

Summary
Kar3, a Saccharomyces cerevisiae Kinesin-14, is essential for karyogamy and meiosis I but also has specific functions during vegetative growth [1–7]. For its various roles, Kar3 forms a heterodimer with either Cik1 or Vik1, both of which are noncatalytic polypeptides [8–11]. Here, we present the first biochemical characterization of Kar3Cik1, the kinesin motor that is essential for karyogamy [8–11]. Kar3Cik1 depolymerizes microtubules from the plus end and promotes robust minus-end-directed microtubule gliding. Immunolocalization studies show that Kar3Cik1 binds preferentially to one end of the microtubule, whereas the Kar3 motor domain, in the absence of Cik1, exhibits significantly higher microtubule lattice binding. Kar3Cik1-promoted microtubule depolymerization requires ATP turnover, and the kinetics fit a single exponential function. The disassembly mechanism is not microtubule catastrophe like that induced by the MCAK Kinesin-13s [12–18]. Soluble tubulin does not activate the ATPase activity of Kar3Cik1, and there is no evidence of Kar3Cik1•tubulin complex formation as observed for MCAK [12, 13, 15, 16, 18]. These results reveal a novel mechanism to regulate microtubule depolymerization. We propose that Cik1 targets Kar3 to the microtubule plus end. Kar3Cik1 then uses its minus-end-directed force to depolymerize microtubules from the plus end, with each tubulin-subunit release event tightly coupled to one ATP turnover.
Results
Kar3 is one of six kinesin-related genes in budding yeast [1, 11, 19]. Like Drosophila Ncd, Kar3 is classified as a Kinesin-14 because its motor domain is at the carboxy terminus, and it promotes minus-end-directed microtubule (MT) gliding in vitro [1, 2, 20]. However, Kar3 is the only Kinesin-14 in S. cerevisiae, and there are specific roles for Kar3 motor activity in both the cytoplasm and the nucleus. The cellular localization is modulated in part by Cik1 and Vik1 to either the cytoplasm or nucleus [8–11]. During conjugation in response to mating pheromone, Cik1 targets Kar3 to astral MTs and to the spindle pole bodies [8–11]. Mating cells that lack either Kar3 or Cik1 fail to interdigitate their MTs and pull nuclei together after cell fusion [1, 8–10] (Figure 1). During vegetative growth, Kar3 regulates MT dynamics that affect both spindle size and spindle position [3, 21]. Because both Kar3Cik1 and Kar3Vik1 function during vegetative growth [7–11, 22], it is difficult to distinguish Kar3 functions driven by Kar3Vik1 from those due to Kar3Cik1.
Figure 1. Model for Kar3Cik1 Function during Karyogamy.
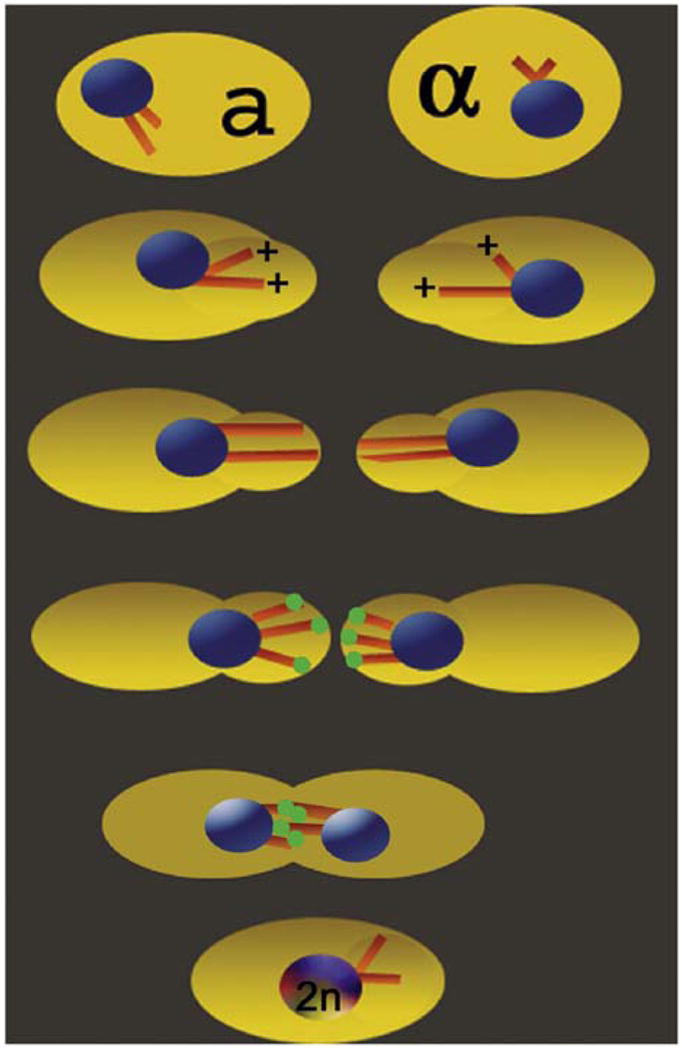
(1) Mating-specific pheromones induce Kar3 and Cik1 expression as well as the formation of the rounded cell protrusions or “shmoos.” (2 and 3) The astral MTs are highly dynamic, and the MT plus ends become associated with the cortex. (4) For MT shortening, Kar3Cik1 motors (green spheres) are targeted to the MT plus ends. Kar3Cik1-promoted MT depolymerization shortens the MTs to pull each nucleus into its shmoo. (5) Localized cell-wall breakdown allows cytoplasm fusion, resulting in interdigitating MTs of opposite polarity. (6) Kar3Cik1 can now use its minus-end-directed force for both MT sliding and MT shortening to form the diploid nucleus.
Initial in vitro studies with GST-Kar3 revealed that this C-terminal kinesin promoted minus-end-directed MT gliding but also MT shortening from the minus end [20]. Historically, it was thought that Kar3 motored to the minus end of the MT and then switched its catalytic activity to MT depolymerization [3, 20, 23]. Subsequent studies with green fluorescent protein fused to α-tubulin (GFP-tubulin) showed that the MT dynamics in yeast appear to occur only at the MT plus ends [24, 25]. Recently, Maddox et al. reported that during mating, GFP-Kar3 couples the MT plus ends to the cortical shmoo tip during MT depolymerization and that the Bim1-Kar9 complex maintains attachment of the MT plus ends during MT polymerization [26]. Bim1 is the EB1 homolog (MT plus-end tip protein), and Kar9 is the MT linker-cortex attachment protein [27, 28].
Budding yeast is an excellent model system to dissect the roles of kinesin-motor-associated proteins for regulation of motor activity and microtubule dynamics. Because Kar3Cik1 is essential for karyogamy and Kar3-Vik1 is not involved [10], our questions are framed around the role of Kar3Cik1 for karyogamy (Figure 1). At least three hypotheses can account for the observations in the literature: (1) After cell fusion, MTs interdigitate, and Kar3Cik1 slides MTs in relation to one another to bring the nuclei together. (2) MTs interdigitate, but only Kar3Cik1-promoted MT depolymerization is required. (3) Nuclear fusion requires both Kar3Cik1-promoted MT sliding and MT depolymerization.
For our in vitro studies, we have addressed the role of Cik1 for Kar3 mechanochemistry. We propose that Cik1 may modulate the catalytic activity of the Kar3 motor domain comparably to a myosin light chain, Cik1 may modulate the Kar3 interaction with the MT directly or indirectly, and/or Cik1 may target Kar3 to the MT plus end. We coexpressed truncated genes of Kar3 and Cik1 (Figure 2). The Kar3 motor domain (Kar3MD) [29] and dimeric D. melanogaster Ncd MC1 [30, 31] were comparative C-terminal kinesins. The Coomassie-stained gel (Figure 2B) shows that the expressed Kar3Cik1 proteins purified with an apparent stoichiometry of one Kar3 per Cik1, and the analytical gel-filtration analysis (Figure 2C) reveals that Kar3Cik1 migrates ahead of dimeric kinesin K401 [32, 33]. These data indicate that the truncated Kar3Cik1 motor is a stable heterodimer.
Figure 2. Kar3Cik1 Characterization.

(A) The three motors used: monomeric Kar3MD, Kar3Cik1 heterodimer, and dimeric D. melanogaster Ncd MC1.
(B) Coomassie-stained gel (2 M Urea, 8% SDS-PAGE) and Western blot showing protein purity and antibody specificity.
(C) Analytical gel filtration of Kar3Cik1, dimeric conventional kinesin K401, and Kar3MD with Mr based on amino acid sequence. Stokes radius of Kar3Cik1 = 4.0 nm, K401 = 3.8 nm, and Kar3MD = 3.4 nm.
(D) Kar3Cik1 minus-end-directed MT motility in the presence of MgATP. Arrowheads denote the bright MT minus end, and the asterisks, the dim MT plus end. The scale bar represents 5 μm.
(E–G) The steady-state ATPase kinetics of Kar3Cik1 as a function of MTs (E) and MgATP (F and G). Both (E) and (F) were performed at 25 μM Taxol, at which the MTs were stable and resistant to Kar3Cik1-promoted depolymerization. (E) Final concentrations: 1 μM Kar3Cik1, 1–40 μM tubulin, 25 μM Taxol, and 1 mM Mg[α32P]ATP. Steady-state parameters: kcat = 0.35 ± 0.02 s−1; K1/2,Mt = 6.3 ± 1.3 μM. Inset: Kar3Cik1 was incubated with increasing concentrations of tubulin as MTs (black), soluble tubulin heterodimer (red), or in the absence of tubulin (green circle), and the reactions were initiated with MgATP. Final concentrations: 0.5 μM Kar3Cik1, 0–3 μM tubulin, ± 3 μM Taxol, and 100 μM Mg[α32P]ATP. The ATPase rate in the absence of tubulin (green) was 0.014 ± 0.006 s−1; soluble tubulin (red) = 0.015 ± 0.002 s−1. (F) Steady-state ATPase at stable MT conditions. Final concentrations: 1 μM Kar3Cik1, 40 μM tubulin, 25 μM Taxol, and 1–700 μM Mg[α32P]ATP. Kar3Cik1: kcat = 0.37 ± 0.005 s−1; Km,ATP = 8.2 ± 0.53 μM. Inset: Initial phase at 1–200 μM MgATP. (G) The steady-state ATPase at MT depolymerizing conditions. Final concentrations: 1 μM Kar3Cik1, 40 μM tubulin, 15 μM Taxol, and 1 mM Mg[α32P]ATP. Kar3Cik1: kcat = 0.26 ± 0.017 s−1; Km, ATP = 71.2 ± 16.2 μM. The table shows the steady-state parameters of Kar3Cik1 in comparison to Kar3MD [29], and dimeric Ncd MC1 [30]. “ND” denotes “not determined.”
Kar3Cik1 Promotes Robust Microtubule Gliding
We used both polarity-marked and unmarked rhodamine-labeled 20 μM Taxol-stabilized MTs to show that Kar3Cik1, like GST-Kar3 [20], was able to promote MT gliding in vitro (see Movie S1 in the Supplemental Data available with this article online). Figure 2D illustrates one example of a polarity-marked MT that moved with its plus end leading, indicating minus-end-directed motion. The polarity of the MTs was confirmed with conventional kinesin, a well-characterized plus-end-directed motor. The rate of Kar3Cik1 MT gliding was 2.40 ± 0.06 μm/min. This rate was faster than the 1.3 ± 0.1 μm/min reported previously for GST-Kar3 [20].
Kar3Cik1-Promoted Microtubule Depolymerization Requires ATP Turnover
The phenotypes of the kar3Δstrain and kar3-1 rigor mutant suggest that Kar3 motor function acts to limit both MT length and number during vegetative growth and karyogamy [1, 3, 26]. To analyze the Kar3Cik1 depolymerase activity, we developed both a solution assay (Figure 3) and a real-time microscopy assay (Figure 4). The MT•Kar3Cik1 complex was preformed with different MT-stabilized substrates, and MT depolymerization was initiated by MgATP (Figure 3). The results for Kar3Cik1 were compared to the kinetic profile of MCAK depolymerases [12–18, 34]. Kar3Cik1 was unable to depolymerize 20 μM Taxol-stabilized or 1 mM MgGMPcPP-stabilized MTs (Figure 3A). Both can be rapidly destabilized by MCAK [12]. Kar3Cik1 was able to depolymerize MTs stabilized with 3 μM Taxol (Figures 3B and 3C). In the absence of Kar3Cik1, the 3 μM Taxol-stabilized MTs showed only trace amounts of soluble tubulin in the supernatant. Upon addition of Kar3Cik1 plus MgATP, there was an exponential increase in tubulin partitioning to the supernatant as a function of time (kobs = 0.07 s−1). Experiments were performed with 1 mM MgAMPPNP to ask whether ATP binding was sufficient to induce Kar3Cik1-promoted depolymerization, as observed for MCAK [12, 14, 15, 17, 18]. No evidence of MT depolymerization by Kar3Cik1 with MgAMPPNP was observed (Figure 3A). These results demonstrate that Kar3Cik1-promoted MT depolymerization requires ATP turnover. In contrast to MCAK, ATP binding is not sufficient for Kar3Cik1 to destabilize MTs. The kinetics of MCAK-promoted depolymerization are sigmoid, which is characteristic of a MT catastrophe mechanism [15, 35]. However, the Kar3Cik1-promoted MT depolymerization kinetics best fit a single exponential function (Figure 3C). These data indicate that Kar3Cik1 does not promote MT catastrophe. Rather, the results suggest a mechanism of sequential release of tubulin subunits from the MT polymer, with each release event tightly coupled to one ATP turnover.
Figure 3. Solution Assays of Microtubule Depolymerization.
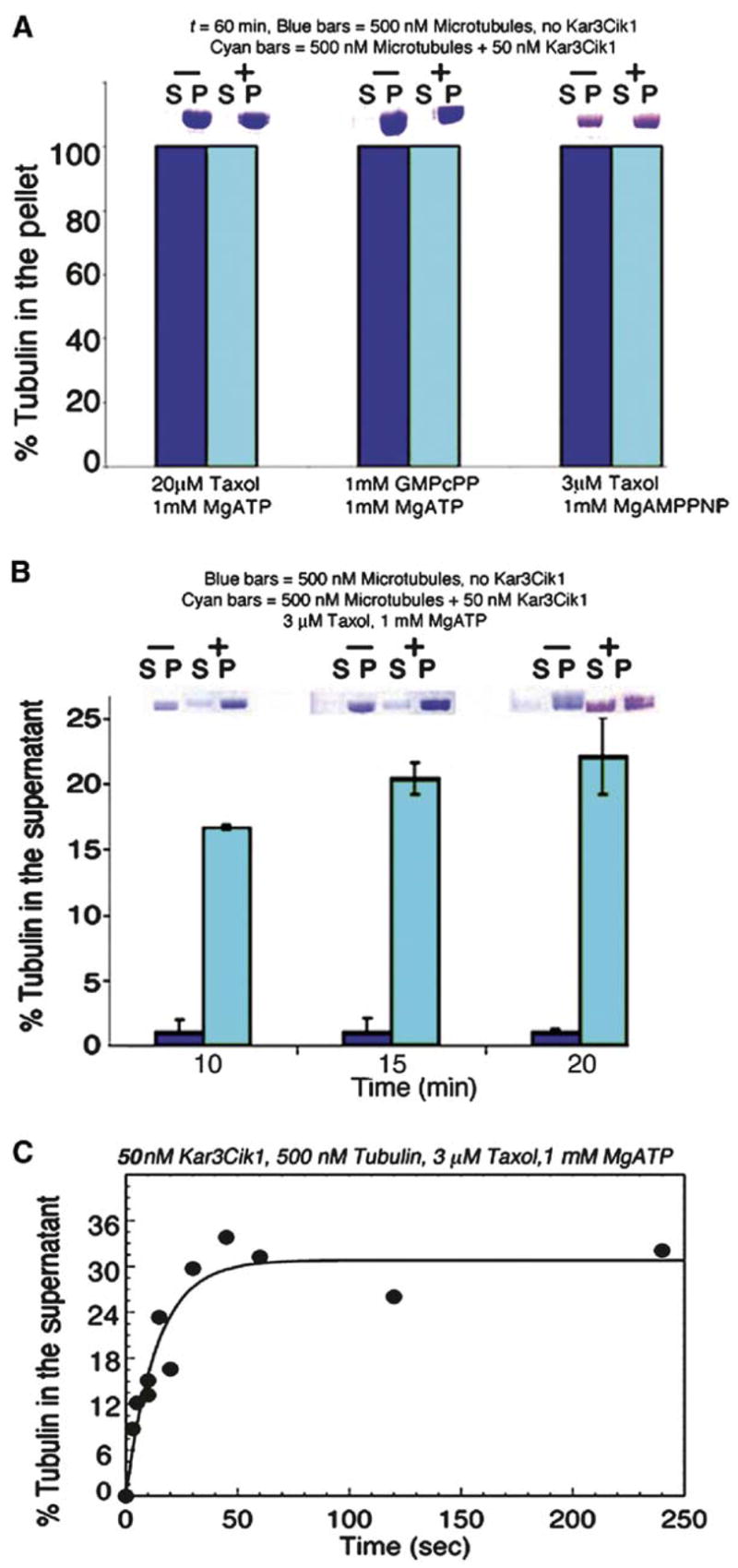
(A and B) Representative gel slices above each set of bars show tubulin partitioning to either the supernatant (S) or to the MT pellet (P). Each bar is the mean of three replicates, and the error bars report the SEM. MTs in the presence (cyan bars) or absence (blue bars) of Kar3Cik1 were incubated with different nucleotides and MT stabilizing agents to determine the substrates for Kar3Cik1 depolymerization. Final concentrations: ± 50 nM Kar3Cik1, 500 nM tubulin, 3 or 20 μM Taxol or 1 mM MgGMPcPP, and 1 mM MgATP or 1 mM MgAMPPNP. (B and C) MTs stabilized with 3 μM Taxol were incubated with Kar3Cik1 in the presence of 1 mM MgATP. The reactions were terminated at specific times with 2 mM MgAMPPNP, and then centrifugation and analysis by SDS-PAGE were performed. The percent of tubulin partitioning to the supernatant was plotted as a function of time, and the data were fit to a single exponential function with kobs = 0.07 ± 0.01 s−1
Figure 4. Microtubule Shortening Promoted by Kar3Cik1, Ncd, and MCAK.

MT•motor complexes were formed in the presence of MgAMPPNP and imaged at t = 0. MT depolymerization was initiated by MgATP plus an ATP regeneration system.
(C, F, and I) Merge of times 0 and 60 min to show MT shortening (yellow) in comparison to the original length (red).
(C) The MTs exhibited shortening from only one end in the presence of Kar3Cik1 plus 1.5 mM MgATP. The scale bar represents 5 μm.
(F) MT shortening from the plus end (*) of the MT in the presence of Kar3Cik1 and 1.5 mM MgATP. The scale bar represents 5 μm. (D)–(R) are at the same magnification.
(G–I) MT shortening from the plus end (*) of MTs in the presence of dimeric Ncd and 1.5 mM MgATP; the MT seed (arrowhead) denotes the MT minus end.
(J–O) MT shortening from both ends promoted by Xenopus MCAK.
(P–R) Conventional kinesin K401 did not promote MT shortening. The MT shortening rate is reported as nm/min ± the SEM.
Kar3Cik1 Steady-State ATPase Is Only Stimulated by Microtubules
We examined the ATPase properties of Kar3Cik1 to compare with Kar3MD [29], dimeric Ncd [30, 31], and MCAK [15, 16]. Traditionally, kinesins in the absence of MTs exhibit a very low ATPase activity, and MTs greatly enhance this rate. This behavior was exhibited by Kar3-Cik1 (Figures 2E–2G). For MCAK, soluble tubulin subunits also activate steady-state ATP turnover, and MCAK can form a stable complex with a tubulin heterodimer in vitro [12–18]. These observations for MCAK have led to a proposed mechanism in which the MCAK•tubulin complex detaches from the MT, and ATP turnover is used to liberate the motor from the tubulin heterodimer [14–18]. We pursued experiments to test this type of mechanism for Kar3Cik1. In the absence of MTs, the Kar3Cik1 steady-state ATPase was 0.014 s−1 (Figure 2E, inset, green circle). When Kar3Cik1 was added to soluble tubulin, there was no activation of this rate by soluble tubulin (Figure 2E, inset, red triangles), yet there was dramatic activation of the steady-state ATPase when MTs were used at the same tubulin concentrations (Figure 2E, inset, black circles). In addition, there was no evidence of complex formation between soluble tubulin and Kar3Cik1 when these were analyzed by gel filtration (data not shown). Our results indicate that Kar3Cik1 does not detach from the MT in association with a tubulin heterodimer.
The Kar3Cik1 steady-state ATPase was activated by stable MTs (25 μM Taxol) and MTs that depolymerized (15 μM Taxol) in the presence of Kar3Cik1 (Figures 2E–2G). Note that the concentration of Taxol used for Kar3Cik1-promoted MT depolymerization in the different experiments varied. The concentration required was determined in each case experimentally through cosedimentation assays and SDS-PAGE. The steady-state ATPase kinetics reveal a slightly higher kcat at conditions under which Kar3Cik1 promoted motility (0.37 s−1) in comparison to conditions for Kar3Cik1-promoted MT depolymerization (0.26 s−1). The two functional activities of Kar3Cik1 exhibited different relative affinities for MgATP: Km,ATP = 8.2 μM for stable MTs and 71 μM for depolymerizing MTs. At this time, we do not know the mechanistic significance of the differences in the steady-state parameters for the two activities—MT depolymerization versus MT gliding.
Kar3Cik1 Promotes Plus-to-Minus-End Microtubule Shortening
We visualized Kar3Cik1-promoted MT depolymerization with rhodamine-MTs immobilized by the MT binding protein GST-XCTK2-NM [36], which acted as a scaffold to hold the MTs away from the glass coverslip. The MTs were imaged, and the reaction was initiated by MgATP (Figure 4). Figures 4A–4C show times 0 and 60 min and the overlay of the two images. The red-tip MTs reflect shortening during the reaction. The MTs generally shorten from only one end (Figure 4C, table). MT shortening depended on the addition of Kar3Cik1 plus MgATP. No change in MT length was observed in the absence of motor or in the presence of Kar3Cik1 plus MgAMPPNP (Figure 4, table). These experiments were consistent with our solution studies in which Kar3Cik1-promoted MT depolymerization required ATP turnover (Figure 3).
The use of polarity-marked MTs showed Kar3Cik1-promoted shortening from the MT plus end (Figures 4D–4F). We also evaluated dimeric Ncd to determine whether MT depolymerization was characteristic of other C-terminal kinesins. Dimeric Ncd also promoted MT shortening from the plus- to the minus end (Figures 4G–4I). The rates of motor-promoted depolymerization were similar for Kar3Cik1 (45 nm/min) and Ncd (53 nm/ min). In contrast, MCAK promoted MT shortening from both ends, and at rates significantly faster (>800 nm/ min) [15].
For the polarity-marked MTs, we saw no examples in which Kar3Cik1 promoted MT shortening specifically from the minus end, although some MTs showed shortening from both ends for Kar3Cik1 and Ncd (Figure 4, table). In contrast, conventional kinesin K401 was not observed to shorten MTs (Figures 4P–4R). We do not know whether the depolymerase activity of Ncd is biologically relevant because there are no genetic or cellular studies reported to date that implicate Ncd in regulating MT dynamics. However, our results document the ability of Ncd to shorten MTs with characteristics similar to Kar3Cik1 in vitro. These observations suggest that this depolymerase activity may be common to the Kinesin-14 C-terminal kinesins.
Kar3Cik1 Has a Higher Affinity for the Microtubule End than Kar3MD
To determine whether Cik1 modulates the binding of Kar3 to the MT, we localized Kar3 on rhodamine-MTs with polyclonal antibodies to the Kar3MD. The affinity-purified Kar3 antibodies recognize both the Kar3MD and the Kar3 subunit of Kar3Cik1 but not tubulin or Cik1, as shown by Western blot (Figure 2B). We localized both Ncd and K401 with an antibody to a conserved sequence in the kinesin catalytic core. In the absence of motor, there was very little nonspecific binding, resulting in low background fluorescence (Figures 5A–5C). Figures 5D–5F illustrate an example of a polarity-marked MT with Kar3Cik1 localized to the plus end, Figures 5G–5I show Kar3MD bound to the MT lattice, Figures 5J–5L demonstrate Ncd binding to the lattice, and Figures 5M–5O show conventional kinesin coating the MT. The Figure 5 table summarizes the localization results for the Kar3MD, Kar3Cik1, dimeric Ncd, and dimeric K401. Very few localization events were scored in the absence of motor, and very few binding events were evident at 50 nM Kar3MD in comparison to 50 nM Kar3Cik1, 50 nM Ncd, or 50 nM K401. These results indicate that the Kar3Cik1 heterodimer shows a higher affinity for MTs than the monomeric Kar3MD. For the Kar3MD at 100 nM, 21.7% of scored MTs showed end binding, 72.9% with lattice binding, and 5.3% with lattice and end binding. In contrast, at both 50 nM and 100 nM Kar3Cik1, 48% of the MTs exhibited end binding, 27% lattice binding, and 25% lattice and end binding. These data indicate that there is a high-affinity site at the MT end for Kar3Cik1, resulting in MT end binding in preference to MT lattice binding. In contrast, the Kar3MD binds more often to the MT lattice. The results for Ncd are more similar to the Kar3MD results, and conventional kinesin K401 shows no preference for the MT ends.
Figure 5. Immunolocalization of Kar3MD, Kar3Cik1, and Ncd.

Mt•motor complexes were preformed in solution in the presence of MgAMPPNP, and glutaraldehyde fixation and centrifugation through a glycerol cushion onto coverslips followed. The coverslips were processed for immunofluorescence. Final concentrations: 50 nM or 100 nM motor, 500 nM tubulin, 3 μM Taxol, and 1 mM MgAMPPNP. All panels represent the same magnification. The scale bar represents 5 μm. The MT seed (arrowhead) marks the MT minus end, and (*) denotes the MT plus end extended from the seed.
(A, D, G, J, and M) Rhodamine-labeled tubulin.
(B, E, H, K, and N) Anti-Kar3/Kinesin fluorescence.
(C, F, I, L, and O) Merge of the two channels to show MT-motor colocalization. The table shows the summary of MT localization events scored for the four motors and the no-motor control.
Discussion
We provide the first in vitro characterization of Kar3Cik1 mechanochemistry. Our results reveal that C-terminal kinesins exhibit a depolymerase activity quite distinct from the MCAK depolymerases. In vitro, Kar3Cik1 binds preferentially to one end of the MT and promotes slow, directed MT shortening that occurs from only the plus end of MTs. This mechanism is novel and critical for the hypothesized roles of Kar3Cik1 during karyogamy, meiosis, and mitosis [1–11, 21–23, 26, 37, 38]. In vivo, most kinesin depolymerases appear to facilitate dynamic instability, catalyzing the rapid MT fluctuations associated with chromosome “search and capture” during mitosis [39–41]. To date, all of these kinesins have been MCAK Kinesin-13s, but other types of MT depolymerization events occur in the cell, for example the very specific unidirectional MT depolymerization that leads to karyogamy. It is plausible to argue that a C-terminal kinesin, possessing minus-end-directed force, makes an attractive candidate to ensure directed, coordinated shortening of the MT from the plus end to carry out events in which MT shortening must be temporally and spatially exact. Kar3Cik1 is the first example of a motor used in such a specialized cellular event.
We propose that Cik1 targets Kar3 to the plus end of MTs in the shmoo tip for MT shortening, and Kar3Cik1-promoted depolymerization acts to pull the nucleus into the shmoo tip (Figure 1). Once the two shmoo cytoplasms become confluent, our results would suggest that Kar3Cik1 uses both its MT gliding activity to slide antiparallel MTs and its depolymerase activity to shorten the astral MTs for nuclear fusion. Our in vitro data provide a mechanism to explain the role of Cik1 for Kar3Cik1 function in karyogamy, but unresolved questions remain. First, Kar3Cik1 appears to be tethered to the cortex much like Bim1-Kar9 for MT polymerization [26], and our in vitro analysis shows that it is the C-terminal domain of Kar3Cik1 that tracks with the shortening MTs. Therefore, our results suggest that it is the N-terminal domain of Kar3Cik1 that binds to a cortex binding partner yet to be identified. Because Kar3-Cik1 can promote both MT shortening and MT gliding in vitro, we cannot rule out a requirement for Kar3Cik1 MT sliding activity for nuclear fusion. Lastly, we do not yet know if other MT- and/or Kar3Cik1-associated proteins participate to regulate Kar3Cik1-promoted MT depolymerization during karyogamy.
Supplementary Material
Detailed Experimental Procedures and a supplemental movie are available at http://www.current-biology.com/cgi/content/full/15/15/1420/DC1/.
Acknowledgments
We thank Sharyn Endow for the Kar3MD and Ncd MC1 clones, Michael Snyder for the yeast genomic DNA, and Claire Walczak for MCAK and GST-XCTK2-NM proteins. We also thank the Walczak group for assisting us in establishing the microscopy assays. This work was supported by National Institute of General Medical Sciences Grant GM54141 and National Institute of Arthritis and Musculoskeletal and Skin Diseases Career Development Award K02-AR47841 to S.P.G.
References
- 1.Meluh PB, Rose MD. KAR3, a kinesin-related gene required for yeast nuclear fusion. Cell. 1990;60:1029–1041. doi: 10.1016/0092-8674(90)90351-e. [DOI] [PubMed] [Google Scholar]
- 2.Middleton K, Carbon J. KAR3-encoded kinesin is a minus-end-directed motor that functions with centromere binding proteins (CBF3) on an in vitro yeast kinetochore. Proc Natl Acad Sci USA. 1994;91:7212–7216. doi: 10.1073/pnas.91.15.7212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Saunders W, Hornack D, Lengyel V, Deng C. The Saccharomyces cerevisiae kinesin-related motor Kar3p acts at preanaphase spindle poles to limit the number and length of cytoplasmic microtubules. J Cell Biol. 1997;137:417–431. doi: 10.1083/jcb.137.2.417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Saunders W, Lengyel V, Hoyt MA. Mitotic spindle function in Saccharomyces cerevisiae requires a balance between different types of kinesin-related motors. Mol Biol Cell. 1997;8:1025–1033. doi: 10.1091/mbc.8.6.1025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bascom-Slack CA, Dawson DS. The yeast motor protein, Kar3p, is essential for meiosis I. J Cell Biol. 1997;139:459–467. doi: 10.1083/jcb.139.2.459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Shanks RM, Bascom-Slack C, Dawson DS. Analysis of the kar3 meiotic arrest in Saccharomyces cerevisiae. Cell Cycle. 2004;3:363–371. [PubMed] [Google Scholar]
- 7.Tanaka K, Mukae N, Dewar H, van Breugel M, James EK, Prescott AR, Antony C, Tanaka TU. Molecular mechanisms of kinetochore capture by spindle microtubules. Nature. 2005;434:987–994. doi: 10.1038/nature03483. [DOI] [PubMed] [Google Scholar]
- 8.Page BD, Snyder M. CIK1: A developmentally regulated spindle pole body-associated protein important for microtubule functions in Saccharomyces cerevisiae. Genes Dev. 1992;6:1414–1429. doi: 10.1101/gad.6.8.1414. [DOI] [PubMed] [Google Scholar]
- 9.Page BD, Satterwhite LL, Rose MD, Snyder M. Localization of the Kar3 kinesin heavy chain-related protein requires the Cik1 interacting protein. J Cell Biol. 1994;124:507–519. doi: 10.1083/jcb.124.4.507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Manning BD, Barrett JG, Wallace JA, Granok H, Snyder M. Differential regulation of the Kar3p kinesin-related protein by two associated proteins, Cik1p and Vik1p. J Cell Biol. 1999;144:1219–1233. doi: 10.1083/jcb.144.6.1219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Barrett JG, Manning BD, Snyder M. The Kar3p kinesin-related protein forms a novel heterodimeric structure with its associated protein Cik1p. Mol Biol Cell. 2000;11:2373–2385. doi: 10.1091/mbc.11.7.2373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Desai A, Verma S, Mitchison TJ, Walczak CE. Kin I kinesins are microtubule-destabilizing enzymes. Cell. 1999;96:69–78. doi: 10.1016/s0092-8674(00)80960-5. [DOI] [PubMed] [Google Scholar]
- 13.Niederstrasser H, Salehi-Had H, Gan EC, Walczak C, Nogales E. XKCM1 acts on a single protofilament and requires the C terminus of tubulin. J Mol Biol. 2002;316:817–828. doi: 10.1006/jmbi.2001.5360. [DOI] [PubMed] [Google Scholar]
- 14.Moores CA, Yu M, Guo J, Beraud C, Sakowicz R, Milligan RA. A mechanism for microtubule depolymerization by KinI kinesins. Mol Cell. 2002;9:903–909. doi: 10.1016/s1097-2765(02)00503-8. [DOI] [PubMed] [Google Scholar]
- 15.Hunter AW, Caplow M, Coy DL, Hancock WO, Diez S, Wordeman L, Howard J. The kinesin-related protein MCAK is a microtubule depolymerase that forms an ATP-hydrolyzing complex at microtubule ends. Mol Cell. 2003;11:445–457. doi: 10.1016/s1097-2765(03)00049-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Moores CA, Hekmat-Nejad M, Sakowicz R, Milligan RA. Regulation of KinI kinesin ATPase activity by binding to the microtubule lattice. J Cell Biol. 2003;163:963–971. doi: 10.1083/jcb.200304034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ogawa T, Nitta R, Okada Y, Hirokawa N. A common mechanism for microtubule destabilizers—M type kinesins stabilize curling of the protofilament using the class-specific neck and loops. Cell. 2004;116:591–602. doi: 10.1016/s0092-8674(04)00129-1. [DOI] [PubMed] [Google Scholar]
- 18.Shipley K, Hekmat-Nejad M, Turner J, Moores C, Anderson R, Milligan R, Sakowicz R, Fletterick R. Structure of a kinesin microtubule depolymerization machine. EMBO J. 2004;23:1422–1432. doi: 10.1038/sj.emboj.7600165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hildebrandt ER, Hoyt MA. Mitotic motors in Saccharomyces cerevisiae. Biochim Biophys Acta. 2000;1496:99–116. doi: 10.1016/s0167-4889(00)00012-4. [DOI] [PubMed] [Google Scholar]
- 20.Endow SA, Kang SJ, Satterwhite LL, Rose MD, Skeen VP, Salmon ED. Yeast Kar3 is a minus-end microtubule motor protein that destabilizes microtubules preferentially at the minus ends. EMBO J. 1994;13:2708–2713. doi: 10.1002/j.1460-2075.1994.tb06561.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hoepfner D, Schaerer F, Brachat A, Wach A, Philippsen P. Reorientation of mispositioned spindles in short astral microtubule mutant spc72Delta is dependent on spindle pole body outer plaque and Kar3 motor protein. Mol Biol Cell. 2002;13:1366–1380. doi: 10.1091/mbc.01-07-0338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Shanks RM, Kamieniecki RJ, Dawson DS. The Kar3-interacting protein Cik1p plays a critical role in passage through meiosis I in Saccharomyces cerevisiae. Genetics. 2001;159:939–951. doi: 10.1093/genetics/159.3.939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Huyett A, Kahana J, Silver P, Zeng X, Saunders WS. The Kar3p and Kip2p motors function antagonistically at the spindle poles to influence cytoplasmic microtubule numbers. J Cell Sci. 1998;111:295–301. doi: 10.1242/jcs.111.3.295. [DOI] [PubMed] [Google Scholar]
- 24.Maddox P, Chin E, Mallavarapu A, Yeh E, Salmon ED, Bloom K. Microtubule dynamics from mating through the first zygotic division in the budding yeast Saccharomyces cerevisiae. J Cell Biol. 1999;144:977–987. doi: 10.1083/jcb.144.5.977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Maddox PS, Bloom KS, Salmon ED. The polarity and dynamics of microtubule assembly in the budding yeast Saccharomyces cerevisiae. Nat Cell Biol. 2000;2:36–41. doi: 10.1038/71357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Maddox PS, Stemple JK, Satterwhite L, Salmon ED, Bloom K. The minus end-directed motor Kar3 is required for coupling dynamic microtubule plus ends to the cortical shmoo tip in budding yeast. Curr Biol. 2003;13:1423–1428. doi: 10.1016/s0960-9822(03)00547-5. [DOI] [PubMed] [Google Scholar]
- 27.Miller RK, Cheng SC, Rose MD. Bim1p/Yeb1p mediates the Kar9p-dependent cortical attachment of cytoplasmic microtubules. Mol Biol Cell. 2000;11:2949–2959. doi: 10.1091/mbc.11.9.2949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Hwang E, Kusch J, Barral Y, Huffaker TC. Spindle orientation in Saccharomyces cerevisiae depends on the transport of microtubule ends along polarized actin cables. J Cell Biol. 2003;161:483–488. doi: 10.1083/jcb.200302030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Mackey AT, Gilbert SP. The ATPase cross-bridge cycle of the Kar3 motor domain. Implications for single head motility. J Biol Chem. 2003;278:3527–3535. doi: 10.1074/jbc.M206219200. [DOI] [PubMed] [Google Scholar]
- 30.Foster KA, Correia JJ, Gilbert SP. Equilibrium binding studies of non-claret disjunctional protein (Ncd) reveal cooperative interactions between the motor domains. J Biol Chem. 1998;273:35307–35318. doi: 10.1074/jbc.273.52.35307. [DOI] [PubMed] [Google Scholar]
- 31.Foster KA, Mackey AT, Gilbert SP. A mechanistic model for Ncd directionality. J Biol Chem. 2001;276:19259–19266. doi: 10.1074/jbc.M008347200. [DOI] [PubMed] [Google Scholar]
- 32.Gilbert SP, Johnson KA. Expression, purification, and characterization of the Drosophila kinesin motor domain produced in Escherichia coli. Biochemistry. 1993;32:4677–4684. doi: 10.1021/bi00068a028. [DOI] [PubMed] [Google Scholar]
- 33.Correia JJ, Gilbert SP, Moyer ML, Johnson KA. Sedimentation studies on the kinesin motor domain constructs K401, K366, and K341. Biochemistry. 1995;34:4898–4907. doi: 10.1021/bi00014a047. [DOI] [PubMed] [Google Scholar]
- 34.Ovechkina Y, Wagenbach M, Wordeman L. K-loop insertion restores microtubule depolymerizing activity of a “neckless” MCAK mutant. J Cell Biol. 2002;159:557–562. doi: 10.1083/jcb.200205089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Walker RA, Pryer NK, Salmon ED. Dilution of individual microtubules observed in real time in vitro: Evidence that cap size is small and independent of elongation rate. J Cell Biol. 1991;114:73–81. doi: 10.1083/jcb.114.1.73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ems-McClung SC, Zheng Y, Walczak CE. Importin alpha/beta and Ran-GTP regulate XCTK2 microtubule binding through a bipartite nuclear localization signal. Mol Biol Cell. 2004;15:46–57. doi: 10.1091/mbc.E03-07-0454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Manning BD, Snyder M. Drivers and passengers wanted! The role of kinesin-associated proteins. Trends Cell Biol. 2000;10:281–289. doi: 10.1016/s0962-8924(00)01774-8. [DOI] [PubMed] [Google Scholar]
- 38.Trelles-Sticken E, Loidl J, Scherthan H. Increased ploidy and KAR3 and SIR3 disruption alter the dynamics of meiotic chromosomes and telomeres. J Cell Sci. 2003;116:2431–2442. doi: 10.1242/jcs.00453. [DOI] [PubMed] [Google Scholar]
- 39.Wordeman L, Mitchison TJ. Identification and partial characterization of mitotic centromere-associated kinesin, a kinesin-related protein that associates with centromeres during mitosis. J Cell Biol. 1995;128:95–104. doi: 10.1083/jcb.128.1.95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Walczak CE, Mitchison TJ, Desai A. XKCM1: A Xenopus kinesin-related protein that regulates microtubule dynamics during mitotic spindle assembly. Cell. 1996;84:37–47. doi: 10.1016/s0092-8674(00)80991-5. [DOI] [PubMed] [Google Scholar]
- 41.Maney T, Hunter AW, Wagenbach M, Wordeman L. Mitotic centromere-associated kinesin is important for anaphase chromosome segregation. J Cell Biol. 1998;142:787–801. doi: 10.1083/jcb.142.3.787. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Detailed Experimental Procedures and a supplemental movie are available at http://www.current-biology.com/cgi/content/full/15/15/1420/DC1/.