

Abstract
Animal models contribute to the understanding of molecular mechanism of cancer, revealing complex roles of altered cellular-signaling networks and deficient surveillance systems. Analogous pathologies are documented in an unconventional model organism that receives attention in research on systems theory, evolution, and aging. The honeybee (Apis mellifera) colony is an advanced integrative unit, a “superorganism” in which order is controlled via complex signaling cascades and surveillance schemes. A facultatively sterile caste, the workers, regulates patterns of growth, differentiation, homeostasis, and death. Workers differentiate into temporal phenotypes in response to dynamic social cues; chemosensory signals that can translate into dramatic physiological responses, including programmed cell death. Temporal worker forms function together, and effectively identify and terminate abnormal colony members ranging from embryos to adults. As long as this regulatory system is operational at a colony level, the unit survives and propagates. However, if the worker phenotypes that collectively govern order become too few or change into malignant forms that bypass control mechanisms to replicate aberrantly; order is replaced by disorder that ultimately leads to the destruction of the society. In this chapter we describe fundamental properties of honeybee social organization, and explore conditions that lead to states of disorder. Our hope is that this chapter will be an inspirational source for ongoing and future work in the field of cancer research.
I. ORDER IN LIVING ORGANISMS
Cancer is a disease of genes (Bishop, 1991) that is characterized by loss of intercellular order (Schafer, 1969). Tumor cells emerge through carcinogenic mutations at critical loci that underlie regulation of cell growth and cell death (Ryan et al., 2000; Thompson, 1995). Disorder arises from the uncontrolled behavior of the mutated cell type, which invades neighboring tissues and spreads malignant cells (Karin and Greten, 2005). The tumorigenic process of initiation, promotion, and progression (Hoeijmakers, 2001; Karin and Greten, 2005), thereby, is a complex product of defective control mechanisms that collectively tear down the order of the living organism.
Animal models contribute broadly to insights into the dynamics of cancer (Van Dyke and Jacks, 2002). Drosophila researchers have developed tools for rapid characterization of tumor genes (Potter et al., 2000), and forward genetic approaches in the zebra fish have uncovered cancer pathways (Shepard et al., 2005). Studies of the mouse, in particular, have highlighted the intricacy of cancer: unraveling the faceted interface between the tumor cells and their “host” organism, and the subtle genetic interactions that influence individual susceptibility to disease (Balmain, 2002; Van Dyke and Jacks, 2002). Our understanding of the molecular systems that drive the onset and progression of carcinogenesis are thus developing rapidly, while research is uncovering mechanisms that are expected to lead to new paradigms for prevention, diagnostics, and therapy (Hoeijmakers, 2001).
Yet, novel insights into complex phenomena can be constrained by the models that are used also when the accumulation of knowledge in a field is remarkable (Golstein et al., 2003). Work on pending problems can be inspired by findings in distant research areas and the models that turn out to be the most valuable in this context are often the least predictable. Here, we argue that lessons at the intersection between order, disorder, and death can arise from biological systems other than the molecular networks that govern organismal order. Our outline focuses on an unconventional model in medical research—a superorganism.
II. SUPERORGANISMS
A superorganism is defined as “a collection of single creatures that together possess the functional organization implicit in the formal definition of organism” (Wilson and Sober, 1989). The concept emerged from studies of the tightly interlinked colonies of eusocial insects: the ants, wasps, bees, and termites (Wheeler, 1911, 1928). Societies of eusocial species are characterized by differentiation of individuals into reproductive and facultative or functionally sterile forms called castes. Colonies consist of numerous overlapping generations of such individual forms that function as an integrative whole; a unique biology that in many aspects mimics the basal anatomy of multicellular organisms (Wilson, 1985). By adopting a definition of Metazoan animals as “composed of cells arranged in at least two nonuniform layers and differentiated into reproductive and somatic cells with different functions” (Kaestner, 1969), the reproductive castes become analogous to the germ line, and the sterile forms become analogous to the soma (Wilson, 1971, 1975). Like somatic cells, the sterile individuals further specialize through processes of differentiation, and communicate to produce coordinated patterns of growth, homeostasis, provisioning, and defense before death.
The usefulness of the superorganism analogy has been debated, in particular, with respect to natural selection on the level of social groups (Mitchell, 2003). However, studies of the organizational principles of social insect societies have led to deep insights into the emergent properties of biological systems (Camazine et al., 2001; Fewell, 2003; Hölldobler, 1995; Theraulaz et al., 2002), and research on the underlying modes of interindividual communication has inspired the design of control algorithms that exhibit high degrees of flexibility and robustness (Bonabeau et al., 2000). Thus, although comparisons between the evolution and development of multicellular creatures and superorganisms are controversial (Mitchell, 2003; Wilson, 1975; Wilson and Sober, 1989), the highly integrative societies of insects are recognized systems for understanding how complex order arises from the actions of individual parts (Fewell, 2003).
The remarkably coordinated and manifold activities of ant, wasp, bee, and termite societies have long been subject to curiosity and awe (Mitchell, 2003). This focus may explain why the dynamics of disorder have received less general attention than the principles that govern the exceptional order of these systems. Initiation, promotion, and progression of states that tear down the well-organized structure of superorganisms nevertheless have been examined, and it is possible that deeper knowledge of such states can lead to conceptual insights of heuristic value. In the following, we take a first step toward such insights by reviewing features of the biology and pathology of the honeybee (Apis mellifera), the best studied eusocial invertebrate to date.
III. HONEYBEE SOCIETY
The honeybee is one of several social bees of the genus Apis. It is widely domesticated as a source of honey and beeswax and a well-known research system for understanding the characteristic differentiation of eusocial individuals into reproductive and sterile forms (Amdam et al., 2004a; Bloch et al., 2002; Robinson et al., 2005). The bee is an established model in systems theory (Mitchell, 2003; Page and Erber, 2002), behavioral ecology (Seeley, 1995), neurobiology (Farooqui et al., 2004; Humphries et al., 2003; Menzel, 1979; Scheiner et al., 2001), and aging (Amdam and Page, 2005; Amdam et al., 2005; Omholt and Amdam, 2004; Seehuus et al., 2006). It has a rising position in molecular research that is fueled by an emerging availability of functional genomic tools (Aase et al., 2005; Amdam et al., 2003; Beye et al., 2002; Omholt et al., 1995; Yu et al., 1997) and a newly annotated genome sequence (www.hgsc.bcm.tmc.edu).
A. Anatomy of Individuals
Honeybees are haplo-diploid (males are haploid). Their sex is determined by a single gene, csd, which has structural similarity to the tra genes of dipteran insects (Beye et al., 2003). Females are derived from fertilized eggs and are heterozygous at the csd locus, whereas males typically develop from unfertilized eggs and are hemizygous at the csd locus. Diploid males occur when the locus is homozygous (Santomauro et al., 2004).
Honeybee females differentiate into two distinct forms: the queen (Fig. 1A) and the worker (Fig. 1B). The queen's anatomy is characterized by an enlarged ovary (180−200 ovarioles per ovary) and a sperm-storage organ, the spermatheca, that can store over 5 million spermatozoa (Kraus et al., 2004; Snodgrass, 1956). The end stylet of the queen's stinging apparatus is smooth so it can be retracted after stinging, and the queen produces a specific signature of C21–C33 cuticular hydrocarbons in addition to a pheromone blend from enlarged glands in the head (mandibular gland), abdomen (Dufour and tergite gland), and tarsus (tarsal gland); major components being (E)-9-oxodec-2-enoic acid, (E)-9-hydroxydec-2- enoic acid, methyl p-hydroxybenzoate, and 4-hydroxy-3-methoxyphenylethanol (Free, 1987; Keeling et al., 2003).
Fig. 1.

The castes of the honeybee. There is only one queen (A) in a honeybee colony. She is the dominant reproductive female and can live up to 3−5 years. The worker (B) is a facultatively sterile female with a typical lifespan of 4−6 weeks. A colony normally consists of 10,000−30,000 worker bees. The male caste, or drone (C), is only raised under favorable conditions when there may be 300−3000 drones present in the society. Few drones survive more than 4−5 weeks.
The worker (Fig. 1B) has 2−16 ovarioles per ovary and no spermatheca. The end stylet of her stinger has series of recurved barbs that ensures that the stinger and venom sac is anchored in the enemy after stinging. The worker has a specialized third hind-leg segment, called the corbicula or “pollen basket,” in which pollen and propolis is carried to the hive. She also has a set of paired head glands, the hypopharyngeal glands, that during the worker's temporal nurse bee stage (Section IV) produces a proteinaceous jelly used for brood rearing (royal jelly). The jelly contains proteins of the MRJP/yellow family, sugars, lipids, vitamins, and free amino acids; and has fractions with disinfectant, antitumor and anti-inflammatory activities (Kamakura and Sakaki, 2005). The worker bee has two specialized abdominal glands: the wax gland and the Nasanov gland. The latter gland releases an attractant aggregation pheromone in response to disturbances; major components being geraniol, nerolic acid, and geranic acid (Winston, 1987). Furthermore, the stinger discharges alarm pheromones (primarily isoamyl acetate) after it has been embedded in the opponent. The worker bee is characterized by a stage-specific profile of C21–C33 cuticular hydrocarbons and pheromones, including eicosenol, ethyl oleate, and 10-hydroxy-decenoic acid that changes as a function of the social role of the animal (Chaline et al., 2005; Dor et al., 2005; Leoncini et al., 2004a,b).
The males, or “drones” (Fig. 1C), do not have a stinger because this character evolved from the female ovipositor. Drones also lack specialized body parts for food collection (Snodgrass, 1956) and are easily recognizable with broad thoraxes, rectangular abdomens, and enlarge compound eyes. They are designed to perform one successful mating flight, and the act of mating is fatal to the drone (Winston, 1987). Diploid drones are normally recognized and selectively killed by workers shortly after hatching from the egg (Section V). In vitro rearing studies indicate that these males are characterized by small testes and diploid spermatozoa (Herrmann et al., 2005). For further details on honeybee anatomy, see work by Snodgrass (1956).
B. Anatomy of the Colony Unit
The honeybee colony dwells in a nest cavity reinforced by the resinous substance propolis (Fig. 2). Brood (eggs, larvae, and pupae) and hoarded foods (honey and pollen) are located in the main nest structure, the wax combs. The combs are arranged in sheets and the nest is organized spatially with the brood located centrally (Winston, 1987). A controlled amount of pollen (Section IV) is stored close to the brood. Pollen is the main amino acid source for the honeybees. Honey is stored in surplus at the periphery of the wax combs and is the society's source of carbohydrates. The pollen, honey, and propolis resin of the nest may, in addition to the jelly, be part of a collective antibacterial defense system. Active compounds are, for example, chrysin and cinnamic acid derivatives, such as baccharin and drupanin, that have growth-inhibitor and also antitumor activities (Mishima et al., 2005). The central nest is thermoregulated at 33°C and 80% relative humidity (Section IV), which is optimal for the growth and development of the brood.
Fig. 2.
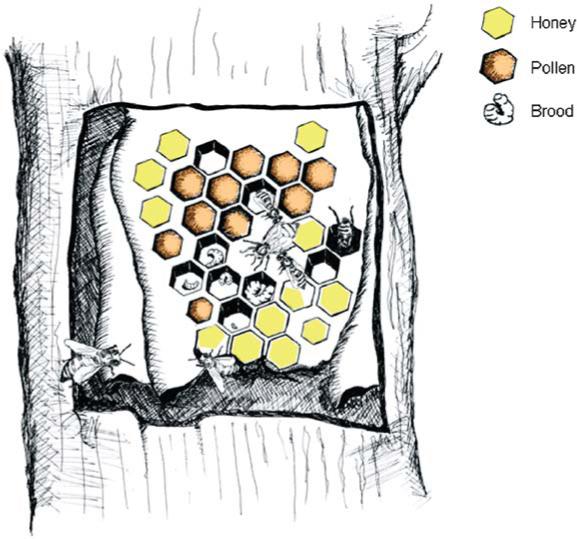
The honeybee colony—schematic illustration of a feral nest inside a trunk cavity. The cross-section shows the spatial arrangement of wax-cells of honey at the periphery and a pollen storage close to the central brood nest.
The queen is most frequently found in the central nest along with the drones and young worker bees. The workers feed developing larvae, and they are responsible for colony hygiene, comb construction, and food processing (Winston, 1987). Later in life, worker bees make a transition from nest tasks to guarding and foraging outside the colony (Seeley, 1982). After this shift, they are seldom observed performing tasks within the nest other than those directly related to foraging such as unloading pollen and nectar and performing recruitment dances. When agitated by alarm pheromones, though, workers of diverse ages and functions can be recruited to the colony's strong defensive response (aggressive stinging). The overall anatomy of the colony is maintained through the temporal and spatial division of labor between the workers; a pattern of organization that, in principle, arises through the emergence of new adults in the central brood nest (Fig. 2) that replaces the bees that shift to tasks in the periphery and eventually die.
IV. REGULATION OF DIFFERENTIATION, GROWTH, AND HOMEOSTASIS
The female forms (Fig. 1A and B) are the fundamental components that underlie superorganismal order in the bee because the drones do not have colony-level functions other than the act of mating. Note, however, that this phenomenon is specific to the Hymenoptera lineage of social insects. In the termites, Isoptera, males and females are diploid, and both sexes have worker and reproductive castes.
A. Differentiation of Female Forms
Segregation of female phenotypes occurs at two levels of biological organization in the honeybee superorganism. A developmental bifurcation separates the queen caste from the worker caste during the larval stage, and adult regulatory pathways subsequently drive the temporal specialization of the mature worker bees.
The caste trajectories of the queens and workers are decoupled during the third larval instar via a feeding-sensitive endocrine switch (Fig. 3). Differentiation, thereby, is ultimately determined by social nursing regimens that are enforced by the adult worker population (Winston, 1987). Ample feeding of proteinaceous jelly to a larva triggers an increase in the circulating titer of the systemic hormone “juvenile hormone” that is secreted by the corpus allatum complex (Hartfelder and Engels, 1998). The resulting signaling cascade gives rise to a queen phenotype. Poorer dietary conditions that result from reduced feeding suppress juvenile hormone signaling and trigger programmed cell death in the developing ovary (Capella and Hartfelder, 1998). The endocrine shift changes global gene expression patterns (Evans and Wheeler, 1999) and yields a worker phenotype characterized by half the body weight of a queen (a queen is ∼200 mg). The role of feeding in caste differentiation has been linked to a proposed growth factor- or hormone-like activity of the honeybee jelly protein p56kP-4, which has close homology to Drosophila Yellow protein (Kamakura and Sakaki, 2005).
Fig. 3.
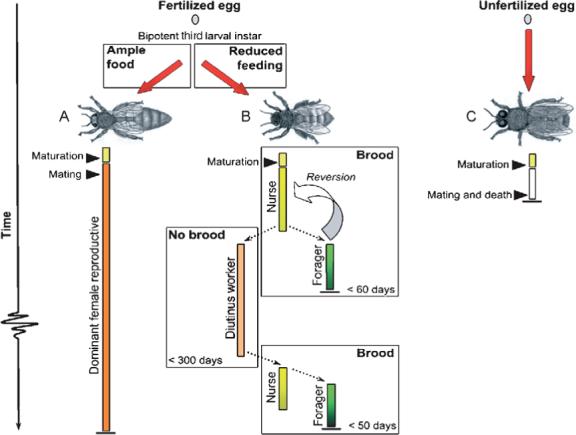
Differentiation of honeybees castes and further specialization of workers. After hatching, a diploid honeybee larva can either become a worker or a queen. The differentiation depends solely on nourishment provided by adult workers during the first days of larval life. Ample feeding leads to a queen phenotype (A). Newly emerged virgin queens go through a maturation period before they mate and become the dominant reproductive female of a nest. Reduced feeding of larvae leads to a worker phenotype (B). After maturation, the worker enters the temporal nurse bee stage (duration of this stage is typically in the range of 7−40 days). Nurse bees can differentiate into foragers or diutinus workers depending on whether brood is present in the colony. As a forager, the bee collects resources in the field (duration of this stage is about 7−21 days), but may also revert to nest tasks if many nurse bees are removed from the colony. As a diutinus worker, the bee survives for several months (up to 280 days) before she differentiates into a nurse or a forager (Omholt and Amdam, 2004). The males, or drones (C), are haploid and die as soon as they have mated with a virgin queen.
The adult workers differentiate into three temporal forms referred to as the “nurse bee,” the “forager,” and the “diutinus worker” stages, respectively (Amdam and Page, 2005). The nurse bee (Fig. 3) is the phenotype that performs tasks within the nest, and the majority of workers mature into nurse bees within 3−4 days after adult emergence (Naiem et al., 1999). They become characterized by enlarged jelly-synthesizing hypopharyngeal glands, hypertrophied abdominal fat bodies (analogous to the mammalian liver and adipose tissue), and they also have a high number of circulating hemocytes (immune cells). Like T cells, insect hemocytes have several important immunological functions in nonself recognition, phagocytosis, encapsulation, nodulation, and wound repair (Millar and Ratcliffe, 1994), and the circulating number reflects the organism's capacity to cope with immunogenic challenges (Doums et al., 2002; Kraaijeveld et al., 2001). Severe infections, that is, more than 1000 microbes per microliter hemolymph (blood), thus, rarely occur in nurse bees (Amdam et al., 2005).
Foragers (Fig. 3) typically develop from nurse bees (Seeley, 1982). They work in the nest periphery and are responsible for collecting nectar, pollen, water, and propolis in the field. Workers in this temporal stage secrete low levels of digestive endopeptidases and are unable to obtain amino acids from the colony's pollen store (Crailsheim, 1990). The protein intake of a forager, therefore, is controlled by the nurse bees that feed the forager in proportion to her foraging activity level (Crailsheim et al., 1999). A forager is characterized by atrophy and apoptosis of the hypopharyngeal glands (De Moraes and Bowen, 2000) and the abdominal fat body (Seehuus and Amdam, unpublished data). She has very few circulating hemocytes (Rutz et al., 1974; Wille and Rutz, 1975) as the immune cells also apoptose during the transition from nurse tasks to foraging activities (Amdam et al., 2005), see Section V for mechanisms. The differentiation of nurse bees into foragers is driven by social signals (Section IV). At the physiological level, these signals modulate a positive regulatory feedback loop between the vitellogenin gene, which is expressed in the fat body, and juvenile hormone (Amdam and Omholt, 2003; Guidugli et al., 2005). Specifically, vitellogenin gene activity suppresses the endogenous juvenile hormone level during the nurse bee stage, whereas reduced signaling from the vitellogenin gene releases juvenile hormone activity during the forager differentiation (Guidugli et al., 2005). This endocrine signal feeds back to the regulatory system to further inhibit the synthesis of vitellogenin protein (Pinto et al., 2000), shift the gene expression pattern of the hypopharyngeal glands (Ohashi et al., 1997), and suppress immunity (Amdam et al., 2005; Rutz et al., 1974; Wille and Rutz, 1975).
Diutinus workers (Fig. 3) develop during unfavorable periods without opportunities for brood rearing (Amdam and Page, 2005; Maurizio, 1950; Omholt and Amdam, 2004). They differentiate from nurse bees that no longer take care of brood (Amdam and Omholt, 2002; Huang and Robinson, 1995; Omholt and Amdam, 2004). A diutinus worker is characterized by hypertrophy of the hypopharyngeal glands and the abdominal fat body (Deseyn and Billen, 2005; Koehler, 1921; Maurizio, 1954). She has a high number of circulating hemocytes, an elevated vitellogenin protein level, and a low juvenile hormone titer (Amdam et al., 2004b; Fluri et al., 1977, 1982). The diutinus stage is described as an oxidative stress-resistant survival form similar to the dauer larva of Caenorhabditis elegans (Seehuus et al., 2006). Diutinus workers develop the physiological and behavioral attributes of nurse bees and foragers when the queen starts to lay eggs at the onset of the next favorable season (Maurizio, 1954; Sekiguchi and Sakagami, 1966).
The temporal differentiation of honeybee workers typically follows the trajectory in which the forager stage follows the nurse stage, as described earlier. Yet, worker bees also have the unique plasticity to revert (Huang and Robinson, 1996; Robinson et al., 1992). This implies that differentiated foragers can go through a physiological reversion (Section IV)—an ontogeny (Fig. 3) that is characterized by reduced juvenile hormone signaling, elevated vitellogenin protein activity (Amdam et al., 2005), reactivation of the hypopharyngeal glands (Amdam et al., 2005; Huang and Robinson, 1996), and reversal of immunosenescence (Amdam et al., 2005).
B. Colony Growth
The honeybee superorganism reproduces as a unit through fission (swarming) and the production of drones. The process of swarming is initiated by workers that appear to respond to a queen-specific pheromone signal that diminishes as a function of colony size. Explicitly, circulation of queen pheromones is a continuous process mediated by food exchange (trophallaxis) and physical contact between workers as well as volatile transmission (Winston, 1987). This distribution process becomes less efficient as the colony grows in size. As the signal from the queen weakens, workers start to rear new queen larvae (Baird and Seeley, 1983). One of the resulting virgin queens takes over the maternal nest while the old queen leaves in a swarm with the majority (50−90%) of the workers to establish a colony at a new nest site (Severson, 1984). The virgin queen uses her retractable stinger to kill all other developing queens (Fig. 4A). Subsequently, she engages in one or several mating flights before she takes over the role as the dominant reproductive female in the old nest.
Fig. 4.
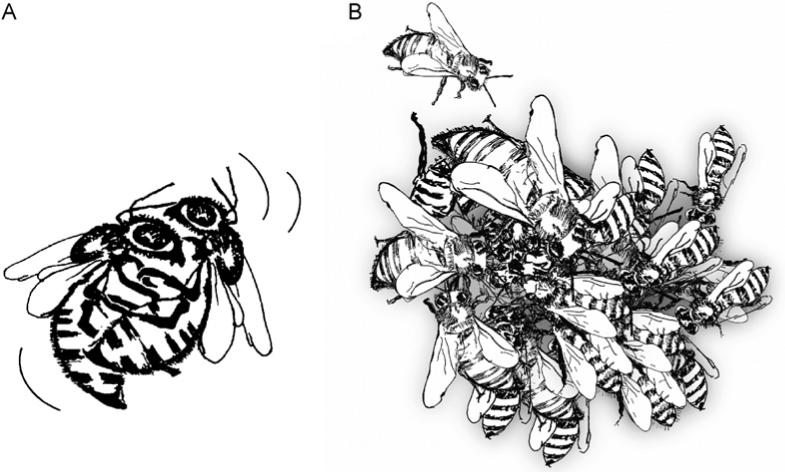
Regulation of death. The illustrations show the lethal fight between two virgin queens (A), and the excessive coating response of honeybee workers that have targeted an adult individual for termination (B). The undesired individual in the center dies of thermal stress.
After swarming, the colony grows through the reproductive effort of the queen and the alloparental investment of the workers. The queen may produce up to 1500 eggs daily, and thus if resources for brood rearing are abundant the colony can grow rapidly (Winston, 1987). Growth continues until the society once more is split by swarming or until the onset of the unfavorable season when the rearing of brood ceases and the stress-resistant diutinus workers emerge. The next favorable period triggers a phase characterized by exclusive production of worker bees that strengthen the colony's work force before drones are reared close to the peak of the colony cycle. Through these simple mechanisms, the development of the super-organism and its offspring societies is regulated.
C. Colony Homeostasis
Over the course of the favorable period, the allocation of workers to the nurse bee stage and the forager stage is not under centralized control. Rather, a dynamic differentiation emerges through social signals produced by the workers themselves. The foragers synthesize a blend of cuticular hydrocarbons and pheromones that are specific to their stage (Leoncini et al., 2004b; Pankiw, 2004a). This signature is transferred by physical contact with the nurse bees (Huang and Robinson, 1992). Nurse bees and foragers antennate, groom, and feed each other (Winston, 1987), and though these social transfers the nurse bees can identify the relative size of the forager population (Huang and Robinson, 1996). As foragers are lost, consequently, nurse bees perceive the decline and some differentiate into foragers to take the place of those that have vanished. Likewise, foragers recognize the presence of the nurse bees, and an experimental removal of the nurse bee population triggers a fraction of the forager population to revert to nurse tasks (Amdam et al., 2005; Huang and Robinson, 1996; Robinson et al., 1992; Toth and Robinson, 2005). Individual response thresholds to social signals probably constitute one factor that determines if a particular bee differentiates in response to demographic changes (Amdam and Omholt, 2003).
The amount of stored pollen in the colony (Fig. 2) is also under dynamic decentralized control. Returning pollen foragers deposit their loads directly into empty cells or cells containing pollen close to the area of the nest where the young larvae are raised. Stored pollen is consumed by nurse bees that convert it into the proteinaceous secretions of the hypopharyngeal glands (Crailsheim et al., 1992). Stored pollen inhibits pollen foraging, while pheromones produced by larvae (methyl palmitate, methyl stearate, methyl linoleate, methyl linolenate, methyl oleate, ethyl palmitate, ethyl stearate, ethyl linoleate, ethyl linolenate, and ethyl oleate) (Slessor et al., 2005) stimulate pollen foraging (Fewell and Winston, 1992; Pankiw et al., 1998). Colonies, therefore, regulate the amount of stored pollen probably through a combination of the inhibiting effects of pollen and stimulating effects of brood pheromone. This regulatory scheme, per se, is put into operation by the workers that specialize on pollen collection. In response to accumulation of pollen in the nest and reduced pheromone signaling from brood, these foragers collect smaller pollen loads or stop foraging altogether (Fewell and Winston, 1992). Equilibrium pollen intake thus equals the pollen consumption that meets the protein demands of the developing larvae. Remarkably, the workers' sensory perception of this equilibrium is determined genetically, and colonies can be selected for high and low levels of pollen hoarding (Page and Fondrk, 1995).
The temperature and humidity of the central brood nest is similarly regulated through the behavior of individual worker bees (Jones et al., 2004). In response to environmental variability (Schmickl and Crailsheim, 2004), workers in the nest fan their wings (cooling), vibrate their thorax muscles (warming), or collect water that is deposited on the combs (cooling and humidifying). Individual response thresholds to environmental fluctuations also have a strong genetic component (Jones et al., 2004). In this connection it has been proposed that the polyandrous mating biology of the honeybee queen promotes environmental homeostasis because it generates a diversity of worker genotypes that produces a fine-tuned rather than an excessive colony-level response to variations. As a whole, the social unit is a homeothermic superorganism (Grodzicki and Caputa, 2005).
V. REGULATION OF DEATH
As in multicellular organisms, order emerges in the honeybee society at the intersection between growth, differentiation, and death. The colony can regulate the lifespan of its members in two different ways. One mechanism is the programmed life termination that is built into the physiology of the temporal forager stage. The other approach is the targeted killing of abnormal and disposable colony members, which is a mechanism that is under social decentralized control.
A. Programmed Disposal of Soma
Reduced rates of tactile interactions between nurse bees and foragers cause the hemocytes, or immune cells, of the nurse bees to terminate through programmed cell death (Amdam et al., 2005). This response is one component of the workers' transition from nest tasks to foraging activities (Section IV). The loss of hemocytes is associated with a complete deterioration of the nodulation immune response (Bedick et al., 2001), a predominant defense reaction to infection in insects (Franssens et al., 2005). Severe infections are more frequently observed in foragers than in nurse bees (Amdam et al., 2005), and in general terms increased susceptibility to pathogens and toxins is a characteristic of the forager stage (Meled et al., 1998). This frailty is nonintuitive from the perspective that foragers, to a much larger extent than other workers, encounter dangerous substances as they engage in numerous foraging trips outside the protected nest: flowers have rich faunas of bacteria and fungi, and some pollens and nectars are poisonous to bees (Maurizio, 1950, 1954).
Having a robust forager caste may not be advantageous for the society, however. If resistant foragers repeatedly brought virulent strains of bacteria and fungi as well as loads of poisonous nectars to the nest, the situation could soon become disastrous for the superorganism. The increased frailty of honeybee foragers, therefore, can be interpreted to reflect a colony-level selection pressure for disease control. Foragers typically return to the same source of pollen or nectar over the course of days (Seeley, 1995) and in being strongly susceptible to substances that potentially could harm the colony unit, an individual forager dies before she can bring substantial amounts of contaminants to the hive. A wide range of physiological stressors, such as cooling, oxygen deprivation, mechanical wounding, and parasite exposure, cause nurse bees to differentiate into foragers (Bühler et al., 1983; Ebadi et al., 1980; Kovac and Crailsheim, 1988; Tustain and Faulke, 1979). Thereby, individuals in poor condition can also be driven into a physiological state in which they rapidly perish (Amdam et al., 2004b).
From a regulatory perspective, the programmed termination of the cellular immune system of the bee is astonishing because it in essence implies that an apoptosis pathway can be conditionally activated upon loss of a social signal, that is, the forager signature of hydrocarbons and pheromones. The response can be provoked in the lab by placing a nurse bee in an isolated cage, and although the bee is provided with ample pollen and nectar she will not survive for many days (Maurizio, 1950). Unfortunately, knowledge on the underlying molecular cascade is limited. What has been determined, however, is that the immune cells of the honeybee are sensitive to the level of zinc in the hemolymph. Programmed cell death is triggered in vitro if hemocytes are cultured in a zinc-deprived medium (Amdam et al., 2004b). Remarkably, the zinc concentration of worker bee hemolymph is a function of the bee's social role, so that foragers have much lower levels (0−4 ppm) than nurse bees and diutinus workers (up to 28 ppm). The vitellogenin molecule appears to be the major circulating zinc ligand in the bee (Amdam et al., 2004b), and thus the dramatic decline in the hemolymph zinc concentration of new foragers can be linked to the feedback suppression of vitellogenin synthesis that occurs as part of the workers' transition from nest tasks to foraging activities (Section IV). This scenario suggests that hemocyte apoptosis, and possibly also patterns of programmed cell death in the hypopharyngeal glands (De Moraes and Bowen, 2000) and abdominal fat body (Seehuus and Amdam, unpublished data), are downstream of a causal chain in which social deprivation triggers a decline in vitellogenin gene activity. At a mechanistic level, the inhibitory effect of vitellogenin-derived zinc availability on hemocyte apoptosis may relate to the finding that zinc also inhibits activation of apoptotic caspases 9 and 3 in cultured cancer cells (Zhao et al., 2004).
B. Collective Termination of Colony Members
Adult worker honeybees have an astonishing ability to detect abnormal character states in the developing brood and their sister workers. Under certain conditions the worker bees also determine that the drones and the queen are no longer productive members of the colony unit. These collective insights unfold as swift responses in which one or several workers target an individual for immediate termination.
1. KILLING OF DEVELOPING BROOD
The ovary of the honeybee worker is typically inactive due to suppression of oogenesis by pheromones from the brood (Winston and Slessor, 1998) and the queen's mandibular gland (Hoover et al., 2003). Yet, a worker bee will occasionally produce an egg and deposit it in the nest. This egg is unfertilized and can develop into a normal haploid drone. Few if any such drones reach adulthood, however, because the worker population has the collective ability to identify and destroy the worker-laid eggs (Ratnieks, 1993). Numerous nurse bees inspect the nest daily by inserting their head and thorax into brood cells, and if they determine that an egg is of worker origin, they eat it. Workers probably establish the source of an egg on the basis of substances that are deposited on the egg's surface when it is laid (Katzav-Gozansky et al., 2001). The queen's eggs have a more complex hydrocarbon coating than worker-laid eggs, and acetates of fatty alcohols, alkenes, and monomethylalkanes are characteristic to eggs that are laid by a queen (Katzav-Gozansky et al., 2003).
Diploid males, which occur when the sex-determining csd locus of the bee is homozygous (Section III), constitute another group that is targeted for termination. Diploid male larvae are viable but are recognized and killed as soon as they hatch from the egg (Woyke, 1980). Adult worker bees identify these drones based on their cuticular hydrocarbon profile. The cuticular profile of newly hatched honeybee larvae is mainly composed of alkanes and squalene, but the diploid drones, the diploid females, and the haploid drones produce these compounds in different amounts (Santomauro et al., 2004). The signature of the diploid females and the haploid drones are accepted by the workers whereas the profile of the diploid males is not.
Odors are also used by the worker bees to detect brood that is infested with pathogens. Sensitivity to these olfactory cues are enhanced by the neuromodulator octopamine (Spivak et al., 2003), and amazingly workers can recognize odors specific to infested brood though the wax capping that is laid down over the comb when bee larvae pupate (Gramacho and Spivak, 2003; Spivak, 1996). When a diseased individual is detected, the workers collectively uncap the brood cell, pull the pupae out of the brood nest, and dispose it of outside the colony (Boecking and Spivak, 1999). The bee's ability to discriminate between olfactory signatures may be of relevance to research on the use of odors in tumor diagnostics (Yamazaki et al., 2002). Honeybee olfactory sensitivity is remarkable, and bees can easily be trained to discriminate between odors that are presented to them (Masterman et al., 2001). Simplified, a bee sticks out her tongue in a “proboscis extension reflex” (PER) when she perceives a specific odor that previously has been paired with a sugar reward. Most bees respond with PER to the odor alone after as little as two to five conditioning trials where the odor and the sugar reward are presented together (Gerber et al., 1998).
2. STINGING, “COATING,” AND EVICTION OF ADULTS
Worker bees are wary of other adults, and if a worker encounters an individual that is not acceptable to her, she takes action (Winston, 1987). The probability of launching such attacks is stage dependent, and nurse bees and diutinus workers are less hostile than bees that typically work in the periphery of the nest (Pearce et al., 2001). The victim is usually a worker in poor condition, one with developed ovaries (a “laying worker”) or one that has lost her way while foraging and entered a colony that is not her own (Pearce et al., 2001; Ratnieks, 1988; Winston, 1987). Such individuals are most likely singled out based on deviations in the hydrocarbon blend of their cuticle (Dani et al., 2005).
The assault unfolds as the attacker bites and tries to sting the worker that is perceived as undesired or foreign. The attacker will attempt to maneuver the bee toward the colony entrance, but the victim will try to flee from its attacker. During this initial phase, however, the target is marked by alarm pheromones that are secreted by the attacker (Pettis et al., 1998). Analogous to effects of cytokines secreted by T cells, the pheromones agitate other workers and attract them to the scene. Thereby, a “coat” forms around the victim that is stung to death or bitten until she is defenseless and can be disposed of. Sometimes the coat grows so large that it takes the shape of a ball of bees that surrounds the target (Fig. 4B). In this case, the attackers do not sting but instead collectively increase the temperature inside the coat until the undesired individual in the core dies of thermal stress (Ken et al., 2005). This defense strategy is also used when predators, such as hornets, attempt to enter the honeybee nest.
Another well-described phenomenon is the massacre of the drones (Maeterlinck, 1901). As the favorable season comes to an end and there are no more virgin queens to be mated, the males are no longer tolerated by the colony. The massacre starts suddenly, and the situation escalates rapidly over the course of a day until all the males in the colony unit are under forceful attack. Without a stinger the drones are defenseless, and they are chased around and bitten before they are dumped on the ground outside the hive. Those that are able to make it back are rejected by the worker bees that guard the entrance of the nest, and soon these last males starve or freeze to death.
Likewise, the queen is not protected from the collective surveillance schemes that are enforced by the workers. The life of the queen is inevitably over when her storage of sperm becomes depleted or when her pheromone blend no longer is perceived to be an acceptable signal of fertility and health. Under these conditions, the worker bees rear a replacement virgin without swarming. The old queen may continue to lay eggs while the new queen is developing, but she is eliminated when the virgin queen has mated and begun laying eggs (Winston, 1987).
VI. ORDER, DISORDER, AND DISEASE
Through the mechanisms that are described in the previous section, superorganismal order arises in the honeybee society. The living parts of the colony are constantly replaced through differentiation and death. Homeostasis is maintained, growth and reproduction is regulated, colony defense systems are in operation, and unproductive individuals are not permitted to become a burden on the society. As in multicellular life forms, however, these states of order can be replaced by states of disorder that ultimately lead to the destruction of the superorganism.
A. Growth Disorder
The size of the worker population is a critical variable that underlies the dynamic organization of the honeybee society (Jeffree and Allen, 1956). Order can be replaced by disorder, consequently, if the number of viable and diverse worker phenotypes declines. Irreversible failure to keep up the production of new worker bees can occur as a result of insufficient sperm transfer to the queen's spermatheca after the completion of her mating flights, and may also arise because of inbreeding.
Postmating sperm migration from the queen's lateral oviducts into the spermatheca is a wasteful process, and the polyandrous mating strategy of the honeybee may be a means of which to ensure that the spermatheca becomes adequately filled (Kraus et al., 2004). If this filling is incomplete, it is probable that the queen's sperm store is depleted prematurely. Once the availability of sperm decreases to critical levels, the nest will soon contain an increasing proportion of unfertilized eggs that develop into haploid drones. It is unlikely that the worker population will attempt to replace the queen with a new virgin when this situation arises during the queen's first year of life (Collins, 2004), see also Section V. Consequently, colony growth is repressed dramatically.
Growth is also inhibited in inbreed populations, which are characterized by reduced brood viability (Beye et al., 2003). Homozygosis at the csd locus is one factor that contributes to reduced amounts of worker brood in inbred groups. Yet, in addition to the constraints that emerge from production of diploid drones (Section V), reduced viability is also found for immature females of inbred strains (Fuchs and Schade, 1994). These individuals develop growth abnormalities (Simôes, personal communication) that in many cases resemble tumorous outgrowths (Fig. 5). Due to the surveillance systems of the colony (Masterman et al., 2001), however, these embryos that die within the egg membrane or hatch as abnormal larvae are efficiently removed. Larvae with growth abnormalities, therefore, are typically only observed in experiments that involve in vitro rearing (Bergem and Kaftanoglu, personal communication). These setups have further shown that embryos exposed to UV light develop morphological characteristics similar to some of those observed in inbred strains (Figs. 5A vs 6B). UV light is a key environmental carcinogen (Maglio et al., 2005). Therefore, it may deserve further attention that UV exposure probably induces mutations (Fig. 6) that worker bees can detect by use of olfaction.
Fig. 5.
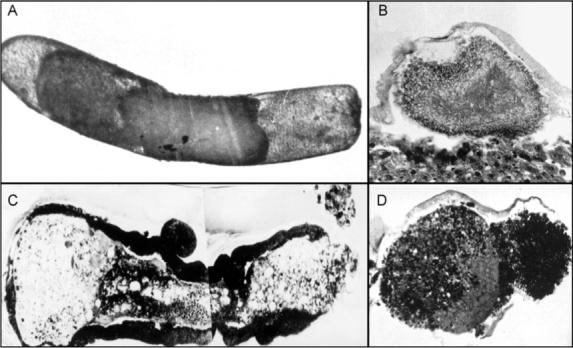
Growth abnormalities in embryos of an inbred strain. Embryos are stained with hematoxylin/eosin. Abnormal development within the egg membrane (A) (photo at 10×) can lead to death of the embryo and the egg never hatches. An older embryo shows severe growth abnormalities (B) (two superimposed photos at 16×) with tumor-like outgrowths (C and D) (close ups at 40×). Resulting larvae may survive for a limited time in vitro (M. Bergem, personal communication). In the colony setting, similar individuals appear to be rapidly killed by adult workers. Image courtesy of Z. L. P. Simôes.
Fig. 6.
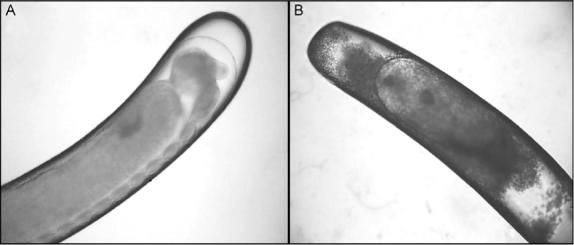
Embryos that were left untreated or UV irradiated (0.1 second at 1 J/cm2) when 4-hour old (photos at 10×). Untreated honeybee embryos (A) show normal development 54 hours after collection, whereas irradiated embryos (B) after 54 hours develop abnormally. Image courtesy of M. Bergem.
Several mechanisms fundamental to the emergent order of the honeybee society fail in the small units that develop as the number of worker bees declines. The colonies' defense systems are destabilized (Schneider and McNally, 1992), the workers become more accepting of abnormal individuals including diploid drones (Polaczek et al., 2000), the colonies' foraging efforts are less efficient (Eckert et al., 1994), and the ability to regulate the nest environment can collapse (Omholt, 1987). The proximate mechanisms responsible for the collective loss of surveillance functions are not well understood compared to the principles that underlie the foraging and thermal properties of healthy honeybee colonies (Anderson and Ratnieks, 1999; Bourke, 1999; Eckert et al., 1994; Free and Racey, 1968; Fukuda, 1983; Seeley, 1995; Wilson et al., 1999). Yet, it is clear that features of the social connectivity of the honeybee society are compromised in units that are very small (Villumstad, 1977). Associated weakening of colony-level functions, such as the defensive response toward abnormal or foreign individuals, increase susceptibility to diseases and raids against the colony's food stores (Atkins et al., 1975; Morse and Nowogrodzki, 1990; Winston, 1987). Loss of homeostatic mechanisms further enhances the risk of superorganismal death in response to changes in ambient conditions (Lee and Winston, 1987).
B. Social Cancer
The reproductive capability of individual worker bees is normally suppressed by pheromone-driven inhibition of oogenesis and nest surveillance schemes enforced by the worker population (Section V). This system can be compromised, however (Martin et al., 2002). The order of the honeybee society is subsequently torn apart by uncontrolled replication of a malignant worker phenotype, a situation that is comparable to a lethal social cancer (Oldroyd, 2002).
Uncontrolled worker replication is initiated if an Apis mellifera capensis (Cape honeybee) worker is able to enter an Apis mellifera scutellata (African honeybee) host colony. No special mechanisms exist that allow A. m. capensis workers to enter the nest, but a key difference between A. m. capensis and all other honeybee subspecies is that the worker caste can reproduce via thelytokous parthenogenesis (Ratnieks, 1988). Thereby, if an unmated worker lays an egg, it can develop into a fully viable diploid replicate of its mother or, if sufficiently nourished, the offspring can grow to become a queen. The ovary of A. m. capensis workers develop rapidly inside the host society, probably implying that these bees do not respond to the A. m. scutellata queen and larval pheromones that normally suppress worker oogenesis (Martin et al., 2002). Additionally, A. m. capensis worker bees provide their eggs with a pheromone coating that mimics that of eggs laid by the A. m. scutellata queen (Section V). Thereby, their offspring is readily accepted by the A. m. scutellata host. This ability to escape detection is in some ways analogous to tumor immune evasion by expression of self-antigens by the tumor cells (Houghton and Guevara-Patino, 2004; Kortylewski et al., 2005).
When reared by A. m. scutellata nurse bees, A. m. capensis brood elicits more food than the host's own larvae (Calis et al., 2002). The resulting A. m. capensis adults often have enlarged ovaries and other morphological and pheromonal characteristics similar to that of queens (Section III). This class of offspring does not engage in collective division of labor, but instead the workers feed off the host society's food stores and start laying their own eggs after as little as 6 days (Martin et al., 2002). The A. m. scutellata queen is killed by the worker bees within 5−6 weeks, and the society subsequently breaks down when the remaining population of A. m. scutellata workers becomes so small that the social connectivity of the colony collapses (Fig. 7). As the host's brood rearing, foraging, and defensive efforts thereby come to an end, the A. m. capensis workers disperse in search of new A. m. scutellata colonies.
Fig. 7.
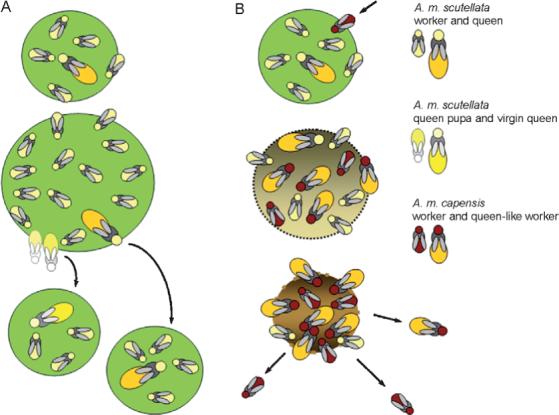
Normal colony development versus social cancer. The propagation unit for a honeybee colony is the swarm (A). Swarming (splitting) occurs after the colony unit has grown to a certain size. Worker bees produce new queen larvae and prior to the adult emergence of the first virgin, the old queen leaves to find a new colony together with a fraction of the worker population. The malignant A. m. capensis phenotype enters a host nest (B), overrides the control mechanisms of the society, and starts laying diploid eggs by thelytokous parthenogenesis. Gradually the resulting female clones, which do work, break down the social integrity of the society and the host colony perishes. A. m. capensis subsequently disperse in search of new host colonies. In many ways, this process is comparable to metastasis.
Note that in the native maternal nest, replication of A. m. capensis worker bees does not come out of control—although the worker caste can lay eggs by thelytokous parthenogenesis as it does in A. m. scutellata host societies. This is probably because all worker offsprings are nourished normally (Beekman and Oldroyd, 2003; Calis et al., 2002). The resulting adults, therefore, are not queen-like and display normal social phenotypes that engage in collective division of labor. Genetic analysis has further shown that all workers with the malignant A. m. capensis phenotype, although collected from different A. m. scutellata colonies, are the parthenogenetic descendants of a single bee (Moritz, 2002). It has been proposed that this clonal population emerged from an extraordinary A. m. scutellata nest infestation; one A. m. capensis worker genotype suited to evade the full array of control mechanisms inherent to the host society.
Within European honeybee populations (A. m. mellifera), a more benign dynamic arises through the behavior of so called “anarchistic workers” (Beekman and Oldroyd, 2003; Oldroyd et al., 1999). Anarchistic bees activate their ovaries despite the presence of inhibitory pheromones from the queen and brood, and they lay unfertilized eggs that are not disposed of by nestmates (Hoover et al., 2003, 2005). Although a considerable number of the drones in such colonies may stem from eggs laid by workers, the queen continues to be the dominant female reproductive. Thus, the society does not progress into the destructive state that emerges under A. m. capensis infestation. Some colony-level functionality may still be lost, however, because anarchistic bees do not work as hard as normal worker bees (Dampney et al., 2004).
VII. CONCLUSIONS
The amazing order of social insect societies is a mass phenomenon that emerges from integration of much simpler individual patterns by means of signal transduction or social communication. We have described how the honeybee superorganism uses tactile, thermal, and chemosensory cues to control growth, differentiation, homeostasis, and death. Individual bees perceive these signals through sensory receptors on their antennae, feet, and proboscis (tongue) (Gould and Gould, 1988; Snodgrass, 1956), and the subsequent integration of peripheral stimuli in the honeybee brain is an area that receives much attention (Belzunces et al., 1996; Dacher et al., 2005; Farooqui et al., 2004; Scheiner et al., 2002).
Chemosensory cues, in particular, play an important role in honeybee social organization (Sections IV–VI), having powerful effects on individual gene expression patterns (Grozinger et al., 2003), behavior (Leoncini et al., 2004b; Pankiw, 2004a; Pankiw and Page, 2003), and physiology (Hoover et al., 2003; Huang and Robinson, 1992, 1996). A unique phenomenon that emerges from integration of social chemosensory cues is the behavioral and physiological differentiation of temporal worker forms (Section IV). The shift from nurse tasks to foraging duties is intriguing because it demonstrates that programmed cell death can be released by signals transferred between individuals (Amdam et al., 2005). Programmed cell death in addition is observed in the ovary of workers inhibited by pheromones from queen and brood (Capella and Hartfelder, 1998), possibly explaining how worker reproduction is controlled. The idea that worker propagation is conditionally suppressed through an apoptosis pathway that is governed by pheromones raises interesting questions relative to the casual basis of anarchistic workers and malignant A. m. capensis phenotypes. We believe that the honeybee in this connection can provide valuable insights into molecular mechanisms that enable interindividual signals to translate into apoptotic responses in target tissues. Such insights may be of general interest since apoptosis contributes to the antitumor activity of many chemotherapeutic drugs (Fisher, 1994; Johnstone et al., 2002).
Insights into honeybee chemosensory signaling are fundamental also for understanding how the biological order of the society can fail. Specifically, destructive dynamics that turn order into disorder are intimately linked to a progressive loss of integration of chemosensory cues—at the level of individual bees and the colony unit. We have described outcomes that emerge when workers responsible for social connectivity become few or when malignant phenotypes override the regulatory machinery of the colony and subsequently replicate in an uncontrollable manner. The latter is the social cancer of A. m. scutellata that is regarded as lethal. Similar infestations by malignant A. m. capensis phenotypes occurred in 1928 and 1977 but eventually petered out (Oldroyd, 2002). The causes of these remissions are not understood but suggest that the dynamics between the current clone and its host may change in future—creating possibilities for insights into how aberrant replication can be restrained and order restored at the level of a superorganism.
Approximately 50 substances synthesized by honeybee queens, workers, and brood have been identified as having functional roles in social organization (Pankiw, 2004b). Remarkable physiological effects of such compounds have been a driving force of studies exploring if interindividual dynamics can translate into patterns of cell growth and cell death (Amdam et al., 2005). Such findings, along with the availability of the honeybee genome sequence, open up for new insights into how emergent biological order arises through control of individual molecular pathways (Robinson et al., 2005). In this context, our chapter is a first initiative to underline that the honeybee, in addition, can become a model for understanding the frailty of underlying regulatory systems—thereby increasing the knowledge of principles of disorder.
ACKNOWLEDGMENTS
We thank M. Amdam, U. Gimsa, and M. Laubichler for helpful reviews of the chapter. We are grateful to M. Bergem, J. Sahertian, Z. L. P. Simôes, and G. Tuven for contributions to illustrations. G.V.A. and S.C.S. were supported by Norwegian Research Council #147085/110 and #171958. Funding was also provided by National Institute on Aging PO1 AG22500 to G. .A.
REFERENCES
- Aase ALTO, Amdam GV, Hagen A, Omholt SW. A new method for rearing genetically manipulated honey bee workers. Apidologie. 2005;36:293–299. [Google Scholar]
- Amdam GV, Omholt SW. The regulatory anatomy of honeybee lifespan. J. Theor. Biol. 2002;216:209–228. doi: 10.1006/jtbi.2002.2545. [DOI] [PubMed] [Google Scholar]
- Amdam GV, Omholt SW. The hive bee to forager transition in honeybee colonies: The double repressor hypothesis. J. Theor. Biol. 2003;223:451–464. doi: 10.1016/s0022-5193(03)00121-8. [DOI] [PubMed] [Google Scholar]
- Amdam GV, Page RE. Intergenerational transfers may have decoupled physiological and chronological age in a eusocial insect. Aging Res. Rev. 2005;4:398–408. doi: 10.1016/j.arr.2005.03.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amdam GV, Simões ZLP, Guidugli KR, Norberg K, Omholt SW. Disruption of vitellogenin gene function in adult honeybees by intra-abdominal injection of double-stranded RNA. BMC Biotechnol. 2003;3:1–8. doi: 10.1186/1472-6750-3-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amdam GV, Norberg K, Fondrk MK, Page RE. Reproductive ground plan may mediate colony-level selection effects on individual foraging behavior in honey bees. Proc. Natl. Acad. Sci. USA. 2004a;101:11350–11355. doi: 10.1073/pnas.0403073101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amdam GV, Simões ZLP, Hagen A, Norberg K, Schrøder K, Mikkelsen O, Kirkwood TBL, Omholt SW. Hormonal control of the yolk precursor vitellogenin regulates immune function and longevity in honeybees. Exp. Gerontol. 2004b;39:767–773. doi: 10.1016/j.exger.2004.02.010. [DOI] [PubMed] [Google Scholar]
- Amdam GV, Aase ALTO, Seehuus SC, Norberg K, Hartfelder K, Fondrk MK. Social reversal of immunosenescence in honey bee workers. Exp. Gerontol. 2005;40:939–947. doi: 10.1016/j.exger.2005.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson C, Ratnieks FLW. Task partitioning in insect societies. I. Effect of colony size on queueing delay and colony ergonomic efficiency. Am. Nat. 1999;154:521–529. doi: 10.1086/303255. [DOI] [PubMed] [Google Scholar]
- Atkins EL, Banker R, Butler CG, Cale GH, Cale GH, Jr., Crane E, Dadant CC. The Hive and the Honey Bee. Datant & Sons; Hamilton, Illinois: 1975. [Google Scholar]
- Baird DH, Seeley TD. An equilibrium theory of queen production in honeybee colonies preparing to swarm. Behav. Ecol. Sociobiol. 1983;13:221–228. [Google Scholar]
- Balmain A. Cancer as a complex genetic trait: Tumor susceptibility in humans and mouse models. Cell. 2002;108:145–152. doi: 10.1016/s0092-8674(02)00622-0. [DOI] [PubMed] [Google Scholar]
- Bedick JC, Tunaz H, Aliza ARN, Putnam SM, Ellis MD, Stanley DW. Eicosanoids act in nodulation reactions to bacterial infections in newly emerged adult honey bees, Apis mellifera, but not in older foragers. Comp. Biochem. Physiol. 2001;130:107–117. doi: 10.1016/s1532-0456(01)00226-5. [DOI] [PubMed] [Google Scholar]
- Beekman M, Oldroyd BP. Effects of cross-feeding anarchistic and wild type honey bees: Anarchistic workers are not queen-like. Naturwissenschaften. 2003;90:189–192. doi: 10.1007/s00114-003-0406-7. [DOI] [PubMed] [Google Scholar]
- Belzunces LP, Vandame R, Gu XF. Modulation of honey bee thermo-regulation by adrenergic compounds. Neuroreport. 1996;7:1601–1604. doi: 10.1097/00001756-199607080-00014. [DOI] [PubMed] [Google Scholar]
- Beye M, Hartel S, Hagen A, Hasselmann M, Omholt SW. Specific developmental gene silencing in the honey bee using a homeobox motif. Insect Mol. Biol. 2002;11:527–532. doi: 10.1046/j.1365-2583.2002.00361.x. [DOI] [PubMed] [Google Scholar]
- Beye M, Hasselmann M, Fondrk MK, Page RE, Omholt SW. The gene csd is the primary signal for sexual development in the honeybee and encodes an SR-type protein. Cell. 2003;114:419–429. doi: 10.1016/s0092-8674(03)00606-8. [DOI] [PubMed] [Google Scholar]
- Bishop JM. Molecular themes in oncogenesis. Cell. 1991;64:235–248. doi: 10.1016/0092-8674(91)90636-d. [DOI] [PubMed] [Google Scholar]
- Bloch G, Wheeler D, Robinson GE. Endocrine influences on the organization of insect societies. In: Pfaff D, Arnold A, Etgen A, Fahrbach S, Moss R, Rubin R, editors. Hormones, Brain and Behavior. Vol. 3. Academic Press; San Diego: 2002. pp. 195–235. [Google Scholar]
- Boecking O, Spivak M. Behavioral defenses of honey bees against Varroa jacobsoni Oud. Apidologie. 1999;30:141–158. [Google Scholar]
- Bonabeau E, Dorigo M, Theraulaz G. Inspiration for optimization from social insect behaviour. Nature. 2000;406:39–42. doi: 10.1038/35017500. [DOI] [PubMed] [Google Scholar]
- Bourke AFG. Colony size, social complexity and reproductive conflict in social insects. J. Evol. Biol. 1999;12:245–257. [Google Scholar]
- Bühler A, Lanzrein B, Wille H. Influence of temperature and carbon dioxide concentration on juvenile hormone titre and dependent parameters of adult worker honey bees (Apis mellifera L.). J. Insect Physiol. 1983;29:885–893. [Google Scholar]
- Calis JNM, Boot WJ, Allsopp MH, Beekman M. Getting more than a fair share: Nutrition of worker larvae related to social parasitism in the Cape honey bee Apis mellifera capensis. Apidologie. 2002;33:193–202. [Google Scholar]
- Camazine S, Deneubourg JL, Franks NR, Sneyd J, Theraulaz G, Bonabeau E. Self-Organization in Biological Systems. Princeton University Press; Princeton, NJ: 2001. [Google Scholar]
- Capella ICS, Hartfelder K. Juvenile hormone effect on DNA synthesis and apoptosis in caste-specific differentiation of the larval honey bee (Apis mellifera L.) ovary. J. Insect Physiol. 1998;44:385–391. doi: 10.1016/s0022-1910(98)00027-4. [DOI] [PubMed] [Google Scholar]
- Chaline N, Sandoz JC, Martin SJ, Ratnieks FLW, Jones GR. Learning and discrimination of individual cuticular hydrocarbons by honeybees (Apis mellifera). Chem. Senses. 2005;30:327–335. doi: 10.1093/chemse/bji027. [DOI] [PubMed] [Google Scholar]
- Collins AM. Functional longevity of honey bee, Apis mellifera, queens inseminated with low viability semen. J. Apic. Res. 2004;43:167–171. [Google Scholar]
- Crailsheim K. The protein balance of the honey bee worker. Apidologie. 1990;21:417–429. [Google Scholar]
- Crailsheim K, Schneider LHW, Hrassnigg N, Bühlmann G, Brosch U, Gmeinbauer R, Schöffmann B. Pollen consumption and utilization in worker honeybees (Apis mellifera carnica): Dependence on individual age and function. J. Insect Physiol. 1992;38:409–419. [Google Scholar]
- Crailsheim K, Riessberger U, Blaschon B, Nowogrodzki R, Hrassnigg N. Short-term of simulated bad weather conditions upon the behaviour of food-storer honeybees during day and night (Apis mellifera carnica Pollmann). Apidologie. 1999;30:299–310. [Google Scholar]
- Dacher M, Lagarrigue A, Gauthier M. Antennal tactile learning in the honeybee: Effect of nicotinic antagonists on memory dynamics. Neuroscience. 2005;130:37–50. doi: 10.1016/j.neuroscience.2004.09.006. [DOI] [PubMed] [Google Scholar]
- Dampney JR, Barron AB, Oldroyd BP. Measuring the cost of worker reproduction in honeybees: Work tempo in an “anarchic” line. Apidologie. 2004;35:83–88. [Google Scholar]
- Dani FR, Jones GR, Corsi S, Beard R, Pradella D, Turillazzi S. Nestmate recognition cues in the honey bee: Differential importance of cuticular alkanes and alkenes. Chem. Senses. 2005;30:477–489. doi: 10.1093/chemse/bji040. [DOI] [PubMed] [Google Scholar]
- De Moraes R, Bowen ID. Modes of cell death in the hypopharyngeal gland of the honey bee (Apis mellifera L). Cell Biol. Int. 2000;24:737–743. doi: 10.1006/cbir.2000.0534. [DOI] [PubMed] [Google Scholar]
- Deseyn J, Billen J. Age-dependent morphology and ultrastructure of the hypopharyngeal gland of Apis mellifera workers (Hymenoptera, Apidae). Apidologie. 2005;36:49–57. [Google Scholar]
- Dor R, Katzav-Gozansky T, Hefetz A. Dufour's gland pheromone as a reliable fertility signal among honeybee (Apis mellifera) workers. Behav. Ecol. Sociobiol. 2005;58:270–276. [Google Scholar]
- Doums C, Moret Y, Benelli E, Schmid-Hempel P. Senescence of immune defence in Bombus workers. Ecol. Entomol. 2002;27:138–144. [Google Scholar]
- Ebadi R, Gary NE, Lorenzen K. Effects of carbon dioxide and low temperature narcosis on honey bees, Apis mellifera. Environ. Entomol. 1980;9:144–147. [Google Scholar]
- Eckert CD, Winston ML, Ydenberg RC. The relationship between population size, amount of brood, and individual foraging behaviour in the honey bee, Apis mellifera L. Oecologia. 1994;97:248–255. doi: 10.1007/BF00323157. [DOI] [PubMed] [Google Scholar]
- Evans JD, Wheeler DE. Differential gene expression between developing queens and workers in the honey bee, Apis mellifera. Proc. Natl. Acad. Sci. USA. 1999;96:5575–5580. doi: 10.1073/pnas.96.10.5575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farooqui T, Vaessin H, Smith BH. Octopamine receptors in the honeybee (Apis mellifera) brain and their disruption by RNA-mediated interference. J. Insect Physiol. 2004;50:701–713. doi: 10.1016/j.jinsphys.2004.04.014. [DOI] [PubMed] [Google Scholar]
- Fewell JH. Social insect networks. Science. 2003;301:1867–1870. doi: 10.1126/science.1088945. [DOI] [PubMed] [Google Scholar]
- Fewell JH, Winston ML. Colony state and regulation of pollen foraging in the honey bee, Apis mellifera L. Behav. Ecol. Sociobiol. 1992;30:387–393. [Google Scholar]
- Fisher DE. Apoptosis in cancer-therapy—crossing the threshold. Cell. 1994;78:539–542. doi: 10.1016/0092-8674(94)90518-5. [DOI] [PubMed] [Google Scholar]
- Fluri P, Wille H, Gerig L, Lüscher M. Juvenile hormone, vitellogenin and haemocyte composition in winter worker honeybees (Apis mellifera). Experientia. 1977;33:1240–1241. [Google Scholar]
- Fluri P, Lüscher M, Wille H, Gerig L. Changes in weight of the pharyngeal gland and haemolymph titres of juvenile hormone, protein and vitellogenin in worker honey bees. J. Insect Physiol. 1982;28:61–68. [Google Scholar]
- Franssens V, Simonet G, Bronckaers A, Claeys I, De Loof A, Broeck JV. Eicosanoids mediate the laminarin-induced nodulation response in larvae of the flesh fly, Neobellieria bullata. Arch. Insect Biochem. Physiol. 2005;59:32–41. doi: 10.1002/arch.20053. [DOI] [PubMed] [Google Scholar]
- Free JB. Pheromones of Social Bees. Chapman & Hall; London: 1987. [Google Scholar]
- Free JB, Racey PA. The effect of the size of honeybee colonies on food consumption, brood rearing and the longevity of the bees during winter. Ent. Exp. & Appl. 1968;11:241–249. [Google Scholar]
- Fuchs S, Schade V. Lower performance in honeybee colonies of uniform paternity. Apidologie. 1994;25:155–168. [Google Scholar]
- Fukuda H. The relationship between work efficiency and population size in a honeybee colony. Res. Popul. Ecol. 1983;25:249–263. [Google Scholar]
- Gerber B, Wustenberg D, Schutz A, Menzel R. Temporal determinants of olfactory long-term retention in honeybee classical conditioning: Nonmonotonous effects of the training trial interval. Neurobiol. Learn. Mem. 1998;69:71–78. doi: 10.1006/nlme.1997.3801. [DOI] [PubMed] [Google Scholar]
- Golstein P, Aubry L, Levraun JP. Cell-death alternative model organisms: Why and which? Mol. Cell. Biol. 2003;4:1–10. doi: 10.1038/nrm1224. [DOI] [PubMed] [Google Scholar]
- Gould JL, Gould CG. The Honey Bee. Scientific American Library; New York: 1988. [Google Scholar]
- Gramacho KP, Spivak M. Differences in olfactory sensitivity and behavioral responses among honey bees bred for hygienic behavior. Behav. Ecol. Sociobiol. 2003;54:472–479. [Google Scholar]
- Grodzicki P, Caputa M. Social versus individual behaviour: A comparative approach to thermal behaviour of the honeybee (Apis mellifera L.) and the American cockroach (Periplaneta americana L.). J. Insect Physiol. 2005;51:315–322. doi: 10.1016/j.jinsphys.2005.01.001. [DOI] [PubMed] [Google Scholar]
- Grozinger CM, Sharabash NM, Whitfield CW, Robinson GE. Pheromone- mediated gene expression in the honey bee brain. Proc. Natl. Acad. Sci. USA. 2003;100:14519–14525. doi: 10.1073/pnas.2335884100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guidugli KR, Nascimento AM, Amdam GV, Barchuk AR, Angel R, Omholt SW, Simões ZLP, Hartfelder K. Vitellogenin regulates hormonal dynamics in the worker caste of a eusocial insect. FEBS Lett. 2005;579:4961–4965. doi: 10.1016/j.febslet.2005.07.085. [DOI] [PubMed] [Google Scholar]
- Hartfelder K, Engels W. Social insect polymorphism: Hormonal regulation of plasticity in development and reproduction in the honeybee. Curr. Top. Dev. Biol. 1998;40:45–77. doi: 10.1016/s0070-2153(08)60364-6. [DOI] [PubMed] [Google Scholar]
- Herrmann M, Trenzcek T, Fahrenhorst H, Engels W. Characters that differ between diploid and haploid honey bee (Apis mellifera) drones. Genet. Mol. Res. 2005;4:624–641. [PubMed] [Google Scholar]
- Hoeijmakers JHJ. Genome maintenance mechanisms for preventing cancer. Nature. 2001;411:366–374. doi: 10.1038/35077232. [DOI] [PubMed] [Google Scholar]
- Hölldobler B. The chemistry of social regulation: Multicomponent signals in ant societies. Proc. Natl. Acad. Sci. USA. 1995;92:19–22. doi: 10.1073/pnas.92.1.19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoover SER, Keeling CI, Winston ML, Slessor KN. The effect of queen pheromones on worker honey bee ovary development. Naturwissenschaften. 2003;90:477–480. doi: 10.1007/s00114-003-0462-z. [DOI] [PubMed] [Google Scholar]
- Hoover SER, Winston ML, Oldroyd BP. Retinue attraction and ovary activation: Responses of wild type and anarchistic honey bees (Apis mellifera) to queen and brood pheromones. Behav. Ecol. Sociobiol. 2005;59:278–284. [Google Scholar]
- Houghton AN, Guevara-Patino JA. Immune recognition of self in immunity against cancer. J. Clin. Invest. 2004;114:468–471. doi: 10.1172/JCI22685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang Z-Y, Robinson GE. Honeybee colony integration: Worker-worker interactions mediate hormonally regulated plasticity in division of labor. Proc. Natl. Acad. Sci. USA. 1992;89:11726–11729. doi: 10.1073/pnas.89.24.11726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang Z-Y, Robinson GE. Seasonal changes in juvenile hormone titers and rates of biosynthesis in honey bees. J. Comp. Physiol. [B] 1995;165:18–28. doi: 10.1007/BF00264682. [DOI] [PubMed] [Google Scholar]
- Huang Z-Y, Robinson GE. Regulation of honey bee division of labor by colony age demography. Behav. Ecol. Sociobiol. 1996;39:147–158. [Google Scholar]
- Humphries MA, Muller U, Fondrk MK, Page RE. PKA and PKC content in the honey bee central brain differs in genotypic strains with distinct foraging behavior. J. Comp. Physiol. [A] 2003;189:555–562. doi: 10.1007/s00359-003-0433-z. [DOI] [PubMed] [Google Scholar]
- Jeffree EP, Allen MD. The Influence of colony size and of Nosema disease on the rate of population loss in honey bee colonies in winter. J. Econ. Entomol. 1956;49:831–834. [Google Scholar]
- Johnstone RW, Ruefli AA, Lowe SW. Apoptosis: A link between cancer genetics and chemotherapy. Cell. 2002;108:153–164. doi: 10.1016/s0092-8674(02)00625-6. [DOI] [PubMed] [Google Scholar]
- Jones JC, Myerscough MR, Graham S, Oldroyd BP. Honey bee nest thermoregulation: Diversity promotes stability. Science. 2004;305:402–404. doi: 10.1126/science.1096340. [DOI] [PubMed] [Google Scholar]
- Kaestner A. Lehrbuch der speziellen Zoologie. G. Fischer Verlag; Stuttgart: 1969. [Google Scholar]
- Kamakura M, Sakaki T. A hypopharyngeal gland protein of the worker honeybee Apis mellifera L. enhances proliferation of primary-cultured rat hepatocytes and suppresses apoptosis in the absence of serum. Protein Expr. Purif. 2005;45:307–314. doi: 10.1016/j.pep.2005.08.004. [DOI] [PubMed] [Google Scholar]
- Karin M, Greten FR. NF kappa B: Linking inflammation and immunity to cancer development and progression. Nat. Rev. Immunol. 2005;5:749–759. doi: 10.1038/nri1703. [DOI] [PubMed] [Google Scholar]
- Katzav-Gozansky T, Soroker V, Ibarra F, Francke W, Hefetz A. Dufour's gland secretion of the queen honeybee (Apis mellifera): An egg discriminator pheromone or a queen signal? Behav. Ecol. Sociobiol. 2001;51:76–86. [Google Scholar]
- Katzav-Gozansky T, Soroker V, Kamer J, Schulz CM, Francke W, Hefetz A. Ultrastructural and chemical characterization of egg surface of honeybee worker and queen- laid eggs. Chemoecology. 2003;13:129–134. [Google Scholar]
- Keeling CI, Slessor KN, Higo HA, Winston ML. New components of the honey bee (Apis mellifera L.) queen retinue pheromone. Proc. Natl. Acad. Sci. USA. 2003;100:4486–4491. doi: 10.1073/pnas.0836984100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ken T, Hepburn HR, Radloff SE, Yusheng Y, Yiqiu L, Danyin Z, Neumann P. Heat-balling wasps by honeybees. Naturwissenschaften. 2005;92:492–495. doi: 10.1007/s00114-005-0026-5. [DOI] [PubMed] [Google Scholar]
- Koehler A. Beobachtungen über Veranderungen am Fettkörper der Biene. Schweiz. Bienen-Zeitung. 1921;44:424–428. [Google Scholar]
- Kortylewski M, Kujawski M, Wang TH, Wei S, Zhang SM, Pilon-Thomas S, Niu GL, Kay H, Mule J, Kerr WG, Jove R, Pardoll D, et al. Inhibiting Stat3 signaling in the hematopoietic system elicits multicomponent antitumor immunity. Nat. Med. 2005;11:1314–1321. doi: 10.1038/nm1325. [DOI] [PubMed] [Google Scholar]
- Kovac H, Crailsheim K. Lifespan of Apis mellifera carnica Pollm. infested by Varroa jacobsoni Oud. in relation to season and extent of infestation. J. Apic. Res. 1988;27:230–238. [Google Scholar]
- Kraaijeveld AR, Limentani EC, Godfray HCJ. Basis of the trade-off between parasitoid resistance and larval competitive ability in Drosophila melanogaster. Proc. R. Soc. Lond. B. 2001;268:259–261. doi: 10.1098/rspb.2000.1354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kraus FB, Neumann P, van Praagh J, Moritz RFA. Sperm limitation and the evolution of extreme polyandry in honeybees (Apis mellifera L.). Behav. Ecol. Sociobiol. 2004;55:494–501. [Google Scholar]
- Lee PC, Winston ML. Effects of reproductive timing and colony size on the survival, offspring colony size and drone production in the honey bee (Apis mellifera). Ecol. Entomol. 1987;12:187–195. [Google Scholar]
- Leoncini I, Crauser D, Robinson GE, Le Conte Y. Worker-worker inhibition of honey bee behavioural development independent of queen and brood. Insect. Soc. 2004a;51:392–394. [Google Scholar]
- Leoncini I, Le Conte Y, Costagliola G, Plettner E, Toth AL, Wang MW, Huang Z, Becard JM, Crauser D, Slessor KN, Robinson GE. Regulation of behavioral maturation by a primer pheromone produced by adult worker honey bees. Proc. Natl. Acad. Sci. USA. 2004b;101:17559–17564. doi: 10.1073/pnas.0407652101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maeterlinck M. The Life of the Bee. Dodd, Mead and Company; New York: 1901. [Google Scholar]
- Maglio DHG, Paz ML, Ferrari A, Weill FS, Czerniczyniec A, Leoni J, Bustamante J. Skin damage and mitochondrial dysfunction after acute ultraviolet B irradiation: Relationship with nitric oxide production. Photodermatol. Photoimmunol. Photomed. 2005;21:311–317. doi: 10.1111/j.1600-0781.2005.00185.x. [DOI] [PubMed] [Google Scholar]
- Martin SJ, Beekman M, Wossler TC, Ratnieks FLW. Parasitic Cape honeybee workers, Apis mellifera capensis, evade policing. Nature. 2002;415:163–165. doi: 10.1038/415163a. [DOI] [PubMed] [Google Scholar]
- Masterman R, Ross R, Mesce K, Spivak M. Olfactory and behavioral response thresholds to odors of diseased brood differ between hygienic and non-hygienic honey bees (Apis mellifera L.). J. Comp. Physiol. [A] 2001;187:441–452. doi: 10.1007/s003590100216. [DOI] [PubMed] [Google Scholar]
- Maurizio A. The influence of pollen feeding and brood rearing on the length of life and physiological condition of the honeybee preliminary report. Bee World. 1950;31:9–12. [Google Scholar]
- Maurizio A. Pollenernahrung und Lebensvorgange bei der Honigbiene (Apis mellifera L.). Landwirtsch. Jahrb. Schweiz. 1954;245:115–182. [Google Scholar]
- Meled M, Thrasyvoulou A, Belzunces LP. Seasonal variations in susceptibility of Apis mellifera to the synergistic action of prochloraz and deltamethrin. Environ. Toxicol. Chem. 1998;17:2517–2520. [Google Scholar]
- Menzel R. Behavioural access to short-term memory in bees. Nature. 1979;281:368–369. doi: 10.1038/281368a0. [DOI] [PubMed] [Google Scholar]
- Millar DA, Ratcliffe NA. Invertebrates. In: Turner RJ, editor. Immunology: A Comparative Approach. John Wiley & Son; West Sussex: 1994. pp. 29–68. [Google Scholar]
- Mishima S, Ono Y, Araki Y, Akao Y, Nozawa Y. Two related cinnamic acid derivatives from Brazilian honey bee propolis, baccharin and drupanin, induce growth inhibition in allografted sarcoma S-180 in mice. Biol. Pharm. Bull. 2005;28:1025–1030. doi: 10.1248/bpb.28.1025. [DOI] [PubMed] [Google Scholar]
- Mitchell SD. Biological Complexity and Integrative Pluralism. Cambridge University Press; Cambridge: 2003. [Google Scholar]
- Moritz RFA. Population dynamics of the Cape bee phenomenon: The impact of parasitic laying worker clones in apiaries and natural populations. Apidologie. 2002;33:233–244. [Google Scholar]
- Morse RA, Nowogrodzki R. Honey Bee Pests, Predators, and Diseases. Cornell University Press; Ithaca, NY: 1990. [Google Scholar]
- Naiem E-S, Hrassnigg N, Crailsheim K. Nurse bees support the physiological development of young bees (Apis mellifera L.). J. Comp. Physiol. [B] 1999;169:271–279. [Google Scholar]
- Ohashi K, Natori S, Kubo T. Change in the mode of gene expression of the hypopharyngeal gland cells with an age-dependent role change of the worker honeybee Apis mellifera L. Eur. J. Biochem. 1997;249:797–802. doi: 10.1111/j.1432-1033.1997.t01-1-00797.x. [DOI] [PubMed] [Google Scholar]
- Oldroyd BP. The Cape honeybee: An example of a social cancer. TREE. 2002;17:249–251. [Google Scholar]
- Oldroyd BP, Halling L, Rinderer TE. Development and behaviour of anarchistic honeybees. Proc. R. Soc. Lon. Ser-A. 1999;266:1875–1878. [Google Scholar]
- Omholt SW. Thermoregulation in the winter cluster of the honeybee, Apis mellifera. J. Theor. Biol. 1987;128:219–231. [Google Scholar]
- Omholt SW, Amdam GV. Epigenic regulation of aging in honeybee workers. Sci. Aging Knowledge Environ. 2004;26:pe28. doi: 10.1126/sageke.2004.26.pe28. [DOI] [PubMed] [Google Scholar]
- Omholt SW, Rishovd S, Hagen A, Elmholdt O, Dalsgard B, Fromm S. Successful production of chimeric honeybee larvae. J. Exp. Zool. 1995;272:410–412. [Google Scholar]
- Page RE, Erber J. Levels of behavioral organization and the evolution of division of labor. Naturwissenschaften. 2002;89:91–106. doi: 10.1007/s00114-002-0299-x. [DOI] [PubMed] [Google Scholar]
- Page RE, Fondrk MK. The effects of colony-level selection on the social organization of honey bee (Apis mellifera L.) colonies: Colony-level components of pollen hoarding. Behav. Ecol. Sociobiol. 1995;36:135–144. [Google Scholar]
- Pankiw T. Worker honey bee pheromone regulation of foraging ontogeny. Naturwissenschaften. 2004a;91:178–181. doi: 10.1007/s00114-004-0506-z. [DOI] [PubMed] [Google Scholar]
- Pankiw T. Cued in: Honey bee pheromones as information flow and collective decision-making. Apidologie. 2004b;35:217–226. [Google Scholar]
- Pankiw T, Page RE. Effect of pheromones, hormones, and handling on sucrose response thresholds of honey bees (Apis mellifera L.). J. Comp. Physiol. [A] 2003;189:675–684. doi: 10.1007/s00359-003-0442-y. [DOI] [PubMed] [Google Scholar]
- Pankiw T, Page RE, Fondrk MK. Brood pheromone stimulates pollen foraging in honey bees (Apis mellifera). Behav. Ecol. Sociobiol. 1998;44:193–198. [Google Scholar]
- Pearce AN, Huang Z-Y, Breed MD. Juvenile hormone and aggression in honey bees. J. Insect Physiol. 2001;47:1243–1247. doi: 10.1016/s0022-1910(01)00109-3. [DOI] [PubMed] [Google Scholar]
- Pettis JS, Westcott LC, Winston ML. Balling behaviour in the honey bee in response to exogenous queen mandibular gland pheromone. J. Apic. Res. 1998;37:125–131. [Google Scholar]
- Pinto LZ, Bitondi MMG, Simões ZLP. Inhibition of vitellogenin synthesis in Apis mellifera workers by a juvenile hormone analogue, pyriproxyfen. J. Insect Physiol. 2000;46:153–160. doi: 10.1016/s0022-1910(99)00111-0. [DOI] [PubMed] [Google Scholar]
- Polaczek B, Neumann P, Schricker B, Moritz RFA. A new, simple method for rearing diploid drones in the honeybee (Apis mellifera L.). Apidologie. 2000;31:525–530. [Google Scholar]
- Potter CJ, Turenchalk GS, Xu T. Drosophila in cancer research—an expanding role. Trends Genet. 2000;16:33–39. doi: 10.1016/s0168-9525(99)01878-8. [DOI] [PubMed] [Google Scholar]
- Ratnieks FLW. Reproductive harmony via mutual policing by workers in eusocial hymenoptera. Am. Nat. 1988;132:217–236. [Google Scholar]
- Ratnieks FLW. Egg-laying, egg-removal, and ovary development by workers in queenright honey-bee colonies. Behav. Ecol. Sociobiol. 1993;32:191–198. [Google Scholar]
- Robinson GE, Page RE, Strambi C, Strambi A. Colony integration in honey bees: Mechanisms of behavioral reversion. Ethology. 1992;90:336–348. [Google Scholar]
- Robinson GE, Grozinger CM, Whitfield CW. Sociogenomics: Social life in molecular terms. Nat. Rev. Genet. 2005;6:257–270. doi: 10.1038/nrg1575. [DOI] [PubMed] [Google Scholar]
- Rutz W, Gerig L, Wille H, Lüscher M. A bioassay for juvenile hormone (JH) effects of insect growth regulators (IGR) on adult worker honeybees. Bull. Soc. Entomol. Suisse. 1974;47:307–313. [Google Scholar]
- Ryan KM, Ernst MK, Rice NR, Vousden KH. Role of NF-kappa B in p53-mediated programmed cell death. Nature. 2000;404:892–897. doi: 10.1038/35009130. [DOI] [PubMed] [Google Scholar]
- Santomauro G, Oldham NJ, Boland W, Engels W. Cannibalism of diploid drone larvae in the honey bee (Apis mellifera) is released by odd pattern of cuticular substances. J. Apic. Res. 2004;43:69–74. [Google Scholar]
- Schafer PW. Centrioles of a human cancer—intercellular order and intracellular disorder. Science. 1969;164:1300. doi: 10.1126/science.164.3885.1300. [DOI] [PubMed] [Google Scholar]
- Scheiner R, Page RE, Erber J. Responsiveness to sucrose affects tactile and olfactory learning in preforaging honey bees of two genetic strains. Behav. Brain Res. 2001;120:67–73. doi: 10.1016/s0166-4328(00)00359-4. [DOI] [PubMed] [Google Scholar]
- Scheiner R, Pluckhahn S, Oney B, Blenau W, Erber J. Behavioural pharmacology of octopamine, tyramine and dopamine in honey bees. Behav. Brain Res. 2002;136:545–553. doi: 10.1016/s0166-4328(02)00205-x. [DOI] [PubMed] [Google Scholar]
- Schmickl T, Crailsheim K. Inner nest homeostasis in a changing environment with special emphasis on honey bee brood nursing and pollen supply. Apidologie. 2004;35:249–263. [Google Scholar]
- Schneider SS, McNally LC. Colony defense in the African honey-bee in Africa (Hymenoptera, Apidae). Environ. Entomol. 1992;21:1362–1370. [Google Scholar]
- Seehuus SC, Norberg K, Gimsa U, Krekling T, Amdam GV. Reproductive protein protects sterile honey bee workers from oxidative stress. Proc. Natl. Acad. Sci. USA. 2006;103:962–967. doi: 10.1073/pnas.0502681103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seeley TD. Adaptive significance of the age polyethism schedule in honeybee colonies. Behav. Ecol. Sociobiol. 1982;11:287–293. [Google Scholar]
- Seeley TD. The Wisdom of the Hive. Harvard University Press; Cambridge, Massachusetts: 1995. [Google Scholar]
- Sekiguchi K, Sakagami SF. Structure of foraging population and related problems in the honeybee, with considerations on division of labour in bee colonies. Hakkaido Nat. Agric. Exp. Sta. Rep. 1966;Report No. 69:1–58. [Google Scholar]
- Severson DW. Swarming behavior of the honey bee. Am. Bee J. 1984;124:204. [Google Scholar]
- Shepard JL, Amatruda JF, Stern HM, Subramanian A, Finkelstein D, Ziai J, Finley KR, Pfaff KL, Hersey C, Zhou Y, Barut B, Freedman M, et al. Zebrafish bmyb mutation causes genome instability and increased cancer susceptibility. Proc. Natl. Acad. Sci. USA. 2005;102:13194–13199. doi: 10.1073/pnas.0506583102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Slessor KN, Winston ML, Le Conte Y. Pheromone communication in the honeybee (Apis mellifera L.). J. Chem. Ecol. 2005;31:2731–2745. doi: 10.1007/s10886-005-7623-9. [DOI] [PubMed] [Google Scholar]
- Snodgrass RE. Anatomy of the Honey Bee. Comstock Publishing Associates; Ithaca, New York: 1956. [Google Scholar]
- Spivak M. Honey bee hygienic behavior and defense against Varroa jacobsoni. Apidologie. 1996;27:245–260. [Google Scholar]
- Spivak M, Masterman R, Ross R, Mesce KA. Hygienic behavior in the honey bee (Apis mellifera L.) and the modulatory role of octopamine. J. Neurobiol. 2003;55:341–354. doi: 10.1002/neu.10219. [DOI] [PubMed] [Google Scholar]
- Theraulaz G, Bonabeau E, Nicolis SC, Solé RV, Fourcassié V, Blanco S, Fournier R, Joly J-L, Fernández P, Grimal A, Dalle P, Deneubourg JL. Spatial patterns in ant colonies. Proc. Natl. Acad. Sci. USA. 2002;99:9645–9649. doi: 10.1073/pnas.152302199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson CB. Apoptosis in the pathogenesis and treatment of disease. Science. 1995;267:1456–1462. doi: 10.1126/science.7878464. [DOI] [PubMed] [Google Scholar]
- Toth AL, Robinson GE. Worker nutrition and division of labour in honeybees. Anim. Behav. 2005;69:427–435. [Google Scholar]
- Tustain RCR, Faulke J. Effect of carbon dioxide anesthesia on the longevity of honey bee in the laboratory. NZJ Exp. Agric. 1979;7:327–329. [Google Scholar]
- Van Dyke T, Jacks T. Cancer modeling in the modern era: Progress and challenges. Cell. 2002;108:135–144. doi: 10.1016/s0092-8674(02)00621-9. [DOI] [PubMed] [Google Scholar]
- Villumstad E. Influence of colony strength on brood rearing and development of honey bee colonies. Agric. Univ. Norway Res. Bull. 1977;56:2–20. [Google Scholar]
- Wheeler WM. The ant-colony as an organism. J. Morphol. 1911;22:307–325. [Google Scholar]
- Wheeler WM. The Social Insects. Harcourt, Brace & Co.; New York: 1928. [Google Scholar]
- Wille H, Rutz W. Beziehungen zwischen Juvenilhormontiter und Hämozyten erwachsener Sommerbienen (Apis mellifera L.). Schweiz. Landwirtschaft. Forsch. 1975;14:339–353. [Google Scholar]
- Wilson DS, Sober E. Reviving the superorganism. J. Theor. Biol. 1989;136:337–356. doi: 10.1016/s0022-5193(89)80169-9. [DOI] [PubMed] [Google Scholar]
- Wilson EO. The Insect Societies. Belknap Press of Harvard University Press; Cambridge: 1971. [Google Scholar]
- Wilson EO. Sociobiology: The New Synthesis. Belknap Press of Harvard University Press; Cambridge: 1975. [Google Scholar]
- Wilson EO. The sociogenesis of insect colonies. Science. 1985;228:1489–1495. doi: 10.1126/science.228.4707.1489. [DOI] [PubMed] [Google Scholar]
- Wilson RL, Abel CA, Lockhart LL, Huss MH. Indoor wintering small honey bee colonies. J. Kansas Entomol. Soc. 1999;72:289–296. [Google Scholar]
- Winston ML. The Biology of the Honey Bee. Harvard University Press; Cambridge, Massachusetts: 1987. [Google Scholar]
- Winston ML, Slessor KN. Honey bee primer pheromones and colony organization: Gaps in our knowledge. Apidologie. 1998;29:81–95. [Google Scholar]
- Woyke J. Genetic background of sexuality in the diploid drone honeybee. J. Apic. Res. 1980;19:89–95. [Google Scholar]
- Yamazaki K, Boyse EA, Bard J, Curran M, Kim D, Ross SR, Beauchamp GK. Presence of mouse mammary tumor virus specifically alters the body odor of mice. Proc. Natl. Acad. Sci. USA. 2002;99:5612–5615. doi: 10.1073/pnas.082093099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu R, Hagen A, Omholt SW. Cryopreservation of totipotent nuclei from honeybee (Apis mellifera) embryos by rapid freezing. Cryobiology. 1997;35:41–45. [Google Scholar]
- Zhao R, Planalp RP, Ma R, Greene BT, Jones BT, Brechbiel MW, Torti FM, Torti SV. Role of zinc and iron chelation in apoptosis mediated by tachpyridine, an anti-cancer iron chelator. Biochem. Pharmacol. 2004;67:1677–1688. doi: 10.1016/j.bcp.2003.12.036. [DOI] [PubMed] [Google Scholar]