

Abstract
Background
Human infections with non-O1, non-O139 V. cholerae have been described from Laos. Elsewhere, non cholera-toxin producing, non-O1, non-O139 V. cholerae have been described from blood cultures and ascitic fluid, although they are exceedingly rare isolates.
Case presentation
We describe a farmer who died with Vibrio cholerae O21 bacteremia and peritonitis in Vientiane, Laos, after eating partially cooked apple snails (Pomacea canaliculata) and mussels (Ligumia species). The cultured V. cholerae were non-motile. PCR detected ompW and toxR gene regions but not the ctxA, ompU, omp K and TCP gene regions. Although the organisms lacked flagellae on scanning electron microscopy, they possessed the Vibrio flagellin flaA gene.
Conclusion
Severe bacteremic non-O1, non-O139 V. cholerae is reported from Laos. The organisms were unusual in being non-motile. They possessed the Vibrio flagellin flaA gene. Further research to determine the reasons for the non-motility and virulence is required.
Background
Cholera is endemic in Laos and O1 and non-O1 V. cholerae (including serogroups 11, 14, 16, 21, 41, 43, 68 and 169), but not 0139, have been recorded [[1,2], Phouthavane et al. unpublished]. Non cholera-toxin producing, non-O1, non-O139 V. cholerae have been described from blood cultures and ascitic fluid, although they are exceedingly rare isolates. Consumption of undercooked shellfish, exposure of open wounds to salt and fresh water, drinking of well water, ascites, cirrhosis, renal failure and haematological malignancies have all been associated with non-O1, non-O139 disease [3-5]. The mortality of recorded non-O1 bacteraemia is high at 24–62% [6-11]. Concurrent infection with organism(s) that may damage the bowel mucosa may predispose patients to V. cholerae non-O1 bacteraemia [12]. Of 28 patients with non-01 V. cholerae bacteremia recorded in Taiwan over 5 years, 95% had ascites and 75% had cirrhosis, perhaps because increased intestinal permeability in cirrhotic patients predisposes them to invasive disease [13]. Unlike O1 biotype El Tor, but like O139 V. cholerae, O21 possess a capsule, which is thought to facilitate survival in the blood stream [12]. V. cholerae 01 bacteremia has also been described, but is more rare than non-01 [14].
Case Presentation
In January 2006, a 20 year old female rice farmer, from Nayxaythong District, on the outskirts of Vientiane City, was admitted to Mahosot Hospital, Vientiane, Lao PDR (Laos) with a 7 day history of abdominal pain, fever, rigors, watery diarrhoea, yellow eyes, leg oedema, anuria and malaena. She had collected many apple snails 'hoy pak kwang' (Pomacea canaliculata) and mussels 'hoy kee' (Ligumia species) from the mud on the edge of a nearby lake and washed them before boiling them for 30 minutes. She ate partially cooked snails as she boiled them and became unwell 7 days later. The fully cooked snails were eaten, with chilli sauce, by another villager who remained well. On admission she was had a fever (39.5°C) with jaundice, abdominal tenderness, splenomegaly, tachycardia, shortness of breath, lower leg oedema and ascites. She had no skin lesions and had no evidence for prior cirrhosis, antacid or steroid use, gastric surgery or haemoglobinopathy. Her haematocrit was 35%, with a peripheral white cell count of 8.9 109/L (66% neutrophils). Serum creatinine, alkaline phosphatase and glucose were within the reference ranges, with raised total serum bilirubin 41 μmol/L (reference <14.5 μmol/L), AST 75 IU/L (<37 IU/L) and ALT 170 IU/L (<40 IU/L). Abdominal ultrasound demonstrated ascites and splenomegaly with normal liver and kidneys. Stool was not cultured. She was treated empirically with intravenous ampicillin, gentamicin and diuretics. Analysis of tapped ascitic fluid revealed many white cells, 90% of which were neutrophils.
Oxidase positive, curved, non-motile Gram negative rods were grown from 2 of 2 blood cultures, after 2 days incubation and the ascitic fluid, after 1 day of incubation. API 20E (bioMeriuex, France) identified Vibrio cholerae with 99.9% confidence. The organism grew in 0% sodium chloride but not in 6% or 10% sodium chloride and did not agglutinate when tested against O1 and O139 antisera, but did against O21 antisera (Department of Bacteriology, National Institute of Infectious Disease in Tokyo, Japan). By disc diffusion testing (Kerby-Bauer, according to NCCLS guidelines 2003) the organisms were susceptible to ampicillin, ciprofloxacin, ceftriaxone and tetracycline. She was taken home moribund, at her relatives' request, after 2 days in hospital and died soon afterwards.
In order to investigate whether we could grow 021 V. cholerae from the probable source of infection, in May 2006 specimens of P. canaliculata and Ligumi species were collected from the same lake as above and cultured in TCBS according to the methods of Ottaviani et al. [15]. No V. cholerae were grown but Aeromonas, Enterobacter, Proteus and Escherichia species were identified.
The PCR techniques of Nandi et al. [16] were used to confirm that the organisms contained the outer membrane protein W (ompW) and regulatory protein (toxR) of Vibrio cholerae and to determine whether they contained cholera toxin (ctxA) and toxin-coregulated pilus (TCP) virulence genes (Table 1 &2). The bacteria from blood culture and ascitic fluid were re-plated on Modified Drigalski Agar (BTB agar, Eiken Kagaku Co Ltd, Tokyo, Japan) to obtain pure growth and DNA extraction was performed (Wizard, A1120, Promega, WI, USA). Biotype Classic O1 Inaba strain 569B and Vibrio cholerae O21, recovered from a patient with diarrhoea in Vientiane Capital, were used as positive control and reference strains, respectively and compared with human isolates of serogroup O21 109-68 from India (from diarrhoeal stool), and environmental isolate 418-03 from the USA (seawater, Chesapeake Bay, Virginia) (both from National Institute of Infectious Diseases (NIID) Tokyo). Primers of Outer membrane protein W (ompW) were prepared according to [16] and outer membrane protein U (ompU), outer membrane protein K (ompK), toxin-coregulated pilus (TCP), cholerae toxin A (ctxA) and Regulatory protein (toxR) primers were designed (Table 2) and DNA amplification performed (i-cycler, BioRad) with analysis using 1.8% agarose gel electrophoresis. V. cholerae O21 cultures from both blood and ascitic fluid from our patient and 109-68 and 418-03 had ompW and toxR gene regions, confirming them to be V. cholerae [16], but had no ctxA, ompU, omp K, TCP gene regions, which were present in the Classic 569B strain (Figure 1, Table 1). Scanning electron microscopy (SEM; S-5200, Hitachi, Japan) of the patient's isolate, serogroup O21 109-68 and 418-03 (Figures 2, 3, 4) demonstrated that, although 109-68 and 418-03 did possess flagellae, the organisms isolated from the patient did not. The flagellin regulation gene flaA was demonstrated by PCR in the patient's isolates and 569B with specific oligonucleotide primers (Table 3), which we designed based on the work of Klose and Mekalanos [17] who demonstrated that immotility could be caused by mutations in the flaA gene alone.
Table 1.
Summary of the gene regions detected in the patients isolates from ascitic fluid and blood culture compared to the 569B reference organism and serogroup O21 609-68 and 418-03 strains kindly provided by the National Institute of Infectious Diseases, Tokyo, Japan.
| Gene | Primer | Product length/bp | Inaba 569B | 609-68 India | 418-03 USA | Patient's ascitic fluid | Patient's blood culture |
| ompW | ompW 1/2 | 588 | + | + | + | + | + |
| ompW | ompW1/4 | 304 | + | + | + | + | + |
| ompW | ompW 2/4 | 336 | + | + | + | + | + |
| ompW | ompW F/R | 373 | + | - | - | - | - |
| ToxR | Tox F/R | 337 | + | + | + | + | + |
| ctxA | ctxA F/R | 354 | + | - | - | - | - |
| ompU | ompU F/R | 283 | + | - | - | - | - |
| ompK | ompK F/R | 310 | + | - | - | - | - |
| TCP | TCP F/R | 265 | + | - | - | - | - |
Notes. Amplification by was done in a 25 μl reaction mixture containing 3 μl of template DNA, 200 μM of each dNTP, 1X reaction buffer, 1.25 units of Taq polymerase (TaKaRa Ex-Taq), and 0.5 μM of each primers. The initial amplification conditions were one cycle at 95°C for 7 minutes, 35 cycles at 95°C for 30 seconds. 55°C for 30 seconds, and 72°C for 30 seconds; and elongation step of 72°C for 5 minutes. Nested PCR was performed with the same method except that the annealing temperature was 60°C and the template was 2 μl of the first PCR product. Electrophoresis was performed on a 1.8% agarose gel.
Table 2.
Primers and primer sequences. From reference [16] and this paper
| Primer name | Sequence | Accession number if designed in this experiment |
| ompW-1 | CACCAAGAAGGTGACTTTATTGTG | [16] |
| ompW-2 | GAACTTATAACCACCCGCG | [16] |
| ompW-3 | CCACCTACCTTTATGGTCC | [16] |
| ompW-4 | GGAAAGTCGAATTAGCTTCACC | [16] |
| ompW-F | GTTTTTGAAGTCCTCGCTGCT | NC_002506 |
| ompW-R | GCATCTGCACCTGCTTTGTA | NC_002506 |
| ctxA-F | TCAGACGGGATTTGTTAGGC | AF463400 |
| ctxA-R | CCTGCCAATCCATAACCATC | AF463400 |
| toxR-F | GATTAGGCAGCAACGAAAGC | M21249 |
| toxR-R | GATGAAGGCACACTGCTTGA | M21249 |
| ompU-F | GCTGTAGCAGACCGTGTTGA | NC_002505 |
| ompU-R | GGTTTTCCATGCGGTAAGAA | NC_002505 |
| ompK-F | GCAACGAACAAAAGCAGTGA | NC_002505 |
| ompK-R | ACCAGTTGGTCGAGATTTGG | NC_002505 |
| TCP-F | TGGGCAGATATTTGTGGTGA | NC_002505 |
| TCP-R | TTTCTGCAACTCCTGTCAACAT | NC_002505 |
Figure 1.

Agarose gel electrophoresis analysis of V. cholerae from patient's ascitic fluid culture and blood culture isolates, 109-68, 418-02 and 569B. Lane M is the 100 bp lambda ladder marker (Toyobo, Japan); lanes 1–4 are ompW (the primer pairs 1/2, 1/4, 2/4, F/R) and lanes 5–9 are toxR, ctxA, ompU, ompK and TCP.
Figure 2.

Scanning electron micrograph of an environmental V. cholerae isolate from USA (418-03).
Figure 3.
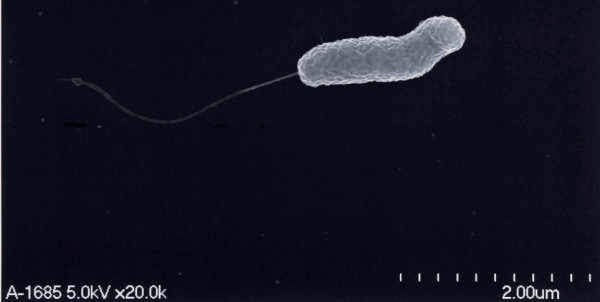
Scanning electron micrograph of a human V. cholerae isolate from India (109-68).
Figure 4.

Scanning electron micrograph of V. cholerae organisms cultured from the patient, demonstrating the absence of flagellae.
Table 3.
Primers used for PCR of the flaA gene of V. cholerae (designed using V. cholerae accession number AF007121 and the JustBio primer soft software)
| Primer name | Sequence | Product length (bp) |
| FlaA_F1 | GACCGCACAACGTTATCTGA | 321 |
| FlaA_R1 | AGTGCCACCGACTCTTCATT | |
| FlaA_F2 | AACCGTATCGCTGAAACCAC | 323 |
| FlaA_R2 | TCCGTTTGACCGTTGATGTA | |
| FlaA_F3 | AAGCTTCGGTTGACCAAGAA | 338 |
| FlaA_R3 | TCCTTCGCAAAATCCGTATC |
The 021 V. cholerae cultured from blood and ascitic fluid of our patient were non-motile. V. cholerae are normally motile, although non-motile variants of Classical O1 may arise in the laboratory [17,18] and non-01 non-motile clinical isolates have been described [19]. The combination of SEM and PCR demonstrated that the organisms lacked flagellae but did possess the important regulatory flagellin gene flaA on the genome. The underlying reasons for the absence of flagellae in the patient's isolates are being investigated.
Non-01 V. cholerae bacteremia has been described from Thailand [9,20] and recently from Laos (Phouthavane et al. unpublished). Our patient may have contracted the infection from the molluscs or from injuries whilst walking barefoot on the edge of the lake. An association between V. cholerae and non-biting chironomid midges (Diptera; Chironomidae) egg masses, which live in fresh water, has been described [21] and such midges may be important in the ecology of cholera. The collection and consumption of snails are an important part of life in Laos, as in rural communities in Australia and USA where non O1 V. cholerae bacteremia has been described amongst people who fish [22]. V. cholerae are usually associated with water of medium salinity as it requires trace amounts of sodium chloride for growth but it can be contracted from fresh water environments [4,8]. Although non-01 and non-0139 V. cholerae are not usually associated with epidemics, in Bangladesh non-01 V. cholerae has caused outbreaks [23]. The optimum treatment remains uncertain but third generation cephalosporins, doxycycline and fluoroquinolones have been suggested [3].
Conclusion
Fatal bacteremic non-O1, non-O139 V. cholerae is reported from Laos. The organisms were unusual in being non-motile. They did however possess the Vibrio flagellin flaA gene. Further research on the determinants of the non-motility is required. Given the importance of fishing and water in Lao society and the high prevalence of cirrhosis secondary to hepatitis B and C infection [24], V. cholerae non-01 bacteraemia may be a more important disease than this case report suggests.
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
RP was in charge of diagnosis and edited the ms, MN performed PCR assays and helped write ms, EA performed diagnosis work up and SEM, and edited ms, VD, MV and OL performed diagnosis work up and edited ms, CEM assisted with diagnostic workup, reviewed molecular analysis and edited ms, SN performed diagnosis workup and helped write ms and PNN assisted with diagnostic workup, wrote the first draft and edited ms. All authors read and approved the final manuscript.
Acknowledgments
Acknowledgements
We are very grateful to the family of the patient for their help and to the Minister of Health, His Excellency Dr Ponmek Dalaloy and the Director of the Curative Department, Ministry of Health, Professor Sommone Phounsavath for their support for this study, which was part of the Wellcome Trust-Mahosot Hospital-Oxford Tropical Medicine Research Collaboration and the 'Sustainable Co-existence of Humans, Nature and the Earth' project of Research Revolution 2002, Government of Japan. We thank Dr Sinthavong Vilavong and Mr Lienag Khamsivilay of the Fisheries Research Centre, Ministry of Agriculture and Forestry, Government of the Lao PDR, for kindly determining the identity of the mollusks and anonymous reviewers and Dr Sharon Peacock for their advice. Written informed consent was obtained from the patient's relative for publication of this study.
Financial Support. Wellcome Trust of Great Britain (RP, VD, MV, OL, CEM, PNN) and Ministry of Education, Culture, Science, Sports and Technology, Japan (MN, EA, SN).
Contributor Information
Rattanaphone Phetsouvanh, Email: rphetsouvanh@yahoo.co.uk.
Masami Nakatsu, Email: nakatsu-masami@umin.ac.jp.
Eiji Arakawa, Email: earakawa@nih.go.jp.
Viengmone Davong, Email: viengmon9@yahoo.com.
Manivanh Vongsouvath, Email: manivanh100@hotmail.com.
Olay Lattana, Email: olay@yahoox.uk.
Catrin E Moore, Email: catrin@tropmedres.ac.
Satoshi Nakamura, Email: snakamura@ri.imcj.go.jp.
Paul N Newton, Email: paul@tropmedres.ac.
References
- Midorikawa Y, Nakamura S, Iwade Y, Sugiyama A, Sisavath L, Phakhounthong R. Bacterial diarrhea in Laos, a region where cholera was endemic. Southeast Asian J Trop Med Public Health. 1996;27:724–7. [PubMed] [Google Scholar]
- Toma C, Nakasone N, Song T, Iwanaga M. Vibrio cholerae SXT element, Laos. Emerg Infect Dis. 2005;11:346–7. doi: 10.3201/eid1102.040794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernandez JM, Serrano M, De Arriba JJ, Sanchez MV, Escribano E, Ferreras P. Bacteremic cellulitis caused by Non-O1, Non-O139 Vibrio cholerae: report of a case in a patient with hemochromatosis. Diagn Microbiol Infect Dis. 2000;37:77–80. doi: 10.1016/S0732-8893(99)00153-4. [DOI] [PubMed] [Google Scholar]
- Rabadan PM, Vilalta E. Non-O:1 Vibrio cholerae bacteremia. Rev Infect Dis. 1989;11:667. doi: 10.1093/clinids/11.4.667. [DOI] [PubMed] [Google Scholar]
- Ichinose Y, Ehara M, Shimizu T, Ohmagari T, Kaku M, Hara K, Tsuji T, Simada T. The characterisation of Vibrio cholerae Non-O1 strain causing perforative pan-peritonitis. Jap J Inf Dis. 2001;75:696–702. doi: 10.11150/kansenshogakuzasshi1970.75.696. [DOI] [PubMed] [Google Scholar]
- Kontoyiannis DP, Calia KE, Basgoz N, Calderwood SB. Primary septicemia caused by Vibrio cholerae non-O1 acquired on Cape Cod, Massachusetts. Clin Infect Dis. 1995;21:1330–3. doi: 10.1093/clinids/21.5.1330. [DOI] [PubMed] [Google Scholar]
- Namdari H, Klaips CR, Hughes JL. A cytotoxin-producing strain of Vibrio cholerae non-O1, non-O139 as a cause of cholera and bacteremia after consumption of raw clams. J Clin Microbiol. 2000;38:3518–3519. doi: 10.1128/jcm.38.9.3518-3519.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson AM, Varkey JB, Petti CA, Liddle RA, Frothingham R, Woods CW. Non-O1 Vibrio cholerae septicemia: case report, discussion of literature, and relevance to bioterrorism. Diagn Microbiol Infect Dis. 2004;49:295–7. doi: 10.1016/j.diagmicrobio.2004.04.016. [DOI] [PubMed] [Google Scholar]
- Suankratay C, Phantumchinda K, Tachawiboonsak W, Wilde H. Non-serogroup O:1 Vibrio cholerae bacteremia and cerebritis. Clin Infect Dis. 2001;32:117–119. doi: 10.1086/319596. [DOI] [PubMed] [Google Scholar]
- Crump JA, Bopp CA, Greene KD, Kubota KA, Middendorf RL, Wells JG, Mintz ED. Toxigenic Vibrio cholerae serogroup O141-associated cholera-like diarrhoea and blood stream infection in the United States. J Infect Dis. 2003;187:866–867. doi: 10.1086/368330. [DOI] [PubMed] [Google Scholar]
- Eltahawy AT, Jiman-Fatani AA, al-Alawi MM. A fatal non-O1 Vibrio cholerae septicemia in a patient with liver cirrhosis. Saudi Med J. 2004;25:1730–31. [PubMed] [Google Scholar]
- Khan AM, Albert MJ, Sarker SA, Bhattacharya MK, Azad AK. Septicemia due to Vibrio cholerae O139 Bengal. Diagn Microbiol Infect Dis. 1995;22:337–8. doi: 10.1016/0732-8893(95)00140-4. [DOI] [PubMed] [Google Scholar]
- Lin CJ, Chiu CT, Lin DY, Sheen IS, Lien JM. Non-O1 Vibrio cholerae bacteremia in patients with cirrhosis: 5-yr experience from a single medical center. Am J Gastroenterol. 1996;91:336–4. [PubMed] [Google Scholar]
- Ninin E, Caroff N, El Kouri D, Espaze E, Richet H, Quilici ML, Fournier JM. Nontoxigenic Vibrio cholerae O1 bacteremia: case report and review. Eur J Clin Microbiol Infect Dis. 2000;19:489–91. doi: 10.1007/s100960000296. [DOI] [PubMed] [Google Scholar]
- Ottaviani D, Masini L, Bacchiocchi S. A biochemical protocol for the isolation and identification of current species of Vibrio in seafood. J Appl Microbiol. 2003;95:1277–84. doi: 10.1046/j.1365-2672.2003.02105.x. [DOI] [PubMed] [Google Scholar]
- Nandi B, Nandy RK, Mukhopadhyay S, Nair GB, Shimada T, Ghose AC. Rapid method for species-specific identification of Vibrio cholerae using primers targeted to the gene of outer membrane protein OmpW. J Clin Microbiol. 2000;38:4145–4151. doi: 10.1128/jcm.38.11.4145-4151.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klose KE, Mekalanos JJ. Differential regulation of multiple flagellins in Vibrio cholerae. J Bacteriol. 1998;180:303–16. doi: 10.1128/jb.180.2.303-316.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mostow P, Richardson K. High-frequency spontaneous mutation of classical Vibrio cholerae to a nonmotile phenotype. Infect Immun. 1990;58:3633–9. doi: 10.1128/iai.58.11.3633-3639.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shehabi AA, Drexler H, Richardson SH. Virulence mechanisms associated with clinical isolates of non-O1 Vibrio cholerae. Zentrabl Bakteriol Mikrobiol Hyg [A] 1986;261:232–9. doi: 10.1016/s0176-6724(86)80040-2. [DOI] [PubMed] [Google Scholar]
- Thisyakorn U, Reinprayoon S. Non-0=O1 Vibrio cholerae septicemia: a case report. Southeast Asian J Trop Med Public Health. 1990;21:149–50. [PubMed] [Google Scholar]
- Halpern M, Broza YB, Mittler S, Arakawa E, Broza M. Chironomid egg masses as a natural reservoir of Vibrio cholerae non-O1 and non-O139 in freshwater habitats. Microb Ecol. 2004;47:341–9. doi: 10.1007/s00248-003-2007-6. [DOI] [PubMed] [Google Scholar]
- Heath CH, Garrow SC, Golledge CL. Non-O1 Vibrio cholerae: a fatal cause of sepsis in northern Australia. Med J Aust. 2001;174:480–1. [PubMed] [Google Scholar]
- Albert MJ, Siddique AK, Islam MS, Faruque AS, Ansaruzzaman M, Faruque SM, Sack RB. Large outbreak of clinical cholera due to Vibrio cholerae non-O1 in Bangladesh. Lancet. 1993;341:704. doi: 10.1016/0140-6736(93)90481-u. [DOI] [PubMed] [Google Scholar]
- Jutavijittum P, Yousukh A, Samountry B, Samountry K, Ounavong A, Thammavong T, Keokhamphue J, Toriyama K. Seroprevalence of hepatitis B and C virus infections among Lao blood donors. Southeast Asian J Trop Med Public Health. 2007;38:674–9. [PubMed] [Google Scholar]