

Abstract
One major contribution of neuroscience to understanding cognitive development has been in demonstrating that biology is not destiny—that is, demonstrating the remarkable role of experience in shaping the mind, brain, and body. Only rarely has neuroscience provided wholly new insights into cognitive development, but often it has provided evidence of mechanisms by which observations of developmental psychologists could be explained. Behavioral findings have often remained controversial until an underlying biological mechanism for them was offered. Neuroscience has demonstrated promise for detecting cognitive problems before they are behaviorally observable—and, hence, promise for early intervention. In this article, we discuss examples drawn from imitation and mirror neurons, phenylketonuria (PKU) and prefrontal dopamine, maternal touch and stress reactivity, and non-genetic (behavioral) intergenerational transmission of biological characteristics.
Keywords: plasticity, epigenesis, mothering, executive functions, animal models, molecular genetics, memory
Neuroscience research has made its greatest contributions to the study of cognitive development by illuminating mechanisms (providing a “how”) that underlie behavioral observations made earlier by psychologists. It has also made important contributions to our understanding of cognitive development by demonstrating that the brain is far more plastic at all ages than previously thought—and thus that the speed and extent by which experience and behavior can shape the brain is greater than almost anyone imagined. In other words, rather than showing that biology is destiny, neuroscience research has been at the forefront of demonstrating the powerful role of experience throughout life. Besides the surprising evidence of the remarkable extent of experience-induced plasticity, rarely has neuroscience given us previously unknown insights into cognitive development, but neuroscience does offer promise of being able to detect some problems before they are behaviorally observable.
PROVIDING MECHANISMS THAT CAN ACCOUNT FOR BEHAVIORAL RESULTS REPORTED BY PSYCHOLOGISTS
Here we describe two examples of behavioral findings by psychologists that were largely ignored or extremely controversial until underlying biological mechanisms capable of accounting for them were provided by neuroscience research. One such example concerns cognitive deficits documented in children treated early and continuously for phenylketonuria (PKU). The second example involves neonatal imitation observed by psychologists and mirror neurons discovered by neuroscientists.
Prefrontal Dopamine System and PKU Cognitive Deficits
Since at least the mid-1980s, psychologists were reporting cognitive deficits in children with PKU that resembled those associated with frontal cortex dysfunction (e.g., Pennington, VanDoornick, McCabe, & McCabe, 1985). Those reports did not impact medical care, however. Doctors were skeptical. No one could imagine a mechanism capable of producing what psychologists claimed to be observing.
PKU is a disorder in the gene that codes for phenylalanine hydroxylase, an enzyme essential for the conversion of phenyl-alanine (Phe) to tyrosine (Tyr). In those with PKU, that enzyme is absent or inactive. Without treatment, Phe levels skyrocket, resulting in gross brain damage and mental retardation. Phe is an amino acid and a component of all dietary protein. PKU treatment consists primarily of reducing dietary intake of protein to keep Phe levels down, but that has to be balanced against the need for protein. For years, children with PKU were considered adequately treated if their blood Phe levels were below 600 micromoles per liter (μmol/L; normal levels in the general public being 60–120 μmol/L). Such children did not have mental retardation and showed no gross brain damage, although no one disputed that their blood Phe levels were somewhat elevated and their blood Tyr levels were somewhat reduced (Tyr levels were not grossly reduced because even though the hydroxylation of Phe into Tyr was largely inoperative, Tyr is also available in protein). Since Phe and Tyr compete to cross into the brain, a modest increase in the ratio of Phe to Tyr in the bloodstream results in a modest decrease in how much Tyr can reach the brain. Note that this is a global effect—the entire brain receives somewhat too little Tyr. How was it possible to make sense of psychologists’ claims that the resulting cognitive deficits were not global but limited to the cognitive functions dependent on prefrontal cortex?
Neuroscience provided a mechanism by which psychologists’ findings made sense. Research in neuropharmacology had shown that the dopamine system in prefrontal cortex has unusual properties not shared by the dopamine systems in other brain regions such as the striatum. The dopamine neurons that project to pre-frontal cortex have higher rates of firing and dopamine turnover. This makes prefrontal cortex sensitive to modest reductions in Tyr (the precursor of dopamine) that are too small to affect the rest of the brain (Tam, Elsworth, Bradberry, & Roth, 1990). Those unusual properties of the prefrontal dopamine system provide a mechanism by which children treated for PKU could show selective deficits limited to prefrontal cortex. The moderate imbalance in the bloodstream between Phe and Tyr causes a reduction in the amount of Tyr reaching the brain that is large enough to impair the functioning of the prefrontal dopamine system but not large enough to affect the rest of the brain. Diamond and colleagues provided evidence for this mechanism in animal models of PKU and longitudinal study of children (Diamond, 2001). That work, presenting a mechanistic explanation and providing convincing evidence to support it, resulted in a change in the medical guidelines for the treatment of PKU (blood Phe levels should be kept between 120 and 360 μmol/L) that has improved children’s lives (e.g., Stemerdink et al., 2000). Also, by shedding light on the role of dopamine in the prefrontal cortex early in development, such work offers insights on the development of cognitive control (executive function) abilities that are relevant to all children.
Mirror Neurons and Neonate Imitation
In 1977, Meltzoff and Moore created a sensation by reporting that human infants just 12 to 21 days old imitated facial expressions they observed adults making (see Fig. 1). That was followed by a second demonstration of such imitation in infants as young as 42 minutes (Meltzoff & Moore, 1983). For years, those reports met strong resistance. Such imitation was thought to be far too sophisticated an accomplishment for a neonate. After all, infants can feel but not see their own mouth and tongue movements, and they can see but not feel the mouth and tongue movements of others. To equate their own motor movements with the perception of those same movements by others would seem to involve high-level cross-modal matching.
Fig. 1.
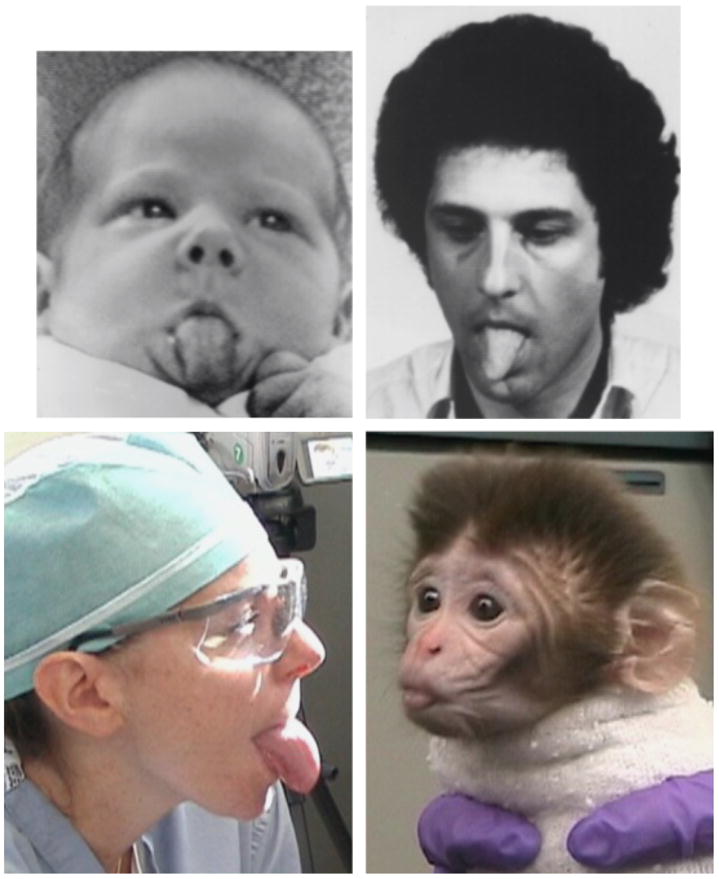
Imitation of a human adult’s tongue protrusion by a neonatal human and a neonatal rhesus macaque. Top pictures are reprinted from “Imitation of Facial and Manual Gestures by Human Neonates,” by A.N. Meltzoff and M.K. Moore, 1977, Science, 198, p. 75. Copyright 1977, American Association for the Advancement of Science. Reproduced with permission. Bottom photos are from the study reported in Ferrari et al. (2006). They are reproduced here with the permission of Annika Paukner and Steve Suomi.
The discovery of mirror neurons by Rizzolatti and his colleagues, Fadiga, Fogassi, and Gallese (for review, see Rizzolatti & Craighero, 2004) provided a mechanism that could conceivably underlie newborns’ ability to show such imitation rather automatically. Mirror neurons fire when an individual executes an action or when an individual observes someone else executing that action. The cross-modal association occurs at the neuronal, single-cell level. It has since been demonstrated that 3-day-old rhesus monkeys also imitate the facial movements of adult humans (Ferrari et al., 2006; see Fig. 1) and that the close link between perception and action is not limited to vision; hearing a sound associated with an action activates mirror neurons associated with that action just as does the sight of that action (Kohler et al., 2002).
Whereas the preceding examples are of neuroscience elucidating possible neurobiological bases for observed psychological phenomena, we move on to describe phenomena—concerning plasticity and environmental influences—that neuroscientists have brought to the attention of developmentalists.
POWERFUL EFFECTS OF EARLY EXPERIENCE ON BRAIN, BODY, MIND, BEHAVIOR, AND GENE EXPRESSION
Ironically, one of the most important findings to emerge from neurobiology is that biology is not destiny. Neuroscience research has shown that experience plays a far larger role in shaping the mind, brain, and even gene expression than was ever imagined. This insight is particularly important in advancing theory in cognitive development, where debates have raged about the importance of nature versus nurture.
Examples of striking experience-induced plasticity abound—for example, the groundbreaking work of Greenough, Merzenich, Maurer, Neville, Pascual-Leone, Taub, Sur, and Kral. Here we highlight work by Schanberg and Meaney, in part because that work emphasizes a sensory system that has received far less attention by psychologists than have vision and audition: the sense of touch.
Nurturing Touch and its Importance for Growth
Two independent, elegant lines of work have demonstrated the powerful effects of touch. Schanberg and colleagues have shown that the licking behavior of rat mothers is essential for the growth of rat pups. If rat pups are deprived of this touch for even just 1 hour, DNA synthesis is reduced, growth-hormone secretion is inhibited, and bodily organs lose their capacity to respond to exogenously administered growth hormone (Butler, Suskind, & Schanberg, 1978; Kuhn, Butler, & Schanberg, 1978). Schanberg and colleagues have identified molecular mechanisms through which deprivation of the very specific kind of touch rat mothers administer to their pups produces these effects (e.g., Schanberg, Ingledue, Lee, Hannun, & Bartolome, 2003).
Nurturing Touch and its Importance for Reducing Stress Reactivity and for Cognitive Development
Meaney and colleagues have demonstrated that rat moms who more frequently lick and groom their pups produce offspring who, throughout their lives, explore more, are less fearful, show milder reactions to stress, perform better cognitively as adults, and preserve their cognitive skills better into old age (Liu, Diorio, Day, Francis, & Meaney, 2000). It is the mother’s behavior that produces these effects rather than a particular genetic profile that produces both a particular mothering style and particular offspring characteristics. Pups of high-licking-and-grooming moms raised by low-licking-and-grooming moms do not show these characteristics, and pups of low-touch moms raised by high-touch moms do show this constellation of attributes (Francis, Diorio, Liu, & Meaney, 1999).
Furthermore, rats tend to raise their offspring the way they themselves were raised, so these effects are transmitted inter-generationally, not through the genome but through behavior. Biological offspring of low-touch moms who are cross-fostered to high-touch moms lick and groom their offspring a lot; in this way the diminished stress response and cognitive enhancement is passed down through the generations (Francis et al., 1999).
Meaney and colleagues have elegantly demonstrated that maternal behavior produces these behavioral consequences through several mechanisms that alter gene expression. Not all genes in an individual are expressed—many are never expressed. Experience can affect which genes are turned on and off, in which cells, and when. For example, methylation (attaching a methyl group to a gene’s promoter) stably silences a gene; demethylation reverses that process, typically leading to the gene being expressed. High licking by rat mothers causes demethylation (i.e., activation) of the glucocorticoid receptor gene, hence lowering circulating glucocorticoid (stress hormone) levels as receptors for the stress hormone remove it from circulation.
Nurturing Touch and Human Cognitive and Emotional Development
Unlike newborn rats, human newborns can see, hear, and smell, as well as feel touch. Yet despite the additional sensory information available to them, touch is still crucial. Human infants who receive little touching grow more slowly, release less growth hormone, and are less responsive to growth hormone that is exogenously administered (Frasier & Rallison, 1972). Throughout life, they show larger reactions to stress, are more prone to depression, and are vulnerable to deficits in cognitive functions commonly seen in depression or during stress (Lupien, King, Meaney, McEwen, 2000).
Touch plays a powerful role for human infants in promoting optimal development and in counteracting stressors. Massaging babies lowers their cortisol levels and helps them gain weight (Field et al., 2004). The improved weight gain from neonatal massage has been replicated cross-culturally, and cognitive benefits are evident even a year later. It is not that infants sleep or eat more; rather, stimulating their body through massage increases vagal (parasympathetic nervous system) activity, which prompts release of food-absorption hormones. Such improved vagal tone also indicates better ability to modulate arousal and to attend to subtle environmental cues important for cognitive development. Passive bodily contact also has substantial stress-reducing, calming, and analgesic effects for infants and adults (e.g., Gray, Watt, & Blass, 2000; see Fig. 2). Thus, besides “simple touch” being able to calm our jitters and lift our spirits, the right kind of touch regularly enough early in life can improve cognitive development, brain development, bodily health throughout life, and gene expression.
Fig. 2.
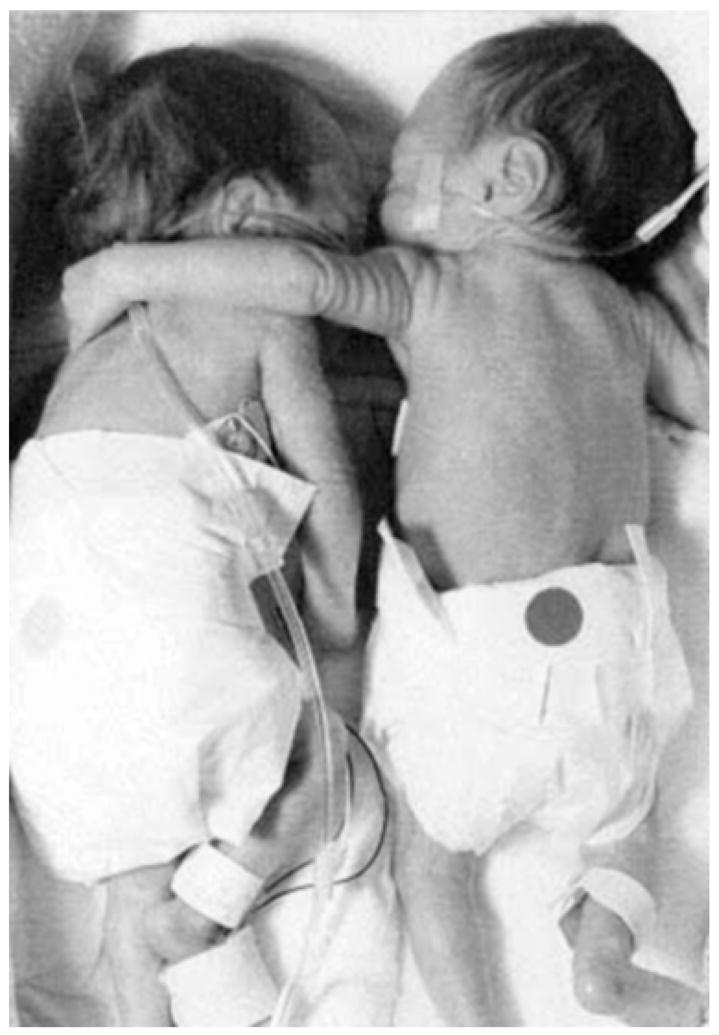
Double-bedded premature twins. Born 12 weeks early, these twins were initially whisked into separate incubators. Kyrie (on the right), the larger by over 2 pounds, slept peacefully, but Brielle (on the left) had breathing and heart-rate problems, didn’t gain weight, and fussed when anyone tried to comfort her. Finally a nurse, acting counter to hospital regulations, put the two sisters together. As Brielle dozed, Kyrie put her arm around her smaller sibling. Brielle began to thrive. Sooner than expected, the girls went home. Today a handful of institutions use double bedding, which reduces the number of hospital days.
FUTURE DIRECTIONS
Neuroscience may be able to make extremely important contributions to child development by building on repeated demonstrations that differences in neural activity patterns precede and predict differences in cognitive performance. Often, when the brain is not functioning properly, people can compensate so their performance does not suffer until the neural system becomes too dysfunctional or until performance demands become too great. Thus, an underlying problem may exist but not show up behaviorally until, for example, the academic demands of more advanced schooling exceed a child’s ability to compensate.
So far, differences in neural activity patterns have been demonstrated to precede and predict differences in cognitive performance only in adults. For example, Bookheimer and colleagues tested older adults (ranging in age from 47 to 82 years) with a genetic predisposition for Alzheimer’s disease, selected because they performed fully comparably to controls across diverse cognitive tasks. Nevertheless, functional neuroimaging revealed that the brains of several of the genetically predisposed individuals already showed predicted differences. Two years later, those individuals showed the cognitive impairments predicted by their earlier neural-activity patterns (Bookheimer et al., 2000). Similarly, adults in the early stages of other disorders may show no behavioral evidence of a cognitive deficit while neuroimaging shows their brains are compensating or working harder to achieve that behavioral equivalence. As the disease progresses, the compensation is no longer sufficient and the cognitive deficit becomes evident (e.g., Audoin et al., 2006).
What this suggests is that functional neuroimaging in developing children may perhaps be able to detect evidence of learning disorders—such as attentional, sensory-processing, language, or math deficits—before there is behavioral evidence of a problem. Already, research is being undertaken to see if infants’ neural responses to auditory stimuli might be predictive of later linguistic problems (e.g., Benasich et al., 2006). The earlier a problem can be detected, the better the hope of correcting it or of putting environmental compensations in place.
Recommended Reading
Diamond, A. (2001). (See References). Summarizes studies with young children and animals showing the role of maturation of prefrontal cortex in the early emergence of executive function abilities and the importance of dopamine for this.
Grossman, A.W., Churchill, J.D., Bates, K.E., Kleim, J.A., & Greenough, W.T. (2002). A brain adaptation view of plasticity: Is synaptic plasticity an overly limited concept? Progress in Brain Research, 138, 91–108. Argues that synaptic, even neuronal, plasticity is but a small fraction of the range of brain changes that occur in response to experience, and that there are multiple forms of brain plasticity governed by mechanisms that are at least partially independent, including non-neuronal changes.
Meaney, M.J. (2001). Maternal care, gene expression, and the transmission of individual differences in stress reactivity across generations. Annual Review of Neuroscience, 24, 1161–1192. Provides an overview of research demonstrating that naturally occurring variations in maternal care modify the expression of genes affecting offspring’s cognitive development as well as their ability to cope with stress throughout life, and that these changes are passed down intergenerationally (epigenetic inheritance).
Meltzoff, A.N., & Decety, J. (2003). What imitation tells us about social cognition: A rapprochement between developmental psychology and cognitive neuroscience. Philosophical Transactions of the Royal Society of London – B: Biological Sciences, 358, 491–500. Reviews the psychological evidence concerning imitation in human neonates and the neurophysiological evidence of a common coding at the single cell level (in mirror neurons) between perceived and generated actions.
Neville, H.J., & Bavelier, D. (2002). Human brain plasticity: Evidence from sensory deprivation and altered language experience. Progress in Brain Research, 138, 177–188. Summarizes research, using behavioral measures and neuroimaging, on individuals with altered visual, auditory, and/or language experience, showing ways in which brain development can, and cannot, be modified by environmental input, and how that varies by the timing of the altered input and by specific subfunctions within language or vision.
Acknowledgments
AD gratefully acknowledges grant support from the National Institute on Drug Abuse (R01 #DA19685) during the writing of this paper.
References
- Audoin B, Au Duong MV, Malikova I, Confort-Gouny S, Ibarrola D, Cozzone PJ, et al. Functional magnetic resonance imaging and cognition at the very early stage of MS. Journal of the Neurological Sciences. 2006;245:87–91. doi: 10.1016/j.jns.2005.08.026. [DOI] [PubMed] [Google Scholar]
- Benasich AA, Choudhury N, Friedman JT, Realpe Bonilla T, Chojnowska C, Gou Z. Infants as a prelinguistic model for language learning impairments: Predicting from event-related potentials to behavior. Neuropsychologia. 2006;44:396–441. doi: 10.1016/j.neuropsychologia.2005.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bookheimer SY, Strojwas MH, Cohen MS, Saunders AM, Pericak-Vance MA, Mazziota JC, et al. Patterns of brain activation in people at risk for Alzheimer’s disease. New England Journal of Medicine. 2000;343:450–456. doi: 10.1056/NEJM200008173430701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Butler SR, Suskind MR, Schanberg SM. Maternal behavior as a regulator of polyamine biosynthesis in brain and heart of the developing rat pup. Science. 1978;199:445–447. doi: 10.1126/science.202031. [DOI] [PubMed] [Google Scholar]
- Diamond A. A model system for studying the role of dopamine in prefrontal cortex during early development in humans. In: Nelson C, Luciana M, editors. Handbook of developmental cognitive neuroscience. Cambridge, MA: MIT Press; 2001. pp. 433–472. [Google Scholar]
- Field T, Hernandez-Reif M, Diego M, Feijo L, Vera Y, Gil K. Massage therapy by parents improves early growth and development. Infant Behavior & Development. 2004;27:435–442. [Google Scholar]
- Ferrari PF, Visalberghi E, Paukner A, Fogassi L, Ruggiero A, Suomi S. Neonatal imitation in rhesus macaques. PLoS Biology. 2006;4:1501–1508. doi: 10.1371/journal.pbio.0040302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Francis D, Diorio J, Liu D, Meaney MJ. Nongenomic transmission across generations of maternal behavior and stress responses in the rat. Science. 1999;286:1155–1158. doi: 10.1126/science.286.5442.1155. [DOI] [PubMed] [Google Scholar]
- Frasier SD, Rallison ML. Growth retardation and emotional deprivation: Relative resistance to treatment with human growth hormone. Journal of Pediatrics. 1972;80:603–609. doi: 10.1016/s0022-3476(72)80057-x. [DOI] [PubMed] [Google Scholar]
- Gray L, Watt L, Blass EM. Skin-to-skin contact is analgesic in healthy newborns. Pediatrics. 2000;105:1–6. doi: 10.1542/peds.105.1.e14. [DOI] [PubMed] [Google Scholar]
- Kohler E, Keysers C, Umiltà MA, Fogassi L, Gallese V, Rizzolatti G. Hearing sounds, understanding actions: Action representation in mirror neurons. Science. 2002;297:846–848. doi: 10.1126/science.1070311. [DOI] [PubMed] [Google Scholar]
- Kuhn CM, Butler SR, Schanberg SM. Selective depression of serum growth hormone during maternal deprivation in rat pups. Science. 1978;201:1034–1036. doi: 10.1126/science.684424. [DOI] [PubMed] [Google Scholar]
- Liu D, Diorio J, Day JC, Francis DD, Meaney MJ. Maternal care, hippocampal synaptogenesis and cognitive development in rats. Nature Neuroscience. 2000;3:799–806. doi: 10.1038/77702. [DOI] [PubMed] [Google Scholar]
- Lupien SJ, King S, Meaney MJ, McEwen BS. Child’s stress hormone levels correlate with mother’s socioeconomic status and depressive state. Biological Psychiatry. 2000;48:976–980. doi: 10.1016/s0006-3223(00)00965-3. [DOI] [PubMed] [Google Scholar]
- Meltzoff AN, Moore MK. Imitation of facial and manual gestures by human neonates. Science. 1977;198:75–78. doi: 10.1126/science.198.4312.75. [DOI] [PubMed] [Google Scholar]
- Meltzoff AN, Moore MK. Newborn infants imitate adult facial gestures. Child Development. 1983;54:702–709. [PubMed] [Google Scholar]
- Pennington BF, VanDoornick WJ, McCabe LL, McCabe ERB. Neuropsychological deficits in early treated phenylketonuric children. American Journal of Mental Deficiency. 1985;89:467–474. [PubMed] [Google Scholar]
- Rizzolatti G, Craighero L. The mirror-neuron system. Annual Review of Neuroscience. 2004;27:169–192. doi: 10.1146/annurev.neuro.27.070203.144230. [DOI] [PubMed] [Google Scholar]
- Schanberg SM, Ingledue VF, Lee JY, Hannun YA, Bartolome JV. PKC mediates maternal touch regulation of growth-related gene expression in infant rats. Neuropsychopharmacology. 2003;28:1026–1030. doi: 10.1038/sj.npp.1300125. [DOI] [PubMed] [Google Scholar]
- Stemerdink BA, Kalverboer AF, van der Meere JJ, van der Molen MW, Huisman J, de Jong LW, et al. Behaviour and school achievement in patients with early and continuously treated phenylketonuria. Journal of Inherited Metabolic Disorders. 2000;23:548–562. doi: 10.1023/a:1005669610722. [DOI] [PubMed] [Google Scholar]
- Tam SY, Elsworth JD, Bradberry CW, Roth RH. Mesocortical dopamine neurons: High basal firing frequency predicts tyrosine dependence of dopamine synthesis. Journal of Neural Transmission. 1990;81:97–110. doi: 10.1007/BF01245830. [DOI] [PubMed] [Google Scholar]