

Abstract
Semantic and affective priming are classic effects observed in cognitive and social psychology, respectively. We discovered that affect regulates such priming effects. In Experiment 1, positive and negative moods were induced prior to one of three priming tasks; evaluation, categorization, or lexical decision. As predicted, positive affect led to both affective priming (evaluation task) and semantic priming (category and lexical decision tasks). However, negative affect inhibited such effects. In Experiment 2, participants in their natural affective state completed the same priming tasks as in Experiment 1. As expected, affective priming (evaluation task) and category priming (categorization and lexical decision tasks) were observed in such resting affective states. Hence, we conclude that negative affect inhibits semantic and affective priming. These results support recent theoretical models, which suggest that positive affect promotes associations among strong and weak concepts, and that negative affect impairs such associations (Kuhl, 2000; Clore & Storbeck, 2006).
Understanding how affect influences memory has long been a critical goal of emotion research. Despite the large literature, however, basic questions remain. For example, does affect promote the recall of mood-congruent memories? Bower famously wrote that, the “activation of an emotion node also spreads activation throughout the memory structures to which it is connected” (Bower, 1981, p. 135). If so, does the mood-congruency assumption apply to all types of memory? The answer is unclear because, “The specific nature of information represented in an emotion network has never been fully specified or empirically tested. Perhaps these networks do not include declarative knowledge about emotions” (Innes-Ker & Niedenthal, 2002, p. 813). Despite the plausibility and power of the mood-congruent memory hypothesis, evidence has been mixed at best, leading some to suggest that, “If mood does not prime semantic information, then mood and memory must be conceptualized differently from the way Bower (1981) conceptualized them” (Weaver & McNeill, 1992, p. 296). If affect does not prime semantic memory in such a content-specific manner, might it activate or direct attention to semantic knowledge more generally? For example, perhaps, “positive affect …is characterized in memory by the activation of wide semantic fields… In contrast, negative affect … is characterized by a more restricted spread of activation to close associates and dominant word meanings” (Bolte, Goschke, & Kuhl, 2003, p. 416; see Isen, 1987 for a similar point).
These questions and quotations from 20 years of the literature underline the logic of the argument proposed in this article about how affect influences memory, especially semantic memory. The mood congruent memory approach dominated early research and is still a commonly held position (Forgas, 2001; Rolls & Stringer, 2001). Initial results were often supportive (see Blaney, 1986, for a review). Typical studies observed that sad individuals recalled more negative events and happy individuals tended to recall more positive events.
Subsequent results, however, were mixed, with successful studies often focusing on episodic memory (Innes-Ker & Niedenthal, 2002). Many studies reporting negative results focused on semantic memory (Clark, Teasdale, Broadbent, & Martin, 1983; Gerrig & Bower, 1982; Weaver & McNeill, 1992). Two new assumptions emerged from these studies, including (1) that affect influences episodic but not also semantic memory processes (Weaver and McNeill, 1992), and (2) that affect influences controlled, but not automatic processing (Gerrig & Bower, 1982; Matthews & Wells, 1999).
An alternative possibility is that affect has general rather than content specific effects on memory. Some theorists, who had earlier assumed mood-congruency in memory, proposed instead that positive affect leads to “substantive” processing (Forgas, 2001) or that it activates semantic associations in memory generally (Ashby, Isen, & Turken, 1999; Kuhl, 2000; Smith and DeCoster, 2000). Others also suggested that positive affect promotes the activation and use of semantic associations, whereas negative affect inhibits their accessibility and use (Storbeck & Clore, 2005; Gasper, 2004). It would appear, then, that affect may regulate general semantic processing, rather than access to mood-specific content (e.g., Storbeck & Clore, 2005). Although this position seems sound, the studies supporting it generally focused on controlled processing, in which people consciously recruit associations related to the task at hand (e.g., Bolte, et al., 2003; Storbeck & Clore, 2005).
The question thus remains whether the influences of affect on semantic memory can be implicit and automatic or only explicit and controlled? We hypothesized that positive affect promotes spreading activation and negative affect inhibits spreading activation. The basis of this prediction is the notion that positive affect confers value on one’s natural, relational processing of verbal material, whereas negative affect indicates that such an orientation may be problematic. To test this hypothesis, in a straightforward way, we employed semantic priming paradigms.
Semantic Priming
Semantic priming is a reliable implicit measure of semantic associations (Neely, 1991), whereas affective priming is an equally reliable measure of affective associations (see Klauer & Musch, 2003). The term priming refers to an increase in speed of response when targets are preceded by a semantically related as opposed to a semantically unrelated prime (see Neely, 1991). For example, individuals typically identify the letter string DOCTOR (target) as a word (rather than a non-word) faster if they have just seen the related prime, NURSE, than if they have just seen an unrelated prime, such as TABLE. In the case of affective priming, targets are responded to faster when preceded by a similarly valenced prime than when preceded by a prime of a different valence (Klauer & Musch, 2003). For instance, the positive word SUNSHINE tends to be evaluated faster when preceded by the positive prime PUPPY than when preceded by the negative prime SNAKE.
The main explanation for semantic, as well as affective, priming has been spreading activation, a mechanism based on network models of memory (Klauer & Musch, 2003; Neely, 1991). Activation has been thought to spread from prime to target when the two share an associative link in semantic memory, thereby influencing decisions about targets. Such activation of associated concepts is assumed to occur automatically and without intention for both semantic priming (Neely, 1991) and affective priming (Bargh, 1997).
Response competition mechanisms can also account for both types of priming effects. Similar prime-target meanings require similar responses, and opposing prime-target meanings require different responses. Priming paradigms that confound stimulus and response compatibility limit the ability to draw conclusions about spreading activation (e.g., Klauer & Musch, 2003; De Houwer, 2003).
Affect and Priming
Priming paradigms have been used previously to ask whether affect influences associations in a mood-congruent manner. The assumption was that mood serves as a node in memory that is linked to other valence-congruent concept nodes. If so, positive moods should activate positive concepts and negative moods should activate negative concepts. Based on these assumptions, Clark et al. (1983) induced happy or sad moods prior to having participants judge whether or not good, bad, or neutral letter strings were words in a lexical decision judgment task. They expected that if moods activate valence-congruent concepts, then happy persons would be faster to judge positive words and sad persons would be faster to judge negative words. Contrary to expectations, mood did not influence the accessibility of valence-consistent targets (see also, Challis and Krane, 1988, for similar results).
Classic priming studies have also been used to examine whether affect influences memory in a mood-congruent manner. Hanze and Hesse (1993) used a lexical decision task and varied the semantic relatedness (high vs. low) between primes and targets. Positive and neutral moods were induced prior to the lexical decision task. Priming was observed for happy individuals only when primes and targets were strongly, but not when weakly, associated. The neutral group failed to demonstrate priming effects. In a subsequent study, Hesse and Spies (1996) induced a negative or neutral mood state before participants completed a lexical decision priming task with a long SOA (500 ms). This longer SOA is thought to facilitate controlled processing of primes and targets, whereas a shorter SOA (<300 ms) is thought to facilitate automatic processing of primes and targets. Hesse and Spies observed priming for the negative mood but not the neutral group, suggesting that negative moods may facilitate controlled processing.
The most comprehensive study of affect and priming, observed that positive, but not negative moods promoted semantic priming in a lexical decision task (Corson, 2002). He induced a positive, neutral, and negative mood followed by a lexical decision task with either a short (150ms) or long (1200 ms) SOA. The positive mood group demonstrated priming at both short and long SOA durations and no other priming effects were observed for the sad and control groups. These results of Henze and Spies, Spies and Hesse, and Corson, suggest that happy moods foster semantic priming in a lexical decision task, whereas sad moods do not. However, priming results with a longer SOA were mixed (Corson 2002; Hesse & Spies, 1996).
These studies suggest that positive moods may promote semantic priming. We assessed this hypothesis about semantic priming and related hypotheses about affective priming in three experiments. We simultaneously varied both the nature of the prime-target connection (affective vs. semantic) and whether task demands favored affective priming, semantic priming, or neither (Storbeck & Robinson, 2004). Prime-target connections were varied by using a comparative priming paradigm, which allows for multiple relations between primes and targets (Storbeck & Robinson, 2004). Word stimuli were selected so that affective and semantic connections between primes and targets were varied orthogonally. For example, the prime word SPIDER and target word JAGGED are congruent in valence (bad-bad), but incongruent in category relations (animal-texture). Task demands were varied by employing three different priming paradigms, including evaluative, categorical, and lexical decision priming tasks.
The comparative priming approach allows investigators to examine what types of associations are automatically influenced by mood, if any. We can observe whether mood acts automatically on affective associations, descriptive (semantic) associations, both, or neither. We thus test the hypothesis common to several theories, which propose that positive affect stimulates semantic associations (Ashby et al., 1999; Kuhl, 2000; Smith & DeCoster, 2000). These include the affect-as-information approach, which proposes that affective cues govern both the activation (e.g., Clore & Storbeck, 2006) and use (e.g., Gasper, 2004) of affective/semantic associations. Specifically, positive affect increases accessibility and use of associations that are task-relevant. Therefore, the nature of the priming effects obtained should depend on the demands of the task. Thus, evaluation tasks should make affective associations more accessible, leading to affective priming, whereas categorization tasks should make category associations more accessible, leading to category priming. In lexical decision tasks, whatever prime-target relation is more accessible as a function of the organization of memory should determine priming. If affective relations are more accessible, then affective priming should be evident; but if semantic relations are more accessible, as proposed by Storbeck and Robinson (2004), then descriptive priming should be evident. For negative affect, the affect-as-information approach proposes a reduction in the accessibility and use of such associations. Thus, sad persons should inhibit such associations between primes and targets, thereby preventing all priming effects.
We predicted that the happy mood group and control group would produce typical priming effects (affective priming in the evaluation task and category priming in the category and lexical decision tasks) and that sad moods would impair such priming effects.
EXPERIMENT 1
We conducted three priming experiments simultaneously, randomly assigning participants to two mood conditions (happy or sad) and three kinds of tasks (evaluative, categorical, and lexical decision).
Method
Participants
In the three experiments, 77 (evaluation task), 78 (categorization task), and 68 (lexical decision task) University of Virginia undergraduates participated to fulfill a course requirement.
Materials
Mood was manipulated between participants by music. The happy mood group listened to Eine Kliene Nacht Musik by Mozart for 12 minutes, while the sad mood group listened to Adagietto by Mahler for 12 minutes. Previous studies find that these musical pieces effectively induce happy and sad mood states, respectively (Storbeck & Clore, 2005). The mood manipulation check consisted of one question, which asked “how were you feeling while listening to the music” on a scale of very unhappy (1) to very happy (7).
For the priming task, we followed the procedures of Storbeck and Robinson (2004). The dependent variable was reaction time. Within participants, we assessed the role of Affective Congruency and Category Congruency between primes and targets. The stimuli included words representing two levels of affective meaning (good vs. bad) crossed with two levels of categorical meaning (animal vs. texture). Each category consisted of twenty words, and within each category there were ten good words and ten bad words (see, Storbeck & Robinson, 2004, for words and evaluative ratings). Each word appeared twice as prime and twice as target for a total of 80 trials.1 Selection of primes and targets were randomized on each trial, so that the affective and categorical relationships were also randomized. The primes were presented for 200 ms followed by a 100 ms blank interval for a 300 ms SOA.
Following Storbeck and Robinson (2004), we replaced the texture word list with a religious word list for the lexical decision task. This replacement introduced 20 new words, 10 positive and 10 negative. 40 non-word targets were introduced for the non-word trials. 160 trials were presented, 80 trials had word targets and 80 trials had non-word targets. Non-words never served as primes, and animal and religious words were presented four times as primes and twice as targets.
Procedure
Participants heard a cover story designed to disguise the purpose of the mood induction. The music was played by a computer through individual headphones. When the music ended, the priming task began. For the EVALUATION task, participants were instructed to ignore the first word (prime) and to evaluate the second word (target) by pressing either the 1 key for good words or the 9 key for bad words. For the CATEGORIZATION task, participants were instructed to ignore the prime and to categorize the target by pressing the 1 key for animal words or the 9 key for texture words. For the LEXICAL DECISION task, participants were instructed to ignore the prime and to decide whether the target was a word (1 key) or a non-word (9 key). After the priming task, participants completed the mood questionnaire.
Results
Mood Manipulation Check
Participants whose moods were not effectively manipulated were removed from the analyses: Evaluation task (Happy = 6, Sad = 2), Categorization task (Happy = 5, Sad = 3), and Lexical Decision task (Happy = 4, Sad = 3) (see, Bower, Monteiro, & Gilligan, 1978, for a similar procedure).
As a result of this procedure, for each task, the happy condition reported they were happier than the sad condition. Evaluation task [F(1, 67) = 15.5, p < 0.001, η = 0.44], Categorization task [F(1, 68) = 16.8, p < 0.001, η = 0.45], and Lexical Decision task [F(1, 59) = 16.9, p < 0.001, η = 0.47]. Table 1 shows self-reported mood scores.2
Table 1.
| Evaluation Task | |||||
|---|---|---|---|---|---|
| Mood Score |
Category Congruency |
Affect Congruency |
|||
| Group | CI | CC | AI | AC | |
| Happy (N = 34) | 5.9 | 2.958 (.1279) | 2.965 (.1348) | 2.973 (.1436) | 2.950 (.1208) |
| Sad (N = 35) | 4.9 | 2.941 (.1121) | 2.938 (.1089) | 2.939 (.1163) | 2.939 (.1063) |
| Control (N = 46) | 2.873 (.0972) | 2.875 (.0964) | 2.883 (.0961) | 2.865 (.100) | |
| Categorization Task | |||||
| CI | CC | AI | AC | ||
| Happy (N = 32) | 6.0 | 2.851 (.0722) | 2.839 (.0613) | 2.843 (.0677) | 2.847 (.0662) |
| Sad (N = 38) | 5.1 | 2.845 (.0714) | 2.848 (.0711) | 2.849 (.0709) | 2.843 (.0709) |
| Control (N = 46) | 2.826 (.0759) | 2.815 (.0826) | 2.820 (.0774) | 2.821 (.0782) | |
| Lexical Decision Task | |||||
| Group | CI | CC | AI | AC | |
| Happy (N = 31) | 5.9 | 2.849 (.0791) | 2.830 (.0731) | 2.840 (.0727) | 2.840 (.0814) |
| Sad (N = 30) | 4.7 | 2.883 (.0822) | 2.877 (.0732) | 2.882 (.0790) | 2.878 (.0812) |
| Control (N = 43) | 2.830 (.0779) | 2.799 (.084) | 2.820 (.0734) | 2.809 (.0860) | |
Notes. CI = Category Incongruent, CC = Category Congruent, AI = Affect Incongruent, AC = Affect Congruent. Standard Deviations are in the parentheses. Means and Standard Deviations are represented in log transformed reaction time values.
Reaction Time Analyses
The first five trials were treated as practice and removed from the analysis. Trials involving incorrect responses were also removed: Evaluation 3.9%, Categorization 3.5%, and Lexical Decision 3.5%. A log transformation was performed on the reaction time data to normalize the distribution (see Fazio, 1990). For ease of interpretation, raw reaction times will be displayed for descriptive purposes only.
We computed four latency means for each participant, two for Category Congruence (congruent vs. incongruent) and two for Affective Congruence (congruent vs. incongruent).
Evaluation Task
To test our prediction that the happy mood group would demonstrate affective priming, we conducted a 2 (Category Congruence) × 2 (Affective Congruence) × 2 (Mood) repeated measures ANOVA. A main effect for Affective Congruence appeared, F(1, 67) = 5.06, p = 0.028, η2 = 0.07. As expected, the Affective Congruence main effect was qualified by a significant interaction with Mood, F(1, 67) = 4.81, p = 0.032, η2 = 0.07. No other effects reached significance. Table 1 shows relevant means.3
Subsidiary analyses of the significant Affective Congruence by Mood interaction found, as predicted, that affective priming occurred in the happy mood group, F(1, 33) = 7.97, p = 0.008, η2 = 0.20, but not in the sad mood group, p = 0.97. All other effects were non-significant. Figure 1 (top panel) shows mean response times.
Figure 1.
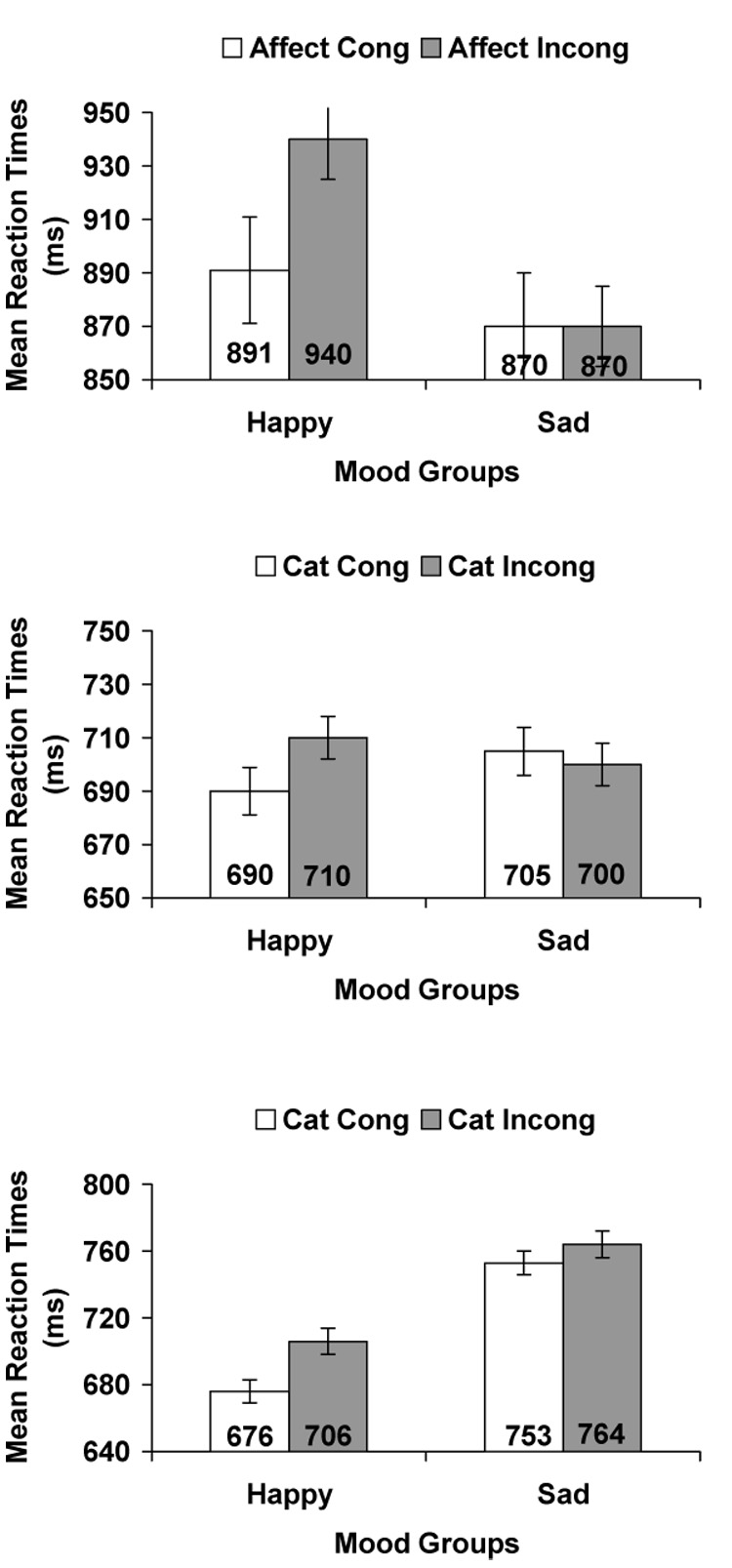
(Top panel) Mean latencies for Affective Congruence by Mood in the evaluation task (bars represent SE of the mean). (Middle panel) Mean latencies for Category Congruence by Mood in the categorization task. (Bottom panel) Mean latencies for Category Congruence by Mood in the lexical decision task.
The happy mood group demonstrated affective priming in the evaluation priming task, but the sad mood group failed to demonstrate any priming effects.
Categorization Task
To test the prediction that the happy mood group would demonstrate category priming, we ran a 2 (Category Congruence) × 2 (Affective Congruence) × 2 (Mood) repeated measures ANOVA. The results revealed only the predicted interaction of Category Congruence by Mood, F(1, 68) = 4.55, p = 0.036, η2 = 0.063. No other effects reached significance. See Table 1 for relevant means.4
To examine the Category Congruence by Mood interaction, we conducted a one-way ANOVA for Category Congruence by mood. As predicted, category priming appeared in the happy mood group, F(1, 31) = 6.3, p = 0.017, η2 = 0.17, but not in the sad mood group, p = 0.57. All other effects were non-significant. Figure 1 (middle panel) shows mean response times.
Thus, the happy mood group demonstrated category priming, whereas the sad mood group did not.
Lexical Decision Task
To test the prediction that the happy mood group would demonstrate category priming, we conducted a 2 (Category Congruence) × 2 (Affective Congruence) × 2 (Mood) repeated measures ANOVA. A main effect for Category Congruence appeared, F(1, 59) = 15.29, p < 0.001, η2 = 0.21. As predicted, it was qualified by a significant Category Congruence by Mood interaction, F(1, 59) = 4.41, p = 0.04, η2 = 0.07. In addition, a main effect for Mood, F(1, 59) = 4.3, p = 0.042, η2 = 0.068, was observed such that the happy group (M = 692 ms) was faster at making lexical decisions compared to the sad mood group (M = 759). No other effects reached significance. Table 1 shows relevant means.5
Subsequent analyses of the Category Congruence by Mood interaction found, as predicted, category priming for the happy mood group, F(1, 30) = 22,62, p < 0.001, η2 = 0.43, but not for the sad mood group, p = 0.26. All other effects were non-significant. Figure 1 (bottom panel) shows mean response times.
Thus, as predicted, the happy mood group showed category priming effects, whereas the sad mood group did not.
The results are quite clear from these three tasks, happy moods led to affective (evaluation task) and category (categorization and lexical decision tasks) priming, whereas sad moods inhibited such priming effects.
Experiment 2
Experiment 1 revealed a consistent pattern, happy mood states produced priming effects, whereas sad mood states failed to produce such priming effects. However, it is unclear as to whether happy moods promote or sad moods inhibit semantic and affective priming effects. To determine how affect influences priming, the same series of priming tasks were run with participants in their natural affective state. We anticipated that the control group, like the happy mood groups, would demonstrate category priming (categorization and lexical decision tasks) and affective priming (evaluation task). Therefore, if this pattern of results was found, it would suggest that sad moods inhibit semantic and affective priming.
Participants
46 University of Virginia undergraduates participated to fulfill a course requirement.
Materials
For each priming task, the stimuli and task features were identical to the analogous priming task in Experiment 1.
Procedure
Participants engaged in an unrelated experiment, and then completed the three priming tasks described in Experiment 1. The task order was randomly determined for each participant. Thus, each task had an equal chance of coming first, second, or third.6 A short break occurred between tasks. The priming procedures within each task were identical to the priming procedures in Experiment 1.
Results
Reaction Time Analyses
We treated the reaction time data as in Experiment 1. The first five trials and incorrect responses [Evaluation 8.5%, Categorization 4.9%, and Lexical Decision 5.3%] were removed from the analysis. The reactions times were then log transformed. For ease of interpretation, raw reaction times will be displayed for descriptive purposes. We computed four latency means for each participant, two for Category Congruence (congruent vs. incongruent) and two for Affective Congruence (congruent vs. incongruent).
Evaluation Task
To examine whether individuals in their natural affective state produce priming effects in the evaluation task, we conducted a 2 (Category Congruence) × 2 (Affective Congruence) repeated measures ANOVA. A main effect for Affective Congruence appeared, F(1, 45) = 6.99, p = 0.011, η2 = 0.13. No category priming effect was observed, p = 0.57, and the category by affect congruence was also non-significant, p = 0.55. Thus, affective priming was observed, such that individuals were faster to respond to targets when primes and targets shared an affective relation compared to when they did not. Table 1 shows relevant means.7
Categorization Task
A 2 (Category Congruence) × 2 (Affective Congruence) repeated measures ANOVA was conducted to examine whether category priming effects would occur for individuals in their natural affective states. A marginal effect for category congruence was observed, F(1, 45) = 3.30, p = 0.076, η2 = 0.068. No effect for affective priming appeared, p = 0.77, and category congruence did not interact with affective congruence, p = 0.41. Thus, tentative evidence was found for category priming during the categorization priming task. See Table 1 for relevant means.7
Lexical Decision Task
Three participants were removed from the analysis for having an accuracy rate near chance (all accuracy means < 54%).
To test the prediction that individuals in their natural affective state would demonstrate category priming, we conducted a 2 (Category Congruence) × 2 (Affective Congruence) repeated measures ANOVA. A main effect for Category Congruence was observed, F(1, 42) = 19.01, p < 0.001, η2 = 0.31. In addition, a marginal main effect for affective priming was found, F(1, 42) = 3.59, p = 0.065, η2 = 0.079 along with a significant interaction between affective congruence and category congruence, F(1, 42) = 12.31, p = 0.001, η2 = 0.23. Table 1 shows relevant means.7
Thus, category priming and affective priming were both observed in the lexical decision priming task. In addition, the category by affective congruence interaction revealed that trials in which primes and targets were not congruent in either dimension produced the slowest target responses.
General Discussion
For three different priming tasks, the results were quite clear; sad moods inhibited both affective and semantic priming effects. On the other hand, both the happy and control groups demonstrated priming across all three tasks. For the evaluative and categorical priming tasks, the happy and control groups demonstrated task-consistent priming effects. However, in the lexical decision task, which does not focus attention on either affective or categorical relations, both groups demonstrated semantic priming and the control group also demonstrated affective priming. Thus, as anticipated, the happy and control groups showed priming effects, whereas sad mood groups failed to show such effects. These findings support assumptions that affect influences the accessibility of semantic associations in memory (Clore & Storbeck, 2006; Isen, 1987; Kuhl, 2000; Smith & DeCoster, 2000).
The sad mood groups turned out to be the most interesting condition. The results suggest that sad moods dampen the activation of semantic associations, whereas happy and control groups maintain such activation, resulting in the same semantic and affective priming effects that are normally found when moods are not induced (Klauer & Musch, 2003; Neely, 1991). Both imaging and behavioral studies find fewer differences between control and happy moods than between control and sad moods (Drevets & Raichle, 1998). This tendency for controls to mimic the results of happy mood groups presumably reflects the fact that most people report quite positive resting moods (Diener & Diener, 1996). We suggest then that sad moods change the default style of processing seen in happy mood groups, which has the effect of eliminating semantic and affective priming.
Mood and Semantic Activation
Priming effects are typically assumed to reflect either spreading activation or response competition. One is an input process (spreading activation), and one is an output process (response competition), but both could plausibly be influenced by mood. In the evaluative and categorical priming tasks, the type of priming observed (affective & categorical) was consistent with the response demands of the respective tasks. These results are therefore compatible with a response competition interpretation. Klauer and Musch (2003) and De Houwer (2003) have suggested that participants are likely to apply the same response rules to primes and targets. As a result, responses should be faster when primes and targets share the same response than when they require different responses. This explanation requires no assumptions about differences in the accessibility of primes and targets in semantic memory. Thus, these findings are compatible with a response competition account as well as with a spreading activation account.
The lexical decision task, on the other hand, requires neither evaluation nor categorization and is therefore not influenced by response compatibility processes. Since the primes were always words, the influence of response compatibility should lead to faster reactions to targets that are words (compatible responses) as opposed to non-words (incompatible responses). However, the obtained category (happy and control groups) and affective (control group) priming effects involve only word trials. Hence, the speed of only one kind of response is involved, and response competition cannot play a role. Spreading activation is, therefore, the more plausible explanation, as traditionally assumed.
This series of results along with other findings in the mood literature (Corson, 2002) suggest that affect influences semantic activation. We find it plausible that response competition factors might also have played a role, especially in Evaluation and Categorization tasks, because these tasks confound the descriptive compatibility of presented stimuli with the motor compatibility of required responses. But, it is possible for spreading activation and response competition to operate simultaneously (Spruyt, De Houwer, Hermans, & Eelen, in press). Therefore, we cannot fully know whether affect influenced one mechanism or the other or both, but there is less ambiguity regarding the lexical decision results.
Prior observations also suggest that affect influences semantic activation (Corson, 2002; Hanze & Hesse, 1993). Specifically, using the Deese-Roediger-McDermott false memory paradigm, Storbeck and Clore (2005) found that affect influenced semantic activation during encoding, but not at the response or retrieval stage. Specifically, they observed false memory effects for both happy and control groups, but not for sad mood groups. It is generally believed that the same semantic activation processes responsible for the Deese-Roediger-McDermott false memory effect also govern semantic priming (Roediger, Balato, & Watson, 2001). Based on the diversity of the priming effects we observed and the fact that they appeared in the happy and control groups and not in sad mood groups, we conclude that negative affect inhibits general levels of semantic activation.
Limitations
Several participants did not report being in the intended mood states. Since an adequate test of the hypothesis required comparing participants differing in mood, data from participants failing to meet that precondition was omitted from the analyses. This procedure is not uncommon in the mood literature (e.g., Bower et al, 1978; Storbeck & Clore, 2005), but it does introduce a nonrandom element into the assignment of subjects to conditions. It should be noted, therefore, that whatever protection against the influence of a third variable that random assignment provides is correspondingly limited.
Another aspect of the data deserving comment is that participants in the sad condition reported moods that appeared more neutral than sad. However, these ratings are not problematic, because people generally report resting moods that are quite positive (Diener & Diener, 1996). Moreover, research shows that people respond to normative or expected affective values, rather than to absolute affective values (Mellers, Schwartz, & Ritov, 1999). Consequently, the affect in the sad mood conditions was functionally quite negative in comparison to the normatively positive reports of most people. Although still lower scores might have yielded different results, we are unaware of any data suggesting such a discontinuity or curvilinearity of affective influence. Indeed, data from our own lab (e.g., Storbeck, 2007) show similar results for mild sad moods (induced with music) and strong sad moods (induced with films).
A final issue concerns whether happy moods actively promote or merely maintain the priming effects typically found in experiments without mood as a factor. A previous study (Corson, 2002) compared happy and sad moods to induced neutral mood and found priming in the happy mood group, but not in the induced neutral and sad mood groups. Those results suggested that happy moods actively promote priming. We compared happy and sad moods to a resting mood control and found priming in happy and resting mood control groups, but not in the sad mood group. It appears that studies that seek to change normatively positive mood to neutral create functionally sad states in which neutrals perform more like those in sad mood conditions. In contrast, in studies using resting mood instead of induced neutral mood, controls perform more like those in happy mood conditions. Therefore, the different conclusions reached from the data of Corson (2002) and our data appear to reflect the different types of control groups employed.
Conclusion
Our findings demonstrate that affect governs semantic and affective priming within a comparative priming paradigm. The experiments suggest that the effects were due to affective influences on semantic activation. Specifically, the relational processing typical of induced and normatively happy mood is assumed to facilitate the accessibility and the use of semantic concepts, whereas sad moods appear to limit the activation and use of such concepts. Response compatibility may also have contributed to some, but not all, of the results. However, the experiments were not designed specifically to isolate the role of response compatibility processes. In summary, our findings are consistent with theoretical assumptions made by Kuhl (2000), Smith and DeCoster (2000), and Ashby et al., (1999), which suggest that happy moods should maintain, and sad moods should dampen, spreading activation.
These results are consistent with those of other recent studies (e.g., Storbeck & Clore, 2005) in suggesting that many of the basic effects from cognitive psychology disappear or are weakened in negative affective states. Given that respondents are usually in positive moods, these results are consistent with the view that cognition and emotion are intertwined phenomena, and that even basic cognitive processes may have an affective trigger.
Acknowledgments
Note: The authors would like to thank Brian Nosek for his helpful comments and insights. This paper was part of a master’s thesis for the first author. Support for this research is acknowledged from National Institute of Mental Health Grant MH 50074 to Gerald L. Clore.
Footnotes
Response compatibility mechanisms are strengthened when the same stimuli serve as primes and targets.
Hypotheses concerned the effect of mood on priming, rather than the effectiveness of inducing moods. Since appropriate comparisons are between individuals for whom the mood manipulation was successful, we included only individuals whose scores were greater than 2 points below the median self-reported mood score in the happy condition and less than 2 points above the median in the sad condition. We also recognize that our experiment loses randomization of participants to conditions and therefore a third variable may be in part responsible for these effects.
To assess whether a speed-accuracy trade-off occurred, we ran a 2 (Category Congruence) × 2 (Affective Congruence) × 2 (Mood) ANOVA on accuracy rates. A significant main effect was found for Category Congruence, F(1, 67) = 4.71, p = 0.033, and a significant interaction was observed for Category by Affective Congruence, F(1, 67) = 4.61, p = 0.035. The category main effect demonstrated that individuals were more accurate with category congruent trials (M = 0.972) than category incongruent trials (M = 0.957). The remaining effects were non-significant.
A 2 (Category Congruence) × 2 (Affective Congruence) × 2 (Mood) ANOVA on accuracy rates. All effects were non-significant‥
A 2 (Category Congruence) × 2 (Affective Congruence) × 2 (Mood) ANOVA on accuracy rates. A marginal main effect for Category Congruence was observed, F(1, 59) = 3.40, p = 0.070. Individuals were less accurate for Category Congruent trials (M = 0.970) compared to Category Incongruent trials (M = 0.977). All other effects were non-significant.
Analyses were run to examine whether order played a role in the observed priming effects. No order effects were apparent for the category and lexical decision task. An order effect was obtained in the evaluation task, such that when the evaluation task was completed first affective priming was observed. However, since the relevant comparison tasks in Experiment 1 also came first (owing to the between groups designs in Exp. 1), the comparisons involving affective priming remain uncompromised.
We ran a 2 (Category Congruence) × 2 (Affective Congruence) repeated measures ANOVA for error rate. Effects for all priming tasks were non-significant.
References
- Ashby FG, Isen AM, Turken AU. A neuropsychological theory of positive affect and its influence on cognition. Psychological Review. 1999;106:529–550. doi: 10.1037/0033-295x.106.3.529. [DOI] [PubMed] [Google Scholar]
- Bargh J. The automaticity of everyday life. In: Wyer RS Jr, editor. The automaticity of everyday life: Advances in social cognition. Vol. 10. Mahwah, NJ: Lawrence Erlbaum Association, Inc.; 1997. pp. 1–61. [Google Scholar]
- Blaney P. Affect and memory: A review. Psychological Bulletin. 1986;99:229–246. [PubMed] [Google Scholar]
- Bolte A, Goschke T, Kuhl J. Emotion and intuition: Effects of positive and negative mood on implicit judgments of semantic coherence. Psychological Science. 2003;14:416–421. doi: 10.1111/1467-9280.01456. [DOI] [PubMed] [Google Scholar]
- Bower GH. Mood and memory. American Psychologist. 1981;36:129–148. doi: 10.1037//0003-066x.36.2.129. [DOI] [PubMed] [Google Scholar]
- Bower G, Monteiro K, Gilligan S. Emotional mood as a context for learning and recall. Journal of Verbal Learning & Verbal Behavior. 1978;17:573–585. [Google Scholar]
- Challis B, Krane R. Mood induction and the priming of semantic memory in a lexical decision task: Asymmetric effects of elation and depression. Bulletin of the Psychonomic Society. 1988;26:309–312. [Google Scholar]
- Clark D, Teasdale J, Broadbent D, Martin M. Effect of mood on lexical decisions. Bulletin of the Psychonomic Society. 1983;21:175–178. [Google Scholar]
- Clore GL, Storbeck J. Affect as information about liking, efficacy, and importance. In: Forgas J, editor. Affect in Social Thinking and Behavior. New York: Psychology Press; 2006. pp. 123–142. [Google Scholar]
- Corson Y. Effects of positive, negative, and neutral moods on associative and semantic priming. Current Psychology of Cognition. 2002;21:33–62. [Google Scholar]
- De Houwer J. A structural analysis of indirect measures of attitudes. In: Musch J, Klauer KC, editors. The psychology of evaluation: Affective processes in cognition and emotion. Mahwah, NJ: Lawrence Erlbaum Associates; 2003. pp. 219–244. [Google Scholar]
- Diener Ed, Diener C. Most people are happy. Psychological Science. 1996;7:181–185. [Google Scholar]
- Drevets W, Raichle M. Reciprocal suppression of regional cerebral blood flow during emotional versus higher cognitive processes: Implications for interactions between emotion and cognition. Cognition & Emotion. 1998;12:353–385. [Google Scholar]
- Fazio RH. A practical guide to the use of response latency in social psychological research. In: Hendrick C, Clark MS, editors. Research methods in personality and social psychology. Newbury Park, CA: Sage; 1990. pp. 74–97. [Google Scholar]
- Forgas J. The Affect Infusion Model (AIM): An integrative theory of mood effects on cognition and judgments. In: Martin L, Clore GL, editors. Theories of Mood and Cognition: A user's guidebook. Mahwah, NJ, US: Lawrence Erlbaum Associates; 2001. pp. 99–134. [Google Scholar]
- Gasper K. Permission to seek freely: The effect of happy and sad moods on generating old and new ideas. Creativity Research Journal. 2004;16:215–229. [Google Scholar]
- Gerrig R, Bower GH. Emotional influences on word recognition. Bulletin of the Psychonomic Society. 1982;19:197–200. [Google Scholar]
- Hanze M, Hesse F. Emotional influences on semantic priming. Cognition and Emotion. 1993;7:195–205. doi: 10.1080/02699939308409184. [DOI] [PubMed] [Google Scholar]
- Hesse F, Spies K. Effects of negative mood on performance: Reduced capacity or changed processing strategy? European Journal of Social Psychology. 1996;26:163–168. [Google Scholar]
- Innes-Ker A, Niedenthal PM. Emotion concepts and emotional states in social judgment and categorization. Journal of Personality & Social Psychology. 2002;83:804–816. [PubMed] [Google Scholar]
- Isen AM. Positive affect, cognitive processes, and social behavior. Advances in Experimental Social Psychology. 1987;20:203–253. [Google Scholar]
- Klauer K, Musch C. Affective Priming: Findings and Theories. In: Musch J, Klauer KC, editors. The psychology of evaluation: Affective processes in cognition and emotion. Mahwah, NJ: Lawrence Erlbaum; 2003. [Google Scholar]
- Kuhl J. A functional-design approach to motivation and self–regulation: The dynamics of personality systems interactions. In: Boekaerts M, Pintrich PR, Zeidner M, editors. Handbook of self–regulation. San Diego: Academic Press; 2000. pp. 105–134. [Google Scholar]
- Matthews G, Wells A. The cognitive science of attention and emotion. In: Dalgleish T, Power M, editors. Handbook of Cognition and Emotion. New York, NY, US: John Wiley & Sons Ltd.; 1999. pp. 171–192. [Google Scholar]
- Mellers B, Schwartz A, Ritov I. Predicting choices from emotions. Journal of Experimental Psychology: General. 1999;128:332–345. [Google Scholar]
- Neely J. Semantic priming effects in visual word recognition: A selective review of current findings and theories. In: Besner D, Humphreys G, editors. Basic processing in reading: visual word recognition. Hillsdale, NJ: Lawrence Erlbaum Association, Inc.; 1991. pp. 264–336. [Google Scholar]
- Roediger H, Balota D, Watson J. Spreading activation and arousal of false memories. In: Roediger H, Nairne J, et al., editors. The nature of remembering: Essays in honor of Robert G. Crowder. Science conference series. Washington, DC: American Psychological Association; 2001. pp. 95–115. [Google Scholar]
- Rolls ET, Stringer S. A model of interaction between mood and memory. Network: Computational Neural Systems. 2001;12:89–109. [PubMed] [Google Scholar]
- Smith ER, DeCoster J. Dual–process models in social and cognitive psychology: conceptual integration and links to underlying memory systems. Personality and Social Psychology Review. 2000;4:108–131. [Google Scholar]
- Spruyt A, De Houwer J, Hermans D, Eelen P. Affective priming of non-affective semantic categorization responses. Experimental Psychology. doi: 10.1027/1618-3169.54.1.44. (in press) [DOI] [PubMed] [Google Scholar]
- Storbeck J. Dissertation. 2007. Affective cues influence cognitive encoding processes: Mood, false memories, and spatial learning. [Google Scholar]
- Storbeck J, Clore GL. With sadness comes accuracy, with happiness, false memory: Mood and the false memory effect. Psychological Science. 2005;16:785–793. doi: 10.1111/j.1467-9280.2005.01615.x. [DOI] [PubMed] [Google Scholar]
- Storbeck J, Robinson MD. Preferences and inferences in encoding visual objects: A systematic comparison of semantic and affective priming. Personality and Social Psychology Bulletin. 2004;30:81–93. doi: 10.1177/0146167203258855. [DOI] [PubMed] [Google Scholar]
- Weaver K, McNeill A. Null effect of mood as a semantic prime. The Journal of General Psychology. 1992;119:295–301. doi: 10.1080/00221309.1992.9917810. [DOI] [PubMed] [Google Scholar]