

Abstract
Intra-fixation location changes were measured when one-line sentences written in lower or aLtErNaTiNg case were read. Intra-fixation location changes were common and their size was normally distributed except for a relatively high proportion of fixations without a discernible location change. Location changes that did occur were systematically biased toward the right when alternating case was read. Irrespective of case type, changes of the right eye were biased toward the right at the onset of sentence reading, and this spatial bias decreased as sentence reading progressed from left to right. The left eye showed a relatively stable right-directed bias. These results show that processing demands can pull the two fixated eyes in the same direction and that the response to this pull can differ for the right and left eye.
Keywords: Fixation Duration, Saccade, Fixation-Change, Attention, Reading
Eye movements during reading consist of discrete jumps (saccades) that are separated by distinct viewing pauses during which the eyes are assumed to be relatively immobile, i.e., ‘fixated’. Detailed oculomotor observations during individual fixations (e.g., Ditchburn, 1955; Ditchburn & Fender, 1955; Martinez-Conde, Macknik, & Hubel, 2004; Riggs, Ratcliff, Cornsweet, & Cornsweet, 1953) showed, however, that the eyes were not completely stable. Instead, they executed miniature movements that were assumed to prevent retinal adaptation, as effective vision is quickly lost when these movements are suppressed or counteracted (Ditchburn, 1955; Ditchburn & Fender, 1955; Inhoff & Topolski, 1994; 1995; Riggs et al., 1953).
Ditchburn and Fender (1955) discriminated two types of intra-fixation movements, drifts and flicks. Drifts are gradual changes in viewing location that move the eyes smoothly across a relatively small distance from a few seconds of arc to about 1 degree of visual angle. Drift during a fixation is not linear and a movement that starts out in one direction may be changed and continued in another direction, the net effect often being an approximately zero-size location change. Flicks, or microsaccades, move the eyes across a similar distance, but they do so rapidly. Microsaccades during prolonged fixations show a biphasic pattern (Engbert & Kliegl, 2004), with early microsaccades moving away from and later microsaccades moving toward the to-be-maintained fixation position. Both types of miniature movement thus create a dynamic balance between two spatial biases, one that shifts the eyes slightly away from a fixated location, presumably to prevent retinal adaptation, and one that directs the eyes back toward the desired viewing position. In this scheme, the eyes execute micro-movements during individual fixations that are directionally neutral. The fixation location at the beginning of a fixation will typically differ little from the fixation location at the end of the fixation, and location changes that do occur should not be subject to a systematic spatial bias.
Visual tasks that require the acquisition of detailed information from different spatial areas involve the execution of a sequence of saccades, as the visual interest area often extends beyond the range of high acuity vision. In reading, useful visual and linguistic information is obtained from a relatively small region, and readers must execute a sequence of saccades. Each saccade typically moves the eyes across 6–10 letter spaces (LS) to identify the sequence of to-be-read words on a line of text. The intervening reading fixations are relatively short, typically between 200 ms to 500 ms, with a grand mean of approximately 220–250 ms (see Rayner, 1998, for a comprehensive description). Binocular measurement in this task showed that the line of gaze of the two eyes typically does not occupy the same spatial location at the onset of a fixation, with the discrepancy often amounting to more than one letter space (LS) (Heller & Radach, 1999). This binocular disparity appears to be the source of another type of intra-fixation movement that seeks to reduce it (Blythe, Liversedge, Joseph, White, Findlay, & Rayner, 2006; Hendricks, 1996; Juhasz, Liversedge, White, & Rayner, 2006; Liversedge, White, Findlay, & Rayner, 2006; Radach, Heller, & Jaschinski, 1996).
In one of the most detailed studies of post-saccadic binocular activity during reading, Liversedge et al. (2006) found that the position of the two eyes differed by more than one LS at the beginning of the majority of fixations (52%), and that the line of gaze of the left eye was to the left of the line of gaze of the right eye on the vast majority of fixations (referred to as uncrossed fixations). The two eyes typically converged toward each other (52% of fixations), but convergence did not eliminate binocular disparity. Instead, it was reduced from mean disparity of 1.3 LS at fixation onset to 1.1 LS at fixation offset. This disparity was not always accomplished by symmetric movements of the eyes toward each other. The convergence could have been accomplished primarily by in intra-fixation movement of one eye that moved toward the other or both eyes moved in the same direction, with one eye moving a larger distance than the other. For uncrossed fixations, vergence occurs, for instance, when the left eye executes a larger shift toward the right than right eye; for crossed fixations, vergence occurs when the right eye executes a larger right-directed shift.
Hendricks (1996) found that convergence of the two eyes increased with genera processing demands during passage reading. Word-specific manipulations of linguistic processing difficulty, via the use of high and low frequency target words and words that were written either in lower case or aLtErNaTiNg case, did not influence retinal disparity (Juhasz et al., 2006; see also Blythe et al., 2006), however. This occurred even though word frequency and letter case manipulations themselves had a profound effect on the time spent viewing individual target words. Vergence movements of the two eyes during reading, therefore, appear to be immune to the demands of visual word recognition during reading.
All prior examinations of intra-fixation movements during reading fixations were concerned with movements of the two eyes in relation to each other. From this perspective, the absolute size and direction of intra-fixation location changes of each eye are relevant only in so far as they influence the spatial distance between the fixation locations of the two eyes. The spatial properties of intra-fixation location changes themselves are abstracted away, which means that potentially useful oculomotor information could be lost. Processing demands during reading could influence the size and direction of intra-fixation movements without effecting binocular disparity
In particular, intra-fixation location changes could be influenced by the location of a fixation on a line of text. Vitu, Kapoula, Lancelin, and Lavigne (2004) showed that saccades to peripheral words or letter strings were pulled toward the screen center. Saccades to equidistant horizontal targets were larger when the saccade was directed toward, rather than away from, the screen center. Analogously, the size and direction of horizontal intra-fixation location changes could depend on the proximity of a fixation to the center. When a fixation is to the left of the screen center, intra-fixation location changes may be pulled toward the right, and when a fixation is to the right of the screen center, intra-fixation location changes may be pulled toward the left. The absolute size of intra-fixation location changes could also change as a function of linguistic processing demands. Intra-fixation location changes of each eye could be relatively small when a word is easy to identify, and they could increase with the difficulty of the word identification task. It could also be the case, however, that each eye becomes increasingly ‘fixed’ as linguistic processing difficulty increases.
Similar to prior studies that established normative data regarding the binocular coordination of the eyes during reading (e.g., Blythe et al., 2006; Liversedge et al., 2006; Juhasz et al., 2006), the current study compared the line of gaze (fixation location) at the beginning of a fixation with the line of gaze at the end of a fixation to determine the magnitude of intra-fixation location changes. However, rather than focusing on changes of the two eyes in relation to each other, we focused on the direction, magnitude, and frequency of intra-fixation location changes of individual eyes, the right eye in Experiment 1 and the right and left eye in Experiment 2. This was predicated on the assumption that the direction and absolute size of intra-fixation location changes of each eye could be influenced by general demands of the reading task. This may occur even though some of these demands did not influence intra-fixation movements of the two eyes in relation to each other in earlier work.
Two major goals were pursued: to establish normative data on the spatial properties of intra-fixation location changes of each eye during reading fixations and to determine whether these location changes are reflexive line of gaze corrections or whether they are subject to the influence of task-related text properties and processing demands. Together, Experiment 1 and Experiment 2 yielded over 65,000 saccade-fixation-saccade sequences and this database was used to describe general properties of intra-fixation location changes of the right and left eye.
Neither Experiment 1 nor Experiment 2 included controlled manipulations of spatial and lexical text properties. Since words are ordered from left-to-right on a line of text, word order was used to index the relative horizontal location of a word on the screen. The initial words of a sentence were always to the left of the screen center and the ending words to its right. If intra-fixation location changes were directed toward the visual screen center, then intra-fixation location changes should be biased toward the right when the beginning words of a sentence are fixated and toward the left when the ending words are fixated.
The length of individual words was used to index ease of visual word recognition. The orthographic, phonological, and morphologic complexity of a word typically increases with its length, and the frequency and length of a word are negatively correlated (New, Ferrand, Pallier, & Brysbaert, 2006). If ease of visual word recognition influenced intra-fixation location changes, then the frequency, direction, and size of these changes should be a function of the length of a fixated word.
Since oculomotor activity was recorded from only the right eye, we could not determine the extent to which its intra-fixation shifts were influenced by movements of the two eyes in relation to each other (see Blythe et al., 2006; Liversedge et al., 2006, for detailed accounts of binocular coordination in reading). To determine whether the magnitude and direction of intra-fixation movements of the recorded right eye are influenced by movements of the left eye, Experiment 1 included a subgroup of six participants who read text under binocular and monocular viewing conditions while movements of the right eye were recorded.
Experiment 1
Method
Participants
Two separate groups of participants were tested both of which consisted of undergraduate students at Binghamton University (State University of New York) who participated in exchange for experimental course credit. One set of participants consisted of twenty undergraduate students who read the experimental materials without any vision constraints. A smaller set of six subjects read the experimental material under binocular and monocular viewing conditions. All participants considered themselves fluent readers, all had normal or corrected to normal vision, and all were naïve with regard to the purpose of the experiment.
Materials
The materials consisted of 160 declarative sentences each of which contained between 10–15 words. All sentences occupied a single line of text, and only one sentence was visible at a time. Except for the first letter of each sentence, all text was written in lower case. All sentences were relatively easy to comprehend for college level readers, and they were devoid of syntactic and linguistic ambiguities. Each sentence contained a specific target word the parafoveal visibility of which was manipulated so that a reader either saw the intact target throughout sentence reading or a length-matched nonword at the target’s location until the eyes crossed an invisible pre-target boundary. The target word was always fully visible when the eyes reached its spatial location, and fixations on the target word were therefore included in the analyses of intra-fixation location changes.
Apparatus and Procedure
All text was shown on a 21-inch flat screen monitor in Courier font with a 1024*768 pixel resolution. Eye movements were recorded with a SR Research Eyelink 1000 tracking system. The head of the reader was held in place with chin and head rests, and eye movements were recorded with a camera that occupies a fixed location. Eye position was sampled at a rate of 1000 Hertz, with a relatively high spatial resolution of approximately .1 LS. All recordings were obtained from a reader’s right eye, as this eye has been preferentially monitored in the majority of published reading studies. Fixation locations were sampled on-line and used off-line to determine saccade size, fixation duration, fixation location, and the size and direction of intra-fixation location changes.
Participants were tested individually. Twenty participants viewed all sentences with both eyes which has been the standard procedure in prior reading experiments. Six additional participants viewed one block comprising half of the experimental sentences with both eyes in a binocular viewing condition and the remaining block of sentences with the left eye occluded in a monocular viewing condition (the order of binocular and monocular sentence blocks was counterbalanced across readers). A horizontal calibration of the eye tracking system preceded the experiment. During this calibration, the reader was asked to fixate a sequence of four fixation markers as they appeared in random order for 1s at the right, left, and center location at the horizontal midline of the screen (the left side location corresponded to the position of the first letter of a sentence). The initial calibration was followed by a validation routine that determined the stability of the calibration. All calibrations and validations of the monitored right eye were performed under binocular viewing conditions in the binocular text viewing condition1 and with the left eye occluded in the monocular viewing condition. To encourage reading for meaning, participants were asked to report the previously read sentence on approximately every 10th trial. All readers were able to report sentence content upon questioning with an accuracy rate of >95%.
Oculomotor measures
Commercially available software (SR Research) was used to parse the continuous stream of fixation locations into fixations and saccades. The software detected the onset of a reading saccade when eye movement velocity was greater than 30 degrees/s and when acceleration exceeded 8000 degrees/s2. The saccade was considered terminated when velocity was less than 30 degrees/s, and the time in-between saccades defined the duration of a fixation. Similar fixation-saccade-fixation parsing criteria have been used in the vast majority of published reading experiments.
To determine the magnitude and direction of intra-fixation location changes, we determined the fixation location (to the nearest pixel location) at the onset of a fixation and then again at the offset of a fixation. Rather than using the two points in time at which the signal stream separated a saccade from a fixation, we used a slightly more conservative estimate of on- and offset locations. Specifically, we determined the mean fixation location slightly after an incoming saccade, during the 6–8 ms period following saccade offset, and the mean location during the 8–6 ms period that preceded an outgoing saccade. For a fixation of 250 ms, this meant that intra-fixation location changes were determined that took place during the ‘middle’ 238 ms of that fixation. This particular measurement solution was chosen to reduce the possibility that intra-fixation position changes were compromised by position changes that were due to movement spillover from incoming or outgoing saccades. On- and offset fixation location measures were converted to a letter space (LS) scale, and location changes to the right were given positive LS values and changes to the left were given negative LS values. No location change was assumed to have occurred when on- and offset locations differed by less than +/−.05 LS from zero, and all other on- and offset differences were measured in .1 LS increments. For instance, a movement size of .1 LS was assigned when on- and offset locations differed by .05 to .14 LS, and a movement size of .2 was assigned when on- and offset locations differed by .15 to .24 LS. In addition, we obtained other oculomotor measures that are typically obtained in studies of reading, including saccade size (and direction), fixation duration, and gaze duration. Gaze duration consisted of the cumulated time spent viewing a word until another word was fixated. If, for instance, a word received three fixations before the eyes moved to a different word, then gaze duration consisted of the sum of the three fixation durations, and it included the time spent on the two intervening saccades.
Binocular vs. monocular sentence reading
Intra-fixation location changes from the twenty readers who read all sentences without any vision impediment were of primary interest. These data were used to obtain normative data on intra-fixation movement dynamics of the right eye and to determine whether intra-fixation location changes are influenced by visual and/or linguistic processing demands. Data from the six additional participants who read under binocular and monocular viewing conditions were collected to determine whether intra-fixation movements of the right eye differ under binocular and monocular viewing conditions.
Data Selection
Overall, the set of raw data from twenty readers who read text without any vision constraint consisted of 40,426 right eye saccades and fixations. From this data base, we excluded all first and last fixations on a sentence, irrespective of the location of these fixations, and we excluded all fixations on the last word of a sentence, as there was no text after that. Furthermore, we excluded all fixations that followed unusually small or large incoming and outgoing saccades of less than 1 or of more than 20 LS, and we removed all outlying fixations with durations of less than 30 ms and of more than 1000 ms. The remaining database consisted of 30,892 saccade-fixation-saccade sequences, or 76% of the full set of data. We first examined fixation durations, saccade sizes, and gaze durations to determine whether oculomotor activity in Experiment 1 was similar to oculomotor activity in previously reported studies of reading. We then examined intra-fixation location changes of the right eye to obtain normative data regarding the frequency of their size and direction. Finally, we examined whether intra-fixation location changes were influenced by visual and linguistic processing demands. The data from six additional readers who read the experimental materials under monocular and binocular viewing conditions were subject to the same selection criteria as the primary set of data. This yielded another 6,876 saccade-fixation-saccade sequences with virtually the same number of fixations in the monocular and binocular conditions (3,402 and 3,474, respectively).
Results
Figure 1 shows the frequency distribution of 30,892 right eye fixation durations from the twenty participants who read all sentences without any vision constraints. Figure 2 shows the corresponding frequency distribution for incoming saccade sizes. Both distributions are quite similar to previously reported data (e.g., Rayner & Pollatsek, 1989). Other indexes of oculomotor activity also reveal a typical pattern of oculomotor activity. The vast majority of saccades was directed toward the right (approximately 92%) and the relative frequency with which words received a fixation (and more than one fixation), increased linearly with word length. Mean fixation (and refixation) rate for words with 3 to 10 constituent letters was 39% (3%), 48% (5%), 75% (10%), 83% (14%), 93% (17%), 96% (19%), 99% (23%), and 100% (26%), respectively. The effect of word length on fixation rate is very similar to previously reported findings (e.g., Brysbaert & Vitu, 1998; Vitu, O’Regan, Inhoff, & Topolski, 1995; see also Rayner, 1998).
Figure 1.

The relative frequency of fixation durations during the reading of lower case text in Experiment 1.
Figure 2.

The relative frequency of saccade sizes during the reading of lower case text in Experiment 1.
Word length also influenced the time spent viewing a word. The mean fixation duration for words with 3 to 10 constituent letters was 243 ms, 246 ms, 253 ms, 250 ms, 280 ms, 260 ms, 282 ms, and 285 ms, respectively, F(7,133) = 31.82, p<.01. Word length had a profound effect on gaze duration which increased linearly with length from 258 ms, to 280 ms, 307 ms, 333 ms, 375 ms, 387 ms, 442 ms, and 481 ms for words with 3 to 10 constituent letters, F(7,133) = 71.16, p<.01.
Figure 3 shows the frequency distribution of intra-fixation location changes of the right eye during reading fixations. The grand mean of the distribution was close to zero, .03 LS, and fixations with no discernible location change were atypically common. The eye was not immobile, however. It moved more than .1 LS on 86.6% of reading fixations, and there was considerable variability in the size of location changes (SD = .85 LS)2. A paired comparison that examined the size of these intra-fixation location changes as a function of movement direction (right vs. left) did not reveal a reliable difference, t(19)<.5, and movements toward the right were not significantly more common than movements toward the left (54% and 46%, respectively), t(19) <.7.
Figure 3.

The relative frequency of location changes during reading fixations.
Intra-fixation location changes toward the left (46%) were much more common than left-directed incoming saccades (8%), indicating that the direction of intra-fixation micro-movements was not an extension of the direction of the incoming saccade. Consistent with this view, left-directed intra-fixation location changes of .1 LS or more were also relatively common (37%) even when the incoming and the following outgoing saccade were right-directed. Correlations of the mean size of in- and outgoing saccades with the mean size of intra-fixation movements (saccades to the left and right were given negative and positive values, respectively) support this view. Across readers, the mean correlations between in- and out-going saccades with the size of intra-fixation location changes were quite small and not statistically reliable, r=.05 (n.s.), and r=.07 (n.s.), respectively.
To determine whether the eyes were pulled toward the center of the screen, location changes were examined as a function of word order. The exclusion of the first fixation on a sentence prevented a meaningful analysis of its first word. Not all sentences contained more than 11 words, and the last word of each sentence was routinely excluded as there was no text after that. Consequently, we analyzed the size of intra-fixation location changes as a function of word order for words 2–10 in each sentence. The results revealed a large and systematic effect of order. During the fixation of the second word at the leftmost examined screen location, intra-fixation locations changed by .18 LS, and this right-directed bias decreased systematically to .14 LS, .11 LS, .06 LS, .07 LS, −.01 LS, −.03 LS, −.02 LS, and −.04 LS for word locations 3 to 10, respectively, F(8,152) = 10.30, p<.01.
More detailed analyses that examined the frequency of location changes further showed that there was a systematic decrease in the proportion of right-directed location changes from words 2 to 10, with corresponding proportions of 63%, to 61%, 60%, 58%, 53%, 52%, 50%, 51%, and 49%, respectively. Moreover, the size of right- and left-directed location changes was a function of word order. Right-directed location changes became smaller and left-directed changes became larger as the eye moved from left to right through a sentence. The corresponding interaction, shown in Figure 4, was statistically significant, F(8,152) = 3.95, p<.01.
Figure 4.

Mean size (in LS) of fixation location changes of the right eye as a function of change direction. Mean values are shown with standard errors of the mean.
In spite of sizable effects of word length on fixation duration and gaze duration, word length did not influence intra-fixation location changes. The mean sizes of location changes were .07 LS, .06 LS, .07 LS, .01 LS, .05 LS, .06 LS, .04 LS, and .04 LS, for words with 3 to 10 constituent letters, respectively, F(7,133) = 1.67, p>.12. Location changes during reading fixations, therefore, do not appear to be influenced by the ease of visual word recognition (see also Juhasz et al., 2006; Blythe et al., 2006).
These intra-fixation location changes occurred under binocular viewing conditions, i.e., when the right and left eye may have fixated slightly different text locations. To determine the extent to which binocular visibility of text influenced intra-fixation movements, we examined the size of these shifts for the six readers who read the experimental materials under binocular and monocular viewing conditions. The two corresponding frequency distributions are shown in Figure 5. As can be seen the two distributions are nearly identical. Intra-fixation movements of the right eye were therefore very similar under monocular and binocular viewing conditions.
Figure 5.
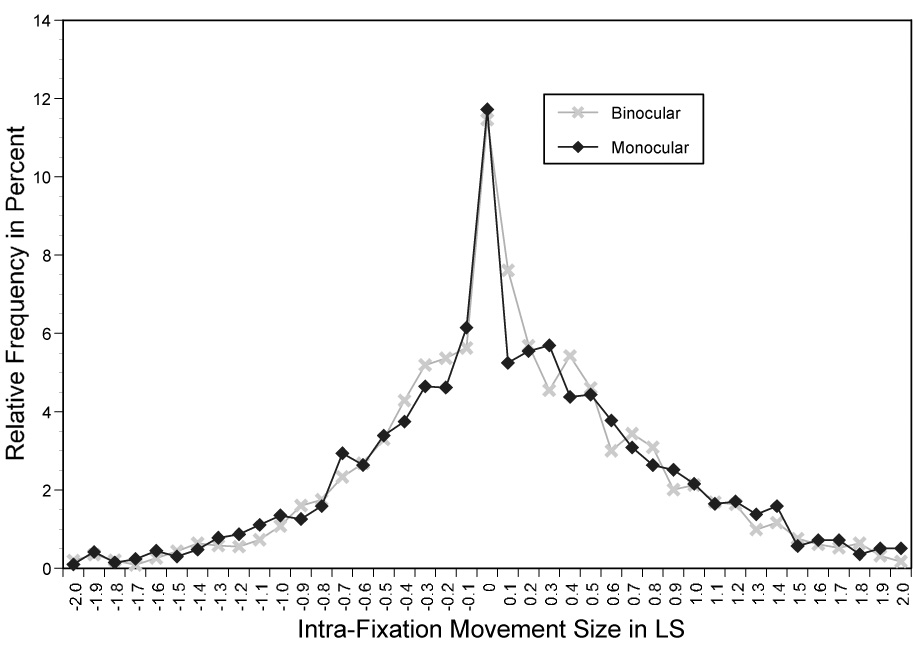
The relative frequency of intra-fixation location changes of the right eye under binocular and monocular viewing conditions.
Discussion
The examination of a relatively large data base of intra-fixation location changes of the right eye revealed an almost standard Gaussian distribution of shift sizes with a mean value close to zero and a SD of .85 LS. The one exception to the normal distribution of shift size frequencies was the relatively high proportion of fixations with no discernible location changes. Intra-fixation micro-movements of the right eye were also independent of the spatial properties of in- and outgoing saccades, and they were immune to the demands of visual word recognition.
Intra-fixation movements were, however, subject to a strong effect of word order. Right-directed location change were relatively large and frequent when the left-most words on a line of text were fixated, and this spatial bias decreased systematically until shift size was close to zero when words five and six were fixated. After that, fixations were pulled slightly in the opposite direction. Since words five and six were typically close to the horizontal screen center, these results are consistent with the assumption that the fixating right eye was pulled toward the center, and that the strength of the pull decreased as the eye moved closer to it.
Other accounts cannot be ruled out, however. The screen center was aligned with the head midline, and the right eye could have been pulled toward that. Moreover, this pull could have been asymmetrical. The right eye crossed the head midline when it fixated a word on the left half the screen but not when it fixated a word near the middle or on the right side of the screen. Consequently, the fixating right eye could have been pulled toward the right when its line of gaze crossed the head midline, and this pull decreased and then ceased as the line of gaze approached and then crossed the head midline. Consonant with this, mean intra-fixation location changes exceeded .1 LS only when words 2, 3, and 4 of a sentence were fixated, i.e., when the line of gaze was likely to have crossed the head midline.
Experiment 2
Experiment 2 differed from Experiment 1 in two critical aspects. First, intra-fixation location changes were obtained from each of the two eyes. If intra-fixation location changes of both eyes were generally pulled toward the screen center or the head midline, then intra-fixation location changes of the right and left eye should be biased toward the right when a fixation is near the left side of the screen, i.e., when the beginning words of a sentence are fixated, and this bias should disappear and eventually reverse as the eyes move from left to right though a sentence. If, however, the location change bias during reading fixations occurred primarily when a fixating eye crossed the head midline, then the right eye should show a more distinct right-directed intra-fixation location change pattern than the left eye when the beginning words of a sentence are read and the left eye should show a more distinct left-directed shift pattern when the ending words of a sentence are read.
Experiment 2 further differed from Experiment 1 in that sentences were written in alternating case. This eliminated the familiar spatial configuration of words and created two sets of same-case letter groups for each word, each of which was likely to receive some individual processing (Mayall & Humpreys, & Olson, 1997; Humpreys, Mayall, & Cooper, 2003). Word recognition, therefore, involved the fusion of these two visually distinct letter groups. Although letter case does not influence the magnitude of binocular vergence (Juhasz et al., 2006), the integration of two spatially discontinuous letter groups into a word could, nevertheless, influence the magnitude of intra-fixation location changes. The integration of visually distinct and interleaved letter groupings requires the use of order information and this could bias intra-fixation location changes in a congruent direction.
Method
Participants
Forty-five undergraduate students at the State University of New York at Binghamton volunteered to participate in the experiment. All were fluent readers, all were native with respect to the purpose of the experiment, and all had normal or corrected to normal vision (contact lenses). The right eye of 15 readers and the left eye of 30 readers was monitored.
Materials
The materials consisted of 87 declarative sentences that had been used in experiments of an earlier study (Inhoff, Eiter, & Radach, 2005). They contained between 11–15 words that occupied up to 80 LS. All sentences occupied a single line of text and only one sentence was visible at a time. All text was shown in alternating case. Participants received practice reading alternating text so that it was relatively familiar. As in Experiment 1, each sentence contained a specific target word whose parafoveal visibility was manipulated so that a reader either saw the intact target throughout sentence reading or a length-matched nonword until the eyes crossed an invisible spatial pretarget boundary. The target word was always fully visible when the eyes reached its spatial location (or the blank space preceding it), and fixations on the target were therefore included in the analysis of intra-fixation movements.
Apparatus and Procedure
The identical apparatus and procedure were used as in Experiment 1. Only one eye was tracked at a time due to limitations of the Eyelink 1000 system. We preferentially recorded movements of the left eye (30 readers) as Experiment 1 provided a relatively large data base of right eye fixations. Text was always visible to both eyes, irrespective of whether the right or left eye was monitored.
Oculomotor measures and Data Selection
The same selection criteria were used as in Experiment 1. The to-be-read number of words in Experiment 2 was slightly less than half of the number of to-be-read words in Experiment 1. Nevertheless, the set of eligible corpus data was almost as large as in Experiment 1, 30,190 saccade-fixation-saccade sequences, due to the larger number of participants in Experiment 2.
Results
Figure 6 and Figure 7 show the relative frequencies of fixation durations and of incoming saccade sizes for the right and the left eye. The frequency distributions for fixation durations and saccade sizes of the right and left eye overlapped substantially. Fixation durations were numerically longer when the left eye was recorded (257 ms) than when the right eye was recorded (244 ms), t(43) = 1.39, p=.79, but there was virtually no difference in the size of forward directed saccades, both approximately 6.8 LS, t~0. Relatively few saccades (8%) of the right and left eye were regressions, as was the case in Experiment 1, and there was no difference between the two eyes, t~0. The fixation rate of words again increased with word length for words with 3–10 constituent letters, with right and left eye fixation rates of 57% (right eye) vs. 58% (left eye), 73% vs. 73%, 87% vs. 87%, 90% vs. 92%, 94% vs. 94%, 97% vs. 96%, 97% vs. 96%, and 98% vs. 98%, respectively. Refixation rates for words with 3 to 10 constituent letters revealed a corresponding word-length effect for the right and left eye, 2.4% vs. 2.6%, 6% vs. 7%, 22% vs. 24%, 19% vs. 21%, 25% vs. 24%, 36% vs. 34%, 38% vs. 39%, and 45% vs. 42%, respectively. Compared to the reading of lower case text in Experiment 1, alternating case resulted in slightly longer fixation durations and distinctly shorter saccades (see also Inhoff et al., 2005; Juhasz et al., 2006). The shorter saccade size also resulted in a higher rate of word fixations and refixations than in Experiment 1. Critically, however, there were no differences between the oculomotor activity of the right and the left eye, all F<1.
Figure 6.

The relative frequency of fixation durations of the right and left eye during the reading of alternating case text.
Figure 7.
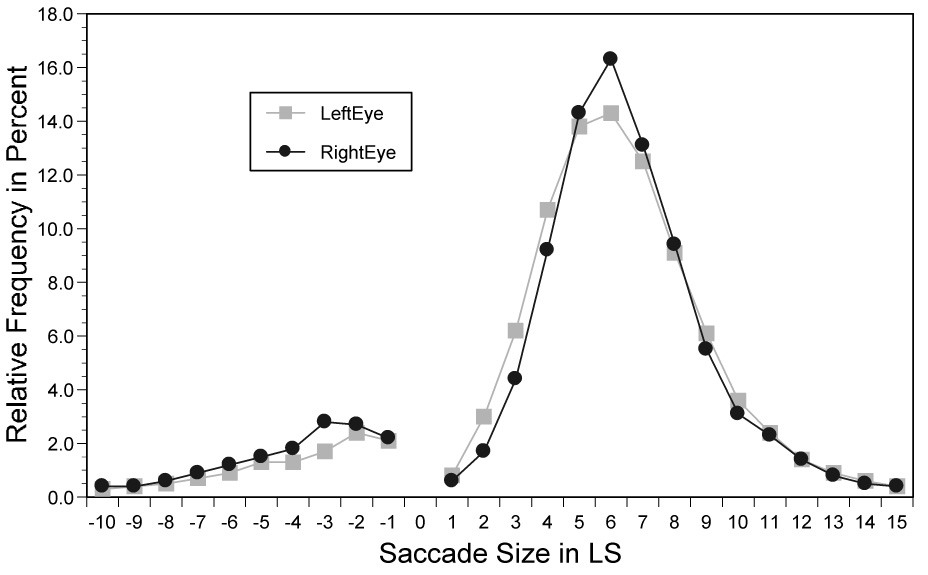
The relative frequency of saccade sizes of the right and left eye during the reading of alternating case text.
As in Experiment 1, word length had a profound effect on mean fixation duration and gaze duration (see Table 1). Similar to Experiment 1, mean fixation duration increased with word length for words with 3–5 constituent letters and then leveled off. An ANOVA with the factors word length (3–10) and eye (left vs. right) revealed a robust main effect of word length, F(7,301) = 12.63, p<.01, and a trend toward slightly longer left eye mean fixation durations, F(1,43) = 2.29, p<.15, especially for long words, which resulted in a marginally reliable interaction, F(7,301) = 1.86, p<.08. The source for this is unclear. Critically, differences between the right and left eye were no longer evident in the gaze data. As was the case in Experiment 1, increases in word length resulted in a near linear increase of gaze duration, F(3,301) = 89.62, p<.01. The main effect of monitored eye (right vs left) and the interaction of word length with monitored eye were now negligible, both F<1.
Table 1.
Mean fixation duration and gaze duration as a function of word length and recorded eye.
| Mean Fixation Duration | Length 3 | Length 4 | Length 5 | Length 6 | Length 7 | Length 8 | Length 9 | Length 10 |
|---|---|---|---|---|---|---|---|---|
| Left Eye | 241 | 257 | 265 | 269 | 266 | 264 | 267 | 258 |
| Mse | 5 | 5 | 5 | 6 | 6 | 6 | 6 | 8 |
| Right Eye | 234 | 246 | 255 | 250 | 259 | 252 | 248 | 233 |
| Mse | 7 | 7 | 8 | 8 | 9 | 8 | 8 | 11 |
| Gaze | ||||||||
| Left Eye | 249 | 281 | 317 | 336 | 353 | 370 | 396 | 390 |
| Mse | 5 | 7 | 9 | 11 | 12 | 13 | 15 | 15 |
| Right Eye | 241 | 267 | 301 | 315 | 342 | 357 | 373 | 363 |
| Mse | 7 | 9 | 12 | 15 | 17 | 19 | 21 | 21 |
Figure 8 shows the frequency distribution of intra-fixation location changes for the right and left eye. In stark contrast to Experiment 1, the grand mean of intra-fixation movements was no longer close to zero. Instead, both eyes revealed a systematic bias toward the right, with larger and more frequent right-directed than left-directed location changes. The grand means of intra-fixation location changes of the left and right eye were .27 LS and .11 LS, respectively, and both location change sizes were significantly larger than zero, t(29) = 5.84, p<.01, and t(14) = 2.99, p<.01. The left eye’s intra-fixation location changes toward the right were also significantly larger than the location changes of the right eye, t(43) = 3.09, p<.05.
Figure 8.

The relative frequency of intra-fixation location changes of the right and left eye during the reading of alternating case text.
In spite of the relatively strong right-directed location change bias, there was again no systematic relationship between spatial properties of in- and out-going saccades and intra-fixation location changes. The correlation between in- and out-going saccades and intra-fixation movements were small for the right eye r=.14 and r=.09 and for the left eye r=.15 and r=.09, respectively (all p>.25).
As in Experiment 1, intra-fixation location changes of the right eye were a function of word order. A relatively large right-directed movement bias during the fixation of word two decreased systematically throughout sentence reading. In contrast to Experiment 1, a small right-directed movement bias was present even during the reading of the final words of a sentence. Intra-fixation location changes of the left eye were more strongly biased toward the right and the size of this bias decreased only slightly with word order (except or the atypical increase from location two to three). The corresponding ANOVA revealed a robust main effect of word order, F(8,344) = 3.96, p<.01, of monitored eye, F(8,344) = 3.30, p<.01, and a significant interaction of the two, F(8,344) = 2.62, p<.01 (see Figure 9).
Figure 9.

Intra-fixation location changes of the right and left eye as a function of the word order. Condition means and mean standard errors are shown.
Although a word’s length influenced the time spent viewing it, word length had once more no discernible effect on intra-fixation location changes (see Figure 10), and neither the main effect of word length nor the interaction of word length with monitored eye were reliable, both F<1. The main effect of recorded eye was reliable, F(1,43) = 4.64, p<.05, due to larger right-directed location changes of the left eye. Ease of visual word identification therefore does not influence intra-fixation location changes.
Figure 10.

Intra-fixation location changes of the right and left eye as a function of word length (in letters). Condition means and mean standard errors are shown.
Discussion
The results of Experiment 2 largely replicated those of Experiment 1: the eyes typically shifted during reading fixations and the distribution of movement sizes was near Gaussian except for the large frequency of near zero-size location changes. As in Experiment 1, the location on a word on a line of text influenced intra-fixation movements and word length did not. Experiment 2 also extended Experiment 1 in two critical aspects. First, it showed that word location influenced intra-fixation movements of the right eye to a larger extent than those of the left eye. Second, it revealed systematic right-directed intra-fixation movements that were considerably larger than in Experiment 1.
The larger effect of word location on intra-fixation location changes of the right than the left eye is theoretically informative, as it disagrees with visual and oculomotor accounts. If micro-movements of during a fixation pulled the eyes toward the screen center, then both eyes should have exhibited a synchronous bias toward the right during the reading of the beginning words of a sentence and toward the left during the reading of the ending words of a sentence which was not the case. If intra-fixation location changes had occurred primarily when the fixating eye crossed the head midline, then the right and left eye should have shown complementary effects of word order which was also not the case.
Differences between the right and left eye could be taken as evidence for the influence of ocular dominance. (Porac and Coren 1976) discriminated between three functionally distinct types two of which, motor dominance and directional dominance, appear relevant3. Motor dominance is related to the movement size of an eye during a fixation, with the dominant eye moving to a smaller extent. This dominance can explain the overall smaller size of location changes of the right eye, assuming that most readers were right eye motor-dominant. It cannot explain, however, the effects of either case type or word order. Additional processing assumptions must be invoked to account for these findings. Direction dominance is typically measured with the porta test in which the observer extends one arm with a raised thumb that is aligned with a distant object and then determines how the alternative opening and closing of each eye aligns the object with the thumb. The dominant eye reveals a more stable object perception, and this could translate into smaller intra-fixation movements if the majority of readers were right eye direction-dominant. But this type of dominance also cannot account for case type or word order effects. Again, additional processing assumptions must be invoked. The view that oculomotor dominance cannot explain the pattern of right and left eye micro-movements in Experiment 2 is also consistent with Liversedge et al.’s (2006) findings according to which oculomotor dominance, measured with a porta-type test, does not influence intra-fixation movements of the right and left eye in relation to each other.
The larger right-directed movement bias when alternating case text was read than when lower case text was read also implies an influence of processing demands on intra-fixation movement dynamics. Alternating case forms two distinct letter groups each of which receives some processing (Humphreys et al., 2003; Mayall et al., 1997). The extraction of linguistic information from words written in alternating case therefore requires integration of the two letter groups, and this integration must be principled, i.e., it must follow letter order. Integration could, therefore, progress with letter order from left to right during reading fixations, and it is plausible to assume that it biased micro-movements of the right and left eye in a corresponding direction.
The effects of alternating case and word length differed, although both have been shown to influence the ease of visual word identification. Alternating case changes a word’s visual configuration but not its linguistic content. Changes in the length of a word, by contrast, are intertwined with changes in linguistic structure. Alternating case and word length are, therefore, likely to engage functionally distinct word recognition processes: an added-on process that involves the integration of letter groups into words and the representation of the linguistic content of the full word, respectively. Only one of these processes, that ordered the elements of visually distinct subword units, appears to influence intra-fixation movements.
The absence of a word length effect on intra-fixation location changes rules out the possibility that the mere duration of a fixation influenced their spatially selectivity. Fixation durations in Experiment 1 and Experiment 2 were relatively short when words with three or four constituent letters were fixated. Yet, the viewing of these words was not accompanied by a diminished right-directed movement bias. Similarly, saccade size per se did not influence intra-fixation movements in Experiment 2. Although incoming saccades were smaller in Experiment 2 than in Experiment 1, it is unlikely that intra-fixation movements sought to extend the relatively short saccades of Experiment 2. Right directed saccades in Experiment 2 were often followed by left-directed intra-fixation movements, and the correlation between the size of incoming saccades and the size of intra-fixation location changes was small and not reliable.
General Discussion
The current investigation pursued two major goals: to establish normative data on the frequency, size, and direction of intra-fixation movements of each eye during reading and to determine whether these location changes are subject to processing demands. With one striking exception, the frequency distributions of intra-fixation location changes were normally distributed with a mean value of .03 LS for the right eye in Experiment 1 and with mean values of .11 LS and .27 LS for the right and left eyes, respectively, in Experiment 2. In both Experiments, negligible movement sizes between −.05 and +.05 LS were relatively common and did not fit the normal distribution. This suggests that there was an underlying tendency to keep each eye at a ‘fixed’ location. The experiments also revealed that micro-movements during reading fixations are not a function of the direction and size of in- and outgoing saccades, that they are similar under monocular and binocular viewing conditions, and that they are not influenced by the success with which a lexical representation can be accessed.
Nevertheless, micro-movements during reading fixations were not directionally neutral. One key finding was a distinctly larger spatial bias toward the right when alternating case text was read than when lower case was read, as the ordered integration of two alternating letter groups pulled the eyes in a congruent direction. The robust effect of word order on intra-fixation location changes of the right and left eye provide further evidence for the influence of text processing on intra-fixation movement dynamics.
Right-directed location changes decreased in size and frequency as the right eye moved from left-to-right through a sentence. The right-directed movement size was .24 LS for the second word location which decreased to .05 LS for the tenth word location. The left eye revealed a similar but much smaller effect of word order, except for a somewhat anomalous reversal of the word location effect for locations two and three. When word two was excluded, the size of right-directed intra-fixation location changes decreased from .32 LS during the viewing of third word of the sentence to .23 LS during the viewing of the tenth word. As argued before, this pattern of word order effects for the right and left eye rules out the possibility that the two eyes were pulled toward either the screen center or toward the head midline, and it rules out the possibility that intra-fixation location changes are spatially biased because a line of gaze that crosses the head midline is pulled toward the midline.
The systematic effect of word order on intra-fixation movements of the right eye implies that the spatial dynamics of micro-movements were open to graded input that changed as sentence reading progressed from left to right. As the eyes moved along a line of text, fewer to-be-identified (to-be-attended) words remained visible in the right visual field where they occupied a progressively smaller spatial area. The two eyes may have been pulled toward the area of yet to be read text, and the strength of this pull decreased as the eyes approached the sentence ending. This account does not explain, however, why word location influenced right eye intra-fixation movements to a much larger extent than left eye movements.
The larger effect of word order on micro-movements of the right than left eye should have had some effect on movements of the two eyes in relation to each other. Since both eyes were more likely to move to a larger extent in the same direction when a fixated word was near the beginning of a line of text than when it was near the ending, convergence of the two eyes may be less successful at the onset of sentence reading than toward the end. The testing of this hypothesis, which requires measurement of binocular disparity as a function of word order, is currently in progress.
Acknowledgements
This work was supported by NIH grant HD043405. We thank Simon Liversedge, Marc Brysbaert, Seth Greenberg, and Ulrich Weger for helpful comments on an earlier version of this manuscript. Inquiries should be sent to Albrecht Inhoff (Inhoff@binghamton.edu).
Footnotes
Monocular calibrations are likely to yield more precise estimates of each eye’s fixation location (Liversedge et al., 2006).
To check this assumption, intra-fixation movements were also analyzed separately for words that had been masked before they were directly fixated. The distribution of intra-fixation location changes during the fixation of these words was virtually identical to the distribution of shifts for other words in the data base.
Motor dominance and direction dominance are statistically independent (Porac & Coren, 1976).
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Blythe H, Liversedge SP, Joseph HS, White SJ, Findlay JM, Rayner K. The binocular coordination of eye movements during reading in children and adults. Vision Research. 2006;46:3898–3908. doi: 10.1016/j.visres.2006.06.006. [DOI] [PubMed] [Google Scholar]
- Brysbaert M, Vitu F. Word skipping: Implications for theories of eye Movement control in reading. In: Underwood G, editor. Eye guidance in reading and scene perception. Oxford, UK: Elsevier Publishers; 1998. pp. 125–148. [Google Scholar]
- Ditchburn RW. Eye-movements in relation to retinal action. Optica Acta. 1955;1:171–176. [Google Scholar]
- Ditchburn RW, Fender DH. The stabilized retinal image. Optica Acta. 1955;3:128–133. doi: 10.1080/713817821. [DOI] [PubMed] [Google Scholar]
- Engbert R, Kliegl R. Microsaccades keep the eyes’ balance during fixation. Psychological Science. 2004;15:431–436. doi: 10.1111/j.0956-7976.2004.00697.x. [DOI] [PubMed] [Google Scholar]
- Heller D, Radach R. Eye movements in reading: Are two eyes better than one. In: Becker W, Deubel H, Mergner T, editors. Current oculomotor research: Physiologial and psychological aspects. New York: Luker Academic Plenum; 1999. pp. 341–348. [Google Scholar]
- Hendricks A. Vergence movements during fixations in reading. Acta Psychologica. 1996;92:131–151. doi: 10.1016/0001-6918(95)00011-9. [DOI] [PubMed] [Google Scholar]
- Humphreys GW, Mayall K, Cooper ACG. The PIG in sPrInG: Evidence on letter grouping from the reading of buried words. Psychonomic Bulletin & Review. 2003;10:939–946. doi: 10.3758/bf03196555. [DOI] [PubMed] [Google Scholar]
- Inhoff AW, Eiter B, Radach R. Time course of linguistic information extraction from consecutive words during eye fixations in reading. Journal of Experimental Psychology: Human Perception and Performance. 2005;31:979–995. doi: 10.1037/0096-1523.31.5.979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inhoff AW, Topolski R. Seeing morphemes: Loss of visibility during the retinal stabilization of compoiund and pseudocompound words. Journal of Experimental Psychology: Human Perception and Performance. 1994;20:840–853. doi: 10.1037//0096-1523.20.4.840. [DOI] [PubMed] [Google Scholar]
- Inhoff AW, Topolski R. Loss of vision during the retinal stabilization of letters. Psychological Research. 1995;58:155–162. doi: 10.1007/BF00419630. [DOI] [PubMed] [Google Scholar]
- Juhasz B, Liversedge SP, White SJ, Rayner K. Binocrular coordination of the eyes during reading: Word frequency and case alternation affect fixation duration but not fixation disparity. The Quarterly Journal of Experimental Psychology. 2006;59:1614–1625. doi: 10.1080/17470210500497722. [DOI] [PubMed] [Google Scholar]
- Liversedge SP, White SJ, Findlay JM, Rayner K. Binocular coordination of eye movements during reading. Vision Research. 2006;46:2363–2374. doi: 10.1016/j.visres.2006.01.013. [DOI] [PubMed] [Google Scholar]
- Martinez-Conde S, Macknik SL, Hubel DH. The role of fixational eye movements in visual perception. Nature Reviews Neuroscience. 2004;5:229–240. doi: 10.1038/nrn1348. [DOI] [PubMed] [Google Scholar]
- Mayall KA, Hymphreys GW, Olson A. Disruption to word and letter processing? The origins of case-mixing effects. Journal of Experimental Psychology: Human Perception and Performance. 1997;13:14–24. doi: 10.1037//0278-7393.23.5.1275. [DOI] [PubMed] [Google Scholar]
- New B, Ferrand L, Pallier C, Brysbaert M. Reexamining the word length effect in visual word recognition: New evidence from the English lexicon project. Psychonomic Bulletin & Review. 2006;13:45–52. doi: 10.3758/bf03193811. [DOI] [PubMed] [Google Scholar]
- Porac C, Coren S. The dominant eye. Psychological Bulletin. 1976;83:880–897. [PubMed] [Google Scholar]
- Radach R, Heller D, Jashinski P. Binocular coordination, fixation disparity and ocular dominance. Perception. 1999;25 Supplement:67. [Google Scholar]
- Rayner K. Eye movements in reading and information processing: 20 years of research. Psychological Bulletin. 1998;124:372–422. doi: 10.1037/0033-2909.124.3.372. [DOI] [PubMed] [Google Scholar]
- Riggs LA, Ratliff F, Cornsweet JC, cornsweet TN. The disappearance of steadily fixated visual test objects. Journal of the Optical Society of America. 1953;43:495–501. doi: 10.1364/josa.43.000495. [DOI] [PubMed] [Google Scholar]
- Vitu F, O'Regan K, Inhoff AW, Topolski R. Mindless reading: Eye movement characteristics are similar in scanning strings and reading text. Perception & Psychophysics. 1995;57:352–364. doi: 10.3758/bf03213060. [DOI] [PubMed] [Google Scholar]