

Abstract
The cluA gene of Dictyostelium discoideum encodes a novel 150-kDa protein. Disruption of cluA results in clustering of mitochondria near the cell center. This is a striking difference from normal cells, whose mitochondria are dispersed uniformly throughout the cytoplasm. The mutant cell populations also exhibit an increased frequency of multinucleated cells, suggesting an impairment in cytokinesis. Both phenotypes are reversed by transformation of cluA− cells with a plasmid carrying a constitutively expressed cluA gene. The predicted sequence of the cluA gene product is homologous to sequences encoded by open reading frames in the genomes of Saccharomyces cerevisiae and Caenorhabditis elegans, but not to any known protein. The only exception is a short region with some homology to the 42-residue imperfect repeats present in the kinesin light chain, which probably function in protein–protein interaction. These studies identify a new class of proteins that appear to be required for the proper distribution of mitochondria.
Mitochondria generate the ATP that is used to power the activities of eukaryotic cells, ranging from metabolism to motility. Mitochondria are commonly found dispersed throughout the cell, although they may also be concentrated at sites of high energy demand, such as along myofibrils and at the base of sperm tails. There are several indications that mitochondria are actively transported to the appropriate sites in a cell, but the mechanism(s) and regulation of this transport remain unclear. In vertebrate neurons, both microtubules and actin filaments have been implicated in the axonal transport of mitochondria (1). An essential role for microtubules in mitochondrial distribution has been demonstrated in fission yeast (2), and kinesin or kinesin-like motor proteins have been found associated with mitochondria in both vertebrates and flies (3–5). In budding yeast, distribution of mitochondria is defective in the absence of a normal actin cytoskeleton, and an actin-based motor activity is associated with mitochondria (6, 7). Current data leave open the possibility that different cytoskeletal elements may be used for different purposes, such as long distance vs. local transport, or at different stages of the cell cycle. There may also be variation in transport or positioning mechanisms from one organism to another. Current information regarding the division and inheritance of mitochondria has been reviewed (8).
The studies described in this report have identified the first member of a novel class of proteins that appear to be needed for the proper distribution of mitochondria. We have named the gene cluA, for clustered mitochondria. The properties of the cluA gene product, CluA, suggest that it is neither a motor protein nor a structural component of the cytoskeleton. It is a new element in the puzzle of mitochondrial distribution.
MATERIALS AND METHODS
Cells and Cell Culture.
Dictyostelium discoideum strains (AX2, AX3, and DH1) and culture conditions have been described (9).
Purification of the CluA Protein.
AX3 cells in HL5 suspension culture were harvested at a density of 1.5 × 107/ml, washed to remove HL5, and swirled in phosphate buffer (20 mM K2HPO4/KH2PO4, pH 6.4) for 7–10 hr. Initial steps in the purification of CluA were essentially as described (10) for the purification of myosin-IC, an unconventional myosin of Acanthamoeba castellanii. During the purification, the CluA protein was monitored by immunoblot using anti-myosin-IC (11) and/or the 10D5 mAb described below. Dictyostelium cells were homogenized in 2 volumes of a buffer containing 30 mM imidazole-HCl (pH 7.5), 75 mM KCl, 235 mM sucrose, 12 mM sodium pyrophosphate, 5 mM DTT, and protease inhibitors. Unbroken cells were removed by centrifugation. Diisopropylfluorophosphate was added to 1 mM, MgCl2 to 2 mM, and ATP to 0.5 mM (final concentrations), and the cells were swirled for 2 hr. The extract was centrifuged at 125,000 × g for 2 hr at 4°C, and the supernatant was collected. Batch adsorption on DE52 resin (Whatman) was carried out essentially as described (10), except that CluA was step-eluted with 240 mM KCl rather than 150 mM KCl. The eluate was pumped as described (10) onto a phosphocellulose (PC11) column equilibrated with 25 mM Hepes-KOH, pH 7.5/1 mM DTT (buffer H), and eluted with a 0–600 mM KCl gradient; the peak of CluA centered around 300 mM KCl. Positive fractions were pooled, diluted with buffer H to reduce the KCl concentration to 0.1 M, and applied to a Mono S column (Pharmacia) equilibrated with buffer H containing 0.1 M KCl. CluA was eluted with a KCl gradient; positive fractions (between 275 and 400 mM KCl) were pooled. The final step was chromatography on a Mono Q column (Pharmacia) equilibrated with 25 mM Tris-HCl, pH 8.8/0.15 M KCl/1 mM DTT; elution was with a KCl gradient, the CluA peak being at 300–330 mM KCl. The yield from 80–100 g of cells (wet weight) was 200–250 μg of purified protein.
Antibodies and Immunoblotting Methods.
CluA was recognized by a polyclonal rabbit anti-peptide antiserum that had been raised against the phosphorylation domain of Acanthamoeba myosin-IC (11), and by our mAb 10D5. The 10D5 mAb was obtained by immunizing BALB/c mice with partially purified CluA (the Mono S pool) and screening the hybridomas by immunoblot to identify those with the desired specificity. Immunoblots were as described (12) except that 0.1% SDS was included in the transfer buffer. Staining conditions for the polyclonal antiserum were as described (12); for 10D5, ascites fluid was diluted 1:350, and the secondary antibody was peroxidase-conjugated goat anti-mouse IgG (1:1000; American Qualex, La Mirada, CA).
Cloning of the cluA Gene.
Microsequence analysis of tryptic peptides from purified CluA (the Mono Q pool) was performed by W. S. Lane (Harvard Microchemistry Laboratory). Two peptide sequences overlapped, yielding a combined sequence of KSNILTEEQQLEQKQKFEQQQQQQQQTEDKEEKETIATEQQQNKK. Degenerate oligonucleotides were prepared for use as primers in PCR. The oligonucleotides encoded sequences near the ends of the peptide (underlined above); flanking restriction sites were added (PstI for the forward primer and SalI for the reverse primer, italicized below). The forward primer was 5′-AAAAAGTCGAC(T/C)AT(T/C/A)(T/C)TIAC(T/C/A)GA(A/G)GA(A/G)CA(A/G)C-3′ and the reverse primer was 5′-GTTGTTCTGCAGC(T/G/A)AT(T/G/A)GT(T/C)TC(T/C)TT(T/C)TC(T/C)TC-3′. Dictyostelium genomic DNA was used as template. The resulting DNA product was purified using the Wizard PCR Preps DNA Purification Resin (Promega), cut with PstI, and ligated to form dimers. This product was cut with SalI and cloned into pBluescript KS(+); DNA sequences were determined using Sequenase (United States Biochemical). A clone of 120 bp was identified that encoded the starting peptide, with an additional glutamic acid residue: ILTEEQQLEEQKQKFEQQQQQQQQTEDKEEKETIA. The 120-bp DNA fragment, labeled by asymmetric PCR, was used to screen a Dictyostelium cDNA library in λZAP (gift of H. Freeze, The Burnham Institute, La Jolla, CA). Positive clones were identified and recovered as recommended by Stratagene. The largest positive clone, ≈2 kb, was sequenced and found to contain an ORF as well as ≈300 bp of 5′ noncoding region. The remainder of the gene could not be found in the λZAP library. Restriction mapping of genomic DNA predicted that it should be contained within a KpnI/BamHI fragment of ≈3 kb. Accordingly, a minilibrary was prepared by digesting genomic DNA with these two enzymes, separating the restriction products on a 1% low-melt agarose gel (SeaPlaque), recovering DNA fragments of the appropriate size, and cloning the DNA fragments into pBluescript SK(+). Transformants were screened by colony hybridization, and plasmids were recovered from positive colonies and screened by Southern blot analysis to verify that they had the expected restriction pattern. A 2.8-kb positive insert was transferred to pBluescript SK(−) and sequenced as described above. The 2.0-kb cDNA clone and the 2.8-kb genomic clone, which overlapped, were combined in pBluescript SK(+) using a shared restriction site. The resulting plasmid, pDM1, contained a 3.9-kb ORF (designated cluA) with ≈300 nucleotides of flanking sequence at each end.
Disruption of the cluA Gene.
A 2.3-kb MunI/NsiI restriction fragment was excised from the central portion of the cluA coding region in pDM1 and replaced with the 1.9-kb pyr5-6 gene (an EcoRI/PstI fragment, having compatible cohesive ends). The pyr5-6 gene, which confers uracil prototrophy, had first been subcloned as a BamHI/PstI fragment from DIV2 (13) into pBluescript to add the desired restriction sites. The resulting plasmid, termed pDMK, contained 1.2–1.3 kb of cluA DNA on each side of the pyr5-6 gene. The plasmid was cut in the polylinker regions flanking cluA, and 8 × 106 DH1 cells were transformed with 40 μg of the linear DNA by electroporation (13). Transformants were selected in minimal medium in 96-well plates (8 × 105 cells/plate). Viable transformants were transferred to 24-well plates and grown to confluence, then screened as described in Results.
Extrachromosomal Expression of cluA.
For complementation studies, the entire cluA gene was recovered from pDM1 as a PCR product using Deep Vent DNA polymerase (New England Biolabs). The forward primer was 5′-GTTCAGATCTATGTCAGAAACTATTGATAACC-3′, which created a BglII restriction site immediately in front of the ATG translation initiation codon, and the reverse primer was 5′-ATGCATTGAAGAGTCGGCAGCGGA-3′, which created an NsiI site within a few codons of the 3′ end of the gene. The PCR product was cloned into the EcoRV site of pBluescript SK(−), yielding the plasmid pDM10. A ClaI/XhoI fragment (containing the 3′ half of cluA) was removed from pDM10 and replaced with the corresponding ClaI/XhoI fragment from pDM1 to obtain a different set of 3′ flanking restriction sites, yielding plasmid pDM11. A BglII/PstI fragment (containing the entire cluA gene and its 3′ flanking region) was excised from pDM11 and cloned into the Dictyostelium expression plasmid pDXA-3H (14) cut with BamHI and NsiI. The resulting plasmid (pDM2) was transformed into a cluA− mutant, and transformants were selected in HL5 containing 10 μg/ml G418. Molecular biology procedures were as described (9, 15).
RESULTS
Purification of the CluA Protein.
The 150-kDa CluA polypeptide was first detected in Dictyostelium (12) by virtue of its cross-reactivity with an anti-peptide antiserum specific for an unconventional myosin of Acanthamoeba castellanii, myosin-IC (11). The CluA protein was purified from Dictyostelium cells using the anti-myosin-IC antiserum to monitor its fractionation behavior. Its fractionation properties were similar to those of myosin-IC (10) during the initial steps, but differed as the purification progressed. The final product was a single polypeptide for which we could detect neither myosin-like enzymatic activity nor associated light chains, although cross-reactivity with the anti-myosin-IC antiserum persisted. The purification of CluA is summarized in Fig. 1.
Figure 1.

Purification and immunoreactivity of CluA. (A) SDS gel (Coomassie-stained) showing fractions from the purification of CluA. Lanes: 1, total lysate; 2, high speed supernatant; 3, DE52 eluate; 4, PC11 pool; 5, Mono S pool; 6, Mono Q pool. Lanes 1–3 received 17 μg of protein, and lanes 4–6, 0.5 μg. (B) Immunoblot of the Mono S and Mono Q pools (0.5 μg each, identical to lanes 5 and 6 in A) stained with anti-myosin-IC or with 10D5, as indicated. (C) Specificity of 10D5 mAb. Lane 1, total lysate from 1 × 106 cells; lane 2, immunoblot of an identical sample stained with 10D5 antibodies. Prestained molecular weight markers (Bio-Rad) are indicated for each gel (myosin, 213,000; β-galactosidase, 119,000; bovine serum albumin, 83,000; ovalbumin, 47,000).
Cloning and Analysis of the cluA Gene.
The partial amino acid sequences of tryptic peptides from purified CluA were determined, and data from two overlapping peptides allowed us to generate a PCR product that was used to isolate a partial cDNA. The remainder of the gene was obtained from a genomic minilibrary (see Materials and Methods for details). The gene corresponding to the 3.9-kb ORF was designated cluA. The nucleotide sequence of cluA is shown in Fig. 2, together with the predicted amino acid sequence of its protein product.
Figure 2.

Nucleotide sequence of cluA and amino acid sequence of its predicted protein product.
Southern blot analysis of genomic DNA suggested that cluA is single copy. Northern blots showed a single RNA band of ≈4 kb, which was present in vegetative cells and increased to higher levels in developing cells, peaking at 8–12 hr (data not shown).
The Protein Encoded by cluA.
The cluA gene encoded a protein of 1,321 residues with a calculated molecular weight of 148,916 and an isoelectric point of 6.1. The predicted amino acid sequence showed no significant homology to myosins or to any other characterized proteins, except as noted below. However, a search of the GenBank database revealed that homologous proteins are encoded by ORFs in the genomes of Saccharomyces cerevisiae and Caenorhabditis elegans. The alignment of the proteins encoded by cluA and these ORFs is shown schematically in Fig. 3. The fraction of identical residues between each pair of proteins is 27–30%, and the overall similarity among them is ≈50%. Near the C terminus of Dictyostelium CluA and its homologues is a region with some homology to a series of 42-residue imperfect repeats found in the kinesin light chain (see Discussion).
Figure 3.

Alignment of the predicted protein sequences encoded by cluA and its homologues in S. cerevisiae and C. elegans. The S. cerevisiae ORF lies on chromosome XIII, locus YM8270.16 (Z48613), and the C. elegans ORF lies on chromosome III, locus F55H2.6 (P34466). In this schematic MACAW (multiple alignment construction and analysis workbench) alignment (16), the greater bar widths indicate regions of homology, and the vertical black lines mark amino acids whose conservation is especially significant.
Disruption of the cluA Gene.
The Dictyostelium cluA gene was disrupted by homologous recombination, replacing the central half of the coding region with a selectable marker (Fig. 4A). An internal 2.3-kb fragment was removed from the cloned cluA gene and replaced with the 1.9-kb pyr5-6 gene, which functions in uracil biosynthesis. The linearized cluA construct was transformed into DH1, a uracil auxotroph, and transformants were selected in minimal medium lacking uracil. Transformants were screened by immunoblot to identify those lacking the CluA antigen; about one-quarter of the transformants contained no detectable CluA. Several of these negative strains were further screened by Southern blot analysis to identify those in which a double crossover had occurred. Restriction patterns suggestive of single-crossover events were more common (data not shown), but 3 of the first 12 strains we examined had the pattern predicted for a double crossover (Fig. 4B). The phenotypic properties described below were common to both types of gene disruptants, but the mutants created by double crossover were expected to be, and proved to be, completely stable even when grown under nonselective conditions. Therefore, further characterization was restricted to these strains.
Figure 4.

Disruption of cluA. (A) A double crossover between the disruption plasmid (Top) and the chromosomal gene (Middle) would lead to the substitution of the selectable marker pyr5-6 for the central portion of cluA (Bottom). A consequence of this event would be a change in the size of the indicated HindIII fragment from 2.9 to 2.5 kb. (B) Southern blot of one of the cluA− transformants with this restriction pattern. The probe was a 1.2-kb PvuII fragment (as indicated below the gene in A).
The growth rate of the cluA− mutants was somewhat slower than that of the parental cells in axenic suspension culture (16 hr vs. 12 hr doubling time). Even when grown on a surface, about half of the cells in the mutant populations were multinucleated, suggesting an impairment in cytokinesis. The mutant cells took one day longer than control cells to clear a bacterial lawn, but they proceeded through development to form fruiting bodies that contained viable spores.
The most dramatic consequence of the lack of CluA became apparent when cluA− cells were stained with 4′,6-diamidino-2-phenylindole (DAPI), a DNA-specific dye that stains both nuclei and mitochondria. Mitochondria are normally dispersed uniformly throughout the cytoplasm of Dictyostelium cells (Fig. 5C), but in cluA− cells, all of the mitochondria were clustered near the cell center (Fig. 5 A and B). This was true for all cells in the mutant population, although there was some cell-to-cell variability in how tightly the mitochondria were clustered.
Figure 5.
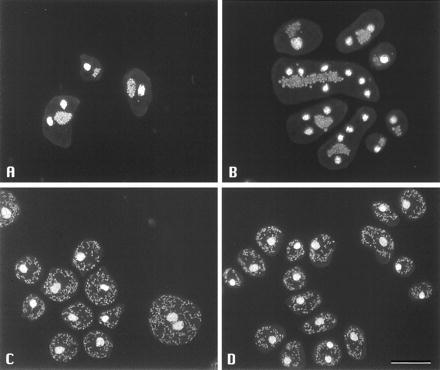
Dictyostelium cells stained with DAPI to visualize nuclei and mitochondria. (A and B) cluA− cells. Mitochondria are clustered near the cell center. (C) Normal cells (DH1). Note scattered mitochondria. (D) Mutant cluA− cells transformed with pDM2, showing complementation of mutant phenotype by constitutive expression of cluA. (Bar = 20 μm.)
The cluster of mitochondria could be observed in living cluA− cells by phase contrast microscopy, indicating that it was not a consequence of fixation. Furthermore, observation of living cells revealed that the clusters changed shape readily as the cells moved about, and that contractile vacuoles occasionally filled and emptied in their midst. Thus, the mitochondria were not tightly associated with one another.
Complementation of cluA− Cells.
To verify that the two phenotypes of cluA− cells (mitochondrial clustering and defective cytokinesis) were directly attributable to the lack of CluA, complementation experiments were performed. The cluA gene was subcloned from pDM1 into a Dictyostelium expression vector in which expression of cluA was controlled by a strong constitutive promoter, act15. This vector, pDXA-3H, is one of the series created by Manstein et al. (14); it is replicated extrachromosomally and carries the neomycin phosphotransferase gene, allowing G418 selection. The pDXA derivative containing cluA was termed pDM2.
The cluA− cells were transformed with pDM2, and transformants were selected with G418. Most of the transformant colonies consisted of cells that were small and uniform in size (Fig. 6B), an appearance easily distinguished from that of the cluA− parental cells (Fig. 6A). Examination of these transformants by DAPI staining confirmed that such colonies consisted predominantly of mononucleated cells, and that mitochondrial distribution had been restored to normal (Fig. 5D). Quantitation in one experiment showed that 45% of the cluA− parental cells growing on glass coverslips were multinucleate, with 55% of all nuclei being found in cells containing four or more nuclei. All of the cluA− cells had clustered mitochondria. In a representative pDM2 transformant under the same conditions, 90% of the cells were mononucleate, and fewer than 1% contained more than two nuclei. In 93% of the cells, the mitochondria were normally dispersed. Thus, both phenotypes associated with the disruption of cluA were complemented by extrachromosomal expression of cluA. Immunoblots indicated that the pDM2 transformants contained levels of CluA similar to or greater than those found in control cells.
Figure 6.

The effect of cluA on cytokinesis. (A) A cluA− cell population contains many multinucleated cells, as indicated by their unusually large size. (B) Transformation of cluA− cells with a plasmid expressing cluA restores normal cell size. The cells were growing in HL5 on tissue culture plates and were viewed with Hoffman modulation contrast optics. (Bar = 20 μm.)
DISCUSSION
The studies presented here demonstrate that a novel protein, the product of the cluA gene, is required for normal mitochondrial distribution in Dictyostelium. In the absence of CluA, all of the mitochondria in the cell are found in a single cluster near the cell center. Probably owing to the small size of Dictyostelium cells, this mutation has only a modest effect on growth rate; in a cell that is only 10 μm in diameter, diffusion may be an adequate mechanism to spread ATP throughout the cytoplasm.
Cytokinesis is partially impaired in cluA− cells, resulting in a population in which a majority of the cells are multinucleated. The impairment of cytokinesis may be due to a physical obstruction presented to the cleavage furrow by the mislocalized mitochondria, or it may reflect some as-yet-unrecognized aspect of the function of CluA. The production of occasional cells devoid of mitochondria no doubt contributes to the slower growth rate of the cluA− mutants.
The clustering of mitochondria could occur because these organelles adhere to one another. However, if they do adhere, observation of living cells indicates that their contacts must be weak and readily reversible. In cluA− cells the mitochondria redistribute themselves readily as the cells change shape and move about, and they become somewhat more scattered in cells undergoing mitosis. Furthermore, thin section electron microscopy of cryofixed amoebae does not show any evidence of adhesion between membranes of contiguous mitochondria (S. D. Fields and M.C., unpublished data).
The central position of mitochondria could arise from active transport to this location or, possibly, from passive accumulation. Studies in mammalian cells have shown that small tracer particles are excluded from regions of the cytoplasm that are especially rich in cytoskeletal filaments, suggesting that a molecular sieving mechanism affects the distribution of such particles and also of membrane-bound organelles such as mitochondria (17). Dictyostelium cells are highly motile, undergoing rapid and extensive rearrangements of the cytoskeleton as they move about. Thus, inert particles might collect near the nucleus, where the height of the cell is greatest and the frequency of cytoskeletal rearrangements is likely to be lower.
If correct, this interpretation would imply that the mitochondria in cluA− cells lack an interaction with the cytoskeleton that is required to produce the uniform mitochondrial distribution characteristic of normal cells. Preliminary data suggest that the cytoskeleton itself is not altered in mutant cells. It does not seem likely that CluA is a motor responsible for mitochondrial transport, since it lacks any homology to motor domains of the known actin- or microtubule-based motors. It contains no obvious calmodulin binding motifs, and it lacks a consensus sequence for the phosphate binding domain (“P-loop”) that is typically responsible for ATP/GTP binding in motor proteins (18). Thus, a motor function seems improbable, although it cannot yet be excluded.
It is possible that CluA may act as a linker between motor molecules and mitochondria. A region near the C terminus of CluA has detectable homology to a series of tandem 42-residue imperfect repeats found in the kinesin light chain, which are structurally related to 34-residue repeats (“TPR motifs”) that function in protein–protein association (19). The tandem repeat region of the kinesin light chain is important for the interaction of kinesin with membrane vesicles (20). Thus, the homologous region in CluA may be a locus through which CluA interacts with a target protein, possibly even the kinesin light chain.
The CluA polypeptide was first detected on immunoblots using a polyclonal antiserum that labeled contractile vacuole and plasma membranes (11, 12). However, as described here (and also unpublished data), CluA fractionates as a soluble protein without obvious enrichment in membranes or mitochondrial fractions. The availability of cluA− cells should have made it possible to determine whether membrane staining persisted in cells lacking CluA, but the original polyclonal antiserum was exhausted long ago. Indeed, although this antiserum recognized purified CluA, as shown in Fig. 1, the basis for that recognition is not clear; the predicted amino acid sequence of CluA contains no good match to the 25-residue Acanthamoeba myosin-IC peptide that was used to generate the antiserum. Since our 10D5 mAb has not proved useful for immunolocalization studies, new antibodies are being raised for this purpose.
Because proteins homologous to the Dictyostelium CluA protein are encoded by S. cerevisiae and C. elegans (and expressed sequence tag databases contain expressed partial sequences for C. elegans, plants, and mammals), CluA appears to be the first recognized member of a new class of proteins involved in the transport or positioning of mitochondria. We have disrupted the cluA homologue in S. cerevisiae (CLU1) and found that the consequences of the null mutation in yeast cells are similar to those in Dictyostelium. The lack of the gene product is not lethal, but in mutant cells the mitochondrial chondriome is clumped in a single mass rather than spread as a reticulum (S. D. Fields, M. N. Conrad, and M.C., unpublished data). Thus, the function of CluA appears to be conserved in at least two organisms with quite different life styles and mechanisms of cell reproduction.
We speculate that the consequences of the lack of a CluA homologue might be more deleterious in a multicellular organism. Larger cell size and the pronounced asymmetry of cells such as neurons could make proper mitochondrial distribution a necessity rather than just an advantage. Similarly, the high energy demands of certain types of tissues and cells require the concentration of mitochondria at specific sites. A deficiency in this targeting ability could have serious consequences. Examining the role of CluA in multicellular organisms is therefore a high priority.
Acknowledgments
We thank Gary Farrell for his assistance in the production of mAbs. This work was supported by grants from the National Institutes of Health (GM29723), the National Science Foundation (MCB-9317286), and the Oklahoma Center for the Advancement of Science and Technology (HR4-055) to M.C.
ABBREVIATIONS
- DAPI
4′,6-diamidino-2-phenylindole
- myosin-IC
an unconventional myosin of Acanthamoeba castellanii
Footnotes
References
- 1.Morris R L, Hollenbeck P J. J Cell Biol. 1995;131:1315–1326. doi: 10.1083/jcb.131.5.1315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Yaffe M P, Harata D, Verde F, Eddison M, Toda T, Nurse P. Proc Natl Acad Sci USA. 1996;93:11664–11668. doi: 10.1073/pnas.93.21.11664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Nangaku M, Sato-Yoshitake R, Okada Y, Noda Y, Takemura R, Yamazaki H, Hirokawa N. Cell. 1994;79:1209–1220. doi: 10.1016/0092-8674(94)90012-4. [DOI] [PubMed] [Google Scholar]
- 4.Elluru R G, Bloom G S, Brady S T. Mol Biol Cell. 1995;6:21–40. doi: 10.1091/mbc.6.1.21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Pereira A J, Dalby B, Stewart R J, Doxsey S J, Goldstein L S B. J Cell Biol. 1997;136:1081–1090. doi: 10.1083/jcb.136.5.1081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Lazzarino D A, Boldogh I, Smith M G, Rosand J, Pon L A. Mol Biol Cell. 1994;5:807–818. doi: 10.1091/mbc.5.7.807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Simon V R, Swayne T C, Pon L A. J Cell Biol. 1995;130:345–354. doi: 10.1083/jcb.130.2.345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Yaffe M P. Adv Mol Cell Biol. 1996;17:341–350. [Google Scholar]
- 9.Liu T, Clarke M. J Cell Sci. 1996;109:1041–1051. doi: 10.1242/jcs.109.5.1041. [DOI] [PubMed] [Google Scholar]
- 10.Lynch T J, Brzeska H, Miyata H, Korn E D. J Biol Chem. 1989;264:19333–19339. [PubMed] [Google Scholar]
- 11.Baines I C, Korn E D. J Cell Biol. 1990;111:1895–1904. doi: 10.1083/jcb.111.5.1895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Zhu Q, Clarke M. J Cell Biol. 1992;118:347–358. doi: 10.1083/jcb.118.2.347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kuspa A, Loomis W F. Methods Mol Genet. 1994;3:3–21. [Google Scholar]
- 14.Manstein D J, Schuster H-P, Morandini P, Hunt D M. Gene. 1995;162:129–134. doi: 10.1016/0378-1119(95)00351-6. [DOI] [PubMed] [Google Scholar]
- 15.Sambrook J, Fritsch E F, Maniatis T. Molecular Cloning: A Laboratory Manual. 2nd Ed. Plainview, NY: Cold Spring Harbor Lab. Press; 1989. [Google Scholar]
- 16.Schuler G D, Atschul S F, Lipman D J. Protein Struct Funct Genet. 1991;9:180–190. [Google Scholar]
- 17.Provance D W, McDowall A, Marko M, Luby-Phelps K. J Cell Sci. 1993;106:565–578. doi: 10.1242/jcs.106.2.565. [DOI] [PubMed] [Google Scholar]
- 18.Saraste M, Sibbald P R, Wittinghofer A. Trends Biochem Sci. 1990;15:430–434. doi: 10.1016/0968-0004(90)90281-f. [DOI] [PubMed] [Google Scholar]
- 19.Gindhart J G, Goldstein L S B. Trends Biochem Sci. 1996;21:52–53. [PubMed] [Google Scholar]
- 20.Stenoien D L, Brady S T. Mol Biol Cell. 1997;8:675–689. doi: 10.1091/mbc.8.4.675. [DOI] [PMC free article] [PubMed] [Google Scholar]