murine melanoma cells J Exp Med 18113331343 [DOI] [PMC free article] [PubMed] [Google Scholar]
- YancopoulosGDDavisSGaleNWRudgeJSWiegandSJHolashJ2000Vascular-specific growth factors and blood vessel formation Nature 407242248 [DOI] [PubMed] [Google Scholar]
- ZhangHTCraftPScottPAZicheMWeichHAHarrisALBicknellR1995Enhancement of tumor growth and vascular density by transfection of vascular endothelial cell growth factor into MCF-7 human breast carcinoma cells J Natl Cancer Inst 87213219 [DOI] [PubMed] [Google Scholar]
- ZicheMMorbidelliLMasiniEAmeriniSGrangerHJMaggiCAGeppettiPLeddaF1994Nitric oxide mediates angiogenesis in vivo and endothelial cell growth and migration in vitro promoted by substance P J Clin Invest 9420362044 [DOI] [PMC free article] [PubMed] [Google Scholar]