

Abstract
Novel 2-(4-aminophenyl)benzothiazoles possess highly selective, potent antitumour properties in vitro and in vivo. They induce and are biotransformed by cytochrome P450 (CYP) 1A1 to putative active as well as inactive metabolites. Metabolic inactivation of the molecule has been thwarted by isosteric replacement of hydrogen with fluorine atoms at positions around the benzothiazole nucleus. The lipophilicity of these compounds presents limitations for drug formulation and bioavailability. To overcome this problem, water soluble prodrugs have been synthesised by conjugation of alanyl- and lysyl-amide hydrochloride salts to the exocyclic primary amine function of 2-(4-aminophenyl)benzothiazoles. The prodrugs retain selectivity with significant in vitro growth inhibitory potency against the same sensitive cell lines as their parent amine, but are inactive against cell lines inherently resistant to 2-(4-aminophenyl)benzothiazoles. Alanyl and lysyl prodrugs rapidly and quantitatively revert to their parent amine in sensitive and insensitive cell lines in vitro. Liberated parent compounds are sequestered and metabolised by sensitive cells only; similarly, CYP1A1 activity and protein expression are selectively induced in sensitive carcinoma cells. Amino acid prodrugs meet the criteria of aqueous solubility, chemical stability and quantitative reversion to parent molecule, and thus are suitable for in vivo preclinical evaluation.
British Journal of Cancer (2002) 86, 1348–1354. DOI: 10.1038/sj/bjc/6600225 www.bjcancer.com
© 2002 Cancer Research UK
Keywords: 2-(4-aminophenyl)benzothiazole, prodrug, CYP1A1
Novel 2-(4-aminophenyl)benzothiazoles possess remarkable and intriguing antitumour properties (Shi et al, 1996; Bradshaw et al, 1998a) representing a mechanistic class distinct from clinically used chemotherapeutic agents. Consistently, they are active against only a specific subset of human cancer cell lines in the National Cancer Institute (NCI) in vitro anticancer drug screen, producing mean graph patterns that are highly characteristic of this class of compounds only. Moreover, growth inhibition in vitro is characterised by a unique biphasic dose-response relationship (Bradshaw et al, 1998a,b). In vivo, 2-(4-amino-3-methylphenyl)benzothiazole compound 1 outperformed the 3′-halogeno counterparts in breast, colon and ovarian xenograft studies.
Compound 1 is efficiently sequestered by sensitive cell lines (e.g. breast MCF-7, MDA-468; renal TK-10) (Chua et al, 1999; Kashiyama et al, 1999). CYP1A1 mRNA (Hose et al, 2001; Monks et al, 2001) activity and protein expression (Chua et al, 2000) are induced exclusively in sensitive cell lines. Covalent binding, detected between compound 1 and recombinant CYP1A1, requires metabolism and is significantly reduced by glutathione (Chua et al, 2000). This suggests that an electrophilic, reactive intermediate species is formed. The C-6 oxidation biotransformation product, liberated into nutrient media, however, is devoid of antitumour activity (Kashiyama et al, 1999). Moreover, this metabolite antagonises cellular uptake of compound 1, covalent binding between CYP1A1 and compound 1, CYP1A1 activity and growth inhibition induced by compound 1. In addition, hydroxy derivatives of compound 1 possess mitogenic properties at μM concentrations (Hutchinson et al, 2001). In contrast, insensitive cell lines (e.g. breast MDA-MB-435; renal A498, CAKI-1; prostate PC-3) neither retain nor metabolise compound 1 (Kashiyama et al, 1999).
Fluorinated analogues of 2-(4-aminophenyl)benzothiazoles have been synthesised which successfully block C-oxidation (Hutchinson et al, 2001). 2-(4-Amino-3-methylphenyl)-5-fluorobenzothiazole (compound 2) is the favoured analogue for clinical consideration possessing enhanced efficacy in vitro and superior potency in vivo against human breast and ovarian tumour xenografts implanted in nude mice. 2-(4-Aminophenyl)benzothiazoles, which are synthetically accessible, small and lipophilic pose a pharmaceutical challenge, as aqueous i.v. formulations would be desired to minimise the possibility of first pass deactivating metabolism and improve drug bioavailability.
Amino acid conjugation may be utilised to enhance the aqueous solubility of amine or alcohol drugs (as amide or ester prodrugs respectively). The primary aromatic amine, dapsone, has been derivatised as amino acid amides fulfilling the criteria for a suitable prodrug–water solubility, chemical stability and rapid, quantitative bioreversion to the parent moiety (Pochopin et al, 1995). The exocyclic primary amine function of 2-(4-amino-3-methylphenyl)benzothiazoles has been successfully conjugated to alanine and lysine residues as the mono- and dihydrochloride salts respectively (Hutchinson et al, 2002); such structural manipulation converts lipophilic benzothiazoles into water soluble prodrugs (Figure 1). Herein, reversion of prodrugs to parent molecule and selective antitumour activity in vitro has been described. In addition, we demonstrate that interaction with the putative molecular target of this class of agent, CYP1A1, has not been compromised.
Figure 1.
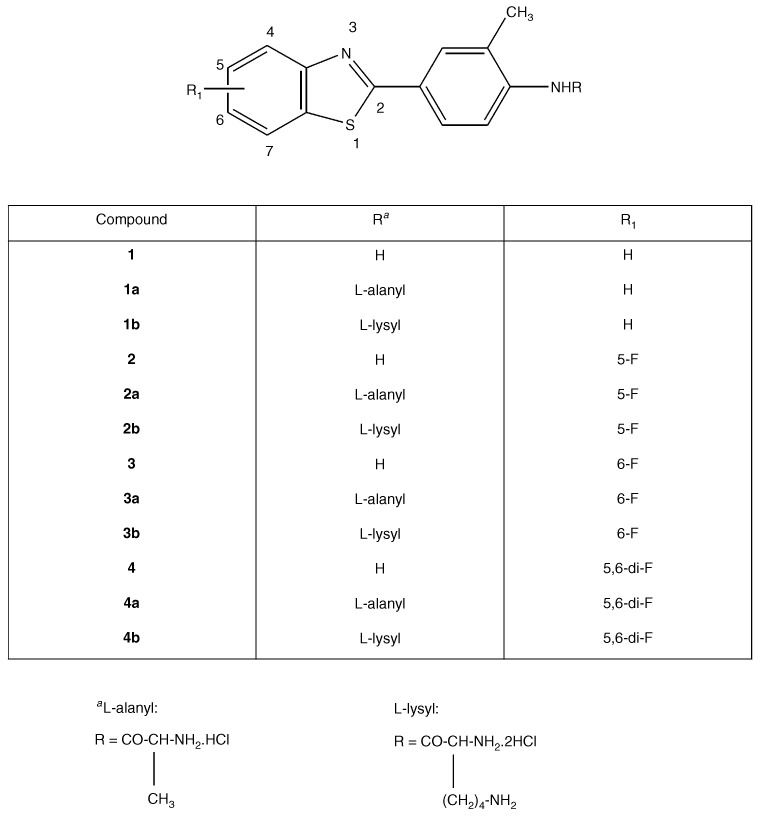
Structures of 2-(4-aminophenyl)benzothiazoles and their amino acid prodrugs.
MATERIALS AND METHODS
Growth inhibitory assays
Prodrugs were prepared as 10 mM top stocks, dissolved in dimethylsulphoxide (DMSO) or sterile ddH2O, and stored at 4°C, protected from light for a maximum period of 4 weeks. MCF-7 (ER+) and MDA 468 (ER−) human derived breast carcinoma cells, cultivated at 37°C in an atmosphere of 5% CO2 in RPMI 1640 medium supplemented with 2 mM L-glutamine and 10% foetal calf serum, were routinely subcultured twice weekly to maintain continuous logarithmic growth. Cells were seeded into 96-well microtiter plates at a density of 5×103 per well and allowed 24 h to adhere before drugs were introduced (final concentration 0.1 nM–100 μM, n=8). Serial drug dilutions were prepared in medium immediately prior to each assay. Viable cell masses at the time of drug addition (time-zero), and following 72 h drug exposure were determined by cell-mediated 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) reduction. MTT was added to each well (final concentration 400 μg ml−1) and plates were incubated at 37°C for 4 h to allow reduction of MTT by viable cell dehydrogenases to an insoluble formazan product. Well supernatants were aspirated and cellular formazan solubilised by addition of DMSO : glycine buffer (pH 10.5) (4 : 1). Cell growth as well as drug activity were determined by measuring absorbance at 550 nm using an Anthos Labtec systems plate reader.
NCI in vitro cytotoxicity assays
Cell culture and drug application procedures have been described previously (Boyd and Paull, 1995). Briefly, cell lines were inoculated into a series of 96-well microtiter plates, with varied seeding densities depending on growth characteristics of each cell line. Following a 24 h drug free incubation, test agents were added at five 10-fold dilutions with a maximum concentration of 100 μM. Cellular protein levels were determined after 48 h drug exposure by sulphorhodamine B colorimetry.
Metabolism studies
MCF-7 cells were seeded into 25 ml flasks at appropriate densities (5×105–5×106). After 24 h, medium was changed and drug introduced at a final concentration of 10 μM. Media samples, collected from flasks at time zero and 24 h intervals, were mixed with 1.5-fold volumes of high performance liquid chromatography (HPLC) grade acetonitrile to precipitate protein and centrifuged at 14 000 g for 5 min. Supernatants (600 μl) were mixed with 400 μl 10% acetonitrile in 1% acetic acid and analysed by HPLC. The system consisted of Beckman System Gold equipment (solvent module 128, autosampler 507e and multiple wavelength UV detector 168). Separation of parent compounds and biotransformation products was effected at room temperature on a Phenomenex Aqua C18 reversed-phase column (150×4.6 mm). The mobile phase was formed by increasing, then decreasing the acetonitrile concentration in 1% acetic acid over 20 min by mixing two solutions: acetonitrile-water-acetic acid (10 : 90 : 1, v v−1) and acetonitrile-water-acetic acid (80 : 20 : 1 v v−1) and delivered at a flow rate of 1 ml min−1. Compounds were detected at 324 nm.
Western blot protocol
Whole cell lysates were prepared for examination of CYP1A1 protein expression from untreated MCF-7, IGROV-1 and HCT 116 cultures and following exposure of cells to compounds 1, 2, 2a, 2b, 3, 3a, 3b, 4, 4a and 4b. Following protein determination (n=3, Bradford, 1976) and addition of sample buffer, samples were boiled at 95°C for 5 min and solubilised proteins (50 μg) were separated by sodium dodecyl sulphate (SDS) polyacrylamide gel (10%) electrophoresis. Proteins were electroblotted to polyvinylidene difluoride (PVDF) membranes and probed for CYP1A1 protein with polyclonal antiserum specific for human CYP1A1/1A2 (Gentest Corporation). Secondary antibody was conjugated to alkaline phosphatase, and CYP1A1 was detected following brief (<10 min) incubation with bromochloroindolyl phosphate and nitro-blue tetrazolium in alkaline phosphatase buffer. Molecular weight markers and a positive control of recombinant CYP1A1 (Gentest Corporation), included in all blots, confirmed detection of 52 kDa CYP1A1 protein.
Determination of ethoxyresorufin O-deethylase (EROD activity)
A sensitive and rapid fluorometric assay was used to measure EROD activity (Burke et al, 1994). Incubation mixtures (total 1 ml) consisted of 100 mM Tris-HCl (pH 7.4), 50 μM MgCl2, 100 μM 7-ethoxyresorufin and 100 μl cell homogenate. Homogenates were prepared following treatment of cells for 24 h with compounds 1, 2, 2a, 2b, 3, 3a, 3b, 4, 4a and 4b or vehicle alone and protein content determined (n=3). Thus induction of ethoxyresorufin O-deethylation by agents under study, catalysed by CYP1A1 activity, could be determined. Alternatively, microsomes expressing recombinant CYP1A1 (0.1 mg ml−1) provided the enzyme source for EROD catalysis, in the presence or absence of drug, in order to determine inhibition of CYP1A1 activity by compounds 1, 2, 2a, 2b, 3, 3a, 3b, 4, 4a and 4b. Incubation mixtures were pre-incubated for 5 min at 37°C before initiation of reaction by addition of NADPH (500 μM). Following further incubations at 37°C (30 min for cell homogenates, 15 min for CYP1A1 microsomes), reactions were terminated by addition of 3 ml ice-cold acetonitrile. Reaction mixtures were centrifuged (1400 r.p.m., 10 min) before analyses of supernatants. Fluorescence was read on a Perkin-Elmer LS-5 luminescence spectrometer (excitation, 530 nm; emission 585 nm). Estimation of resorufin reaction product (pmoles mg−1 protein, pmoles 100 μg−1 microsomes), as a measure of CYP1A1 activity, was determined following performance of a resorufin standard curve.
RESULTS
Prodrug uptake and conversion to parent amine
Uptake and metabolism of compounds 1a and 1b were examined in sensitive breast (MCF-7 and T47D) and insensitive renal (A498) cells using HPLC conditions which allow simultaneous detection of prodrug, parent amine and metabolites. When incubated at 0.1, 1 or 10 μM over 7 days at 37°C, the lysyl-amide compound 1a was most readily removed from the media supporting MCF-7 and T47D cells (Figure 2). The decline of each prodrug in media was accompanied by the appearance of compound 1. Media levels of regenerated compound 1 reached a peak at day 3, thereafter declining as oxidative metabolism by MCF-7 and T47D sensitive cells yielded its inactive 6-hydroxy metabolite. In contrast, negligible net removal of effluxed compound 1 occurred from media supporting the growth of inherently resistant A498 cells, which fail to sequester and metabolise 2-(4-aminophenyl)benzothiazoles (Kashiyama et al, 1999). Thus, compound 1, regenerated from the amide prodrugs accumulated harmlessly in the media of A498 cells over the 7 day incubation period.
Figure 2.
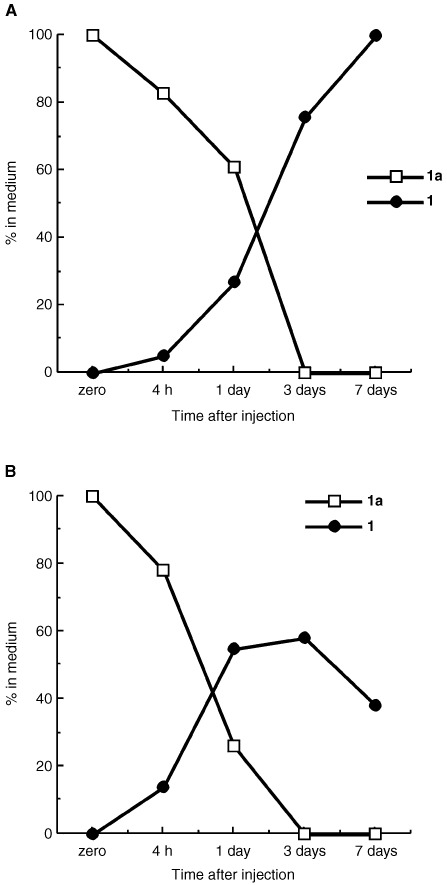
Uptake and conversion of 10 μM compounds 1a to 1 in: (A) inherently insensitive A498 renal cells and (B) sensitive ER+ T47D breast cells. Compound 1a is rapidly and efficiently hydrolysed by both cell lines. Only sensitive cells (B) sequester regenerated compound 1.
Amino acid prodrugs of compounds 2, 3 and 4 were stable in medium alone at 37°C>7 days: detection of parent drug during this time was negligible. Uptake and metabolism of compounds 2a, 2b, 3a, 3b, 4a and 4b (10 μM) by MCF-7 cells were examined; conditions for HPLC analysis allowed detection of prodrug, parent compound and metabolites. All alanyl- and lysyl-amide prodrugs were rapidly depleted from nutrient media in the presence of cells and conversion of prodrugs to parent amines was apparent after 24 h. Release of compound 2 into nutrient medium was significant after 24 h and continued to increase throughout the 7 day period. No biotransformation products could be detected (Figure 3A). In contrast, emergence of an oxidative metabolite accompanied detection of compounds 3 and 4 (days 1–7, Figure 3B,C).
Figure 3.
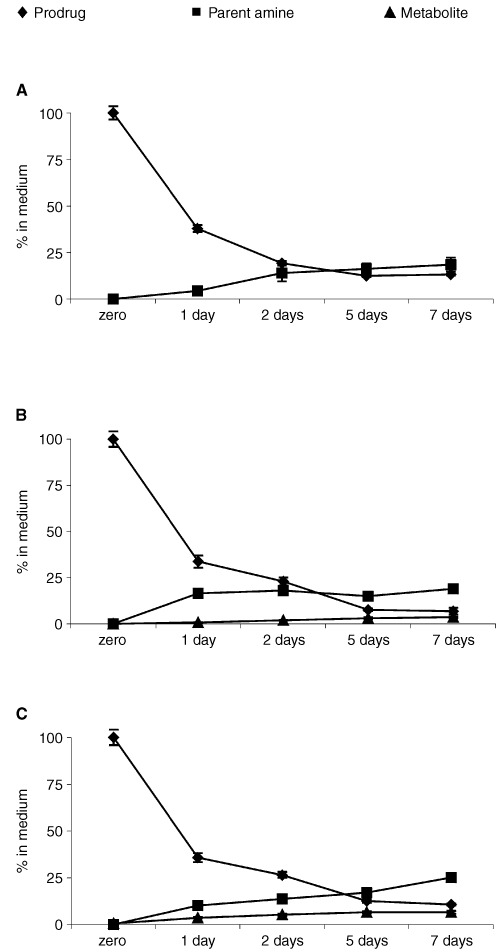
Depletion of (A) compound 2a, (B) compound 3a and (C) compound 4a (10 μM) from nutrient media supporting MCF-7 growth, conversion of prodrugs to the corresponding parent amine and further metabolism of compounds 3 and 4 only by MCF-7 cells (B and C).
In vitro cytotoxicity
In the NCI panel of human-derived carcinoma cell lines, tested amino acid salts of compounds 1, 2 and 3 retained the selective growth inhibitory properties of their parent amines; mean graphs demonstrated activity against certain ovarian, renal and breast cell lines following standard 48 h exposure (e.g. Figure 4). However, as expected considering the prodrug nature of compound 2b, following 48 h exposure, potency at the GI50 level in sensitive cell lines was reduced. Data obtained following examination of activity of amino acid salts of compounds 1, 2, 3 and 4 in MCF-7 and MDA 468 breast cancer cells corroborates this observation: typically, after 72 h exposure, GI50 values were two orders of magnitude greater than the parent amine (Table 1). However, neither alanyl nor lysyl prodrugs evoked the biphasic dose response characteristic of sensitive cell lines exposed to non-conjugated parent amines compounds 1, 3 or 4.
Figure 4.
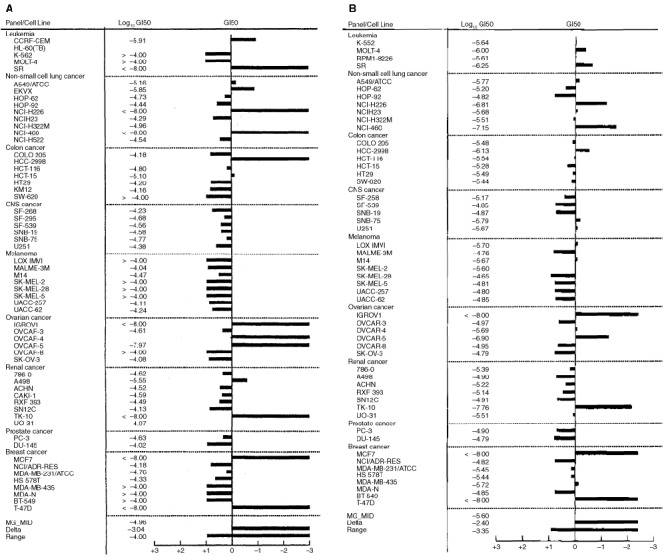
Mean GI50 graph demonstrating the selective nature of growth inhibition by (A) compound 2 and (B) compound 2b in the NCI in vitro panel of human derived cancer cell lines. Cells were exposed to drug for 48 h before viability was determined using the sulforhodamine B assay. Display of data utilises the mean graph format where the average GI50 concentration is plotted on the centre line. Using a log scale, the sensitivity or resistance of a particular cell line is represented by deflections to the right or left, respectively, from the mean.
Table 1. Retention of biological activity following conjugation of alanyl and lysyl residues to 2-(4-amino-3-methylphenyl)benzothiazole compounds 1, 2, 3 and 4.
Mechanism of action studies
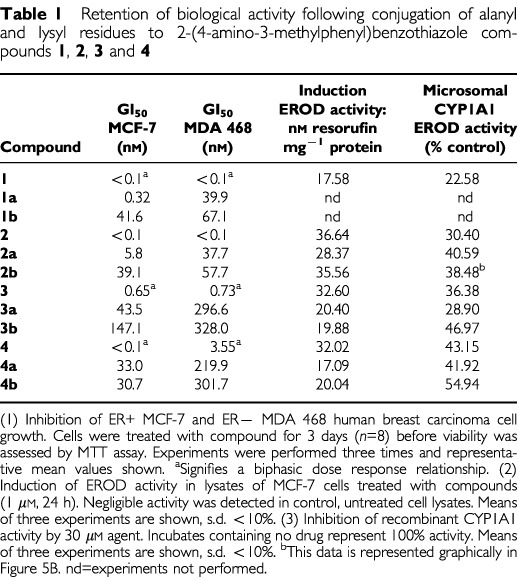
Figure 5 demonstrates selective induction of CYP1A1 protein expression in lysates of MCF-7 cells treated with benzothiazole prodrugs and their parent amines (1 μM, 24 h). No CYP1A1 was detected in lysates of HCT 116 or untreated MCF-7 cells. In addition, expression of CYP1A1 protein was clearly observed in lysates of IGROV-1 cells after exposure to concentrations of compound 2b between 10 nM and 10 μM (24 h, result not shown). Similarly, only lysates of sensitive cells exposed to prodrugs effected deethylation of ethoxyresorufin, indicative of CYP1A1 activity (Table 1). Figure 6A demonstrates the biphasic dose response relationship between CYP1A1-catalysed conversion of ethoxyresorufin to resorufin and compound 2b concentration in lysates of IGROV-1 cells exposed to this agent for 24 h. Maximum induction of EROD activity followed treatment of cells with 1 μM compound 2b. It has been speculated that these agents are metabolised by CYP1A1 to a reactive intermediate that covalently binds to, and then inactivates the enzyme; the activity of CYP1A1 microsomes was irreversibly inhibited by compound 1 (Chua et al, 2000). Indeed, significant inhibition of CYP1A1 microsomal EROD activity was monitored when incubates included compounds 1, 2, 2a, 2b, 3, 3a, 3b, 4, 4a, 4b (Table 1, Figure 6B).
Figure 5.
Western Blot detection of CYP1A1 protein induced in lysates of (A) MCF-7 cells exposed to compounds 1, 2, 3, 4, 2a, 2b, 3a, 3b, 4a and 4b (1 μM, 24 h). CYP1A1 expression was not detected constitutively in lysates of untreated MCF-7 or (B) HCT 116 cells and not induced in HCT 116 cells exposed to compounds 1, 2, 3 or 2b. For each sample, 50 μg total protein was loaded onto the gel.
Figure 6.
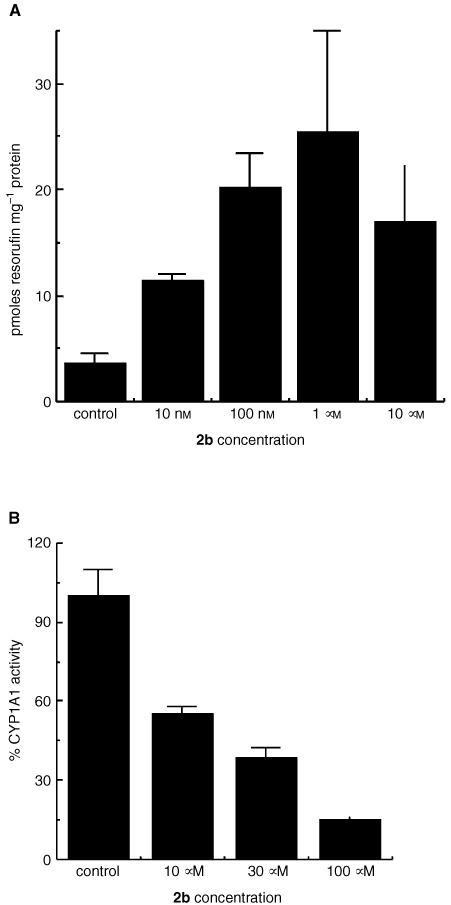
(A) EROD activity in homogenates of IGROV-1 cells treated with concentrations of compound 2b for 24 h. Mean values of three experiments are shown; bars, s.d. (B) Inhibition of CYP1A1 microsomal EROD activity by compound 2b. Resorufin product was measured after coincubation of CYP1A1 microsomes with compount 2b for 15 min at 37°C. Mean values of three experiments are shown; bars, s.d.
DISCUSSION
Data have been presented describing the in vitro biological properties of alanyl and lysyl amide derivatives of 2-(4-amino-3-methylphenyl)benzothiazoles. Prodrugs are rapidly depleted from nutrient media, generating their parent species in the presence of cells (Figures 2 and 3). In the absence of cells, no spontaneous hydrolysis of the amide bond occurs to release the amine from its amino acid salt. The cleavage of amino acid amides may be catalysed by ubiquitously expressed aminopeptidases, but subsequent bioactivating steps, essential for the mechanism of action of aminophenylbenzothiazoles, take place selectively in sensitive cell lines (e.g. Figure 5). Compounds 1a and 1b were stable when incubated at 37°C with human plasma; specifically, no free amine was detected. Amino acid amide stability, assessed in aqueous solution over a pH range of 4–9, was found to be pH dependent with more acidic pH favouring greater stability (Hutchinson et al, 2002).
The selective depletion of amine (e.g. compound 1, Figure 2b) from nutrient media in vitro, and accumulation in sensitive cells only has been reported previously (Kashiyama et al, 1999; Chua et al, 2000).
The remarkably potent and selective antitumour activity characteristic of this class of agent is retained; total growth inhibition (TGI) and cytocidal effects (LC50) were elicited by nM concentrations following amino acid conjugation coupled with monofluoro substitution. In vitro, prodrug compound 2b exhibits a superior antitumour profile. In vivo, plasma concentrations of compound 2, regenerated from lysylamide prodrug compound 2b, sufficient to elicit cytocidal activity against human mammary carcinoma cells persist >6 h (Bradshaw et al, 2002). Indeed, compound 2b suppresses significantly the growth of breast and ovarian xenografts in vivo.
Planar, hydrophobic 2-(4-aminophenyl)benzothiazoles fulfil structural requirements for binding to the aryl hydrocarbon receptor (AhR), and are potent AhR agonists (Loaiza-Perez et al, 2002). Subsequent induction of CYP1A1 mRNA (Hose et al, 2001; Monks et al, 2001) activity and protein expression (Chua et al, 2000; Hutchinson et al, 2001) only in cell lines sensitive to the growth inhibitory properties of 2-(4-amino-3-methylphenyl)benzothiazoles has been reported. That the role of CYP1A1 as a putative target for the mechanism of action of this class of compound was not compromised following prodrug modification of the 2-(4-aminophenyl)benzothiazole structure has been verified (Table 1, Figures 5 and 6). Induction of CYP1A1 protein expression and EROD activity in lysates of sensitive MCF-7 breast, IGROV-1 ovarian cells was observed following exposure to prodrugs. Lysates of insensitive HCT 116 cells treated with prodrugs neither expressed CYP1A1 protein nor effected ethoxyresorufin O-deethylation.
Induction of CYP1A1 activity and protein expression probably leads to generation of a reactive electrophilic species and benzothiazole-derived covalent binding to CYP1A1, resulting in enzyme inactivation (Chua et al, 2000). We have established that at prodrug compound 2b concentrations >1 μM, EROD activity, induced in lysates of treated sensitive cells, begins to decline (Figure 6A). Moreover, μM prodrug concentrations inhibit microsomal CYP1A1 activity (Figure 6B), substantiating the thesis that these agents are inducers, substrates and suicide inhibitors of CYP1A1. Generation of a CYP1A1-dependent reactive electrophilic species preceeds detection of DNA adducts (Stevens et al, 2001), and ultimately cell death.
In conclusion, alanyl- and lysyl-amide prodrugs of 2-(4-amino-3-methylphenyl)benzothiazoles undergo reversion to their parent amine. In vitro efficacy and selectivity are retained. Stark antitumour selectivity can be rationalised, and involvement of a crucial molecular target of this class of agent demonstrated. In addition, chemical stability and aqueous solubility is exhibited rendering these synthetically accessible prodrugs suitable for further preclinical development.
Acknowledgments
This study was supported by Cancer Research UK, and the National Cancer Institute. This is part 19 of the series ‘Antitumour 2-(4-aminophenyl)benzothiazoles’. The authors thank Cancer Research UK, for support to the Cancer Research UK Experimental Cancer Chemotherapy Research Group, Nottingham. We also thank the University of Nottingham, UK, for a scholarship (to M-S Chua). We gratefully acknowledge extensive collaborations, support and discussions with the staff of the Developmental Therapeutics Program of the NCI and members of the Screening and Pharmacology Group within the European Organisation for Research and Treatment of Cancer (EORTC).
References
- BoydMRPaullKD1995Some practical considerations and applications of the National Cancer Institute in vitro anticancer drug discovery screen Drug Development Res 3491109 [Google Scholar]
- BradfordMM1976A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding Anal Biochem 72248254 [DOI] [PubMed] [Google Scholar]
- BradshawTDWrigleySShiD-FSchultzRJPaullKDStevensMFG1998a2-(4-Aminophenyl)benzothiazoles: novel agents with selective profiles of in vitro antitumour activity Br J Cancer 77745752 [DOI] [PMC free article] [PubMed] [Google Scholar]
- BradshawTDShiD-FSchultzRJPaullKDKellandLWilsonAGarnerCFiebigHHWrigleySStevensMFG1998bInfluence of 2-(4-aminophenyl)benzothiazoles on growth of human ovarian carcinoma cell lines in vitro and in vivo Br J Cancer 78421429 [DOI] [PMC free article] [PubMed] [Google Scholar]
- BradshawTDBibbyMCDoubleJAFichtnerICooperPAAlleyMCDonohueSStinsonSFTomaszewjskiJESausvilleEStevensMFG2002Preclinical evaluation of amino acid prodrugs of novel 2-(4-amino-3-methylphenyl)benzothiazoles Mol Can Therapeutics 45239246 [PubMed] [Google Scholar]
- BurkeMDThompsonSWeaverRJWolfCRMayerRT1994Cytochrome P450 specificities of alkoxyresorufin O-dealkylation in human and rat liver Biochem Pharmacol 48923936 [DOI] [PubMed] [Google Scholar]
- ChuaM-SShiD-FWrigleySBradshawTDHutchinsonIShawPNBarrettDAStanleyLAStevensMFG1999Antitumour benzothiazoles. 7. Synthesis of 2-(4-acylaminophenyl)benzothiazoles and investigations into the role of acetylation in the antitumour activities of the parent amines J Med Chem 42381392 [DOI] [PubMed] [Google Scholar]
- ChuaM-SKashiyamaEBradshawTDStinsonSFBrantleyESausvilleEAStevensMFG2000Role of CYP1A1 in modulation of antitumour properties of the novel agent 2-(4-amino-3-methylphenyl)benzothiazole (DF 203, NSC 674495) in human breast cancer cells Cancer Res 6051965203 [PubMed] [Google Scholar]
- HoseCRivieraMSausvilleEMonksA2001Induction of cytochrome P450 CYP1A1 and cytochrome P450 CYP1B1 by 2-(4-amino-3-methylphenyl)benzothiazole (BZ) in 60 human tumour cell lines: correlation with BZ toxicity Proc Am Assoc Cancer Res 422749511 [Google Scholar]
- HutchinsonIChuaM-SBrowneHLTrapaniVFichtnerIBradshawTDWestwellADStevensMFG2001Antitumour benzothiazoles. 14. The synthesis and unique antitumour properties of fluorinated 2-(4-aminophenyl)benzothiazoles J Med Chem 4414461455 [DOI] [PubMed] [Google Scholar]
- HutchinsonIJenningsSAVishnuvajjalaBRWestwellADStevensMFG2002Antitumour benzothiazoles. 16. Synthesis and pharmaceutical properties of antitumour 2-(4-aminophenyl)benzothiazole amino acid prodrugs J Med Chem 45744747 [DOI] [PubMed] [Google Scholar]
- KashiyamaEHutchinsonIChuaM-SStinsonSFPhillipsLRKaurGSausvilleEABradshawTDWestwellADStevensMFG1999Antitumour benzothiazoles. 8. Synthesis, metabolic formation, and biological properties of the C- and N-oxidation products of antitumour 2-(4-aminophenyl)benzothiazoles J Med Chem 4241724184 [DOI] [PubMed] [Google Scholar]
- Loaiza-PérezAITrapaniVHoseCSinghSSTrepelJStevensMFGBradshawTDSausvilleEA2002The AhR mediates sensitivity of MCF-7 breast cancer cells to the antitumour agent 2-(4-amino-3-methylphenyl)benzothiazole Mol Pharmacol 611319 [DOI] [PubMed] [Google Scholar]
- MonksARiveraMIHarrisEHoseCSausvilleEA2001Gene expression changes induced by a novel antitumour agent, 2-(4-amino-3-methylphenyl)benzothiazole, in MCF-7, a sensitive breast cancer cell line, using cDNA microarrays Proc Am Assoc Cancer Res 423417635 [Google Scholar]
- PochopinNLCharmanWNStellaVY1995Amino acid derivatives of dapsone as water soluble prodrugs Int J Pharmaceutics 121157167 [Google Scholar]
- ShiD-FBradshawTDWrigleySMcCallCJLelieveldPFichtnerIStevensMFG1996Antitumour benzothiazoles. 3. Synthesis of 2-(4-aminophenyl)benzothiazoles and evaluation of their activities against breast cancer cell lines in vitro and in vivo J Med Chem 3933753384 [DOI] [PubMed] [Google Scholar]
- StevensMFGHeydonRTMartinEAFarmerPBBradshawTDHutchinsonIWestwellADBrowneHLTrapaniV2001Induction of CYP1A1 by 2-(4-aminophenyl)benzothiazoles leads to DNA adducts in sensitive tumour cells Proc Am Assoc Cancer Res 421754325 [Google Scholar]